КОМПЕТЕНЦИЯ
БИХЕВИОРИЗМ: СТИМУЛЫ, ОТВЕТЫ И ПОДКРЕПЛЕНИЕ
В предыдущих главах мы проводили различие между движением и действием. Движения, то есть организованные мышечные -сокращения, создают набор поз, двигательный контекст или матрицы, на основе которых формируются акты поведения. В нервный механизм, контролирующий действие, включена информация, поступающая из внешней среды, которая создает нечто вроде силовых полей, но в ней не содержатся сведения о длине той или другой мышцы и о расположении суставов. Таким обра-зом, любое направленное действие может выполняться относительно широким набором потенциальных движений. Перефрази-руя утверждение Гранита относительно рефлексов (на которое мы ссылались в гл. XII), можно сказать, что основными проблемами поведения являются не проблемы движений, как таковых, хотя поведение осуществляется посредством движений. Основные проблемы связаны с процессами настройки, или установки, различных сервомеханизмов, посредством которых вводятся в действие мотонейроны. Эти процессы настройки, как правило, включаются в действие под влиянием факторов внешней среды, действующих в данной ситуации.
Неожиданно оказалось, что вопрос, поставленный нейрофизиологом, входит в сферу интересов психолога. Специфика этих внешних условий, определяющих действие, и есть то, что всегда изучает бихевиорист, пытающийся построить свою концепцию на анализе организации внешней среды, значимые связи которой получают объяснение через поведение организма.Бихевиористы правильно считают, что стимулы, ответы и подкрепление получают свое значение на основании внешних, а не внутренних условий существования организма, хотя необходимо учитывать и поведение организма, без которого картина останется неполной. Почему тогда психологам не ограничиться только «изучением доведения»?
Отношения между стимулом, ответом и подкреплением сложнее, чем это представляется бихевиористу. Хотя центр интересов бихевиориста меняется, все бихевиористы – за небольшим исключением – представляют себе эту связь как прямую последовательность. Стимул -› ответ -подкрепление является той самой последовательностью, которая видна наблюдателю. Однако то, что эта очевидная последовательность есть результат более сложных взаимоотношений, стало ясно только тогда, когда обнаружилось, что мозг обеспечивает этот порядок.
Возьмем, например, тот факт, что рефлекс организуется не па схеме дуги S – R, a с помощью сервосистемы. Этот факт оказал важное влияние на понимание сущности стимула и ответа. Существование механизмов обратных и предваряющих связей усложняет всем известные представления Ньютона и Шерринг-тона о линейной связи стимула и реакции. В результате перед бихевиористами возникли две возможности: либо игнорировать внутреннюю сложность этой системы, либо пытаться объяснить ее и дать трактовку тем изменениям, которые претерпевает эта система. В большинстве случаев бихевиористы игнорировали сложность системы. Однако они не могут уйти от того, что стимулы сами находятся под воздействием возникающих ответов и г напротив, что ответ можно полностью объяснить, только учитывая вызвавшие его условия. Иначе говоря, с точки зрения бихевиоризма стимул и ответ взаимно предполагают друг друга. Эта дилемма может быть разрешена только в том случае, если признать взаимосвязь между S и R. Схема S ‹- / -› R имеет отношение не только к неврологии, но и к логике. Математический подход раскрывает глубокое значение этого взаимодействия (Estes 1959). Предполагается, что стимулы являются элементами этой системы и что их отдельные составляющие соответствуют ответам (рис. XIV-1). Можно сказать, что стимулы – это систематизированные объекты, а ответы – процесс классификации этих стимулов. Однако решение этой дилеммы (S – R) предъявляет очень высокие требования к тому, как освещать факты. В современной психологии большинство расхождений в формулировках происходит из-за неспособности до конца понять эти реципрокные взаимоотношения между стимулом и ответом. В физиологической психологии основные противоречия возникают между теми, кто описывает полученные данные на языке стимулов, и теми, кто предпочитает язык ответов. И конечно, противоречия часто происходят из-за смешения этих языков.
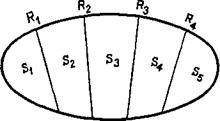
Рис. XIV-1. Концепция, согласно которой реальность описывается в терминах стимулов (Sn) и ответов (Rm). Каждому ответу соответствует свой стимул.
Согласно такому логически и неврологически обоснованному пониманию взаимоотношений между S и R при описании фактов следует придерживаться либо языка стимулов, либо языка ответов, но не обоих сразу. Противоречия возникают потому, что слова, используемые каждым языком, те же самые, хотя их смысл совершенно различен. Большинство отраслей психологии избежало этого противоречия, ограничиваясь лишь тем или другим типом анализа. Так, психологи, занимающиеся восприятием, и психофизики пользуются при описании полученных данных .языком стимулов: механизмы ответов выводятся из организующих свойств стимулов. Напротив, психологи, придерживающиеся концепции оперантного обусловливания, и сторонники психологии познавательных процессов пользуются языком ответов: стимулы определяются теми их свойствами, которые вызывают ответ. Таким образом, возникают две дополняющие друг друга объяснительные схемы, и физиологические данные иногда относятся то к одной, то к другой из них (рис. XIV-2, 3). Путаница возникает тогда, когда схема рассуждения четко не определена. Исходя из этого, рассматривая во второй части книги субъективный опыт, мы придерживаемся языка стимулов, принятого у психофизиков, занимающихся сенсорными процессами, и психологов, исследующих проблемы восприятия. В третьей части мы воспользуемся языком ответов, принятым у психологов, занимающихся оперантным обусловливанием и познавательными процессами, так как этот язык больше соответствует анализу поведения.

Рис. XIV-2. Пример используемых в психологии «языков»: язык стимулов, язык ответов и нейрофизиологический язык.
Возможно, наиболее интересным для анализа этой проблемы « позиций бихевиоризма является объективность, достигнутая в определении подкрепления, когда схема S-› R, или концепция флекторной дуги, заменяется теорией сервомеханизмов. Агентом, запускающим сервомеханизм, является проба на соответствие-или несоответствие между внешними образами, текущим «состоянием» и характером энергии, воздействующей на это состояние. Тйким образом, то, что психологи называют ближайшим («проксимальным») стимулом, зависит от состояния системы, открытой" для ввода информации, то есть от «установок» и «ожиданий» – по терминологии психологии познавательных процессов. Другие рассуждения ставят подкрепляющие свойства среды в зависимость от совпадения условий, которые вызывают данное-поведение, с теми условиями, которые оно само вызывает. В этой главе, а также в гл. XV и XVI излагаются различные данные нейрофизиологии и нейропсихологии, полученные при изучени» процессов подкрепления – механизма, посредством которого осуществляется регуляция поведения. Теперь мы должны приступить к анализу организации процесса поведения во времени.
СОВПАДЕНИЕ И КОНТЕКСТ
Помимо очень большого вклада в технику эксперимента,, бихевиористский подход к изучению поведения значительно обогатил науку своим анализом модификации поведения. Однако объект исследования бихевиоризма ограничен. Бихевиоризм изучал только те изменения поведения, которые непосредственно вызывались стимулами, исходящими от экспериментатора и побуждающими животное к действию. Бихевиористы-экспериментаторы чаще всего не принимали во внимание изменений в поведении, возникающих в результате участия эмоциональных процессов в механизме самоконтроля. Они не придавали должного значения и структурным законам, лежащим в основе изменчивости поведения (Pribram, 1970). Результаты их экспериментов, проведенных главным образом на животных, обедняли теорию бихевиоризма, которая в настоящее время нуждается в уточнении за счет фактов, связанных с деятельностью мозга. Это и есть тот узкий круг вопросов, которые мы будем обсуждать.
С позиций бихевиоризма, изменения поведения происходят благодаря подкреплению. А что же такое подкрепление? Перед последовательными сторонниками бихевиоризма подобный вопрос не возникает: их интересует только описание тех явлений окружающей среды, которые вызывают эффект подкрепления. (Skinner, 1969). Однако заметим, что, даже согласно такому узкому определению, подкрепление должно быть каким-то процессом. Этот процесс должен протекать внутри организма и, таким образом, быть доступным для психологического исследования.
Бихевиористы, занимавшиеся изучением природы процесса научения, провели множество сложных теоретических и экспериментальных исследований, в которых представлены главные точки зрения. Согласно одной из них, известной как теория ассоциаций, предполагается, что научение достигается благодаря повторению событий.
Согласно другой позиции, которой придерживаются сторонники теории влечений и установок, для достижения определенных результатов в научении необходимо изменение внутреннего состояния организма. Как мы увидим дальше, «одна лишь ассоциативная» концепция оказывается несостоятельной для анализа того, что же происходит в мозгу при научении.
Сторонники теории ассоциаций подразделяются на две категории: одни из них предпочитают язык ответов, другие – язык стимулов. И те и другие полагают, что поведение определяется только набором стимулов, организованных по принципу вероятности, причем значение этих стимулов определяется их вероятностью (Estes, 1959). Сторонники теории стимулов полагают, что значение стимулов определяется условиями эксперимента, то есть главным образом вычисленными отношениями между стимулами. Сторонники теории ответов считают, что стимулы приобретают значение благодаря многократно осуществляющимся актам поведения, вследствие чего само поведение становится «саморегулируемым» (Guthrie, 1942).
Подход к проблеме регуляции поведения с позиций теории простого совпадения стимулов и подкрепления сталкивается с рядом фундаментальных вопросов: 1) может ли происходить научение при отсутствии действия (то есть подкрепленного поведения) и, если это возможно, 2) что способствует научению, когда животное выполняет действие?
Мы уже знаем, что научение может осуществляться и без действия. Когда в относительно постоянной ситуации организму многократно предъявляются экспериментально контролируемые сигналы, происходит привыкание организма.
В 60-х годах нейрофизиологи много занимались проблемой привыкания. В результате их исследований наше понимание этого процесса радикально изменилось. Рассмотрим кратко очень важный эксперимент, выполненный Е. Н. Соколовым в Москве (1963). Испытуемому, у которого записывалась электроэнцефалограмма, кожио-гальваиическая реакция и плетизмограмма, с нерегулярными интервалами предъявлялись звуковые сигналы определенной интенсивности и длительности (см. рис. III-1). В начале эксперимента наблюдались характерные изменения указанных физиологических показателей. Эти реакции, известные как ориентировочные, сопровождали изменение поведения. По ходу эксперимента показатели ориентировочного рефлекса постепенно ослабевали, пока звуковой сигнал вообще не перестал вызывать какой-либо эффект. Испытуемый привык к сигналам. Тогда Соколов уменьшил интенсивность звука, не изменив других характеристик сигнала. У испытуемого сразу же снова, появились ориентировочные реакции, проявлявшиеся в указанных показателях.
Рис. XIV-3. Некоторые значения понятий «стимул» и «ответ» в различных языках.
Таким образом, рассуждал Соколов, привыкание не может быть просто каким-то утомлением сенсорных и нервных элементов. Скорее всего, в центральной нервной системе должна сформироваться некая модель стимула, с которой сравниваются все поступающие сигналы. Любые изменения сигнала вызывают ориентировочную реакцию. Соколов проверил свое предположение, предъявив испытуемому, у которого уже обнаружилось привыкание, более короткие звуковые сигналы. Ориентировочная реакция вновь появлялась в момент, когда такие укороченные звуки прекращались. В этом случае реакции, проявляющиеся в ЭЭГ, КГР и плетизмограмме, возникали в ответ на неожиданное наступление тишины.
Как мы уже знаем, множество фактов подтверждает существование самоадаптирующегося механизма, своего рода фильтра, через который проходят поступающие сигналы и с которым они сопоставляются. Этот фильтр можно рассматривать как закодированное представительство (образ) ранее действовавших сигналов, возникающих в результате взаимодействия организма со средой. Этот фильтр формируется постепенно, он изменяется под влиянием сигналов рассогласования (то есть лишь частичного совпадения) и приводит организм к состоянию «ожидания» внешних изменений. Нервные процессы, лежащие в основе этого самоадаптирующегося механизма, были уже детально рассмотрены, и Брунер (1957) в работе о «перцептивной готовности» изложил свои взгляды на значение для психологии экспериментально подтвержденных процессов ожидания. В данной работе я буду говорить об этих результатах, пользуясь терминами «готовность ткани», «компетентные участки мозга» в том смысле, в каком их употребляют эмбриологи при описании развития организма.
Биологическая концепция, согласно которой ткань должна быть «компетентной», для того чтобы дифференцироваться, является результатом ряда работ, авторы которых пытались наглядно показать, как возникает и контролируется процесс дифференциации ткани, то есть как начинается ее развитие. Толчком к развитию служит, в сущности, химическое взаимодействие между «компетентной» тканью и организующими воздействиями среды, которые направляют развитие этой ткани. Классическим примером является один из ранних экспериментов, в котором развитие хрусталика определялось глазным пузырьком. Контакт между этим пузырьком и внешним слоем эпидермы стимулирует пузырек к образованию хрусталика на месте контакта Если глазной пузырек удален, эпидерма не способна превратиться в хрусталик (рис. XIV-4).
