АКТИВАЦИЯ КАК РЕАКЦИЯ НА НЕОПРЕДЕЛЕННОСТЬ
Что порождает эту неуравновешенность, это подавленное состояние духа? Факты говорят о том, что даже процесс зарождения таких состояний не прост.
Переживания «интересов», мотивов (потребностей) и эмоций (аффектов) являются результатом состояний, возникающих, когда организм сталкивается с новизной – новизной, которая создается вследствие того, что непрерывно меняющийся «мир внутри нас» приходит в соприкосновение со всегда изменяющимся «миром вне нас». В результате исследований, проведен-вых в моей лаборатории с использованием техники разрушения отдельных участков мозга, удалось выделить по крайней мере два компонента ориентировочной реакции: один из них указывает на процесс исследования и опробования, а другой выявляется, когда организм фиксирует новизну. Только после такой фиксации возникает привыкание.
Результаты этих исследований были получены примерно в следующей последовательности. Несколько лет назад мы показали, что изменения темперамента и личности, легко вызываемые удалением височных долей мозга, связаны с удалением амигда-лы; эта структура височной доли, регулирующая потребности, обсуждалась в предыдущей главе (Pribram and Bagshaw, 1953; см. рис. Х-3, стр. 215). Дальнейший анализ показал, что амиг-дала участвует в различных типах поведения, которые мы обозначили как четыре фактора, развивая идеи Кеннона о реакциях «агрессии и бегства», возникающих при электрической стимуляции участков гипоталамуса (Pribram, 19606). Наши четыре фактора, дополняющие классификации Кеннона, включают пищевое и половое поведение. Поскольку между амигдалой и гипотала-мическими структурами существуют тесные анатомические связи, эти результаты казались вполне приемлемыми до тех пор, пока я не почувствовал недостаточность описательной корреляции между анатомией мозга и поведением и не попытался понять механизм этих отношений.
Если бы я удовлетворился тем, чтобы изучать поведение, как таковое, я, как и другие исследователи, поставил бы перед собой вопрос, не реализуется ли пищевое поведение, агрессия, бегство и половое поведение различными участками амигдалы. Когда были проведены эксперименты (с удалением или стимуляцией), направленные на разрешение этого вопроса, был получен, в сущности, отрицательный ответ. Но то, что я хотел узнать, касалось психологического процесса, того общего, что характеризовало эти виды аффективного поведения, поскольку одиночное разрушение (даже какой-нибудь целой анатомической формации) могло бы изменить одним ударом все эти различные типы поведения. Концепция «инстинкта» (Beach, 1955), несмотря на ее правдоподобность, по многим причинам была неудовлетворительной. Например, она не может объяснить, почему решающее влияние на изменение агрессивного поведения после удаления амигдалы оказывают социальные факторы (рис. XI-2). Это могут объяснить различные формы концепции «влечения» вследствие наличия сильных связей между механизмами амигдалы и гипоталамуса. Однако, как отмечалось ранее, такие концепции «влечения» также не в состоянии объяснить влияние разрушения и стимуляции гипоталамуса. Поэтому я решил избрать противоположный подход к проблеме и поставить вопрос следующим образом: будет ли поведение, которое явно не является врожденным или управляемым влечениями, испытывать на себе влияние удаления амигдалы?
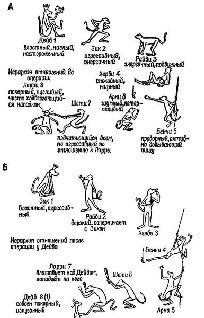
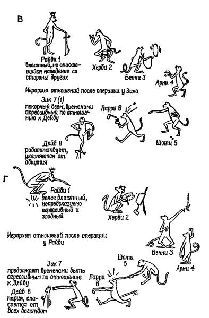
Рис. XI-2. А -иерархия господства в колония из восьми обезьян (резус)-самцов предподросгкового возраста до какого-либо хирургического вмешательства в мозг животных; Б-иерархия господства после двустороннего удаления амигдалы у Дейва. Обратите внимание, что он переместился на самую низкую ступень иерархии; В – иерархия господства после двустороннего удаления амигдалы у Зика; Г – окончательная социальная иерархия, установившаяся после двустороннего удаления амигдалы у Дейва,Зика и Райви. Следует отметить, что для Райви невозможно перемещение вниз по иерархической лестнице. Незначительные различия в размерах удаленного участка мозга у разных животных не могут объяснить различий в поведении, вызываемых этой операцией. Расхождение последних результатов с результатами предыдущих экспериментов обусловлено отсутствием агрессивности в характере Херби, занимавшего второе место на иерархической лестнице (Pribram, 1962).
Проведенные эксперименты не имели прямого отношения к исследованию упомянутых форм аффективного поведения. В сотрудничестве с Дж. Шварцбаумом (Schwartzbaum and Pribram, 1960), Мюриель Бэгшоу (Bagshaw and Pribram, 1965) и Э. Херс-том (Hearst and Pribram, 1964a, 19646) я провел эксперименты по переносу навыка. В одной серии экспериментов обезьян обучали выбирать из двух интенсивностей серого цвета более светлый. Затем во второй серии экспериментов, в которой проверялся перенос навыка, более светлая панель пары предъявлялась в сочетании с еще более светлой. Взятые для контроля нормальные обезьяны в соответствии с выработанным у них навыком продолжали выбирать из двух панелей более светлую. Обезьяны же с удаленной амигдалой во время тестовых испытаний действовали случайным образом, как если бы они решали совсем новую задачу (рис. XI-3). В другом эксперименте на перенос навыка обезьян ббучали выбирать из двух нарисованных квадратов квадрат большего размера, а затем проверяли, будут ли обезьяны выбирать большую из двух окружностей. Контрольные животные решали эту задачу, а животные с удаленной амигдалой – нет. Кроме того, в ситуации выработки оперантных условных реакций было проанализировано явление генерализации стимула. Обезьяны обучались регулировать освещение в экспериментальной камере, нажимая с разной частотой на педаль. Обезьяны с удалением амигдалы не отличались от контрольных и обнаружили такой же эффект генерализации, когда освещение в камере ступенчато менялось по сравнению с тем, на которое они первоначально были натренированы. (Это поразительное отсутствие связи между переносом навыка и явлением генерализации уже само по себе интересно, но к рассматриваемому вопросу не имеет отношения.)
Рис. XI-3. Схема эксперимента для проверки переноса навыка у обезьян. После того как обезьяны были обучены выбирать из двух одновременно предъявляемых панелей более светлую (левый рисунок), им предъявлялся тест (правый рисунок) для проверки переноса навыка. В тестовом эксперименте более светлая панель оказывалась теперь более темной, что, естественно, создавало трудности для определения того, за какой панелью скрывается награда. Нормальные обезьяны продолжали выбирать более светлую панель независимо от ее абсолютной яркости. Обезьяны с удаленной амигдалой выбирали панель случайно, показывая тем самым, что они воспринимают тестовую ситуацию как «новую» и не связывают ее с предшествующим обучением (количественные данные см. на рис. XV-9).
При выборе этих задач мы исходили из предположения, что их решение не требует участия гипоталамуса. Поскольку в обоих экспериментах при удалении амигдалы перенос навыка нарушался, один из моих выводов состоял в том, что процесс, контролируемый амигдалой, отличен от тех, которые обычно связывают с функцией гипоталамуса.
Ключом к разгадке того, что же представляет собой этот процесс, могут служить наблюдения за обезьянами, выполняющими задание с переменой стимулов. Как отмечалось ранее, обезьяны, лишенные амигдалы, не способны ни к переносу навыка, ни к выбору стимула по абсолютной величине. Во время тестовых экспериментов они вели себя, как в совершенно новой ситуации, и действовали на начальной стадии наугад (Douglas, 1966; Schwartzbaum and Pribram, 1960).
Продолжая эти наблюдения (Bagshaw and Benzies, 1968; Bagshaw and Coppock, 1968; Bagshaw, Kimble and Pribram, 1965; Bagshaw and J. Pribram, 1968; Kimble, Bagshaw and Pribram, 1965), мы показали, что в действительности при удалении амигдалы менялись реакции обезьян на новизну. Как по поведенческим, так и по электроэнцефалографическим показателям привыкание было длительным. С другой стороны, разрушение амигдалы вело к устранению висцеральных показателей ориентировочного рефлекса (КГР, изменение в частоте сердцебиения и дыхания) при отсутствии повреждения самих механизмов, ответственных за возникновение висцеральных реакций (рис. XI-4).
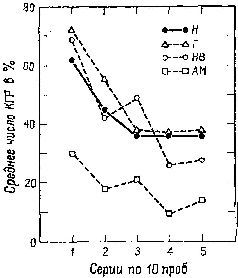
Рис. XI-4. Кривые, показывающие количество кожногальванических реакций – КГР (в %) за первые 50 предъявлений коротких тонов у нормальных обезьян (Н) и трех групп животных с двусторонним удалением гиппокампа (Г), нижневисочной коры (НВ) и амигдалы (AM) (Bagshaw, Kimble and Pribram, 1965).
Эти результаты говорят о том, что ориентировочный рефлекс на новизну проходит через две гипотетические стадии: первая, характеризующаяся поведенческими ориентировочными реакциями, «сравнивает», устанавливает новизну; вторая, связанная с висцеральными реакциями, ведет к «фиксации» в нервной системе нового раздражителя и тем самым создает основу для привыкания (Pribram, 1969). Таким образом, нарушения в поведении агрессии и бегства, пищевом и половом поведении, наблюдаемые в результате удаления амигдалы, я истолковал как результат тех затруднений, которые возникают в работе механизма, фиксирующего следы в нервной системе. Этот механизм необходим для организации поведения во времени. Проверка этой интерпретации предполагала, что поведение отсроченного чередования (выбор между двумя одинаковыми ящиками для пищи, которые заполнялись в последовательности справа – слева – справа – слева и т. д. при последовательных пробах) (Pribram, Lim, Poppen and. Bagshaw, 1966) также окажется нарушенным, и это предсказание подтвердилось.
Судя по этим результатам, амигдала выполняет гораздо большее число функций. Но это еще не все. Обращение к психологическому процессу фиксации помогло объяснить до сих пор не объяснимое наблюдение (Miller, Galanter and Pribram, 1960, гл. 14). В гл. X говорилось о больной, у которой была произведена двусторонняя амигдалэктомия. За год после операции она сильно прибавила в весе. Казалось, что этот случай давал блестящую возможность выяснить непосредственно у больной, каковы были ее ощущения, заставлявшие ее есть так много. Она обычно отвечала, что ощущает очень немногое – она не чувствует, что очень голодна, то есть она не ощущала работу контролирующей и фиксирующей системы. Такая потеря регистрации – обычное явление в клинике у больных эпилепсией, возникающей вследствие повреждений вокруг амигдалы, повреждений, которые также порождают известные феномены deja vu (неадекватное переживание чего-то как уже знакомого) и jamais vu (неадекватное переживание известной ситуации как незнакомой). Таким образом, можно сказать, что контролирующие Образы характеризуют наши переживания, касающиеся «мира вне нас», во многом так же, как перцептивные образы характеризуют его свойства.
Эти эксперименты и их анализ позволяют подойти к рассмотрению второй, более важной и широко распространенной точки зрения на то, каким образом возникают переживания мотивов и эмоций, и прежде всего те переживания интереса, побуждений и аффектов, которые зависят от механизма активации (arousal). Только что упомянутые результаты экспериментов ставят вопрос о том, при каких условиях активация (измеряемая с помощью той же самой техники и тех же критериев, которые были использованы при исследовании амигдалы) приводит к фиксации и привыканию и при каких – к их нарушению. Классический ответ, который был дан на этот вопрос (Lindsley, 1951; Hebb, 1955), состоит в том, что уровень активации определяет его конечный эффект. Сейчас к этому можно добавить, что в результате изучения ориентировочных реакций стало известно, что уровень активации зависит от организации, конфигурации ожиданий, от состояния мозга, с которыми сопоставляется новый сигнал. Таким образом, уровень активации следует понимать как степень соответствия и несоответствия между конфигурациями, степень организации или дезорганизации, а не как величину возбуждения, которое меняется само по себе (см. Hebb, 1949; Лурия, 1960). С тех пор как для характеристики активации начали привлекать понятие степени организации, стало возможным выразить уровень активации через понятия информации и неопределенности. Количество информации или неопределенности выражается числом ответов «да» – «нет», необходимым для ответа на вопрос, являются ли два объекта воспринимаемого или переживаемого поля одинаковыми или различными. Неопределенность – обратная сторона информации; термин «неопределенность» употребляется как противоположный термину «информация», которая прямо пропорциональна определенному числу бит (двоичных единиц «да» – «нет»). Чем больше это число, тем меньше неопределенность системы. Таким образом, можно было бы сказать, что такая точка зрения на теорию активации внесла в нее некоторую меру неопределенности.
Само понятие «уровень» у сторонников теории активации, судя по их анализу, весьма неопределенно. Уровень активации – это не какое-то количественное изменение интенсивности, энергетического уровня центральной нервной системы, а нарушение равновесия, организации, измеряемой количеством неопределенности (и, следовательно, количеством информации), которое характеризует системы. На нейронном уровне активация сопровождается изменением динамических структур возбуждения в ретикулярной формации ствола мозга (см. Huttenlocher, 1961) и в коре (Burns, 1968, гл. 5), а не общего количества спайковых разрядов нейронов. Даже первоначальное определение активации как электроэнцефалографической реакции основано на том наблюдении, что в ЭЭГ возникает низковольтная высокочастотная активность и что такая активность указывает на десинхронизацию. Как было отмечено Линдсли (1961), это предполагает, что во время десинхронизации нервные элементы становятся функционально независимыми друг от друга и, таким образом, способными функционировать в качестве отдельных каналов, проводящих различную информацию. Эту возросшую сепарацию можно описать как увеличение степени организации или в терминах теории информации как увеличение неопределенности (рис. XI-5).
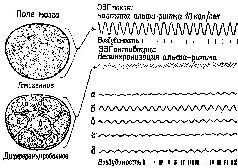
Рис. XI-5. Схема гипотетических полей мозга и связанных с ними типов ЭЭГ. Альфа-ритм покоя на частоте 10/сек характеризует состояние гомогенности, или релаксации, мозга. Десинхронизированная, активированная ЭЭГ связана с переходом к дифференцированной структуре динамических полей мозга. Активированная ЭЭГ характерна для ситуации, требующей внимания и решения задач и вообще является характеристикой более эффективного восприятия и успешной деятельности. Исходя из того, что цикл возбудимости мозга связан с подъемом и падением амплитуды волн ЭЭГ, очевидно, что при ЭЭГ покоя сменяющие друг друга периоды возбудимости и невозбудимости могли бы возникать только с частотой 10 раз в сек. В случае же десинхронизированной ЭЭГ, возникающей в результате дифференциации мозгового ноля, периоды возбудимости следуют почти непрерывно (Linds1еу, 1961).
Естественным следствием такого понимания было бы предположение, что активация представляет собой состояние, при котором независимая активность нейронных ансамблей создает свободу для «резонанса» с другими, которые, несмотря на пространственную удаленность, имеют с ними общие характеристики. Таким образом, рассредоточенная информация в состоянии активации легче направляется по нужному адресу.
Гипотеза о том, что активация является в какой-то мере механизмом неопределенности, может показаться нам привлекательной. Но это еще не все, что известно в отношении активации. Например, в эксперименте, направленном на изучение того, какое влияние в ранний период жизни оказывает ограничение движений мы обнаружили, что инактивация младенца сопровождается увеличением лабильности автономной нервной системы (Konrad and Bagshaw, 1970).
Но прежде чем будут выяснены отношения между неопределенностью и контролем за поведением, нам необходимо еще раз вернуться к теории сервомеханизмов.
Результаты Шехтера еще не получили своего объяснения: усиления того или другого переживания в результате инъекции адреналина можно достигать путем количественного изменения состояния неопределенности, вызываемого введением определенного агента; специфичность же таких чувств, как враждебность или дружелюбие и т. д., еще требует объяснения.
КОРТИКАЛЬНЫЙ КОНТРОЛЬ И КИБЕРНЕТИКА ПРОЦЕССОВ УПРАВЛЕНИЯ
Специфичность чувств предполагает, что они являются скорее мотивами и эмоциями, чем просто количественным нарушением равновесия и неопределенностью или только нарушением протекания психических процессов. И конечно, в их основе лежит работа более сложной системы, чем гипоталамический механизм. Разнообразие переживаний интереса свидетельствует о том, что в устранении нарушений равновесия, в установлении специфических типов контроля, имеющих дело со специфическими ожиданиями (см. Zimbardo, 1969), участвует множество различных процессов.
Единственная возможность у организма управлять собой – это что-то делать, воздействовать на свое окружение. Как будет подробно описано в гл. XVI, всякий раз, когда сервосистема приобретает стабильность, появляются новые чувствительные индикаторы ее состояний и вводятся новые средства для управления ими. Например, когда термостаты были впервые установлены в домах, жители вначале столкнулись с более точными показателями снижения температуры: остывание наружных стен с закатом солнца вызывало поглощение тепла этими стенами. Поэтому были установлены термостаты для наружных стен, которые обеспечили более многосторонний контроль за отоплением в домах. Тот факт, что механизмы контроля развиваются как бы по спирали, обычно упускается из вида в самых распространенных определениях биологического гомеостатического процесса, что дает основание для возражений, выдвигаемых против биологической теории сервомеханизмов.
Но организм может достигать различных форм контроля не только посредством действия. Он может управлять, осуществляя самоконтроль, то есть он может создавать на основе своей неврологической системы внутренние регуляторы, – регуляторы, которые будут приводить к восстановлению равновесия даже и без внешнего действия. Положение, из которого мы исходим, заключается в том, что эти внутренние механизмы приспособления являются тем, что мы переживаем как эмоции.
Сейчас мы располагаем надежными нейрофизиологическими данными о том, что такие внутренние регуляторы не только возможны, но что указание на них стало уже общим местом. Было проведено множество экспериментов, чтобы показать, что входные каналы организма и даже сами рецепторы являются объектом контроля со стороны центральной нервной системы. Серии экспериментов, недавно выполненные в моих лабораториях, показали наличие кортикофугальных (кортико-подкорковых) влияний так называемых ассоциативных областей на периферию: на кохлеарное ядро и зрительный тракт (Dew-son, Nobel and Pribram, 1966; Nobel and Dewson, 1966; Spinelli and Pribram, 1966, 1967; Spinelli, Pribram and Weingarten, 1965; Spinelli and Weingarten, 1966; Weingarten and Spinelli, 1966). Было показано, что при стимуляции коры на этих уровнях нервной системы изменяется электрическая активность, вызванная щелчком и вспышкой света; в этих случаях изменяется как величина, так и форма рецептивных полей нейронов зрительной системы (см. рис. XI-6).
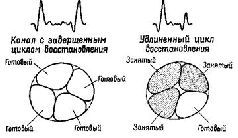
Рис. XI-6. Объяснение значения удлинения цикла восстановления возбудимости в афферентном канале после электрической стимуляции мозга. Эксперимент состоял в предъявлении пар стимулов, разделенных интервалами различной длительности. На график наносились изменения амплитуды ВП на второй стимул относительно амплитуды ВП на первый (в %) в зависимости от величины интервала, разделяющего эти стимулы (см. рис. XI-7). Предполагается, что, когда амплитуда ответа на второй стимул становится меньше амплитуды ответа на первый стимул, это означает, что афферентный канал в основном еще занят обработкой последствий действия первого стимула (показано на правой части рисунка). Обратите внимание на сходство этого рис. с рис. XI-5.
Данные об электрических ответах на вспышку света и щелчок представляют особый интерес. Когда реакция нервной системы вызывается двойным щелчком или двойной вспышкой света, амплитуда ответа на второй стимул служит индикатором длительности, в течение которой часть системы занята обработкой первого входного сигнала. Подавление амплитуды второго ответа пары указывает таким образом на более длительный процесс восстановления – на большее время проведения сигнала по каналу. Такое увеличение времени свидетельствует о десинхронизации в канале, от которого зависит реакция на повторные воздействия: меньшее число волокон готово для проведения любого другого сигнала, предъявляемого в сериях. Удлинение цикла восстановления уменьшает, следовательно, избыточность, уменьшает число волокон, проводящих тот же самый сигнал по каналу. Таким образом, в любой момент через эту систему может быть проведено большее количество информации, большее число различных сигналов при условии, что система имеет достаточный резерв избыточности, чтобы препятствовать снижению эффективности, связанному с уровнем шума. Существует надежное доказательство, что это происходит именно так (Attneave, 1954; Barlow, 1961).
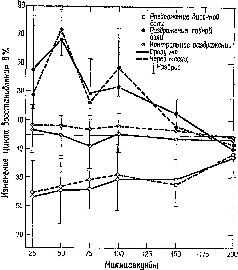
Рис. XI-7. Изменение цикла восстановления ответа афферентного канала под влиянием кортикальной стимуляции {рис. XI-6). Кора в течение нескольких месяцев подвергалась непрерывной электрической стимуляции (8 гц). Для контрольной стимуляции была использована теменная кора. Записи B1I производились немедленно после начала стимуляции и с этого времени повторялись еженедельно. На графике показаны функции восстановления при первых записях и через месяц. Вертикальные полосы показывают фактический разброс данных, полученных для каждой группы из четырех обезьян (Spinelli and Pribram, 1967).