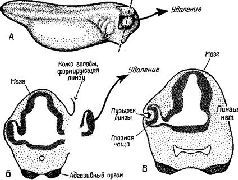
Рис XIV-4 Экстирпация глазного пузырька для демонстрации процесса индукции хрусталика в глазе, Б – поперечное сечение через А по оси X – Y, В -то же самое через несколько дней (Hamburger, 1961).
Этот эксперимент поставил целый ряд проблем, из которых родилось новое направление исследований в экспериментальной эмбриологии, имевшее поразительное сходство с направлением исследований в экспериментальной психологии и этологии. Сначала предполагалось, что индуктор действует только как пусковой механизм (триггер); так, в классическом примере считалось, что эпителий головного конца уже «предрасположен» к образованию хрусталика и требуется только сигнал к старту Эта концепция индуктора была опровергнута двумя видами данных. Во-первых, было доказано методом трансплантации, что зачаток глаза превращается в хрусталик не обязательно в коже, взятой из головного конца эмбриона, но, например, и из эпителия туловища. Во-вторых, в других экспериментах по трансплантации было показано, что та область головного конца, в которой обычно формируется хрусталик, является полипотенциалъной и, следовательно, вовсе не предназначена для формирования только одного хрусталика. Например, если участок головной эпидермы, иэ которой обычно формируется хрусталик, соединяется с индуктором слухового органа, то иэ этого участка разовьется ухо; если он соединяется с обонятельным индуктором, то образуется орган обоняния (рис. XIV-5).
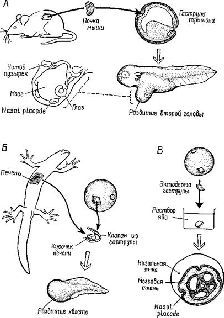
Рис. XIV- 5. A – после имплантации кусочка почки мыши в гаструлу тритона, мозг, расположенный рядом с навальными placodes, глазными и ушными пузырьками, уже начал формироваться; Б – кусочек печени тритона, имплантированный в оболочку эктодермы гаструлы и обработанный раствором яда, дифференцируется на мозговой пузырек и несколько назальных placodes (Holtfreter and Chuang, см. Young, 1957).
Кроме этих фактов, существуют другие, которые показывают, что реагирующая ткань должна быть «готова» или «компетентна», то есть она должна находиться в соответствующем состоянии реактивности, чтобы индукция стала эффективной. Например, ткань, процесс превращения которой в различные специализированные структуры уже «запущен», совершенно не отвечает на индукцию. Далее, было обнаружено, что индукторы действуют независимо от принадлежности животного к тому или другому виду Один и тот же индуктор может быть эффективным по отношению к ткани, принадлежащей животным различных видов, родов и даже отрядов Следовательно, можно предположить, что в основе действия индукторов лежат химические свойства, общие для многих организмов (мы будем подробнее говорить об этом дальше). По-видимому, химические агенты определяют общий характер воздействия индуцируемой структуры, тогда как наследственные механизмы клеток этой структуры определяют ее точную форму. Например, если участок кожи, взятый с боковой поверхности туловища эмбриона лягушки, трансплантируется на голову саламандры, чтобы сформировать соответствующие структуры из ткани саламандры, то у эмбриона саламандры будет развиваться голова саламандры, но с рядом признаков, типичных для лягушки (рис. XIV-6 и XIV-7).
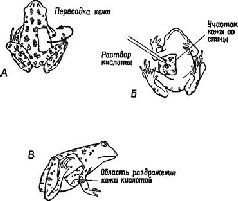
Ответы лягушки при раздражении участка кожи, пересаженного со спины на живот
Рис. XIV-6. Результаты перемещения лоскутов кожи молодой лягушки. Подобные пересадки способствуют развитию врожденных механизмов пигментации лягушки, что происходит после метаморфозы, как это видно на рис. А и Б, где изображена взрослая лягушка сверху и снизу. Вследствие перемещения лоскута кожи со спины на живот прерванные волокна дорсальных корешков спи-пальных нервов регенерируют в коже живота, и наоборот. При раздражении пересаженного участка кожи возникают противоположные по характеру поведенческие реакции (потирание спины вместо живота, и наоборот). Это значит, что центральная нервная система благодаря развитию сенсорных аксонов в пересаженном участке кожи получает знания скорее о локальном участке кожи, чем об общей топографии поверхности тела и топологии центральной нервной системы (Sperry, см. Eddc, 1967).
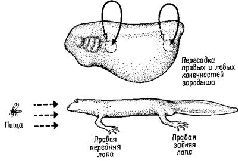
Рис. XIV-7. Эксперимент по изучению моторики, дополняющий опыт, показанный на рис. XIV-6. Пересаженные конечности действуют (во время плавания) так, как если бы они были на своем первоначальном месте. Если правые лапы пересаживаются на место левых, они продолжают работать как правые конечности, несмотря на новую иннервацию. Возникающее в результате этого поведение может не соответствовать требуемому, как это видно на рисунке, где животное пытается плавать, но совершает при этом движения, отдаляющие, а не приближающие его к пище И в этом случае приобретенная компетентность определяется периферической тканью. Эти результаты трудно объяснить с точки зрения обычных представлений о нервных связях. Я объясняю эти факты (Pribram, 1961, 1965) тем, что пространственно-временные структуры нервных импульсов становятся теми кодами, которые опознаются периферической тканью. Подобные пространственно-временные структуры начинают декодироваться в центральной нервной"т:истеме по мере ее развития независимо от путей, по которым они передаются Эти структуры импульсов специфичны по отношению к периферическим структурам, так как каждая периферическая иннервация характеризуется специфическим размером волокон, которые определяют скорость проведения нервного импульса (Quilliam, 1956; Thomas, 1956; см Weiss, 1950, Hamburger, 1961)
В результате большого числа химических экспериментов постепенно утвердилось мнение, что рибонуклеиновая кислота (РНК) является наиболее вероятным, а возможно, и единственным веществом, которое ответственно за эффект индукции (см. Niu, 1959), хотя и рибонуклеопротеины, и стероиды не могут быть полностью исключены из этого процесса. В большинстве случаев рибонуклеаза разрушает эффект индукции, хотя остается нерешенным вопрос, оказывает ли рибонуклеаза какие-либо иные воздействия на индуцируемую ткань, в которой нарушается процесс дифференциации. Однако более прямые доказательства роли РНК в процессе индукции были получены в опытах с демонстрацией эффекта индукции РНК, выделенной из различных органов. Было показано и то, что выделенная из различных органов РНК способна стимулировать реципиентную ткань к тому, чтобы она превращалась в различные специфические структуры. Эти эксперименты свидетельствуют о том, что в организме существует много видов РНК, каждый из которых имеет свои специфические функции.
Итак, сторонники теории ассоциаций считают, что изменения поведения возникают всякий раз, когда события (стимулы) происходят одновременно или если такие одновременно происходящие стимулы (события) возникают благодаря ответам организма. Факты привыкания показывают, что «стимулы» с позиций теории ассоциаций должны рассматриваться как «проксимальные»; воздействие становится стимулом вследствие частичного совпадения «входной» информации с состоянием центров, которое в свою очередь формируется благодаря предшествующим частичным совпадениям центральных состояний и информации, поступающей от внешних воздействий. Таким образом, в любой отрезок времени центральные механизмы должны быть «компетентны», готовы к тому, чтобы обеспечить фон, на котором возникают стимулы. Следовательно, связь стимулов нужно рассматривать не как некую неопределенную, случайную, вероятную «ассоциацию» а как биологически детерминированный процесс, Который организует взаимоотношения контекста (фона) и воздействия стимула. Если это так, то теория совпадения стимулов и теория ожидания сближаются с точки зрения процессов, происходящих в центральной нервной системе.
Таким образом, простого случайного сочетания, как такового- как фактора научения, – не существует, и эта концепция должна быть заменена биологической концепцией, основанной на принципе «компетентности»; наборы воздействий-стимулов (по существу, врожденных, хотя они и модифицируются в результате опыта) кодируются в центрах, которые приобретают «компетентность» по отношению к последующим наборам воздействий, создаваемых стимулами.
Этот процесс модификации поведения напоминает процесс эмбриогенеза, последний определяется унаследованными, врожденными свойствами, пробуждаемыми к жизни стимулирующей функцией среды, в которой растут клетки, Стимулирующая функция сама по себе специфична, но эта специфичность несколько иная, чем генетический потенциал. Генетическая «компетентность» индивидуально специфична (она специфична для каждого вида, рода, отряда). Наследственные факторы предполагают общность прошлого с будущим, допуская определенные вариации внутри каждого отдельного поколения. Индукторы, напротив, неспецифичны по отношению к индивидам, видам и т. д., поскольку существуют относительно простые химические агенты – РНК, общие для всех живых организмов. Таким образом, индукторы обеспечивают ту присущую всему живому общность, которая открывает возможность модификаций механизмов «компетентности» в отдельных поколениях в соответствии с требованиями момента.
СМЕЖНОСТЬ И ПОСЛЕДОВАТЕЛЬНОСТЬ
Второй вопрос, который возникал при бихевиористском подходе к проблеме, был следующим: что способствует научению, когда совершается действие? Можно сформулировать теперь этот вопрос иначе и спросить, происходит ли привыкание только при повторении стимулов или этот принцип применим к более широким проявлениям поведения. Хотя мы еще не можем дать точный ответ на этот вопрос – для этого необходимы новые исследования, – было высказано предположение, что процесс привыкания и угашение поведения имеют общие факторы. Например, Д. Примак (Premack and Collier, 1962), анализируя случаи ненодкреп-ления, отражающиеся на вероятности ответов, счел нужным заявить следующее:
«Существует по крайней мере ряд данных о безусловных ответах, которые не сохраняются полностью при их повторном вызывании. Хотя эта проблема мало изучена, в одном из немногих имеющих отношение к данному вопросу исследований Доджа (1927) сообщается о небольшом, но явно необратимом уменьшении величины и увеличении латентных периодов даже такого безусловного рефлекса, как коленный. Далее, в биологической литературе о привыкании содержатся описания нескольких случаев, когда безусловные ответы уменьшались по мере повторения, а их латентные периоды увеличивались… Возникает вопрос… не имеет ли этот факт некоторого несомненного уменьшения безусловных рефлексов более широкое значение для понимания поведения, чем это принято считать» (р. 15).
Только признав, что поведение регулируется механизмом, сходным с тем, который контролирует сенсорные процессы, можно, по-видимому, объяснить и правильно понять подобные факты. Однако остается вопрос: почему же поведенческие реакции не всегда ослабевают при их повторении? Первый приблизительный ответ состоит в том, что при нормальном ходе событий должно происходить «прерывание» процесса привыкания и угашения и что одна из функций подкрепления как раз и заключается в том, чтобы вызывать такое «прерывание». Эксперимент С. Гликмана и С Фельдмана (1961) показывает, что подобное «прерывание» действительно происходит. Этих исследователей интересовало, будет ли угасать реакция активации в ЭЭГ, которая возникает при стимуляции структур ствола мозга как компонент ориентировочного рефлекса (или бодрствования), при повторении внешних стимулов. Они обнаружили, что активация в ЭЭГ, вызванная стимуляцией ствола мозга, угасает, если только электроды не введены в ту область, в которой возникает эффект самораздра-жения (рис. IX-8). Повторное раздражение этих зон, по-видимому, непрерывно возбуждая животное, создавая и изменяя установки, как это показано в гл. X, действует как фактор, прерывающий процесс привыкания.
Подводя итоги проблемам, поставленным сторонниками теории ассоциаций или совпадения стимулов с подкреплением (ассоциаций по смежности), следует сказать, что ни теория «отбора стимулов», ни теория «саморегуляции поведения» не могут дать адекватного ответа на вопрос о модификации поведения, вероятно, потому, что эти теории не принимают во внимание важность организации процесса научения во времени. Организмы реагируют не налюбые, одновременно происходящие смежные явления. Их поведение направляетсяпредварительно сформированной «компетентностью» мозга, организующей стимулы и включающей в действие лишь те, которые важны для поведения. Таким образом, стимулы представляют собой воздействия, детерминированные нервной системой и «отобранные» на основании центральной «компетентности» организма (или его нервной «установки»), которая в свою очередь детерминирована предшествующим опытом и другими центральными процессами. Поведение организма направляется не только стимулами, но и посредством самоадапти-рующегося фильтрующего механизма, который в определенных случаях сам индуцирует стимулы (то есть вызывает ориентировочную реакцию). Это происходит в тех случаях, когда результат поведения лишь частично совпадает с центральной «компетентностью», которая и программирует поведение. В этих условиях как раз и имеет место подкрепление и поведение становится саморегулируемым.
Интересный эксперимент, проведенный Э. Джоном (John and Morgades, 1969), показал, что организм отвечает должным образом не только в силу того, что воздействия происходят одновременно. Три кошки были обучены нажимать на один рычаг при вспышках света частотой 2 кол/сек, и на другой, когда частота вспышек была равна 8 кол/сек. Регистрировалась электрическая активность в латеральном коленчатом теле – эвене зрительной системы. Было показано, что, когда животное правильно выполняло задание на различение вспышек, возникающие электрические ответы четко различались по форме волн. У четвертого необученного (контрольного) животного подобных различий в форме вызванных ответов не наблюдалось. Когда кошка ошибалась, то есть «огда давался неверный двигательный ответ при каком-либо сигнале, исчезало также и различие в форме электрических колебаний на этот сигнал. Джон пришел к выводу, что за наблюдаемые различия, коррелирующие с правильными ответами, должны быть ответственны определенного рода динамические процессы (с точки зрения концепции, развиваемой в этой книге, по-видимому, нейронные голографические процессы).