Наши эксперименты показали далее, что стимуляция определенных участков коры височной доли (не амигдалы, а нижневисочной области) уменьшает избыточность, тогда как стимуляция другого участка коры (лобной) увеличивает избыточность в зрительной системе (рис. XI-7). Эти противоположные эффекты, по существу, либо «открывают» организм для воздействий со стороны его окружения, позволяя проводить большее число различных сигналов, большее количество информации в любой момент времени, либо, наоборот, «закрывают» входные каналы, создавая ограниченную проводимость для более узкого набора различных сигналов, для меньшего количества информации.
Результаты этих экспериментов говорят о том, что организм располагает по крайней мере двумя способами внутренне регулировать, контролировать неопределенность, но ни один из них не ведет к полной устойчивости поведения. Одним способом является увеличение скорости, с которой до него доводится информация, другим – снижение этой скорости. Один механизм делает организм «открытым» для внешних воздействий, другой ограничивает их. Управление посредством такого внутреннего контроля может, таким образом, быть достигнуто по крайней мере двумя путями: либо путем усиленного контроля за внешним окружением, либо путем доведения числа внешних воздействий до минимума и перехода к внутренним формам регуляции.
ПЕРЕЖИВАНИЯ И ИХ ВЫРАЖЕНИЕ
Теперь мы должны попытаться сделать еще один шаг в объяснении мотивация и эмоции, проблемы, которая стоит перед новой неврологией. Возможно, легче всего прибегнуть к традиционному делению психологических процессов на три класса и задать вопрос, каково различие между восприятием, действием и мотивацией – эмоцией. Сейчас имеется достаточно данных о том, что, когда организм воспринимает что-либо, он формирует внутренний образ своего окружения. Миллер, Галантер и я в книге «Планы и структура поведения» (1960, русский перевод 1965) утверждали, что, когда организм действует, он тем самым осуществляет внешнее выражение своих планов, нервных программ (сложных кодов) своего мозга. Ранее Толмен (1932) и Скиннер (1938) высказали мысль, что действие должно определяться через его внешние реакции, что оперантные условные «ответы» являются как бы метками поведения, которые можно зарегистрировать с помощью специальной записи. Теперь эти внешние проявления можно рассматривать как выражение нейронного процесса, сложившегося на основе тех условий, которые вызывают торможение неадекватных ответов, и в соответствии с ожиданиями организма, возникшими в результате прошлого действия сигналов и подкрепляющих агентов.
Если рассматривать мотивацию и эмоции в соответствии с этой схемой, становится очевидным, что они выражают отношения между восприятием и действием.
По-видимому, эти отношения состоят в следующем: когда разнообразие поступающей информации в значительной мере превосходит совокупность действий, которыми располагает организм, он чувствует себя «заинтересованным», у него возникают мотивы и он делает попытки расширить набор своих действий. Всякий раз, когда его попытка оканчивается неудачей, не подкрепляется, вызывает фрустрацию или прерывается, организм непременно переживает эмоции, или, иначе говоря, вступают в действие механизмы, управляющие саморегуляцией, самоконтролем. Более того, возможно, что эмоции возникают тогда, когда, судя по прошлому опыту, вероятность подкрепления выполненного действия расценивается как низкая.
Противоположная ситуация порождает относительно «спокойное» состояние, не вызывающее мотивации и эмоций. Если репертуар возможных действий превосходит разнообразие имеющихся восприятий, наблюдается довольно любопытный ход событий. Весь репертуар действий, которыми располагает организм, начинает все больше определять богатство той информации, которую он получает. Так происходит до тех пор, пока этот процесс не будет завершен и не произойдет обратное, пока внешняя среда не начнет определять поведение. Такая ситуация возникает только тогда, когда организм ограничивает поступающую информацию и становится «закрытой системой». Чтобы снова стать открытой системой, чтобы вызвать интерес, организм должен открыть доступ разнообразным влияниям.
Мы предполагаем, что обычно интересы, переживания мотивов (влечений) и эмоций (аффектов) возникают тогда, когда организм пытается распространить свой контроль за пределы того, что он воспринимает. Пока эта попытка кажется осуществимой, организм испытывает влияние мотиваций: когда же эта попытка кажется невыполнимой, появляются аффекты, которые можно рассматривать как механизмы саморегуляции, направленные либо на то, чтобы преодолеть состояние, не поддающееся контролю, либо на то, чтобы подготовиться к другой попытке. Мотивация и эмоция возникают одновременно. Но мотив предполагает действие, формирование его внешнего выражения, эмоция же – противоположное, то есть отказ от данного действия, уход от него. Мотивация и эмоция, действие и переживания, определяющие эффективность поведения и его аффективную окраску, являются полярными, контролирующими эффект, механизмами. Они вступают в действие тогда, когда организм воспринимает больше, чем он может выполнить.
Те термины, которыми мы обозначаем переживания, связанные с появлением интереса, могут служить как для анализа эмоций, так и для анализа мотивов, хотя некоторое различие сохраняется: так, «быть влюбленным» относят к эмоции, «любить» – к мотивации; страх как эмоция имеет свое дополнение в страхе как мотиве. Эмоции и мотивы обычно тонко разграничены. Но при слишком сильном пассивном или активном выражении заинтересованности может происходить нарушение приспособления: слишком сильная эмоция ведет к разрушению или негибкости, срыву поведения.
Наконец, несколько слов о поведенческом выражении эмоциональных переживаний.
Согласно моему анализу, выражением мотивов являются действия, тогда как выражение эмоций отражает процесс внутреннего контроля. Для окружающих (как и для самого организма) такие выражения служат коммуникативными сигналами, которые полезно учитывать. Эти аффективные сигналы указывают на заинтересованность организма, однако они предполагают, что действие в данный момент по той или другой причине невозможно или не дает нужного эффекта.
Представленная здесь точка зрения на мотивы и эмоции как на результат регулируемых корой состояний формирования Образа, которые осуществляют и контролируют отношения между системами гомеостатов и получаемой информацией, исходящей не только из внутренней среды, но и из внешнего мира, дает нам новый подход к пониманию ряда важных психологических явлений.
РЕЗЮМЕ
Эксперименты, проведенные на людях, показывают, что возникновение мотивов и эмоций связано не только с механизмами ствола мозга. Мотивы и эмоции, как и эмоциональная заинтересованность, возникают, как правило, всякий раз, когда где-либо в мозгу нарушаются отношения между организованными нейронными структурами. Кибернетическая теория мотивации и эмоций, основанная на нейрофизиологических экспериментах, показывает, что контроль над этими отношениями осуществляет кора мозга.
ЧАСТЬ ТРЕТЬЯ. НЕРВНЫЙ КОНТРОЛЬ И МОДИФИКАЦИЯ ПОВЕДЕНИЯ
«На более высокой стадии понимания поведения организма «исторические» переменные заменяются «причинными». Если мы сможем наблюдать текущее состояние организма… [и] если мы сможем непосредственно создавать или изменять это состояние, то мы сможем использовать зги знания для контроля за поведением».
Б. Ф. Скиккер
«Вместо постоянного повторения того, что с помощью подкрепления можно достигнуть контроля над поведением, я хотел бы подчеркнуть, что подкрепление может привести к … [достижению результата, основанному на] компетентности организма. И … я бы предпочел говорить… о завершении действия [как] о функции вознаграждения, связанной с достижением результата…»
Дж. Миллер
Глава XII
ДВИЖЕНИЯ
Единство всех «последовательных реакций является краеугольным камнем формирования индивидуума, сохранение целостности которого и составляет особую задачу нервной системы» (Шеррингтон, 1969, стр. 227). «Основной проблемой неврологии в значительной части, если не исключительно, является проблема превращения афферентного набора импульсов в эфферентный… все усвоенные действия, по-видимому, относятся к той же самой проблеме серийной организации во времени мышечных сокращений…» (Lashley, 1951). В основе этой части книги лежат мудрые высказывания, определяющие тот круг вопросов, с которыми должен быть связан нейрофизиологический подход к организации поведения. Суть вопроса состоит в следующем. Принцип организации моторных систем (рис. XII-1, XII-2) позволяет в каждый отрезок времени осуществлять лишь один этап действия. Однако, когда вы или я готовимся писать, или печатать на машинке, или говорить, или играть на рояле, мы уже заготовили в нашем мозгу детальное представление о том, как будут осуществляться все эти действия. Доказательством существования подобных заранее заготовленных представлений могут служить обмолвки при разговоре или досадные ошибки при игре на музыкальных инструментах, инверсии порядка слов в предложении или букв в слове, определенные отклонения в последовательности изложения мыслей и т. п. Эта проблема не является вполне ясной. Вопрос состоит в том, что представляют собой тр промежуточные процессы, которые участвуют в действии, когда одна программа превращается посредством нервных операций в другую. С этой проблемой мы уже встречались в предыдущих главах.
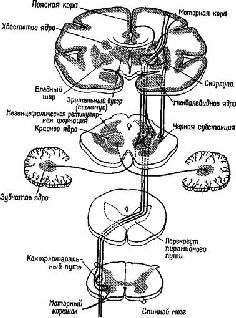
Рис. ХII-1. Схема основных связей моторной системы.
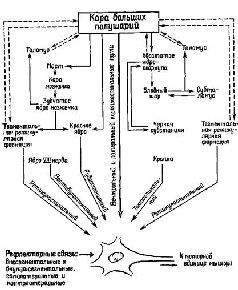
Рис. ХII-2. Некоторые нисходящие системы, воздействующие на активность «общего конечного пути», то есть на активность мотонейрона. Модификация схемы Рэнсона и Кларка (1959). Ср. с рис. ХП-1, где показано анатомическое расположение указанных структур.
В гл. III, IV, VII и VIII обсуждалось формирование программ, состояний в мозгу как функции рецепторов и системы, перерабатывающей поступающую информацию. Мы видели, что формирование подобных программ зависит от образования некой конфигурации из ряда однородных элементов, различные наборы которых размещаются в ней в том порядке, в котором они появляются в этой конфигурации, причем каждый из элементов конфигурации в отдельности представляет всю структуру в целом. В качестве примера служил «разговор» с компьютером, обладающим двоичным контролем, то есть таким компьютером, переключатели которого могут находиться только в двух позициях. Такой компьютер с трудом различает последовательности типа ААВВВА и АВВААВ, но кодирование этих последовательностей в виде рядов восьмеричной или десятичной системы делает общение с данным компьютером возможным. В равной степени важно – с точки зрения проблем, поставленных в этой главе, – что декодирование восьмеричной (или десятичной) системы в двоичную должно осуществляться прежде, чем оператор будет работать с компьютером. Операция декодирования противоположна той, которая требовалась при кодировании; теперь набор различных элементов должен быть превращен в более элементарную структуру, состоящую из однородных элементов. Это и есть проблема последовательной организации поведения, иначе говоря, – проблема единства действия во времени.
Модификация структуры должна осуществляться как по пространственным, так и по временным характеристикам, что и создает соответствующий код. Подобно тому как симфония композитора должна одновременно декодироваться в движения смычка и его остановки, определенная конфигурация в мозгу должна быть превращена в организованные по пространственному и временному принципам структуры нервных импульсов, которые регулируют степень сомкнутости голосовых связок, чтобы изменять прохождение через них воздуха. Даже при простом сгибании передней лапы собаки набор сигналов должен генерироваться таким образом, чтобы во время реципрокно связанных сокращений и расслаблений сгибателей и разгибателей все остальные части тела оставались неподвижными. Рассматривая с этой точки зрения организацию поведения, его последовательный характер следует понимать не как соединение движений в цепи, а как их дифференциацию, процесс декодирования ранее сформированной пространственной конфигурации.
Таким образом, во многих отношениях проблема поведения является обратной стороной проблемы Образа. Образ – это отражение внутри организма его окружения, а поведенческий акт – это отражение в окружающей среде того, что имеется внутри организма.Поведенческие акты – это достижение результатов; так, например, построение гнезда или написание книги совершается во внешнем мире, как оно было запланировано организмом, то есть соответствует его намерению или установке. То, как организуется поведенческий акт, и составляет предмет этого раздела нашей книги.
Вернемся еще раз к модели продуцирования слов, о которой мы говорили в начале этой книги. Предложение «Я вас люблю», произнесенное машиной, есть некий акт. Правда, здесь смоделирован только периферический голосовой акт, иными словами, электронная машина заменила собой уникальный голосовой аппарат человека. У человека настройка голосового инструмента должна осуществляться с помощью мозга. Тем не менее этот инструмент действует таким образом, что генерация даже такой сложной активности, как произнесение предложения, может осуществляться лишь посредством модификации двух мозговых процессов. Каким же образом тогда осуществляются эти модификации?
Я считаю, что организация действия есть в значительной мере управление рецепторными механизмами, содержащимися в сокращающейся ткани (в мышечных веретенах) или в сухожилиях, которые прикрепляют мышцы к костям и сочленениям. Эти рецепторы реагируют не только на сокращения мышц независимо от того, вызываются ли они внешними силами или нервными импульсами, возникающими в мозгу (через систему эфферентных волокон большого диаметра, известных под названием альфа-системы), но также и на возбуждения, приходящие к ним непосредственно через гамма-эфферентную систему волокон. Следовательно, непосредственная организация движения прямо связана с процессами, в которые вовлечены рецепторы, афферентные пути, идущие от этих рецепторов к спинному мозгу, и эффекторные пути, идущие от спинного мозга к сокращающимся мышцам и рецепторам (см. рис. V-2, V-6).
Всякий раз, когда этот процесс завершается стабильным ответом на стимуляцию, мы отождествляем его с рефлексом. В этой главе я впервые остановлюсь на некоторых интересных проблемах, касающихся осуществления рефлекса, который возникает благодаря наличию центрального контроля над рецепторами. Затем я перейду к детальному анализу мозговых механизмов управления функциями мышечных рецепторов.
СЕРВОКОНТРОЛЬ МЫШЕЧНЫХ СОКРАЩЕНИЙ
В связи с существованием непосредственного контроля со стороны центральной нервной системы над рецепторными процессами посредством гамма-системы афферентных волокон, нейронную организацию рефлекса нельзя больше рассматривать как дугу или простую последовательность «стимул – реакция». Существует множество данных, свидетельствующих о том, что и сами рецепторы контролируются центральной нервной системой – особенно те из них, которые воспринимают мышечные сокращения. Этот центральный контролирующий механизм действует как система обратных связей и процессов предвосхищения, что, по существу, и определяет корригирующие возможности сервомеханизма. Получено так много доказательств справедливости этого положения, что Р. Гранит, один из первых исследователей как моторных, так и сенсорных механизмов, вынужден был недавно заявить следующее:
«… Что касается мотонейронов, то основные проблемы связаны не с рефлексами, хотя рефлексы всегда были и все еще остаются полезным средством анализа, необходимым в данной области знания. Основные проблемы касаются настройки или установки различных механизмов, посредством которых управляются мотонейроны – рефлекторно или как-либо иначе. Существует настройка мотонейронов с помощью нейромышечной интрафузальной системы гамма-волокон [находящейся в мышечных веретенах], а также посредством особых систем нейронных связей и нейрогормонов, действующих на альфа- и гамма-мотонейроны из высших уровней нервной системы» (Granit and Kellerth, 1967).
Относительно сервоконтроля движений многое стало известно после эксперимента, проведенного в духе нейропсихологических исследований и давшего парадоксальные результаты.
«Если мышца длительное время находится в состоянии сокращения под влиянием разрядов моторных импульсов, идущих от спинного мозга, удар тока, нанесенный на моторный нерв, приведет к возникновению судорожных всплесков на тензозаписи. Во время судорожного сокращения наблюдается прекращение разрядов моторных импульсов, идущих к мышце (рис. ХII-3). Это – период молчания. Это – рефлекторный акт (обсуждение этого вопроса см. Merton, 1951), который возникает, по-видимому, вследствие прекращения разрядов от мышечных веретен во время судороги. Именно эти разряды в обычных условиях вызывают рефлекс растяжения мышцы, и при их исчезновении мотонейроны замолкают. С точки зрения сервотеории, период молчания можно интерпретировать следующим образом: электрический ток размыкает замкнутую сервосистему и тем самым возбуждает поток добавочных моторных импульсов, который и вызывает сокращение мышцы; следовательно, механизм отрицательной обратной связи отключает спинальные моторные разряды до тех пор, пока мышца посредством расслабления не восстановит своей первоначальной длины.
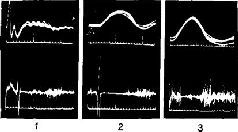
Рис. ХII-3. Периоды молчания в работе подошвенной мышцы человека, регистрация с помощью игольчатых электродов (нижняя запись). Под пяткой испытуемого закрепляется петля, и натяжение разгибателей щиколотки (верхняя запись) регистрируется при' давлении вниз на палец ноги. Время: 10 и 100 мсек; 5 суперпозиций в каждой записи; 1 – период молчания во время подергивания сухожилия, возникающего при ударе молоточком по ахиллесову сухожилию. При контакте молоточка с кожей возникает «всплеск» на кривой; 2 – запись во время рефлекса сокращения, вызванного электрическим раздражением афферентных волокон в подколенной ямке, 3 – запись во время судороги в латеральной части икроножной мышцы, вызванной стимуляцией выше этой мышцы. Обратите внимание, что подошвенная мышца сама по себе не возбуждается стимуляцией. Отметка времени (нижняя кривая на каждой записи) указывает 10 и 100 мсек (Granit, 1955).
Надо отметить, что отрицательная обратная связь не только нейтрализует эффекты изменений нагрузки или интенсивности добавочного потока импульсов, но делает работу мышц независимой от утомления или изменения синаптической возбудимости, точно так же, как усилитель, сконструированный по принципу обратной связи, не чувствителен к изменениям интенсивности поступающего тока или к напряжению электронных ламп. Стабильность – вот единственное требование, предъявляемое к системе обратных связей: в усилителе это сеть обратных связей со стабильным сопротивлением, в мышцах же это неутомляемые сенсорные органы. Как известно из работ Мэтгьюза, мышечные веретена как раз и обладают этими свойствами…
Возвращаясь к проприоцептивному механизму периода молчания, следует сказать, что мышечные, веретена прекращают свои разряды во время судороги потому, что они соединены параллельно с главными мышечными волокнами. При сокращении мышцы натяжение веретен уменьшается и они не посылают больше разрядов. Это «параллельное» соединение веретен и является ключом ко всей теории. Важная роль этого механизма состоит в том, что он дает возможность веретенам посылать сигнальную информацию о длине мышц, тогда как если бы веретена были соединены последовательно, они могли бы отвечать только на их напряжение. Таким механизмом, как серворефлекс растяжения, который столь очевидно обеспечивает поддержание определенной длины мышц, а не их напряжения, должны обязательно обладать рецепторы, сигнализирующие о длине мышц» (Merton, 1955, р. 248-259).
Существуют, конечно, и другие рецепторно-эффекторные связи, контролирующие сокращения мышц. Есть рецепторы в сухожилиях и суставах – нельзя не учитывать также и растяжение кожи вокруг мышцы и суставов как источник информации для контроля над движениями. Хотя о, рамма-системе известно больше, чем о других механизмах контроля движений, иннервация, поступающая от мышечных веретен, является, по всей вероятности, основным механизмом, на котором надстраиваются другие контролирующие механизмы (рис. ХII-4).
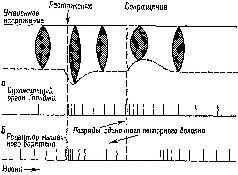
Рис. XII-4. Примеры ответов сенсорных нервных волокон, идущих от сухожильного органа Гольджи (а) и рецептора мышечного веретена (б) и обеспечивающих растяжение и сокращение мышцы. Обратите внимание, что ответ рецептора мышечного веретена (который действует посредством у-системы) выражается как в форме растяжения, так и в форме удлинения мышцы. См. текст на р и с. ХII-5, где обсуждаются эти эффекты (Granit, 1955, см . Thompson, 1967).
Как было установлено, гамма-волокна бывают двух типов. Один тип реагирующих волокон обнаруживает постоянную частоту разрядов, если мышца сохраняет постоянную длину. Другой тип, напротив, обнаруживает уменьшение частоты разрядов во время завершения динамической фазы растяжения. Мэттьюз (1964) в обширном обзоре, посвященном этой теме, приходит к заключению, что такая двойная гамма-иннервация каждого мышечного веретена «обеспечивает относительно независимый контроль за общим режимом работы и отключением сервосистемы», поскольку каждый тип контроля раздельно управляется с помощью центральной стимуляции (рис. ХII-5).
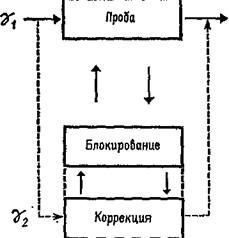
Рис. ХII-5. Двойная иннервация мышечного веретена с позиций концепции Т – О – Т – Е.
Этот анализ важен с двух точек зрения. Во-первых, он привлекает наше внимание к тому факту, что любое, даже самое простое мышечное сокращение должно управляться набором сигналов, которые адресуются либо только к мышечным рецепторам, либо идут одновременно с теми сигналами, которые поступают к сократительным элементам мышцы. Факты (см. следующие разделы) свидетельствуют о том, что некоторые области мозга, а именно базальные ганглии и передняя часть мозжечка, функционируют совместно, непосредственно управляя сократительной альфа-системой и опосредованно воздействуя на рецепторы, контролирующие гамма-разряды, благодаря чему и достигается подобная согласованность движений.