«У тренированных в разной степени животных форма вызванных потенциалов, возникающих под каждым электродом в ответ на два различных стимула, отчетливо различается. Эти различия в вызванных ответах наблюдались при регистрации реагирующих ансамблей клеток в пределах ограниченных областей мозга. Таким образом, клеточные ансамбли, сообщающие о наличии дифференцируемых сигналов, широко распространены по обширным областям мозга и постоянно обнаруживают два типа ответов на два различных стимула. Анализ показывает, что информация, содержащаяся в усредненных ответах каждого из этих двух локальных клеточных ансамблей, достаточна для различения двух периферических сигналов. Ансамбль из многочисленных реагирующих клеток обнаруживает поразительную стабильность в ответах, говорящую о существовании неизменного типа разрядов в ответ на каждый отдельный стимул. Напротив, активность одиночной клетки в высшей степени вариативна, проявляясь в виде разнообразных типов ответов при действии тех же самых стимулов. Эти факты говорят о том, что наборы разрядов обширных нейронных ансамблей служат гораздо более надежным источником информации, чем наборы разрядов одиночных клеток.
V обученных животных наблюдалось поразительное сходство в форме волн электрических ответов, регистрируемых в различных мозговых структурах. Анализ степени наклона кривых выявил, что эти сходные ответы широко распространены в головном мозгу, исключая проводящие пути. Динамическая природа этого феномена была проиллюстрирована фактами нарушения сходства при ошибочных ответах или при предъявлении новых стимулов. Эти факты свидетельствуют о том, что сходств» электрической активности в различных областях мозга отражает протекание некоторого эндогенного процесса, происходящего при научении, а не пассивно проявляющегося при действии физического стимула, как такового» (John and Morgades, 1969, p. 205-206).
Такое понимание процесса подкрепления начинают признавать даже бихевиористы. Так, в недавнем обзоре (Perkins, 1968) утверждается, что концепция «привлекательности» объединяет целый ряд ранее разрозненных фактов. Согласно этой теории, «все классические условные рефлексы могут быть описаны как подготовительные ответы, которые повышают характерную привлекательность комплекса стимулов, совпадающих с предъявлением безусловных стимулов». Другие авторы употребляют термин «желательность», «побудительная ценность» или «побуждение»; я пользуюсь в этой книге термином «интерес» как субъективным эквивалентом «привлекательности». Однако общий смысл: этих терминов сводится к тому, что назначением процесса подкрепления является обращение к некоторым предшествующим состояниям, к некой компетентности организма, что в свою очередь приводит к изменению этих состояний.
КОМПЕТЕНТНЫЙ МОЗГ
Что же тогда представляют собою эти состояния компетентности, которые регулируют поведение? Это иерархически организованные механизмы (логические модули), включающие серво-процессы, программы или планы, направленные на достижение внешнего эффекта, действия. В гл. XII и XIII была подробно описана нейронная организация механизма компетентности, участвующего в таких действиях, как, например, забивание гвоздя; анатомические структуры с однозначным соответствием между мышцами и корой мозга организуются в образ, который контролирует любые движения, диктуемые внешним или «силовым нолем» для достижения результата. Этот результат кодируется в образе, в состоянии, в микроструктуре медленных потенциалов соединений; все это происходит посредством оценки мгновенных экстраполяции тех изменений в текущем процессе регулирования, которые появляются при сопоставлении соответствующих силовых полей с достигнутым результатом.
Несколько более простой, но аналогичный механизм может быть описан по отношению к другим, более «внутренним» состояниям компетентности. Если у животных структуры ствола мозга (например, гипоталамус) стимулируются дискретными импульсами, возникают отдельные движения, имеющие отношение к еде, питью и т. д. Участки, ответственные за подобные отдельные движения, по-видимому, распределены в случайном порядке по довольно широкой области мозга. Так, например, движение облизывания губ может быть вызвано раздражением участка, расположенного вблизи зоны, ответственной за движение выгибания спины (Robinson, 1964). Широко рассеянными оказываются, по-видимому, даже участки мозга, являющиеся субстратом генетической памяти, связанной с инстинктами. Однако при более сильной стимуляции или раздражении более широких зон эти отдельные движения интегрируются в структуры поведения, которые связаны с той или другой инстинктивной потребностью или аффектом. Таким образом, движения захватывания чего-нибудь ртом, жевание и заглатывание образуют комплекс движений при «де; пригибание и отворачивание в сторону указывает на начало полета; выпускание когтей и оскаливание зубов, выгибание спины, расширение зрачков означает агрессию. Можно предположить, что сильная электрическая стимуляция выявляет те движения, которые в обычных условиях возникают при активации тех же самых структур во время аффективных состояний и влечений.
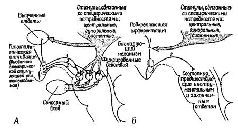
Рис. XIV-8. А – подкрепление моторных структур. Схема взаимосвязей между «подкрепляющей системой мозга» и системой ответов, основанная, на результатах экспериментов; В – роль сенсорного входа. Схема иллюстрирует взаимоотношение между специфическими потребностями и «непосредственным подкреплением», получаемым через сенсорный вход (Valenstein, 1970).
Это предположение родилось в результате серии изобретательных экспериментов, выполненных Э. Валенстайном, которые, однако, в свою очередь поставили ряд новых вопросов. В этих экспериментах электроды вживлялись в ту область гипоталамуса крыс, электрическое раздражение которой вызывает движения еды, питья и попыток грызть кусок дерева. Когда один из таких поведенческих актов наблюдался особенно явно и стабильно, внешние условия эксперимента менялись таким образом, что первоначальный результат нельзя было получить. Так, в опыте создавались такие условия, когда в течение целой ночи животное через нерегулярные интервалы подвергалось стимуляции. На следующее утро та же стимуляция ранее раздражаемого участка вызывала либо первоначальное, либо измененное поведение, что зависело от внешних условий, и, когда были испробованы все возможные варианты, обнаружилось, что часто доминировали новые выработанные движения. Валенстайн пишет:
«В стандартных тестах на выявление обусловленного стимулами поведения удалялся объект (цель, к которой стремилась крыса), и животное-переводилось на ночной режим, когда ему предъявлялись два других целевых объекта. Если, например, крыса во время стимуляции в первой серии опытов пила, то в ночное время сосуд с водой удалялся и в клетке сохранялись только кусочки пищи и дерева. Параметры стимула не менялись. Если животное не проявляло нового поведения, оно переводилось во второй, третий и т. д. раз на ночной режим. Однако в большинстве случаев-одного перевода на ночные условия было достаточно, чтобы возникало новое поведение. После появления нового поведения животным предъявлялись два стандартных теста, в которых начальный целевой объект отсутствовал, и третий стандартный тест (контрольный) со всеми тремя целевыми объектами. В тех случаях, когда животное не подвергалось стимуляции, всегда имелись все три целевых объекта, и, следовательно, животные могли насытиться пищей, напиться воды и удовлетворить свою потребность в том, чтобы грызть кусок дерева, еще до начала опыта. Одиннадцать животных прошли полностью через это испытание, что и составило первый эксперимент…
Вторая серия опытов была проведена после применения различного числа тренировочных стимуляций в условиях отсутствия целевого объекта, к которому прежде стремилось животное… в большинстве случаев вторично сгимулированное поведение появлялось так же закономерно, как и прежнее. Во время предъявления контрольного теста, когда одновременно присутствовали все три целевых объекта, в большинстве случаев с равной? вероятностью наблюдались два типа поведения. Как правило, один тип поведения возникал при действии первых трех-четырех серий стимулов, а затем возникало другое поведение в приблизительно одинаковых по длительности сериях опытов. При предъявлении контрольных тестов животные обнаруживали несколько видов поведения за время предъявления некоторых из 20 стимулов.
Мы хотели выяснить, станет ли доминирующим первый тип поведения, если мы дадим серию контрольных тестов. Хотя в этих опытах наблюдались разнообразные виды поведения, мы пришли к заключению, что как только закреплялся второй вид обусловленного стимулами поведения, печальное поведение, вызванное стимуляцией, переставало быть доминирующим. И, действительно, у ряда животных обнаружилось явное преобладание второго вида поведения во время предъявления контрольных тестов (Valenstein, Cox and Kakolewski, 1969, p. 247-249).
Валенстайн объяснил эти результаты тем, что стимуляция гипоталамуса не вызывает специфических мотивационных состояний типа голода или жажды. Исходя из фактов взаимодействия двух типов явлений – эффекта самораздражения по Олдсу и эффекта вызова с помощью стимуляции определенного вида поведения (еды, питья или попыток грызть дерево), – он приходит к выводу, сходному с тем, который был изложен в гл. X, а именно что электрическая стимуляция, воздействуя на механизмы гомеоста-зиса, создает установку, изменяющую характер ответов.
Обратимся к одному примеру. Мендельсон (1967) описал поведение животных при стимуляции латерального гипоталамуса, которое проявлялось одновременно и в виде самораздражения, и в виде питья. Могенсон и Стивенсон (1967) приводят сходные данные. Мендельсон отобрал животных, у которых реакции питья появлялись при более слабой стимуляции, чем та, которая необходима для поддержания эффекта самораздражения. На этом низком уровне стимуляции животные не нажимали на рычаг, когда не было воды, однако когда жажда утолялась, они также не нажимали на рычаг для получения воды, если только не стимулировался гипоталамус. Мендельсон сделал следующий вывод: «Таким образом, если у крысы вызвать состояние жажды, она будет нажимать на рычаг, чтобы получить воду, а если крысе давать воду, то она будет нажимать на рычаг, чтобы вызвать жажду». Эти результаты очень важны, однако их интерпретация – предположение, что стимуляция вызывает жажду, – не кажется нам достаточно убедительной. Мы могли бы с уверенностью сказать, что у этих животных можно посредством электрической стимуляции вызвать обусловленное стимулом пищевое поведение, и что то же самое поведение можно затем получить и с помощью пищи. Из работ Кунза и Крюса (1968) известно, что животные, проявляющие в ответ на стимулы пищевое поведение, будут подвергать себя самораздражению на уровне ниже «порога вознаграждения», если они получают пищу. Поэтому более разумно было бы предположить, что подкрепление, возникающее результате выполнения вызванного поведенческого акта, обусловливает не голод или жажду, а суммируется с подкреплением, возникающим непосредственно в результате самой стимуляции. Из этого следует, что сумма покрепления от выполнения поведенческих реакций и подпорогового подкрепления, вызванного стимуляцией мозга, может оказаться достаточной, чтобы поддерживать инструментальное поведение. Действительно, ранее Мендельсон (1966) показал, что сытое животное обнаруживает вызванное -стимулами пищевое поведение скорее при комбинации пищи и стимуляции мозга, чем при одной мозговой стимуляции (Valenstein, Cox and Kakolewski, 1969, p. 267-277).
Однако, предполагая, что стимуляция мозга не вызывает состояния жажды, Валенстайн может выбросить вместе с водой и ребенка. У. Роберте (1969) указывает, что сильная электрическая стимуляция может разрушить четко организованное поведение и расширить гипоталамическую зону, при раздражении которой постоянно наблюдаются частные движения, включенные в тот или иной поведенческий акт. Роберте обвиняет Валенстайна в том, что он стоит па позициях Лешли, отрицая специфичность нервных центров. Эта позиция, как он считает, должна быть пересмотрена, потому что, как отметил П. Милнер (Roberts, I960, р. 17-19), она «заставила многих исследователей на несколько десятилетий отказаться от изучения проблемы функциональной специфичности и локализации функций». Вопреки этому Роберте считает, что компетентная ткань состоит из «перекрывающих друг друга, но специфических по своим функциям механизмов».
Этот аргумент не нов, однако смысл его изменился. Почти сто лет поборники теории локализации функций спорили со сторонниками теории пластичности мозга; полем боя были зрительная и моторная кора. Теперь поле сражения переместилось в гипоталамус.
Как при анализе восприятия и действия, решение вопроса и в этом случае надо искать в микроструктурах медленных потенциалов соединений. Роберте признает существование данной проблемы, но не может дать ей неврологического объяснения.
«Большая частота ответов, имеющих незавершенный характер… подтверждает мнение о том, что [локальная] стимуляция гипоталамуса обычно возбуждает только небольшую часть нейронов данного механизма. Эти факты говорят также о том, что существует очень мало постоянных взаимосвязей между нейронами, контролирующими различные элементы этих ответов, внутри гипоталамуса или в его эфферентных путях. Таким образом, тенденция этих элементов возникать одновременно при нормальном поведении должна быть результатом несколько иного типа интеграции, например, результатом общих гуморальных влияний, как это наблюдается в случае терморегуляторного поведения (Roberts, et al., 1969), или результатом общего нейронного входа от других центральных структур или периферических рецепторов» (Roberts, 1969).