Во-вторых, управление рефлекторным сервомеханизмом легко представить себе, поскольку этот процесс разыгрывается на уровне рецепторов. Следовательно, нет необходимости в специальном устройстве типа клавиатуры фортепиано, с помощью которого импульсы, идущие от мозга, передавали бы мышцам сообщения; дело вовсе не в том, чтобы осуществить сначала одно, а потом другое сокращение, изменить длину того или иного из мышечных волокон. Существующее состояние сокращения мышечного волокна предопределяет вариативное состояние его рецепторов. Набор нервных импульсов взаимодействует с этим фоновым состоянием, изменяя режим работы мышечной сервосистемы и таким образом регулируя мышечное сокращение. Мозг все время должен посылать сигналы, организованные по пространственному и временному принципам, однако сообщения, передаваемые этими сигналами, не должны быть закодированы таким образом, чтобы прямо соответствовать сокращению или расслаблению мышцы, которое эти сообщения вызывают, поскольку эта информация уже содержится в вариативном состоянии рецептора. В следующей главе мы детально проанализируем эти сообщения, которые должны содержать подобный код.
Подведем итоги. Нейрофизиологические данные свидетельствуют о существовании прямых анатомических афферентных путей, идущих от спинного мозга к мышечным рецепторам. Наиболее фундаментальным фактом является то, что основным принципом организации даже самого простого рефлекса является не дуга: стимул – центральная нервная система – реакция, а замкнутая сервосистема в виде последовательности: проба (test) – операция (operate)-проба (test)-результат (exit) (Т-О-Т-Е). Когда с помощью центральной нервной системы рефлексы объединяются в Сложные движения, эта интеграция не может быть эффективной при посылке серий сигналов непосредственно или исключительно сокращающимся мышцам, вызывая в них мелодии, как на клавиатуре фортепиано. Подобные сигналы только бы нарушали сервопроцессы. Чтобы предотвратить эти нарушения, наборы сигналов должны передаваться либо только мышечным рецепторам, либо одновременно с другими сигналами адресоваться непосредственно к мышечным волокнам. Таким образом, интегрированное движение в основном зависит от режима работы, настройки мышечных рецепторов.
ПОДГОТОВКА К ОТВЕТУ
Теория, излагаемая в этой книге, вполне согласуется с взглядами Ф. Меттлера (1967). Последний считает, во-первых, что «тонический фон движения служит своего рода канвой… на которой разыгрывается само движение». Этот фон находится под влиянием системы афферентных импульсов, которые отражают «потенциально доминирующие» мозговые процессы. Эта «канва», это фоновое состояние формируется благодаря работе ряда периферических сервомеханизмов, контролирующих мышечные сокращения.
Далее, Меттлер признает существование «ассоциированных массивных движений (предмет данного раздела), на фоне которых вырисовываются отдельные детали…». Массивные процессы регуляции мышечного тонуса осуществляются базальными ганглиями, самыми передними структурами ствола мозга. Наконец, «на этом фоне статических и глобально действующих сил кора мозга ответственна за точность и интенсивность движений, з также за изменение скорости уже начавшегося движения и той скорости, с которой это движение усиливается и прекращается».
Статическое состояние функций мышцы называется ее тонусом. Однако тонус – это не величина спастичности или вялости сократительной ткани. Это, скорее, состояние готовности всего нейромышечного аппарата к действию, предварительное условие всякого действия.
Центральными структурами с замкнутыми системами связей, участвующими в регуляции тонической мышечной активности, являются базальные ганглии (рис. XII-6) и передняя часть мозжечка (рис. ХII-10). Поражение этих структур заметно изменяет состояние готовности мышечного аппарата: так, удаление переднего отдела мозжечка ведет к вялости, потере мышечного тонуса, а удаление базальных ганглиев – к полной обездвиженности, невыразительности мимики, маске Паркинсона и тремору конечностей, когда последние находятся в состоянии покоя. Филогенетически эти структуры мозга развивались, когда позвоночные впервые приспособились к передвижению по земле. Таким образом, эти структуры интимно участвуют в механизмах, регуляции позы, создающих то состояние готовности к ответу, тот фон, на котором может начаться дискретное действие.
Рис. ХII-6. Расположение базальных ганглиев внутри больших полушарий головного мозга (Krieg, 1966).
Регуляция мышечной активности с помощью замкнутых кольцевых связей базальных ганглиев и передних отделов мозжечка была предметом большого числа исследований, однако в этом вопросе до сих пор нет полной ясности. По-видимому, в этих процессах регуляции участвует прежде всего гамма-система, хотя, как уже говорилось выше, альфа- и гамма-влияния участвуют также в интеграции движений. Наиболее существенные характеристики системы в значительной мере зависят от количественного и временного соотношения тех реципрокных взаимосвязей, которые установились между альфа- и гамма-активностью, а также от пространственной координации повторяющихся наборов сигналов в данной системе (рис. XII-7 и ХII-8). Эти характеристики трудно определить в экспериментальных ситуациях, в которых обычно применяются анестезирующие средства, что делает бесполезными попытки воспроизвести естественные механизмы регуляции позы. Тем не менее современные знания убеждают в том, что модуляция состояний готовности в мышечном аппарате осуществляется сервосистемой «базальные ганглии – передние отделы мозжечка» прежде всего посредством угашения непрерывных колебаний, присущих периферическому сервомеханизму с отрицательными обратными связями – механизму, определяющему рефлекторный акт. Это угашс-ние колебаний, вероятно, возникает благодаря пространственной координации, торможению и совместному действию реципрокных аппаратов различных сервомеханизмов. С помощью этих механизмов тоническое фоновое состояние моторной системы приобретает изменчивость и гибкость.
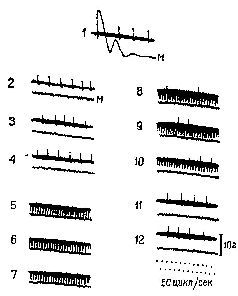
Рис. XII-7. Торможение разряда мышечного веретена при раздражении передней доли мозжечка. Верхние кривые на каждой записи показывают разряд веретена, распространяющийся вверх от черты, и артефакты от применения стимула, которые направлены вниз. Нижние кривые в каждой записи – миограммы. Децеребрированное животное. Икроножная мышца. Начальное давление – 66 гр. Миограмма записана при максимальном усилении (см. запись 12), за исключением 1-й записи, в которой сокращение (клони-ческое) в ответ на единичное раздражение электрическим током икроножного нерва выявляет молчащий период; 2-4 – контрольные записи перед стимуляцией; 5-10 – во время стимуляции мозжечка током 140 имп/сек при длительности импульса – 1 мсек, в течение 26 мсек; 5-7 – через 18-20 сек; 8-10 – непосредственно после прекращения стимуляции. Обратите внимание на падение частоты разрядов веретена от 20 имп/сек до нерегулярных разрядов порядка 5 имп/сек (Granit, Kaada, 1952; Granit, 1955),
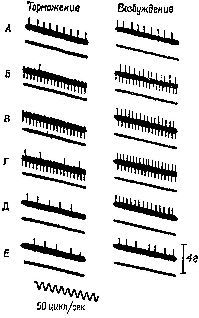
Рис. XII-8. Записи активности веретена подошвенной мышцы. Влияние стимуляции тормозных и активирующих участков ба-зальных ганглиев и других структур ствола мозга. Как и на рис. ХП-7, верхняя линия иа каждой записи показывает разряд веретена (подъем линий вверх) и артефакт от стимуляции (опускание линий вниз). А – средняя линия; В – первая запись во время стимуляции (обратите внимание на артефакты при электрическом раздражении) ; В – во время стимуляции; Г – последняя запись перед прекращением стимуляции; Д, Е – сразу же после прекращения стимуляции. На нижней кривой – миограмма. Начальное давление – 55 гр. Кошка под дозированным хлоролозовым наркозом (Eldred, Granit, Merton, 1953; Granit, 1955),
ФАКТ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ
Сервоконтроль имеет и другой аспект. Иногда сервомеханизм не только регулирует движение «на выходе», но и создает предварительную установку к следующему этапу, то есть обеспечивает то предшествующее двигательной активности состояние, которое нуждается в стабилизирующих влияниях сервомеханизма. Эти процессы предвосхищения (рис. XII-5) широко распространены в моторной системе и обозначаются как произвольные, волевые или преднамеренные движения и действия. Современный уровень разработки проблем, связанных с произвольной активностью, еще не дает возможности понять механизмы нервного контроля поведения; хорошим началом в этом отношении могут служить исследования гамма-системы. Приведем еще одно-высказывание Мертона.
«Что же происходит во время произвольного сокращения? Легко осуществить произвольное сокращение мышц при устойчивом уровне двигательной активности; эти движения совершаются даже быстрее, чем самые быстрые судорожные сокращения. Скорость возникновения небольшого судорожного сокращения мышц лишь в малой степени соответствует скорости произвольного усиления. Здесь возникает очень интересная проблема: если во время такого усиленного сокращения мышц сервомеханизм будет продолжать действовать так же, как прежде, то он явно будет способствовать противоположному эффекту. Сразу же после начала произвольного сокращения мышечное веретено будет укорачиваться и возникающее в результате этого прекращение возбуждающих афферентных импульсов будет интенсивно подавлять мотонейроны… как если бы сокращение мышцы вызывалось небольшой стимуляцией двигательного нерва. Это значит, что дополнительные моторные разряды, возникающие при возбуждении мотонейронов спинного мозга, должны вызывать гот же самый эффект в замкнутой сервосистеме, что и импульсы, возникающие вокруг этой системы в их аксонах под влиянием электрического тока. В любом случае сервомеханизм должен энергично действовать, чтобы прекратить эти механические эффекты. Если эти выводы из экспериментов верны, то это означает, что во время произвольного сокращения следует преодолеть или устранить сопротивление, оказываемое сервосистемой… необходимость возбуждения основных мотонейронов, связанных с интрафузальными мышечными веретенами посредством нисходчщих импульсов, идущих из спинного мозга, исчезает. Возбуждение целиком поступает к небольшим мотонейронам (рецепторам мышечных веретен), н сервомеханизм запускает затем в действие основные мышцы. Таким образом, в произвольном сокращении мышцы участвует тот самый механизм, который обеспечивает постоянное фоновое сокращение, а именно: сервомеханизм, поддерживающий в мышце такой же ритм, как и в ее веретенах; следовательно, мышца находится под точно таким же стабилизирующим влиянием обратных связей, как и мышечные веретена. Предположение, что возбуждение поступает по основным мотонейропам, явно ошибочно, поскольку именно этот тип вмешательства, замкнутая система обратных связей и должна нейтрализовать. Замкнутая система наилучшим образом активируется в результате изменения режима работы ее «гасящих» детекторов (null-detektor).
Эта гипотеза в известной мере подтверждается экспериментами, недавно проведенными на нервно-мышечных приборах, которыми снабжена грудпая мышца. Хант (1951), в частности, наблюдал во время рефлекторного сокращения этой мышцы разряды моторных импульсов, посылаемых к интрафузальным мышцам (рецепторам веретен), предвосхищающие появление активности в основных мотонейронах» (Merton, 1953, р. 251-253).
Таким образом, произвольные движения, как правило, начинаются с активации интрафу зальных мышечных веретен.
Исключением из этого правила является тот случай, когда движения возникают неожиданно. Вследствие сравнительно малого диаметра гамма-волокон и большой длины всего пути скорость проведения слишком мала, чтобы объяснить внезапные мышечные сокращения. Однако подобные движения относительно просты и гамма-система быстро включается в контроль над этими движениями.
Следовательно, нельзя полностью игнорировать прямой путь к основным мотонейронам. Ограниченный объем предварительной информации, которая программирует процесс «сокращения – расслабления», может быть передан по этим путям, если интервал между началом и окончанием процесса будет достаточен для того, чтобы могла включиться в действие гамма-сервосистема. Работы Дж. Бермана и его сотрудников (Taub, Bacon and Berman, 1965) установили как верхнюю, так и нижнюю границы такого рода предпрограммирования движений. Берман использовал в эксперименте обезьяну, у которой спинной мозг ла всем его протяжении был полностью деаффектирован посредством удаления дорсальных корешков. Эта операция вызывает нарушение замкнутого сервомеханизма, обслуживающего движения. Однако после деафферентации у этих обезьян все же сохранялись условные реакции (сгибания конечностей). У человека же подобная деафферентация, широко практикуемая в настоящее время, чтобы снять боли и другие сенсорные нарушения, приводит к серьезным расстройствам автоматизированных движений, выполняемых соответствующей конечностью.
Итак, управление рефлекторными и интегрированными произвольными движениями независимо от того, вызываются ли они внешними или внутренними факторами, осуществляется в основном посредством настройки рецепторов мышечных веретен, включенных в гамма-сервосистему. Центральный контроль над движениями сводится, следовательно, к центральному контролю за рецепторными процессами. Эту проблему мы уже обсуждали во второй части, более подробно рассмотрим ее в четвертой части книги. Теперь мы сформулируем парадокс, который не укладывается в рамки узкого бихевиоризма: даже относительно простые, доступные для наблюдения двигательные механизмы связаны с рецепторными процессами, регулируемыми центральной нервной системой.
БЫСТРОДЕЙСТВУЮЩИЕ МЕХАНИЗМЫ ПРЕДВОСХИЩЕНИЯ
Признав факт существования произвольных движений, мы должны ответить на вопрос: как же они осуществляются? Ответ на этот вопрос можно дать на основе анализа процессов предвосхищения, с одной стороны, и раскрытия сущности предпрограммирующих процессов, подготавливающих результаты ряда движений прежде, чем они будут реализованы, – с другой.
Вспомним, что в гл. V указывалось, что для процессов предвосхищения необходим только приблизительный план, черновой набросок тех операций, которые должны быть выполнены. Как только будет выработан такой приблизительный план движений, дальнейшую их отшлифовку может взять на себя обычная сервосистема обратных связей. На основании нейро-анатомических и нейрофизиологических данных можно допустить, что некоторые аппараты нервной системы идеально приспособлены для обеспечения этой общей планирующей функции; и, как показывают результаты поведенческих экспериментов, эти функции действительно осуществляются такими аппаратами. Центральным звеном этой управляющей системы являются филогенетически новейшая часть мозжечка, эта удивительная структура, панорама которой открывается взору лишь при определенном сечении (рис. XII-9, XII-10).
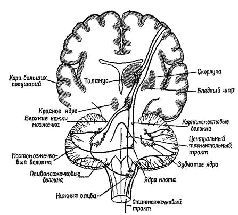
Рис. XII-9. Срединное сеченне через полушария и ствол мозга, на котором видно располошение и главные связи мозжечка Модифицированная схема Крига (1966).
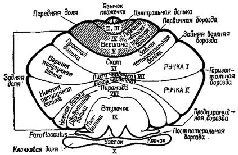
Рис. XII-10. Схематическое изображение борозд и долек мозжечка (Lar-sell, 1951; Jansen, Brodal, 1958; Angevine et al., 1961). Части мозжечка, расположенные сзади по отношению к постлатеральной борозде, составляют клочок (archicerebellum), а части мозжечка, расположенные спереди от первичной борозды, составляют переднюю дольку (paleocerebellum). Neoce-rebellum расположен между первичной и постлатеральной бороздами. Римские цифры обозначают только разделы червя мозжечка (Truex, Carpenter, 1969).
Как уже отмечалось выше, передняя часть мозжечка развилась при переходе морских позвоночных к наземному образу жизни. До этого момента мозжечок функционировал главным образом как дополнительный вестибулярный контролирующий механизм. Рыбы поддерживают ориентацию в соответствии с осью своего тела; их движения начинаются от этой основной линии. Амфибия и другие земноводные имеют дополнительную основную линию, это земля под ними, «низ», определяемый посредством гравитационных сил. Поэтому для организма становятся полезными новые вертикальные ориентиры позы и вследствие этого развивается передняя часть мозжечка. Наконец, приматы в известной мере освобождаются от гравитационной основной линии благодаря развитию двигательных навыков и приспособляются к древесному образу жизни и прямой позе. Теперь полушария мозжечка становятся доминирующими. У человека повреждение этих отделов мозга ведет к нарушению координации движений, к грубым промахам при слежении за целью и к атаксии при выполнении произвольных движений.
К счастью для нейрофизиологии, полушария мозжечка имеют удивительно гомогенное строение (рис. XII-11) и поэтому относительно просты для анализа. Их основными связями являются: 1) связи через спинной мозг со структурами, обеспечивающими афферентацию движений; 2) связи с корой больших полушарий (рис. ХП-9). Благодаря этим связям полушария мозжечка могут функционировать, как компаратор, сопоставляющий сигналы, поступающие от коры больших полушарий, с сигналами, идущими от периферии.
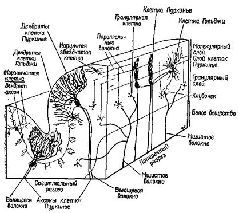
Рис. XII-11. Схематическое изображение коры мозжечка в сагиттальной и поперечной плоскостях, показывающее расположение клеток и волокон. Ср. с рис. XII-12, где дано изображение тех же клеток и волокон в поперечном разрезе (Truex, Carpenter, 1969).
Сущность этих процессов сравнения становится особенно интересной в свете последних детальных анатомических и физиологических данных (рис. XII-12). Полушария мозжечка .имеют кору, которая представляет собой как бы «прямоугольную решетку», напоминающую по своей конфигурации вертикальные колонки, обнаруженные в коре больших полушарий. Правда, в отличие от коры все эти мозжечковые решетчатые структуры фактически идентичны. Решетка построена таким образом, что, минуя один или два синапса, все поступающие в мозжечок импульсы преобразуются в тормозные сигналы. Этот невероятно развитый тормозной механизм должен служить следующим целям:
«Это исключительное превращение всех поступающих сигналов в тормозные посредством самое большее двух синаптических реле придает мозжечку с точки зрения воздействия на входной сигнал характер аппарата, заглушающего колебания в системе. Вследствие этого становится невозможным непрерывное хранение информации посредством импульсов, циркулирующих по сложным нейрональным путям, как ото имеет место в коре больших полушарий или на путях между корой и базальными ганглиями. Через 30 мсек после поступления сигнала в кору мозжечка вызванные разряды уже не регистрируются.
… остальные отделы центральной нервной системы требуют от мозжечка, по-видимому, не осуществления сложных реверберационных циклов в пределах самого мозжечка, а быстрой и четкой реакции на поступление какого-либо частного вида информации» (Eccles, Ito, Szentagothai, 1967, p. 311).
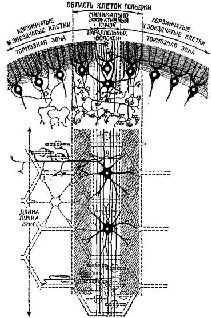
Рис. XII-12. Схема, показывающая распространение тормозных полей (затемненные области) в случае возбуждения узкого пучка параллельных волокон. Верхняя часть рисунка – поперечный разрез листа червя мозжечка; нижняя часть – вид на поверхность листа червя сверху. Предполагается, что существует «минимально эффективный пучок» одновременно возбужденных параллельных волокон, который, по-видимому, способен стимулировать организованные в ряды клетки Пуркинье, звездчатые и корзинчатые клетки. Клетки Гольджи, имеющие самую развитую сеть дендри-тов, вероятно, наиболее приспособлены к возбуждению большими группами одновременно возбужденных параллельных волокон. Следовательно, эффективная стимуляция клеток Гольджи – как отрицательная обратная связь – может перекрыть всю «входную информацию», поступающую по мшистым волокнам. Таким образом, систему клеток Гольджи можно рассматривать как «фокусирующее» устройство, которое ограничивает – или усиливает – активность гранулярных нейронов (через систему параллельных волокон) в относительно узких группах (Eccles, Ito and Szentagothai, 1967).
Быстрое стирание содержания записи предполагает, что последовательность операций сличения выполняется тоже довольно быстро. Эта особенность системы выходит за рамки обычного сервомеханизма обратных связей, в котором для длительного поддержания устойчивого состояния требуется постоянная информация о результатах движения. Т. Рач в своей работе, опубликованной в «Экспериментальной психологии» (1951), правильно сформулировал эту проблему, причем значительно раньше, чем были собраны соответствующие факты.
«Процесс циркуляции возбуждения по замкнутому кругу от коры больших полушарий к [нео]-мозжечковым структурам выполняет функцию не столько устройства, корригирующего ошибки, сколько составляет часть механизма, который может мгновенно осуществлять предвосхищение влияния. Подобный замкнутый контур, хотя и не информированный о конечных результатах движения, мог бы, как говорится, «вчерне» программировать движение и таким образом сократить досадные издержки, связанные с коррекцией движения посредством аппарата обратных связей, который использует поступающую извне информацию» (Ruch, 1951, р. 205).
Основная функция мозжечка, связанная с объемом регистрации, заключается в том, чтобы составить схему движения без детальной его разработки. В действительности такое кратковременное состояние, которое аналогично тому, что создается порой больших полушарий, формируется на фоне спонтанной активности.