Общий вход, интеграция создается, конечно, в экспериментах Валенстайна благодаря комбинации электрической стимуляции моэга и подкрепления от окружающих условий, что – согласно концепции, изложенной в этой книге, – формирует окончательную структуру поведевия, которая опирается на динамическую систему медленных потенциалов соединений. Эти микроструктуры медленных потенциалов, эти кратковременные установочные состояния и регулируют поведение. Конечно, эти микроструктуры не обязательно локализованы в стволе мозга. Активирующий латеральный гипоталамическии механизм является основным перекрестком путей, проводящих сигналы от многих областей мозга, включая и кору больших полушарий.
Результаты, полученные Валенстайном, как и повседневные наблюдения, говорящие о том, что заученные навыки сохраняются надолго, требуют, однако, более стабильной основы, чем эти кратковременные состояния, создаваемые микроструктурами соединений. Субстратом окончательного образа результата не могут быть чисто электрические процессы. Должны возникать какие-то длительные модификации, химические или нервные.
Таким образом, мы вернулись к вопросам, рассмотренным во II главе. В следующей главе мы еще раз обсудим эти вопросы и проанализируем модификацию поведения с точки зрения мотивации процессов памяти, то есть проблему подкрепления.
РЕЗЮМЕ
В моторных механизмах мозга кодируются условия внешней среды, которые организуют поведение. Для решения проблемы, как это происходит, одинаково важны и работа нейрофизиологов, и усилия бихевиористов, направленные на изучение модификации поведения в результате научения, и в частности на анализ тех процессов, с помощью которых внешние условия регулируют изменение поведения и становятся взаимно релевантными. В соответствии с принципами работы нервной системы механизмы, контролирующие поведение, даже те, которые имеют отношение к восприятиям и чувствам, представляют собой сложные системы, связанные с широким классом функций. В этих широких системах формируются компетентные структуры, особые функции которых зависят от опыта организма в данной внешней среде. Внешнее поведение организма определяется сложно организованным механизмом, сходным с тем, который продуцирует образы желаемых результатов. Простая последовательность воздействий не влияет на результат поведения; условия должны воздействовать на внутреннюю компетентность организма или регулировать поведение на основе предшествующего опыта данного организма и ожидания нужных результатов.
Глава XV
ПОДКРЕПЛЕНИЕ И ЗАВЕРШЕНИЕ ДЕЙСТВИЯ
ПОДКРЕПЛЕНИЕ КАК ИНДУКЦИЯ
Сформулированное в гл. XIV утверждение о том, что процесс подкрепления индуцирует изменения в механизмах компетентности мозга, нуждается в тщательной проверке. Имеется ли какое-либо сходство между процессами индукции, изучаемыми эмбриологами, и процессами подкрепления, изучаемыми бихевиори-стами? В гл. II процесс индукции рассматривался в качестве-модели хранения следов памяти. Какие нервные механизмы могли бы приходить в действие во время процесса подкрепления, который вызвал бы процесс индукции в мозгу взрослого организма?
Наиболее спорная трактовка подкрепления, предлагавшаяся как бихевиористами, так и психоаналитиками, заключалась в попытке рассматривать подкрепление как фактор, уменьшающий влечение. Эта концепция основывается на двухфакторной теориш влечения: физиологические потребности приводят к состоянию напряжения в организме, которое проявляется в повышении общей» активности или в усилении нервной активации; поведение, направленное на уменьшение этого напряжения, и есть подкрепление. Некоторые авторы (например, Sheffield et al., 1955) считают, что организмы, напротив, стремятся к повышению напряжения и что изменения поведения сопровождаются ростом: напряжения. Этот аргумент устраняет необходимость во втором факторе. Однако влечение и подкрепление по-прежнему рассматриваются как взаимосвязанные явления. Между тем Эстес (1958) привел убедительные доводы в пользу концепции, согласно которой поведение строится по схеме «побуждение – стимул», а не-по схеме «побуждение – напряжение», однако эта концепция: оставляет без ответа как раз вопрос о том, какие же побуждающие стимулы управляют поведением. Ответ на этот вопрос сходен с тем, который был дан в предыдущей главе, а именно вопрос,, что такое «стимул», – не так прост, как кажется. Побуждающий стимул, как и сенсорный стимул, обусловлен работой корригирующего сервомеханизма, или гомеостата. Гомеостаты снабжены рецепторами, чувствительными к возбуждению в системе «внешний мир – внутренняя среда». Особые зоны мозга, чувствительные к осмотическим колебаниям, изменениям температуры, содержанию эстрогена, глюкозы и уровню углекислоты, находятся вокруг желудочков мозга; они расположены по средней линии; зти области связаны с механизмами, контролирующими поступление и расход агентов, к которым они чувствительны (гл. IX, X).
Помимо этих центральных механизмов, в гомеостатических процессах принимают участие также другие, более периферические механизмы. Центральные гомеостатические процессы часто дополняются вторичными механизмами, которые обеспечивают более точно дозированную регуляцию указанных агентов. Примерами могут служить сокращения желудка в механизме пищевой регуляции во время голода или сухость во рту при жажде, а также регуляция циркуляции крови в сосудах кончиков пальцев для большего или меньшего охлаждения тела. Основные гипота-ламические терморегуляторы дифференцированно управляют температурой пальцев посредством кровообращения.
Если взять в качестве модели голод, то можно видеть, что мозговой гомеостатический механизм с его центральными и периферическими чувствительными аппаратами имеет два реципрокно действующих компонента. Один из них сигнализирует о недостатке какого-либо агента и запускает регуляторный процесс, другой сигнализирует о насыщении организма и останавливает этот процесс. Можно также видеть, что фаза запуска регулятор-ного процесса характеризуется наличием потребности, а фаза прекращения вызывает состояние аффекта.Следовательно, оказывается, что потребности и аффекты, чувство заинтересованности представляют собой мотивационные и эмоциональные (в отличие от перцептивных) стимулы, иначе говоря, то, что бихевиористы называют влечениями. На языке этой книги потребности и аффекты являются управляющими образами, индикаторами процессов, связанных с состояниями мозга и таким путем: влияющих на организацию поведения во времени (гл. II).
Каким образом механизмы, обеспечивающие состояния интереса, потребности и аффекта, модифицируют поведение организма и служат подкреплением? Путем вовлечения механизма памяти, которым обладает организм. В гл. II была сформулирована гипотеза, согласно которой структуры, ответственные эа долговременную память, как бы стимулируются в мозгу, подобно тому как стимулируются тканевые структуры в процессе эмбрионального развития. Внешнее сходство между процессами индукции, изучаемыми в эмбриональных тканях, и процессами подкрепления, изучаемыми в ситуации выработки условных рефлексов, легко сформулировать в следующих положениях: А) индукторы пробуждают и организуют генетический потенциал организма. Подкрепляющие агенты пробуждают и организуют поведенческие возможности организма. Б) Индукторы сравнительно специфичны в отношении действия, которое они вызывают, но, как правило, неспецифичны по отношению к индивидам и тканям. Подкрепляющие агенты довольно специфичны по отношению к тому поведению, которое они обусловливают, но, как правило, неспецифичны по отношению к индивидам и задачам. В) Индукторы детерминируют широкий набор индуцированных эффектов, детали уточняются процессами, протекающими в субстрате, по мере развития ткани. Подкрепляющие агенты определяют решение проблемной ситуации, детали поведенческого репертуара (конкретные формы поведения), используемого для достижения результата, сугубо индивидуальны. Г) Индукторы не являются просто агентами, запускающими развитие, они представляют собой нечто большее, чем мимолетные стимулы. Подкрепляющие агенты не просто агенты, запускающие поведение; они представляют собой особый класс стимулов. Д) Индукторы должны контактировать с субстратом, для того чтобы быть эффективными. Совпадение во времени – обязательное требование для того, чтобы реализовать подкрепление. Е) Простой контакт, хотя он и необходим, недостаточен, чтобы вызвать эффект индукции Индуцируемая ткань должна быть готовой к реакции, должна быть компетентной. Простое сочетание, хотя оно и необходимо, недостаточно, чтобы вызвать подкрепление; подготовленное состояние, недостаток информации, депривация, готовность, контекст, ожидание, внимание, создание гипотезы – вот только некоторые из терминов, используемых для описания тех факторов, которые создают компетентность организма, без чего подкрепление не может стать эффективным. Ж) Индукция обычно осуществляется благодаря двустороннему взаимодействию – благодаря химическому взаимодействию. Подкрепление является наиболее эффективным в оперантной ситуации, где последовательность действий самого организма немедленно используется как регулятор его последующего поведения.
НЕЙРОХИМИЯ ПОДКРЕПЛЕНИЯ
После всего, что уже сказано, остается вопрос, указывает ли описанное выше сходство процессов индукции и подкрепления на сходство их механизмов? С моей точки зрения, да. Какие же факты это подтверждают? Какие нервные процессы начинают действовать во время индукции?
Для этого требуются анатомические пути, функционирующие во многих областях мозга, чтобы запустить в действие подкрепляющий «индуктор», химические агенты, стимулирующие реци-пиентную ткань к дифференциации. Пример подобного любопытного синтеза мы находим в нейрохимическом исследовании, проведенном Л. Стайном (рис. XV-1, XV-2).
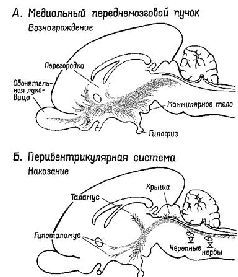
Рис XV-1 Схема медиального переднемозгового пучка (верхний рисунок – система запуска) и перивентрикулярной системы волокон (нижний рисунок – система остановки) в мозгу млекопитающих (сагиттальный разрез) (Stein, 1968, Le Gros Clark et al, 1938)
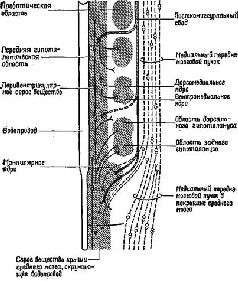
Рис. XV-2. Схема медиального переднемозгового пучка (пунктир) и пе-ривентрикулярной системы волокон в горизонтальной плоскости. На схеме гипоталамус представлен как три продольно расположенных столба – перивентрикулярный поясной слой, медиальная зона и латеральная зона. Пучки волокон в перивентрикулярной и латеральной зонах примыкают к медиальной зоне, которая лишена крупных пучков волокон, но содержит несколько известных ядер. Наклонные линии обозначают систему свода (Stein, 1968; Sutin, 1966).
«Все приведенные факты отвечают представлению о том, что механизм подкрепления содержит систему адренэргических Синапсов, которые в высшей степени чувствительны к фармакологическим воздействиям Усиление порадренэргической передачи в этих сииаптических аппаратах вызывает эффекты облегчения в поведении, ухудшение норадренэргической передачи подавляет поведение При всех условиях эти синаптические аппараты являются важнейшим участком работы мозга, где амфетамин и хлорпромазин оказывают влияние на целенаправленное поведение.
Где локализуются эти синапсы? Сели в медиальном переднемозговом п\чке действительно находится система вознаграждения, как эю уьазыва.-лось выше, то данные адренэргические синапсы, очевидно, уже описаны группой сотрудников Каролинскою института (Fuxe, 1965, Hillarp, Vuxe and Dahlstrom, 1966) Используя гистохимическую технику для обнар›же ния катехоламинов на клеточном уровне, эти исследователи сообщают о системе нейронов, содержащих порадреналин; клеточные тела этих нейронов берут начало в мезенцефальных отделах мозга, а волокна их восходит к медиальному иереднемозгопому пучку и оканчиваются в адренэргиче-ских синапсах гипоталамуса, лимбической доли и неокортекса… Используя совершенно иную технику, независимо от предыдущих авторов Геллер, Сайдеи и Мур (1966) подтвердили существование этой восходящей системы волокон. Эти исследователи разрушали медиальный переднемозговой пучок па одной стороне на уровне латерального гипоталамуса и затем проверяли изменение содержания норадреналина в различных областях мозга. Поскольку в медиальном переднемозговом пучке аксоны в большинстве случаев не перекрещиваются, оказалось возможным сравнить уровень содержания норадреналина на пораженной и интактной сторонах. Было обнаружено, что на пораженной стороне – и только в структурах пучка мозга – норадреналина было значительно меньше; контрольные разрушения медиального гипоталамуса не привели к существенному уменьшению норадреналина. Эти результаты подтверждают существование в медиальном переднемозговом пучке волокон, содержащих норадреналин; кроме того, авторы приходят к выводу, что эти волокна входят в восходящую систему, так как уменьшение уровня норадреналина наблюдается только в структурах, расположенных выше уровня поражения…» (Stein, 1968, р. 110.)