«Таким образом, следует предположить, что даже в условиях минимального сенсорного «входа» существует состояние динамического равновесия в уровне активности различных типов нейронов мозжечка … Отрицательный образ обобщенного результата работы коры мозжечка формируется в ходе процесса, аналогичного созданию скульптуры из камня. Форма, обладающая пространственно-временными признаками, достигается постепенно посредством набора тормозных импульсов, подавляющих «бесформенные» фоновые разряды нейронов, расположенных на более низких уровнях, подобно тому как вечные формы в скульптуре создаются путем очень избирательного удаления резцом всего лишнего из первоначально бесформенного камня» (Eccles, Ito, Szentogothai, 1967).
Подробнее я остановлюсь на этом приборе и на его аналоге-в коре больших полушарий в гл. XIII. Здесь же мы должны только констатировать, что механизмы предвосхищающего' схематичного планирования движения можно рассматривать в качестве эквивалента быстродействующего компьютера, оценивающего конечный результат последовательной серии движений. Сходство между голограммой и процессом формирования образа заставляет предположить, что здесь имеет место процесс типа преобразования Фурье, однако и об этом речь будет идти в следующей главе. Во всяком случае, результаты предварительного расчета движений всегда немедленно поступают как к периферической мускулатуре, так и в моторную область коры больших полушарий через различные мозжечковые пути. С инженерной точки зрения подобный быстрый расчет ожидаемого «выхода» в настоящее время считается исключительно полезным приспособлением (Kelley, 1968). По-видимому, система «мозг-поведение» уже давно овладела им.
РЕЗЮМЕ
Проблема мозговой организации поведения во многих отношениях является обратной стороной формирования Образа (восприятий и чувств). Вследствие повсеместного существования в нервной системе механизма обратной связи (и механизма предвосхищения) контроль над движениями достигается не непосредственно с помощью сигналов, адресующихся к сокращающимся мышечным волокнам, а посредством сигналов, адресующихся к рецепторам, которые регулируют мышечное сокращение. Таким образом, мышечные рецепторы становятся частью гармоничной системы (сервомеханизма Т-О-Т-Е), назначением которой является регуляция мышечных реакций в ответ яа изменения внешней среды. Короче говоря, нервная регуляция поведения осуществляется посредством воздействия на ре-щепторные процессы, а не через непосредственный контроль мышечных сокращений.
Глава XIII
ДЕЙСТВИЯ
МОТОРНАЯ КОРА
Четкие, избирательные движения, лежащие в основе поведения, осуществляются моторной корой. Эта кора, расположенная в прецентральной извилине полушарий (рис. XIII-1), связана с базальными ганглиями и мозжечком. Каковы же функции этой моторной коры?
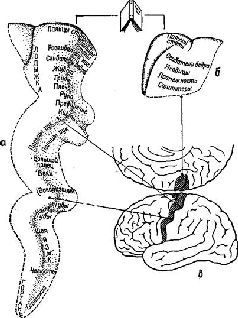
Рис. XIII-1. Представительство моторики в прецентральной коре человека, в – схема поверхности левого полушария мозга, показывающая расположение прецентральной извилины; о – увеличенная прецентральная извилина, видимая в том же ракурсе, что и на рис. в; б – медиальная поверхность прецентральной извилины. В действительности у различных людей наблюдается значительное перекрытие зон и их вариативность, однако в целом эта схема сохраняется. Схема составлена на основе данных Форстера, Пенфилда, Болдри и др. по электрической стимуляции лрецентральной коры и наблюдений за возникающими при этом движениями (Krieg, 1966).
Уже на протяжении целого столетия идет горячий спор об особенностях организации центральной моторной системы. Некоторые авторы (Woolsey, Chang, 1948) придерживаются точки зрения, согласно которой в моторной зоне существует анатомическое «точка в точку» представительство мышц и даже волокон и моторная кора играет роль клавиатуры для всякой другой церебральной активности и прежде всего для всех произвольных действий. Напротив, другие авторы (например, Phillips, 1965) указывают, что рецептивные поля соседних корковых нейронов охватывают широкий набор мышц, хотя большинство активных мышц, зарегистрированных при раздражении какого-либо одного участка коры, связано лишь с определенным суставом. С таким представлением согласуется тот факт, что электрическое раздражение моторной коры вызывает движения, интегрированные в определенную последовательность мышечных сокращений, и что движения, вызываемые отдельным возбуждением, частично зависят от состояния мозга и положения конечностей, что и отражается на их реакциях при стимуляции. Эти нейрофизиологические данные всегда интерпретировались таким образом, что в моторной зоне коры представлены движения, а не мышцы. Несколько лет назад я еще раз вернулся к этому спору, повторил многие критические эксперименты и накопил новые наблюдения с помощью некоторых дополнительных технических устройств (Pribram, et al., 1955-1956).
Результаты этих экспериментов и наблюдений показывают, что основная роль моторных зон коры заключается не в контроле за индивидуальными мышцами или специфическими движениями. Моторная кора, скорее, играет более важную роль в организации действия – действия, определяемого не с точки зрения участвующих в нем мышц, а с точки зрения обеспечения внешнего выражения психологической установки или плана. Это навело меня на мысль, что центральный моторный механизм является в некотором смысле подобием сенсорной системы и что поражение моторной коры вызывает «скотомы в действиях» по аналогии со скотомами в зрительном поле, возникающими при поражении зрительной коры. Появляется все больше данных в пользу этой точки зрения. Вначале я столкнулся со следующим анатомическим парадоксом. Клетка моторной коры получает импульсы через дорсальный та-ламус – структуру ствола мозга, которая расположена дорсально от коры (о чем говорит и само ее название). Следовательно, эта структура аналогична дорсальной части спинного мозга, которая, как показали Белл (1811) и Мажанди (1822), является «сенсорной» частью периферических нервов (рис. XIII-2). Почему же моторная кора должна быть так тесно связана с сенсорными структурами противоположной стороны тела?
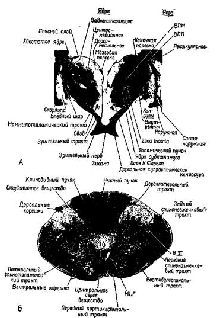
Рис. XIII-2. А – фотография поперечного сечения мозга через; таламус, гитлаламус и базальные ганглии на уровне зрительной хиазмы. Гб означает гипоталамическую борозду в стенке третьего-желудочка. ВПМ и ВПЛ означают вентральное постмедиальное и вентральное постлатеральное ядра таламуса. Методика окрашивания миелина по Вейгерту; Б – фотография поперечного сечения самых верхних отделов спинного мозга месячного ребенка. Методика окрашивания миелина по Вейгерту. Отметьте сходство в расположении и конфигурации задних рогов спинного мозга и задних отделов таламуса, а также передних рогов спинного мозга и базальных ганглиев (и субталамуса) (Truex, Carpenter, 1969)
Этот анатомический парадокс особенно ярко проявился в ходе экспериментов, в которых изменения электрических потенциалов вызывались в коре посредством электрической стимуляции периферических нервов. Однажды совершенно случайно было замечено, что такого рода изменения потенциалов возникают в моторной коре. Эти явления слишком противоречили известным фактам, чтобы сразу поверить им. Однако в действительности оказалось, что они были получены и другими авторами, хотя ни один из них не решился их опубликовать и имелось лишь одно упоминание об этом как о возможном артефакте, да и то – в сноске. Между тем наши эксперименты (Malis, Pribram and Kru-ger, 1953, рис. XIII-3) и работы других авторов (Albe-Fessard, 1957; Penfield, Boldrey, 1937) показали, что входные импульсы от периферии действительно достигают моторной коры, что они проходят через дорсальный таламус, а не через уже давно изученные сенсорные зоны коры, такие, как соматосенсорная кора, примыкающая к моторной, и что эти импульсы не зависят от мозжечка. Кроме того, было показано, что входные импульсы возникают не только в тех нервных волокнах, которые иннерви-руют мышцы, но также и в тех, которые связаны исключительно с кожей.
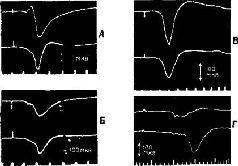
Рис. XIII-З. А - ответы в коре больших полушарий, вызванные стимуляцией седалищного нерва, перед удалением постцентральной коры и мозжечка. Верхняя кривая – ответь в постцентральной коре; нижняя – ответы в прецентральной коре. Время – 10 мсек. Б - ответы сразу после удаления обоих полушарий мозжечка; В - ответы после дополнительного удаления передней доли мозжечка; Г - ответы после дополнительного удаления обеих постцентральных извилин. Заметьте, что в постцентральной области теперь регистрируются только ответы белого вещества мозга (Pribram, Kruger, 1953).
Факты, полученные в других наблюдениях и экспериментах, также подтверждают сенсорную природу моторной коры. Обезьяны и человек, у которых была удалена моторная кора, могли выполнять всякого рода движения при соответствующей организации опыта. Инвалиды войны, у которых уже много лет были парализованы руки вследствие поражения моторной коры, находясь в запертой комнате, при пожаре поворачивают дверную ручку парализованной рукой. Когда обезьянам после удаления моторной коры показывали кинофильмы в замедленном темпе, на которых демонстрировались другие обезьяны, неуклюже выполнявшие заученный ряд движений, чтобы открыть щеколду на ящике с орехами, то они без затруднений делали точно такие же движения рукой и пальцами, которые возникают у них при лазании по решетке клетки или при уходе за собой. Затруднены были только некоторые акты, некоторые движения, но эти затруднения не были связаны со специфическим набором необходимых в данном случае движений (Pribram et al., 1955-1956). Каким же образом моторная кора участвует в превращении движения в действие?
ОБРАЗ РЕЗУЛЬТАТА
Чтобы ответить на вопрос о том, как движение превращается в действие, надо понять, какого рода процессы, связанные с формированием образов, должны происходить в моторной коре и как образ становится мгновенным Образом ожидаемого результата, который содержит в себе всю входящую и выходящую информацию, необходимую для того, чтобы осуществилось следующее звено поведения. Чтобы проверить эту гипотезу, необходимо показать, что, подобно тому как в зрительной коре кодируется незрительная информация (например, образ правой или левой стороны панели, успеха и ошибки), так и моторная кора должна кодировать иную информацию, чем движения. Вероятность того, что такое кодирование должно происходить, велика, так как значительная часть информации, поступающей в моторную кору, исходит из коры мозжечка – структуры, имеющей обширные связи со всеми сенсорными рецепторами.