Шейбел и Шейбел (1967, рис. XV-3 и XV-4) также описали эту систему нейронов; по мнению этих авторов, она функционирует как система неспецифических афферентных путей к коре больших полушарий. Характерно, что аксоны этих клеток длинные и на их окончаниях имеются разветвления, обвивающие апикальные дендриты пирамидных клеток, свободно соединяющиеся с дендритами, в противоположность хорошо ограниченным синапсам, типичным для специфических афферентов. Эти данные, а также эксперименты Фукса, Гамбургера и Хокфельта (1968), показавшие, как отмечает Стайн, что неспецифические соединения являются в большинстве случаев адренэргетическими (то есть, что аксоны и их клетки содержат норадреналин), говорят о том, что эта система нейронов ответственна за запуск индуктивных процессов посредством стимуляции РНК и облегчения синтеза протеина. Стимулирующее влияние норадреналина на синтез, протеина, по-видимому, опосредствовано циклом адениномонофосфата (см. «The Role of Cyclic AMP in the Nervous System» in «Neurosciences Research Program Bulletin», July 1970, vol. 8). Кити как-то отметил: «Интересно, что стимуляция протеи-нокиназы посредством адениномонофосфата может значительно усиливаться с помощью ионов магнезии или калия и тормозиться кальцием, что показывает, каким образом эффект адренэргической стимуляции может по-разному воздействовать на умеренно активные и неактивные синапсы!»
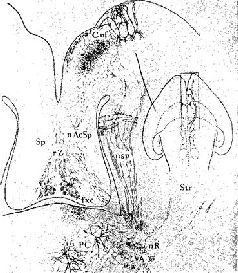
Рис. XV-З. Ростральные проекционные пути неспецифической талампческой системы, идущие через нижние ножки таламуса (часть медиального переднемозгового пучка) в кору. Аксоны от передних неспецифических полей, включающих парацентральное ядро (PC) и, возможно, медиальную часть вентрального переднего ядра (VA), проходят рострально через нижние ножки таламуса как неспецифические проекционные пути (nsp) и через медиальный сектор каудального отдела полосатого тела (Str) к основанию орбитофронтальной коры (Cof). Эти аксоны широко ветвятся в субгризеальном белом веществе, и некоторые из них лродолжаются в ядрах ограды -п. accumbens (nAcSp). Остальные сокращения означают: ретикулярное ядро таламуса (nR), столбы свода (fxc), ограда (Sp) и радиация Цукеркэндла (rZ). Эпидермальная нейроглия (g) тянется вдоль боковых желудочков. Горизонтально-наклонное сечение, модифицированная методика быстрого окрашивания по Гольджи. 50-дневный, частично демиелинизированный мозг мыши. X150. В репродукции масштаб сокращен до Х0.75 (Scheibel, 1967).
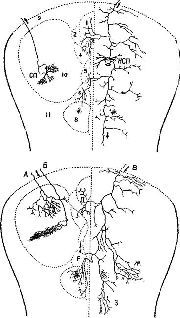
Рис. XV-4. Сравнение организации аксонных элементов специфической и неспецифической таламических систем. Верхний рисунок – таламофугальные элементы. Нейрон специфического вентробазального комплекса ядер (СП) посылает вертикально направленный без коллатералей аксон (10) к коре, исключение составляет лишь одна маленькая возвратная ветвь. Нейрон неспецифической системы (НСП) посылает аксон, который разветвляется на ростральные и каудальные отростки, богато снабженные коллатералями и идущие ипсилатерально и контр-латерально. Нижний рисунок – нисходящие таламопетальные элементы. А и Б – аксоны, идущие от корковых нейронов к комплексу различных специфических вентральных ядер. А – характерная объемная пограничная структура; Б – плоский древовидный нейрон, имеющий форму диска; В – аксон, идущий из коры к неспецифическим структурам. В диффузную, богатую коллатералями систему входят ветви, идущие к области ретикулярного ядра и к вентральному переднему ядру (Г), к неспецифическим структурам другого полушария (Д и Е), к заднему комплексу ядер (Ж) и мезодиенцефальным образованиям (3). Цифры обозначают: 1 – parataential; 2-anterior ventral; 3 – interanteromedial; 4 -r anterior medial; 5 – paracentral; 6 – central lateral; 7 – central medial; 8 -› centre median-parafascicularis complex; 9 – n. reticularis. Рисунок составлен на основании изучения ряда срезов мозга крыс и мышей до методу Гольджи (Scheibel, Scheibel, 1967).
Я подчеркнул бы еще и особое значение нейронной матрицы, на которой реализуется аминэргический подкрепляющий эффект. Эта матрица (нейронная сеть), чувствительная главным образом к холинэргическим веществам, и создает то, что в гл. XIV описывалось как компетентность мозговой ткани. Существуют данные, говорящие (рис. XV-5) о том, что адренэргическая стимуляция оказывает влияние на холинэргический механизм или по крайней мере – на активирующие и тормозящие механизмы мозгового ствола. Вспомним также, что в экспериментах Креча, Розенц-вейга и Беннета (рис. П-6) было обнаружено, что ацетилхолин участвует в эффекте влияния обогащенной среды на рост коры. Могут ли запускаться эти более продолжительные холинэргические процессы с помощью адренэргического механизма подкрепления или эти процессы протекают независимо от последнего?
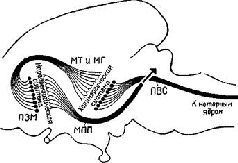
Рис. XV-5. Схема, показывающая возможные взаимосвязи между механизмами вознаграждения и наказания, составленная на основе экспериментов Маргулиса, Стайна и др. (1967, 1968) по химической стимуляции. Стимулы вознаграждения «запускают» поведение, освобождая его от тормозящих механизмов с помощью перивентрикулярной системы (ПВС), причем имеет место следующая последовательность явлений: 1) Активация медиального пе-реднемозгового пучка (МПП) с помощью стимулов, предварительно ассоциированных с наградой (или избежанием наказания) приводит к высвобождению норадреналина в амигдале и других подавляющих зонах переднего мозга (ПЗМ). 2) Тормозное действие норадреналина подавляет активность, сокращая тем самым опосредованное холинэргином возбуждение медиального таламуса и гипоталамуса (МТ и МГ). 3) Ослабление холинэргической синапти-ческой передачи в МТ и МГ уменьшает активность перивентику-лярной системы и таким образом снижает ее тормозное влияние на моторные ядра ствола мозга (Stein, 1968).
Гликман и Шифф (1967) только на основе обширного обзора результатов, наблюдаемых в экспериментах по изучению влияния электрической стимуляции мозга на поведение, дали частичный ответ на этот вопрос. В этом обзоре выделяется два типа врожденных ответов, один из них состоит в «приближении», а другой в «удалении» от объектов-стимулов. Авторы установили корреляцию между участком электрической стимуляции в стволе мозга, откуда запускается подобный тип поведения, и положительным и отрицательным подкрепляющими эффектами при электрическом самораздражении (см. рис. V-1 и V-2). Они также высказали предположение относительно некоторых механизмов, посредством которых контролирующие кортикальные влияния (тормозящие и облегчающие) могут регулировать работу центрального эффекторного механизма ствола мозга. Однако они не поняли важности проблемымодификации поведения посредством подобных контролирующих корковых влияний. Это объясняется тем, что авторы не располагали фактами, которые указывали бы на вариабельную природу механизмов, приводящих к реакции (гл. XIV), в связи с чем возникает необходимость в особом, способном к модификациям состоянии, имеющем интегративные свойства. В настоящее время анатомическая основа подобных способных к модификации интегративных процессов найдена благодаря открытию системы адренэргических афферентов, берущих свое начало в стволе мозга и достигающих апимальных дендритов. коры больших полушарий.
Факты, на которых основываются гипотезы о сущности процесса индукции, получены почти исключительно в опытах на животных. Однако недавний обзор проблемы мозговых поражений у человека, вызывающих серьезные нарушения процессов памяти, неожиданно подтвердил существование механизма, подобного тому, который здесь описан. Ограниченные поражения коры больших полушарий (включая гиппокамп) или базальных ганглиев не всегда коррелируют с нарушениями памяти. Однако г когда патологический процесс затрагивает структуры ствола мозга, у больных наблюдается «неспособность к образованию новых следов памяти, то есть антероградная амнезия (когда продолжительное заучивание не помогает)», несмотря на «сохранность активного состояния внимания, уровня бодрствования-мозга» и «нормальную способность мыслить, решать задачи и т. д.»; в таких случаях при выздоровлении интеллектуальная деятельность лишь «немного ухудшается по сравнению с нремор-бидным уровнем, несмотря на грубые нарушения памяти». Анатомически «ядра, разрушенные поражением, во всех случаях располагаются по средней линии, а именно в 2-3 мм от эпендимы. Иными словами, их расположение в зоне третьего желудочка (и сильвиева водопровода) оказывается более важным как фактор, определяющий их чувствительность, чем все другие свойства этих ядер» (Adams, 1969, р. 98-102).
Нейрофизиологи также внесли известный вклад в изучение мозговых механизмов, определяющих процессы индукции как у человека, так и у животных. Несколько лет назад Грей Уолтер (Walter et al., 1964; Walter, 1967) проанализировал медленно протекающие изменения электрической активности мозга в различных ситуациях. Он обнаружил отрицательное отклонение электрического потенциала при действии одного сигнала, подготавливающего организм к Ответу на следующий, ожидаемый стимул. В наших экспериментах (Donchin et al., 1971; рис. XV-6 и XV-7) подобные отрицательные отклонения вызывались с помощью разнообразных подготовительных состояний ожидания и место возникновения изменений потенциала зависело от типа задания, требующего этого подготовительного состояния, например: если обезьяна или человек готовится, ничего не делая, к~ выполнению ответа, максимальная CNV (контингентная негативная вариация) появляется в лобной коре; если требуется подготовиться к моторному ответу, отрицательный потенциал появляется прежде всего в моторной коре и предшествует осуществлению движения; если требуется выполнить длительное ответное действие, например продолжительный подготовительный нажим на рычаг, максимальный отрицательный потенциал возникает в со-матосенсорной коре. Когда же наконец появляется ожидаемый второй стимул, отрицательный потенциал завершается резким положительным отклонением. В других экспериментах в ситуациях выработки оперантных условных рефлексов эти завершающие позитивные электрические потенциалы были связаны с появлением подкрепляющего агента (Clemente et al., 1964; Grandstaff, 1969; Lindsley, 1969). Таким образом, CNV с ее завершающим быстрым положительным потенциалом сигнализирует об ожидании-и его завершении, о подготовительных фоновых процессах, определяющих судьбу стимула, которые обсуждались в гл. XIV.
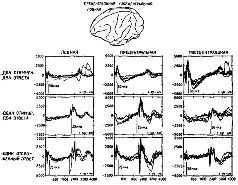
Рис. XV-6. Сравнение транскортикальной негативной вариации (TNV) – в лобной, прецентральной и постцентральной областях коры обезьяны-ре-зус во время выполнения различных заданий на ожидание. В ситуации предъявления двух сигнальных стимулов, требующих двух ответов, обезьяна должна нажимать на рычаг в пределах 400 мсек после включения светового стимула, держать рычаг опущенным в течение двух серунд и отпускать его в пределах 400 мсек после включения звука. В ситуации предъявления одного сигнального стимула и двух ответов обезьяна должна была спонтанно начинать опускание рычага без стимула, держать рычаг опущенным в течение двух секунд и быстро отпускать его при включении звука. В ситуации отсроченных реакций свет и звук предъявлялись с интервалом в 2 сек; обезьяна должна была воздерживаться от ответа до тех пор, пока не появится звук. Каждая запись – усредненные компьютером 40-50 ответов. В качестве подкрепления обезьяны получали за каждый правильный ответ маленькие кусочки пищи. Ср. с рис. XV-7 (Donchin et al., 1971).
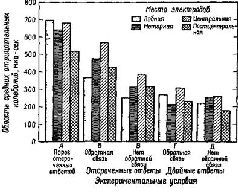
Рис. XV-7. Сравнение контингентных негативных отклонений (CNV) в различных участках мозга у человека в 5 различных экспериментах на время реакции. Во всех опытах предъявлялись 2 зрительных стимула с интервалом в 1,5 сек. В экспериментах А-В испытуемый должен был нажимать на кнопку большим пальцем ведущей руки в ответ иа второй стимул.
В экспериментах Г и Д от испытуемого требовалось нажимать на кнопку сразу же после включения первого стимула, держать ее нажатой в течение интервала между стимулами и отпускать при появлении второго стимула. В качестве обратной связи использовался красный или зеленый свет, который зажигался после каждого правильного или ошибочного ответа. В экспериментах В – Д испытуемые должны были реагировать в пределах 350 мсек. В этих условиях ограниченного времени время реакции сокращалось настолько, что испытуемые допускали до 25% ошибок. Область отрицательных колебаний оценивалась методом суммирования всех отрицательных отклонений (относительно исходной средней линии), возникающих в течение двухсекундного интервала, следующего за включением первого стимула. Электронегативные мозговые волны оценивались, усреднялись и интегрировались с помощью компьютера системы LINC-8.
Непосредственный эффект может быть получен также с помощью отрицательной (катодной) и положительной (анодной) электрической стимуляции коры больших полушарий во время научения (Stamm, 1961; Morrell, 1961). Отрицательная стимуляция повышает скорость научения, положительная оказывает противоположное влияние.
Все эти эксперименты дают убедительное доказательство того, что изменения электрического потенциала возникают в мозгу и до и во время подкрепления. Однако мы фактически ничего не знаем о тех механизмах, которые вызывают эти изменения электрического потенциала. Столь же мало знаем мы и о нейрохимических процессах, сопровождающих эти электрические изменения. Возможно, что CNV отражает или мобилизует «холинэргиче-скую компетентность», а положительные потенциалы возникаюг вследствие активации адренэргических путей или вызывает эту активацию. Тот факт, что электрические изменения связаны с подкреплением, открывает пути для изучения данной проблемы на поведенческом, гистологическом и химическом уровнях – пути, которые в течение ряда лет были полностью закрыты. Понимание подкрепления как процесса индукции выдвигает гипотезы, которые могут быть проверены в биологических исследованиях и помогут разгадать ту загадку, над которой полвека безуспешно бились бихевиористы.
РЕГИСТРАЦИЯ ПОДКРЕПЛЕНИЯ
Вспомним еще раз эксперименты с угашением (гл. III, XI), Регистрируя ряд физиологических индикаторов, мы видим, что-организм отвечает на новый стимул ориентировочной реакцией. Эта реакция угасает при повторном предъявлении стимула. Вспомним также, что после удаления миндалевидного ядра физиологические индикаторы ориентировочного рефлекса, такие, как кожногальваническая реакция (КГР), изменение частоты сердцебиений, дыхания, не обнаруживаются, несмотря на то, что-в других условиях эти физиологические ответы остаются сохранными. Как это ни парадоксально, поведенческое выражение ориентировочной реакции в этих случаях не только сохраняется, но и не угасает. Я интерпретирую этот факт таким образом, что после амигдалэктомии ориентировочный процесс не регистрируется в мозгу так, как это происходит в норме. В ситуации выработки условных рефлексов также отсутствует регистрация этих показателей, как это было обнаружено в опытах с обезьянами, перенесшими подобную операцию (Bagshow and Coppock, 1968; рис. XV-8). Если у нормальных животных наблюдается постепенное увеличение числа электрофизиологических ответов, возникающих во время предъявления стимула, и удлинение периода, во время которого появляются предвосхищающие ответы, то у оперированных животных подобных предвосхищающих КГР и увеличения их числа не наблюдается. Эти результаты говорят о том, что миндалевидное ядро интимным образом связано с организацией процессов во времени, обеспечивая процессы, участвующие в ответе на повторные события, иначе говоря, процессы серийной регистрации в памяти значимых событий. В нормальном организме существует как бы «внутреннее эхо» событий, без которого соответствующая регистрация не происходит.


Рис. XV-8. Среднее числа кожногальванических реакций, возникающих за 10-секундный период между включением света и его выключением в первых 40 и во вторых 40 пробах по каждой группе обезьян. Нормальные контрольные животные научаются предвосхищать выключение света, а у животных с удаленным миндалевидным ядром эта способность нарушается (Bagshaw and Coppock, 1968).
Таблица XV-2
Влияние удаления ампгдалы на перенос навыка иа новое но связанное с прежним задание» (см. объяснение к рис. XI-3). Обезьяны после удаления ампгдалы обучаются, выполнению второго задания, как совершенно нового, тогда как нормальные контрольные животные действуют в этих условиях на основе своего прошлого опыта (Schwartzbanm, Pribram, 1960)

То, что миндалевидное ядро участвует в процессе усвоения опыта, подтверждает и поведение животных (Douglas and Pribram, 1966). В простых задачах на различение часто повторяющихся объектов эти затруднения в регистрации событий почти не заметны. Однако, если порядок подкрепления изменяется таким образом (Barrett, 1969), что начинает подкрепляться ранее не подкреплявшийся стимул (переделка дифференцировки) или вводится новый стимул, как в том эксперименте, когда обезьяна должна была использовать опыт, полученный в другой, сходной, но не идентичной ситуации, животные, перенесшие амигдалэкто-мию, не могут решить задачу (Schwartzbaum and Pribram, 1960; рис. XV-9; Bagshow, Pribram, 1965). В таких опытах обезьяны, подвергшиеся амигдалэктомии, обнаруживают повышенную реактивность на новизну (по сравнению с контрольными животными) и эта реактивность сохраняется длительное время после того, как нормальные животные уже привыкли к ситуации. Эти результаты, а также результаты, полученные при изучении физиологических индикаторов ориентировочного рефлекса, говорят о том, что миндалевидное ядро обычно контролирует «контрастное усиление» нервных процессов, нервные механизмы латерального-торможения во входных каналах, принимающих информацию (что происходит всегда, если работает много нейронных аппаратов). Действие той системы, частью которой является амигда-лярный комплекс, состоит в подавлении эффекта разбаланси-ровки, возникающего при быстром изменении ситуации. Эти изменения ситуации приводят к смене режима работы: самоподавление возбуждения сменяется латеральным торможением в том широко распространенном в нервной системе реципрокном содружестве, которое составляет нейронный тормозный механизм- своего рода «заслонку» в системах «входа» (рис. XV-9)… Короче говоря, наличие нормально функционирующего миндалевидного ядра, по-видимому, вводит в действие «внутреннее эхо», что и приводит к регистрации соответствующих, то есть совпадающих с контекстом, событий; это происходит с помощью подавления эффекта воздействия каждого нового стимула, вызывающего значительную разбалансировку организма.
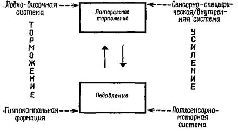
Рис. XV-9. Модель коркового контроля над афферентными подавляющими и тормозными процессами (в лобно-височную систему включается и миндалевидное ядро).
Сам факт регистрации событий посредством своего рода «внутреннего эха», удлиняющего время воздействия на мозг совпадающих событий, наводит на мысль о нейронном механизме-индукции. Работу амигдалярной системы можно понимать как усиление процесса индукции посредством предохраняющей интерференции (Pribram, Douglas and Pribram, 1969) и, таким образом, предоставление достаточного времени для того, чтобы «внутреннее эхо» могло проявиться. Только тогда, когда эти процессы интерференции контролируются, и может осуществляться завершение поведенческого акта и поведение непрерывно регулируется.
УПРАВЛЕНИЕ ПРАВИЛЬНЫМИ И ОШИБОЧНЫМИ ОТВЕТАМИ
Помимо механизма регистрации, существует еще один важнейший механизм, обеспечивающий длительное управление поведением, который мы называем завершением действия (commitment). Этот более тонкий процесс относится к тем явлениям, которые математические психологи называют «негативными случаями». Этот процесс следует за самой ранней фазой процесса регистрации и характеризуется началом активного игнорирования организмом тех явлений, которые уже происходили однажды, во теперь больше не подкрепляются. Имеются надежные доказательства того, что в этом процессе участвует гиппокамп, часть переднего мозга, примыкающая к миндалевидному ядру. Так, например, мы показали, что, если в ситуации различения число не-подкрепляемых (то есть негативных) стимулов варьирует от одного до четырех, выполнение задания нормальными животными почти не изменяется. Однако после удаления гиппокампа обезьяны значительно дольше обучаются заданию, в котором предъявляются четыре неподкрепляемых стимула, и скорость их научения пропорциональна числу подобных неподкрепляемых агентов (Douglas et al., 1969; рис. XV-10). Затем нормальные животные переходят ко второй фазе обучения: они учатся игнорировать новые неподкрепляемые стимулы. У обезьян, перенесших операцию удаления гиппокампа, механизм этой второй фазы нарушается; они не способны оценивать свои ошибки, если таковые делаются вследствие неподкрепления. Эта неспособность к оценке ошибок особенно заметно проявляется в тех ситуациях, когда требуется подавить предварительно заученные формы поведения (Douglas, 1967, Kimble, 1969). Однако изучение такого подавления выдвигает свои проблемы, которые составляют содержание следующей главы.
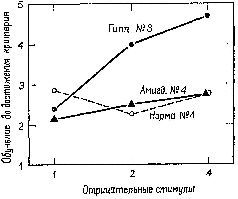
Рис. XV-10. График результатов экспериментов с изменением количества отрицательных стимулов в задачах на различение (Douglas et al., 1969).
Механизм, обеспечивающий активное игнорирование ошибок, несомненно, действует в более широком диапазоне ситуаций, чем те, которые изучаются в лабораторных условиях. Во всех подобных ситуациях должна совершаться быстрая оценка соответствия наличных стимулов прежним результатам поведения (прежним подкреплениям). Структура коры гиппокампа, столь сходная во многих отношениях со структурой мозжечка, анатомически очень подходит для этой цели (рис. XV-11). Мгновенный подсчет вероятности ошибки или рассогласования (основанный на оценке текущих состояний гомеостазиса и влияния на него результатов выполнения последней или нескольких последних задач) может, следовательно, служить в качестве контекста (фона), по отношению к которому и определяется это соответствие, и таким образом может быть оценена правильность ответа. С этой точки зрения гиппокамп можно рассматривать как механизм регуляции подкрепления и выполнения задачи, аналогичный мозжечку.
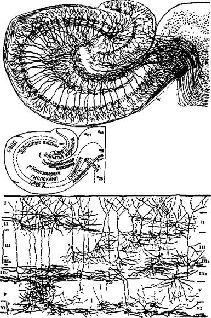
Рис. XV-11. Цитоархитектоника гиппокампальной извилины: зубчатая извилина и гиппокамп. Верхний рисунок из работы Кахала (1911), нижний – из работы Лоренте де Но (1949).
Теперь обратимся к третьей группе проблем, связанных с модификацией поведения: модификации в процессе выполнения задания. В гл. XIV рассматривалась компетентность организма в процессе научения. В настоящей главе мы уточнили нервные механизмы процессов подкрепления, обеспечивающие завершение действия, и рассмотрели мозговые механизмы, участвующие в завершении задачи. Однако завершение задачи еще не означает достижение результата. В следующей главе мы покажем, как завершенные действия превращаются в акт поведения.
РЕЗЮМЕ
Длительные изменения компетентности могут возникать благодаря процессу, сходному с тем, который стимулирует эмбриональные ткани к дифференциации. Характерные черты эмбриональной индукции и поведенческих процессов, связанных с подкреплением, удивительно сходны. Факты показывают, что адренэргические химические процессы индукции стимулируют образование РНК и синтез протеина в механизме соединений весьма разветвленной нервной системы. Механизм индукции нуждается в защите от вмешательства на всех стадиях развития, иначе не будет получен нужный эффект. Большое число мозговых процессов обеспечивает завершение серии непрерывно подкрепляемых звеньев поведенческого акта после того, как они уже введены в действие.
Глава XVI
ДОСТИЖЕНИЯ РЕЗУЛЬТАТА
ТЕОРИЯ ВЫПОЛНЕНИЯ ДЕЙСТВИЯ: ЗНАЧИМОЕ ПОВЕДЕНИЕ
В 50-х и 60-х годах XX в. классическая теория научения получила существенное развитие в результате экспериментов на животных; согласно этой теории, прочность реакции, то есть вероятность ее повторения, должна быть прямо пропорциональна силе подкрепления и его близости к сигналу и обратно пропорциональна тем усилиям, которые требуются, чтобы получить вознаграждение. Однако в некоторых случаях этого не наблюдается.
«За последнее время было собрано значительное число экспериментальных данных, говорящих о том, что эти общие предположения, лежащие в основе теории научения, не получили адекватного подтверждения в изменениях интенсивности ответа. В действительности оказалось, что в соответствующих условиях изменение вознаграждения, времени его отставле-ния и усилий, требующихся для получения вознаграждения, может дать даже противоположные результаты по сравнению с теми, которые ожидаются на основании этих предположений…» (Lawrence, Festinger, 1962, p. 6.)
С этим парадоксом часто сталкиваются те, кто работает в области физиологии. Поражение вентромедиальной зоны гипоталамуса, приводящее к повышенной реактивности, не стимулирует животное «длительно работать за пищу», напротив, такие животные дают меньше реакций, чем контрольные в тех же самых условиях опыта (лишение пищи). После всестороннего изучения условий, которые вызывают такое расхождение результатов, эти факты получили рациональное объяснение при различении двух систем: систем «запуска» и «остановки», мотивации и эмоций.
Изучение условий, приводящих к угашению поведенческих реакций, в этих случаях уменьшает трудности, с которыми сталкивается теория научения. Согласно этой теории, выполнение действия должно отражать «усилие», затрачиваемое организмом, когда он чему-то обучается. Скорость научения как мера «усилия» прямо пропорциональна легкости задания и числу и временной близости подкреплений. Поэтому интенсивность ответа – как показатель скорости научения – должна прямо отражаться на скорости угашения данной реакции в новой ситуации. В действительности же происходит обратное. Чем скорее вырабатывается навык, тем быстрее наступает угашение: скорость научения обратно пропорциональна стойкости к угашению.
Эксперименты показывают, что стойкость к угашению не просто связана с неспособностью обучаться новому, неспособностью вырабатывать новые дифференцировки, неэффективностью усвоения информации или образованием конфликтующих ответов. Разные авторы по-разному интерпретировали эти факты. Например, Д. Лоуренс и Л. Фестингер (1962) предполагали, что информация, необходимая для осуществления «конфликтных», «рассогласованных», или «диссоциированных», ответов, вызывает более стойкий навык, чем это можно было ожидать. А. Эмзель (1958) подчеркнул состояние фрустрации, которое переживает организм в подобных «рассогласованных» ситуациях. Большинство экспериментов и споров было связано с изучением состояния, которое возникает в подобных ситуациях, вызывающих рассогласование и фрустрацию. Так, было установлено, что рассогласование и фрустрация возникают в том случае, когда результат действия не приводит к таким последствиям, которые в достаточной мере его «оправдывают». Однако если вся информация, предшествующая действию, была бы налицо и был бы свободный выбор, действие вообще не осуществилось бы. Иначе говоря, рассогласование и фрустрация возникают только тогда, когда результаты поведения не совпадают с ожидаемыми, то есть когда организм не может регулировать свое поведение. Таким образом, поведение становится выражением эмоциональпого процесса, а не регулируется – через мотивационные каналы – его результатами. Но исчерпали ли мы тем самым полностью проблему?
Хорошо известно, что у морфинистов часто сила их влечения, стремления к наркотику пропорциональна величине затраты той «энергии», которая требуется для получения наркотика. Действительно, в большинстве случаев больные, у которых наблюдались симптомы отказа от наркотика, испытывают понятное отвращение к нему. В экспериментах с обезьянами-резусами обнаруживается сходная взаимосвязь между влечением и режимом подкрепления у животных (Clark and Polish, 1960; Clark, Schuster and Brady, (1961). Конечно, существенную роль играют индивидуальные различия, однако законы, регулирующие потребление морфия (например, действующие в Англии), определяются прежде всего соотношением «энергия – влечение»; в Англии наркоманы получают наркотики по медицинским показаниям и поэтому проявляют мало поведенческих нарушений. Вследствие этого контрабанда наркотиками не имела успеха и проблема наркомании была сведена к минимуму. (К сожалению, в последнее время картина несколько изменилась из-за появления нового типа наркотиков. Тем не менее эта проблема не приобрела таких размеров, как в США.)
Эти наблюдения приводят нас к тому же самому, до сих пор не разрешенному вопросу, а именно: может ли тот факт, что поведение находится под контролем эмоциональных процессов, быть единственным объяснением стойкости к угашению? Какие другие объяснения здесь возможны?
Обратимся к теории информации, которую подтверждают некоторые весьма интересные эксперименты, показывающие, как организм привыкает к наркотикам или как у него вырабатывается влечение к ним. В большинстве случаев организм способен осуществить ряд альтернативных ответных реакций. Когда поступающая информация уменьшает число таких ответов, говорят, что информация исчерпана. Именно объем информации определяет сокращение числа известных альтернатив, количество неопределенности, которое организм может освоить в процессе выбора. Как мы уже отмечали в гл. XI, теория информации связывает, таким образом, количество информации с количеством неопределенности. Термин «информация» имеет отношение к будущему (какое количество информации может быть получено?), тогда как термин «неопределенность» имеет отношение к прошлому (насколько уменьшилась неопределенность в результате действий?). Когда организм решает уже знакомую «му задачу, его действия не уменьшают эту неопределенность. Тем не менее регуляция поведения каким-то образом продолжается. Подобная ситуация наблюдается тогда, когда поведение контролируется центральным гомеостатическим механизмом. Так, у животных, испытавших когда-то голод, жажду или сексуальное влечение, действия, направленные на удовлетворение этих потребностей, имеют тенденцию к продолжению. Влечения и аффекты этих животных обнаруживают повторный и циклический характер. Каким же образом эти гомеостатические механизмы осуществляют контроль над поведением?
Ответ предполагает, что действия регулируются тем значением, которое эти действия имеют для организма. Изменение значения тех факторов, которые управляют поведением – в обычном смысле этого слова, – конечно же, обеспечивается информацией. Однако, согласно более узкому пониманию теории информации, этот процесс аналогичен процессу настройки сервомеханизма, поскольку он устанавливает ценность поведенческого акта, который стабилизирует этот процесс. Точный смысл слова «ценность» идентичен тому смыслу, который обычно придается «тому слову в социальных науках (см., например, Zimbardos, 1969). На различии между «информацией» и «ценностью» мы остановимся несколько позднее.
Р. Уолен (1961) провел следующий эксперимент с крысами, продемонстрировавший разницу между информацией и ценностью. Он показал, что у самца выбор между двумя коридорами лабиринта зависит от завершения его контактов с сексуально доступной самкой, но скорость бега в лабиринте продолжает варьировать в прямой зависимости от числа подобных контактов, несмотря на то, что животное не получает никакой новой информации о местонахождении самки после первых нескольких проб. Спепс (1956) получил сходные результаты, используя в качестве подкрепления кусочки пищи.
Результаты Уолена и Спенса показывают, что, как только начинается действие, подкрепляющие агенты обнаруживают интересные свойства: увеличение числа подкреплений в определенных пределах ведет к равномерному увеличению скорости выполнения задания. Таким образом, ценность поведения определяют подкрепляющие агенты, благодаря им корригируется поведение. Следовательно, результаты поведения могут совпадать с установкой организма, с его влечениями или же расходиться с ними. Это означает, что поведение регулируется, скорее, опережающим процессом, а не процессом обратной связи (рис. XVI-1).
Рис. XVI-1. Схема научения как процесса обратной связи и выполнения как опережающего процесса. Концепция Т – О – Т – Е.
ПАРАДОКС СИСТЕМЫ «СРЕДСТВА – ЦЕЛИ»
Тот фактор, который определяет, контролируется ли поведение процессами обратной связи или опережающими процессами, прямо вытекает из анализа отношения «средства – цель» и их парадоксального изменения. Дж. Мейс (1962) указывал на те факторы, которые вызывают такое изменение отношений.
«Что происходит, когда у человека или у животного нет необходимости работать, чтобы поддерживать свою жизнь?… Простейший случай – жизиь домашнего кота – является ярким примером изобильной жизни, более обеспеченной, чем жизнь лошади или коровы. Все основные жизненные потребности домашнего кота полностью удовлетворяются прежде, чем они проявятся. Он защищен от опасностей и непогоды. Пища и питье даются ему прежде, чем он проголодается или захочет пить. Что же ему остается делать? Чем заполнить свое время?
Мы могли бы ожидать, что получив пнщу, не приложив к этому ни малейшего труда, кот свернется в клубок на своей подушке и будет спать до тех пор, пока неотчетливые внутренние ощущения не дадут ему информацию о потребности в следующей порции даровой пищи. Однако он вовсе не спит. Он бродит по саду и лесу, ловит птенцов и мышей. Он наслаждается жизнью на свой собственный лад. Тот факт, что жизнь может доставлять удовольствие – и даже большое – многим живым существам при наличии в изобилии домашней пищи, указывает на радикальные изменения, происходящие в работе аппаратов жизнедеятельности на определенной стадии эволюционного процесса. Это и есть изменение отношений между средствами и целями в поведении. В природных условиях кот должен охотиться, чтобы жить. В состоянии изобилия он живет для того, чтобы охотиться. То же происходит и с людьми. Когда у людей нет необходимости работать, чтобы жить, у них в общем остается только два дела. Они могут «играть» и они могут заниматься искусством. Таковы два способа получать удовольствие от жизни. Правда, многие люди работают потому, что они очень любят работать, однако в этом случае слово «работа» меняет свое значение. Работа становится формой «игры». «Игра» – это деятельность, в которой доставляет удовольствие сам процесс – безотносительно к пользе или какому-либо результату. «Работа» же – это деятельность, при которой усилия направлены на производство чего-то полезного самым простым и легким способом. Отсюда и вытекает значение эргономики и изучения труда с целью уменьшения затрат и сбережения времени. В игре деятельность часто направлена на достижение бессмысленной цели трудным путем. Так, например, игрок в гольф, используя особые приспособления, направляет маленький мяч в небольшое отверстие с далекого расстояния, преодолевая умышленно созданные препятствия для того, чтобы как можно больше усложнить операцию. Это и создает обратные отношения в системе «средства – цели». «Цель» – загнать мяч в отверстие – является средством для новой цели, реальной цели – получения удовольствия от самого процесса преодоления трудностей» (Масе, 1962, р. 10, 11).
Некоторые условия, при которых происходит перестановка в системе «средства – цели», уже известны. Д. Примак (1959) провел эксперименты с выработкой оперантных условных рефлексов. Он измерял частоту ответов в значимой для организма ситуации (например, частоту актов еды), затем сравнивал эту величину с другой независимой величиной (например, частотой нажатия на рычаг). Полученные им результаты говорят о том, что подкрепление возникает всякий раз, когда ответы с низкой независимой частотой (нажатия на рычаг) совпадают – в определенных границах – со стимулами, которые управляли осуществлением ответов с высокой независимой частотой (еда).
Остроумная постановка эксперимента подтвердила эту точку зрения; один из этих опытов особенно показателен.
«Были созданы такие условия для крыс, которые вызывали в одном случае большую вероятность питья, чем бега, а в другом – большую вероятность бега, чем питья. У тех же самых животных бег служил подкреплением для питья, а питье – подкреплением для бега. Такие отношения означали, что «вознаграждался» попросту любой ответ, который сам по себе более вероятным, чем другой» (Premack, 1962, р 255; рис. XVI-2).
Отключение наказания
Рис. XVI-2. Опыты с оперантными условными рефлексами (крысы). Рисунок показывает, что число лизаний, необходимых для поворота колеса, зависит от 1) среднего числа нажатий на рычаг в серии; 2) средней продолжительности лизания; 3) средней продолжительности побе-жек. Треугольник в левой части верхнего рисунка означает исходную продолжительность лизания (Premack, 1965).
Очень интересен эксперимент, посвященный изучению активности животного в приспособлении с колесом, которое было снабжено тормозом и механизмом, прекращающим поступление жидкости.
«Преобладание питья над бегом обеспечивалось путем отключения этого механизма, освобождением колеса и предоставлением возможности чаще пить, чем бегать. Обратное достигалось остановкой колеса включением механизма, прекращающего поступление жидкости, и предоставлением возможноеги крутить колесо, то есть бегать чаще, чем пить» (Premack, 1962, р 255)
К сожалению, Примак рассматривает только связь между ответами. Ему не удалось полностью выявить те непосредственные операции, которые определяют ответ. Ответ в ситуации оперантных условных реакций указывает на то, что организм действует в зависимости от данной ситуации. Действие (как об этом говорилось в гл. XIII) подготавливается ие набором мышечных сокращений (движений) организма, а совокупностью внешних условий, вызывающих это движение. В действительности оперантный «ответ» как показатель действия является одним из компонентов этой совокупности. Следовательно, «частота ответа» – объект изучения Примака – имеет отношение не столько к быстроте движений, сколько к той скорости, с которой некоторые действительно наблюдаемые последствия этих движений могут быть зарегистрированы организмом. Например, в одних экспериментах наблюдалась обезьяна Цебус. Эта обезьяна прекрасно умела чмокать губами, кружиться в клетке и кувыркаться; все это были иррелевантпые движения, которые не должны были регистрироваться в данной ситуации как ответы, поскольку объектом изучения были манипуляции с рычагом, дверью и корзиной. Даже частные движения, включенные в эти действия, были совершенно несущественны в этих экспериментах – обезьяна могла пользоваться для выполнения действия правой или левой рукой, ногами или даже головой.
Короче говоря, оперантные условные ответы фактически представляют собой звенья какой-то последовательности, часть целого действия. Следовательно, вклад Примака в теорию научения состоит в том, что он установил, что частота осуществления последовательных действий определяет отношение «средства – цели». Таким образом, степень разнообразия действия может определяться количеством связанных с ситуацией последствий поведения, согласующихся с намерением, хотя, вероятно, частота ответа является не единственной переменной, определяющей это разнообразие. В данный момент мы можем установить по крайней мере, что при прочих равных условиях организм стремится управлять своими действиями, не просто уменьшая несоответствие между намерением и результатом поведения, а активно создавая такое соответствие. Достижение этого соответствия является не случайным: поведение строится таким образом, что последовательность звеньев действия организуется по законам иерархии – более близкие цели формируются в системе более отдаленных. По существу, эта иерархическая организация действия создает план или программу. Важность планов для организации поведения уже подробно обсуждалась в работе Миллера, Галантера и Прибрама (1965). Здесь я рассмотрю лишь один аспект этой проблемы, а именно значение планов для решения проблемы «средства – цели» и теории выполнения действия в целом.
ЗАЧЕНИЕ СТИМУЛА И ФОРМИРОВАНИЕ ПЛАНОВ
В книге «Планы и структура поведения» (Миллер, Галантер, Прибрам, 1965) обсуждались два аспекта проблемы мотивации: планы и значимость поведения. Планы строго направляют поведение, то есть выбор программ выполнения этих планов. Значимость играет несколько иную, менее специализированную роль в запуске планов, их выполнении и завершении. Если рассматривать значимость как фактор, определяющий выполнение и завершение действия, то понятие значимости поведения становится более ясным. Таким образом, эффект действия, подкрепляющий его результаты, обнаруживает две особенности: он может обеспечивать информацию, то есть сокращать количество неопределенности в ситуации или направлять поведение организма, определяя его значимость (рис. XVI-1). Рассмотрим две эти особенности подкрепляющего эффекта действия более подробно.
Во второй части книги мы показали, что сенсорные возбуждения вызывают длительные изменения в работе центральной нервной системы. Возникшие констелляции нервных процессов могут быть описаны как процессы, происходящие в сенсорном, «входе». Если эти сенсорные потоки адресуются к одной из «компетентных» систем, они превращаются в воспринимаемую информацию. Таким образом, количество воспринимаемой информации может быть определено как величина совпадения между входом информации и компетентностью систем. Это определение следует из теории информации, согласно которой количество воспринимаемой информации соответствует количеству информации, передаваемой по каналам, пропускная способность которых соответствует нервной компетентности.
В гл. XIV, однако, было отмечено, что мобилизация компетентности организма зависит от гибкого, способного к модификациям механизма, который описан в гл. XV как процесс подкрепления, ведущий к завершению поведенческого акта. Необходимость в завершении действия возникает потому, что нервный субстрат поведения (по крайней мере у млекопитающих) не содержит локальных механизмов, работа которых автоматически ведет к выполнению акта. Вместо этого существуют механизмы, запускающие частные виды активности, связанные с различными отделами нервной системы; для выполнения акта необходима интеграция, организация этих частных видов активности. Такое распределение частных функций, очевидно, свойственно всей нервной системе, и в главах VI, VII и VIII мы рассматривали влияние этого неврологического факта на процесс переработки сенсорной информации (и необходимость таких явлений, как топографическая трансформация и т. д.). Теперь я хочу добавить к этому следующий важный вывод: переработка сенсорной информации, поскольку она является подкреплением, формирует вначение стимула. Как известно, раньше все проблемы психологии сводились к двум вопросам: какова природа сходства стимулов и какова природа подкрепления. В действительности же это вовсе не две проблемы: сходство и подкрепление представляют любой одно и то же явление, хотя в одном случае оно описывается преимущественно на языке стимулов (сходство), а в другом- главным образом на языке ответов (подкрепление). Подкрепление последовательно увеличивает способность к различению стимулов и уменьшает их генерализацию; процесс различения стимулов, то есть приобретения ими определенного значения, и есть подкрепление.
Нервные процессы, участвующие, согласно этой точке зрения, в механизме научения, будут коротко описаны ниже.
«Возможно, что эта модель является прекрасным описанием того, что представляет собой подкрепление для организма. Этот нервный механизм в силу иерархической природы его избирательного контроля над его же собственными модификациями допускает изменение Образа результата в ходе последовательных актов поведения. Всякий раз, когда поступающая информация такова, что не происходит полного совпадения этой информации с Образом результата, последний модифицируется для того, чтобы включить эту информацию, и поведение продолжается. Таким образом, организм может в относительно неизменных или мало изменяющихся внешних условиях осуществлять поиск во внешней среде той дополнительной информации, которая ему необходима для уменьшения неопределенности. Эта нервная модель могла бы быть ответственной за процесс поиска информации организмом, нуждающимся в такой информации, до тех пор, пока корригирующие изменения в Образе результата ие прекратятся, то есть до тех пор, пока не будет достигнута стабильность» (Pribram, 19606, р. 8).
Акты рассматривания, принюхивания, прислушивания, ощупывания и т. п. являются, по сути дела, подкрепляющей активностью, с того момента, как организм начал выполнение задачи. Обезьяны в процессе решения задачи отказываются от пищи, которая дается им в награду за правильный ответ, потому что их защечные мешки и руки полны пищей, и все же они будут упорно продолжать решение задачи. Так, обезьяна будет засовывать в свой защечный мешок земляной орех, который она получает в награду за правильное выполнение задания, только для того, чтобы потом снова вытащить и съесть его, когда она совершит свою следующую ошибку. Главным фактором, управляющим поведением, становится не пища, а стремление получить информацию. Однако, для того чтобы информация стала фактором, регулирующим поведение, то есть чтобы информация етала значимой для организма, обезьяна должна превратить поступающую информацию в последовательные информационные звенья, связанные с ее компетентностью и стремлением к завершению задачи, подобно тому, как в пищеварительной системе заглатывание пищи предшествует ее перевариванию. Таким образом, значение стимула формируется иерархически организованным механизмом, перерабатывающим информацию.
В главах XII и XIII проводилось подробное сравнение сенсорных и моторных механизмов мозга. Чему соответствует в моторном механизме процесс формирования значения, когда информация перерабатывается с помощью сенсорного механизма? Можно предположить, что в моторном механизме происходит иерархический процесс, аналогичный тому, который характерен для сенсорных систем. Образ результата формируется не «объектами» или «интересами», а с помощью тех факторов, которые продуцируются самим действующим организмом. Эти факторы должны обусловливать то стремление к завершению действия, которое подготавливает моторные механизмы к результату. Более того, благодаря работе быстродействующего мозжечкового механизма Образ результата предвосхищает действие. Итак, известно, что: 1) нервный механизм вследствие его избирательного контроля над его же собственными модификациями допускает изменение Образа Результата в ходе последовательных актов поведения и что 2) всякий раз, когда «не происходит полного совпадения поступающей информации с Образом Результата, последний модифицируется для того, чтобы включить эту информацию, и поведение продолжается… до тех пор, пока корригирующие изменения в Образе Результата не прекратятся»; следовательно, любая последовательность предвосхищающих образов, но существу, представляет собой программу или План. Таким образом, достижение результата осуществляется в процессе выполнения действия, и, признав это, мы тем самым признаем теорию, объясняющую выполнение действия через результат. Эта теория объясняет различия в функциях подкрепляющих агентов во время научения (когда они обеспечивают информацию) и во время осуществления действия (когда они оценивают и управляют поведением). По крайней мере один класс переменных – частота ответов, – как выяснилось, играет главную роль в определении тех условий, при которых организм больше не обучается, хотя и продолжает выполнять действие по обратной схеме: вместо «средства – цель» – «цель – средства». Выполнение действия достигается благодаря иерархической природе процессов подкрепления (на языке стимулов – благодаря процессам различения): значения возникают, если информация по иерархическим законам перерабатывается в сенсорных системах, а планы и программы формируются в процессе переработки информации в предвосхищающем моторном механизме. Следующая (четвертая) часть книги содержит подробное изложение неврологии тех процессов, которые лежат в основе формирования значения и планирования, в ней описывается взаимодействие рецепторных и моторных мозговых механизмов и каждого из них с мозговыми механизмами других существ.
ЗАКЛЮЧЕНИЕ
Прежде чем продолжить изложение, подведем итог сказанному. В первой части книги обсуждался двухпроцессный кодирующий механизм работы мозга, который имеет дело с состояниями и с тем, как протекают операции на фоне этих состояний. Состояния кодируются в виде микроструктуры медленных потенциалов, которые генерируются в соедипениях нейронов. Перекодирование операций осуществляется посредством нервных импульсов, которые возникают при сдвиге этих состояний; импульсы продолжаются до тех пор, пока не будет восстановлено исходное состояние. Модификации микроструктурных синаптических процессов (память) бывают трех видов: длительные, временные и мгновенные.
Во второй части двухпроцессный механизм работы мозга рассматривался в связи с субъективным опытом. Помимо детекторов (врожденных) признаков и особого аналитического (изменчивого) механизма, нервный аппарат обладает гибкой системой распределения информации. Это свойство обеспечивается синап-тической микроструктурой медленных потенциалов и в функциональном отношении напоминает те механизмы переработки зрительной информации, которые называются голограммами.
Кроме того, во второй части было описано, каким образом с помощью билатеральной симметричной сенсорной стимуляции конструируется образ внешней «реальности», а также то, каким образом такие специфические состояния, как голод и жажда, настораживание и сонливость, активное стремление к цели и пассивность, обусловливаются работой нейронных и химических механизмов ствола мозга. Каждый механизм ствола мозга состоит из двух частей: активирующей и тормозящей; активирующая часть связана с «инстинктивными», мотивационными ощущениями, а тормозящая – с «аффективными», эмоциональными ощущениями. Однако эти инстинктивные потребности, аффекты и чувство интереса имеют более широкую неврологическую основу, чем только механизмы ствола мозга; она включает большое число контролирующих кортикальных мозговых процессов, в том числе и такие, которые контролируют аппараты ствола мозга и сенсорные системы. Здесь мы развивали «кибернетическую» теорию мотиваций и эмоций, основанную на двухпроцес-сном механизме функционирования мозга.
В третьей части мы анализировали нейронные механизмы регуляции поведения. Мы проводили различие между движением и действием. Движение, состоящее из мышечных сокращений, контролируется двухпроцессным механизмом, сходным с тем, который контролирует сенсорные процессы. Это сходство основано на существовании в рефлекторной организации системы эфферентных волокон, которая проводит сигналы от центрального нервного аппарата к рецепторам, расположенным в мышечной ткани. Эта система волокон организована по принципу кольцевого рефлекторного сервомеханизма Т – О – Т – Е, а не по принципу дуги «стимул – ответ». В процессе контроля за сокращением мышц должны, следовательно, включаться сигналы, идущие не только от сокращающейся мышцы, но и от этих рецепторов: если информация от них не поступает, у укороченной мышцы возникает «молчащий период» в возбуждении рецепторов, что мешает плавному выполнению движения, В действительности движение может контролироваться только посредством изменений в установках, которые воздействуют на спонтанные разряды мышечных рецепторов, подобно тому как работа отопительной системы в доме контролируется регуляцией температуры в термостате.
В данной части обсуждалось значение этого типа контроля над выполнением действий. Действия определяются как последовательности движений, обусловленные внешней средой. Озадачивающие результаты экспериментов, посвященных изучению роли двигательных отделов коры мозга, нашли свое объяснение, когда были получены данные, подтвердившие важность сенсорной коры для действия и показавшие, что моторный образ (представляющий собой статическую часть двухпроцессного механизма) связан с рядом факторов, вызывающих возбуждение мышечных рецепторов, а не с какой-то общей мозговой функцией, контролирующей специфику работы тех ияи иных мышечных волокон. Этот образ по своей природе подобен голограмме и принципиально не отличается от других сенсорных образов. Однако имеется одно важное различие, а именно: аппараты, связанные с действием, в отличие от сенсорных аппаратов характеризуются поступлением информации от мозжечка, где осуществляется мгновенная оценка предстоящего следующего звена действия, если действие уже запущено и не должно прерываться. Поэтому моторные механизмы кодируют результаты действия, а не ощущения или восприятия.
Нейронная основа модификации поведения обсуждалась с точки зрения результата. Способность к модификации поведения во времени зависит от того, что элементы, лежащие в основе моторного образа, компетентности организма для выполнения того или другого Акта, распределяются в нервном субстрате точно так же, как и элементы (биты информации) – в сенсорном образе. Эти элементы комбинируются, интегрируются в различные структуры, причем возникают подвижные комбинации. Процесс подкрепления делает эту интеграцию более длительной. Можно предположить, что подкрепление «индуцирует» непрерывные изменения в нейронных соединениях. Однако эти изменения могут приводить к устойчивым модификациям поведения только тогда, когда стимулируемое у организма стремление к завершению действия не разрушается вследствие отвлечения. Существуют два защитных мозговых механизма: один – регистратор подкрепления соответствия результатов поведения замыслу. Эта функция осуществляется с помощью механизма, который позволяет «внутреннему эху» освободиться от влияния последовательности новых событий. Другой механизм оценивает результаты, которые больше не подкрепляются. Эти нервные механизмы обеспечивают организму устойчивое состояние готовности к подкреплению, благодаря чему и происходит научение.
Вследствие этого прежнее поведение продолжается (хотя и остается способным к новым изменениям с помощью подкреплений) тогда, когда осуществление действия не дает ничего нового, никакой новой информации. Теория, объясняющая выполнение действия с помощью результатов, исходит из этого различения эффектов подкрепления во время научения и во время выполнения действия. Эта теория подтверждается данными бихевиори-стов относительно возможности перестановки членов в системе «средства – цель» (например, результатами экспериментов с уга-шением), а также наблюдениями над наркоманами и фактами, показывающими, что частота ответа влияет на взаимоотношение между типами поведения. Все эти факты прямо подводят нас к рассмотрению того, каким образом мозг формирует значение и контролирует планы; эти два процесса существенно зависят от достижения результата.
Таким образом, мы пришли к выводу, что исследование регуляции поведения должно быть сосредоточено не на изменении поведения организма в результате влияния внешних условий, а на изменении этих внешних условий посредством последовательного поведения, осуществления действий и достижения результата. Существует одно важное различие между восприятием и чувствами, с одной стороны, и действием – с другой. Восприятие без вмешательства действия имеет тенденцию к простой ассимиляции внешних условий организмом (в том смысле, в котором употреблял этот термин Пиаже); чувства же, даже если они находят внешнее выражение, имеют тенденцию быть простой проекцией организма во внешнюю среду (как это понимал Фрейд). Действие, скорее всего, представляет собой процесс достижения согласования (или аккомодации, по терминологии Пиаже) между восприятием организма и его чувствами.
Действие имеет и другие последствия. Благодаря действиям достигается согласование между организмами. Именно эта коммуникативная функция действия и является предметом обсуждения в четвертой части книги. В этой сфере особую проблему составляет проблема реализации человека в результатах его действий. Рассмотрим же теперь, каковы те нервные механизмы, которые обеспечивают коммуникативную функцию действий, а это необходимо нам, чтобы выяснить, что же делает человека Homo Sapiens.