Но каким образом кора мозга становится местом моментального возникновения Образа ожидаемого результата? Каким образом входная информация, поступающая от периферических структур, участвующих в выполнении движения, организуется так, чтобы предвосхитить результат движения, а не просто служить регистратором компонентов движения? Классические эксперименты, выполненные в 30-х годах Н. А. Бернштейном (1966) в Советском Союзе, помогают ответить на этот вопрос. Наряду с последними сведениями о значении гамма-эфферентного контроля над движением теперь начинает проясняться и этот важный и весьма интригующий аспект организации действия. Итак, каким образом организм создает предваряющий Образ, предвосхищение последствий своего поведения?
Н. А. Бернштейн проанализировал выполнение таких действий, как ходьба, бег, работа молотком, напильником или печатание на машинке, с помощью кинематографической записи. Испытуемые были одеты в черные костюмы с белыми полосами, чтобы выделить суставы конечностей (рис. XIII-4). В результате снятый фильм представлял собой последовательно развертывающийся рисунок движений, состоящий из отдельных колебаний, которые могли быть математически проанализированы. Бернштейн обнаружил, в частности, что любое ритмическое движение может быть представлено посредством тригонометрической функции и что каждое следующее звено подобного движения может быть предсказано с точностью до нескольких миллиметров по формуле суммы трех или четырех гармонических колебаний – по так называемой тригонометрической сумме Фурье (рис. XIII-5). Из того факта, что эти компоненты движения могут быть описаны в математических терминах и охарактеризованы как «временная голограмма», следует, что именно такого рода Образы, а не предполагаемый клавиатурный механизм, организуют, вероятно, поведение. Следовательно, совокупность явлений, происходящих в моторной коре, можно рассматривать как отражение своего рода голографической трансформации или проекции предшествующих внешних сил, определяющих действие.
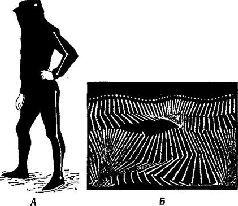
Рис. XIII-4. А – испытуемый в черном костюме с белой тесьмой; Б – кинематографическая запись ходьбы. Направление движения слева направо. Частота-26 экспозиций в секунду (Н. А. Бернштейн, 1966).
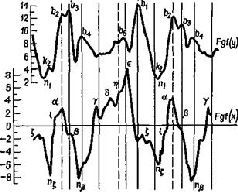
Рис. XIII-5. Кривые усилий в центре тяжести бедра при нормальной ходьбе. Наверху – вертикальные компоненты; внизу – горизонтальные компоненты (Н. А. Берн-штейн, 1966).
Бернштейн резюмирует эту точку зрения следующим образом:
«Совершенно очевидно, что каждый из этих вариантов движения (например, рисование круга, маленького или большого, перед собой или сбоку, на горизонтальной бумаге или вертикальной доске) требует совершенно иной мышечной формулы, более того – зачастую совершенно иного ассортимента включенных в дело мышц. Близкая к одинаковости легкость и точность выполнения всех этих вариантов свидетельствует о том, что все эти варианты определяются в конечном счете одной и той же верховной руководящей энграммой, по отношению к которой энграммы размера и положения занимают подчиненное место. Те энграммы, которые определяют далее мышечную структуру каждого из конкретных вариантов, лежат, очевидно, еще ниже…
Отсюда приходится сделать вывод, что верховная энграмма, которую можно было бы назвать «энграммой данного топологического класса», уже чрезвычайно структурно далека (а потому, вероятно, и локализационно далека) от какого бы то ни было сродства с мышечно-суставными схемами: она целиком геометрична, представляет собой очень абстрагированный моторный образ пространства. Это заставляет думать, пока еще в порядке гипотезы, но очень настойчиво напрашивающейся, что область локализации этих верховных моторных энграмм обладает и сама топологической упорядоченностью по типу внешного пространства или моторного поля (во всяком случае – отнюдь не по типу мышечно-суставного аппарата). Иными словами, есть немалые основания полагать, что в верховном моторном центре мозга (очень возможно, что это есть кора больших полушарий) локализационно отображено не что иное, как какая-то проекция самого внешнего пространства в том виде, в каком оно моторно дано субъекту. Эта проекция, по всему предыдущему, должна быть конгруэнтной с внешним пространством, но конгруэнтной только топологически, а совсем не метрически. Этим соображением вполне устраняются все опасения насчет возможности компенсирования перевернутой проекции сетчатки… и многие другие в том ше роде. Конкретизировать в большей мере, как именно возможна такая топологическая проекция пространства в центральной нервной системе, не представляется сейчас возможным, хотя думается, что это затруднение есть для физиологии только вопрос времени. Надо только оговориться, что топологические свойства проекции пространства в центральной нервной системе могут на поверку оказаться очень неожиданными и странными: не следует надеяться увидеть в головном мозгу что-либо вроде фотографического снимка пространства, хотя бы и очень деформированного. Самый же факт наличия в верховном отделе центральной нервной системы проекции именно пространства, а не мышц и не сочленений представляется мне сейчас более вероятным, чем что бы то ни было другое» (Бернштейн, 1966, стр. 69-70).
Как мы вскоре увидим, предположение Бернштейна о том, что в моторной коре представлена топологическая проекция пространства, оказалось ошибочным. На самом деле в моторной коре предстазлены силы, возбуждающие мышечные рецепторы. Однако это не умаляет значения идеи Бернштейна о том, чта в коре больших полушарий кодируются свойства внешней среды а не расположение мышц и суставов.
МОТОРНЫЙ МЕХАНИЗМ
Как мы уже говорили, наблюдения Бернштейна показывают, что единство действия может быть достигнуто независимо от частных видов движения или количества сокращений каких-либо частных мышц или групп мышц. Действие может осуществляться лишь потому, что движения согласуются с «полем внешних сил», вовлеченных в действие. Как мы уже отмечали при рассмотрении гамма-эфферентной сервосистемы, существенные изменения в этой системе компенсируются указанным механизмом, так что постоянно сохраняется состояние готовности к движению. Таким образом, стабильность состояния достигается за счет специальных приспособлений и компенсаций внешних сил, а представительство в коре больших полушарий этого механизма включает эти параметры. Фактически центральный нервный механизм и состоит в основном, из этих приспособлений и компенсаций; следовательно, центральное представительство должно быть организовано таким образом, чтобы быть «зеркальным отражением» поля внешних сил (аналогичный вывод был сделан и в гл. XII).
Явное доказательство того, что нейроны моторной коры – особенно те, от которых идут пути на периферию, – действительно чувствительны к воздействиям, вызывающим мышечные сокращения, было получено в серии других остроумных опытов. В этих экспериментах Э. Эвартс (1967, рис. XIII-6) обучал обезьян работать рычагами, к которым были подвешены различные грузы, чтобы затруднить движения. Когда обезьяны научались двигать рычаг, в момент выполнения задания регистрировалась активность нейронов, расположенных в моторной коре. Опыты показали, что эти нейроны активируются до внешних признаков начала движения (по миографической записи). Тщательный анализ установил, что эта электрическая активность зависит от величины необходимых усилий, а не от величины смещения рычага. Более того, нейроны реагировали главным образом на изменение требуемой силы, хотя не исключено, что некоторая активность связана и с величиной необходимого усилия (рис. XIII-7).
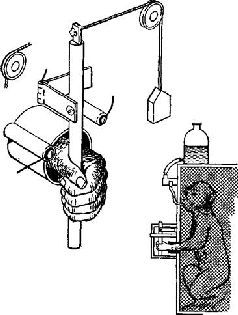
Рис. XIII-6. Аппарат для изучения поведения обезьяны с одновременной регистрацией моторного эффекта в клетках прецент-ральной коры. Левая рука обезьяны просовывается через отверстие в прозрачной перегородке, прикрепленной к передней стенке клетки. Чтобы получить фруктовый сок в качестве вознаграждения, обезьяна должна схватить вертикальный стержень, присоединенный к шарниру, и передвигать его взад и вперед между двумя упорами. Чтобы сработал соленоидный приемник, который «выдает» фруктовый сок, интервал между соприкосновениями стержня с контактами должен составлять 400-700 мсек и предварительное движение в другом направлении также должно совершаться в это же время (Evarts, 1968; см. Милнер, 1973).
Тот факт, что моторная кора предвосхищает параметры усилия, очень важен. Поскольку рефлексы организуются сервомеханизмами, центральное представительство этих рефлексов составляет не запись длины или напряжения мышц, а запись параметров приспособлений и компенсаций, соответствующих тем изменениям внешних сил, которые вовлечены в движение. Вследствие конвергенции этих процессов в центральном представительстве движений формируется образ не только прежних и текущих изменений внешних сил, воздействующих на систему, но – благодаря быстродействующим расчетам движений, совершающимся в мозжечке, – и тех изменений, которые должны произойти при продолжении двигательной активности.
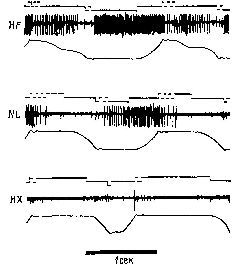
Рис. XIII-7. Активность клетки моторной коры обезьяны в условиях, изображенных на рис. XIII-6. Показана активность клетки при трех видах нагрузки. Использовались следующие три вида нагрузки: 400 г на сгибатели (верхняя запись), 400 г на разгибатели (нижняя запись) и без нагрузки (средняя запись). Видно, что активность нейрона значительно возрастает при нагрузке на сгибатели, то есть при увеличении силы сгибания. Напротив, клетка почти полностью «замолкает» в те периоды, когда движение совершается при нагрузке на разгибатели (Evarts, 1967).
Таким образом, моторную кору можно рассматривать как сенсорную по отношению к действию. Она участвует в пространственной модуляции состояний готовности благодаря своим связям с базальными ганглиями и с механизмом быстродействующего-расчета параметров конечного эффекта действия, осуществляющегося при участии замкнутой мозжечковой системы. Формирование Образа результата зависит – как и всюду в коре – от того, что мы называем «нейронным топографическим представительством», то есть «от в высшей степени избирательного протекания активности в горизонтально расположенных корковых сетях, которые возбуждают и тормозят кортикофугальные нейроны» (Phillips, 1965). То, что это моторное представительство действительно во многих отношениях сходно с тем, которое формируется в сенсорных зонах мозговой коры, может быть установлено при исследовании рецептивных полей нейронов. Результаты этих экспериментов (Welt et al., 1967, табл. XIII-1) показали, что «сенсорная конвергенция в моторной (или сенсорной) коре накладывается на топографически сходные выходные структуры в виде радиальных рядов, диаметр которых, как оказалось, равен 0,1-0,4 мм. Таким образом, нейроны с фиксированными локальными рецептивными полями образуют радиально ориентированную структуру (референтную систему) по отношению к одним и тем же периферическим выходам… Вперемежку с этими клетками, имеющими локальные поля и составляющими 3/4 общей массы клеток, расположены нейроны с широкими, вертикально ориентированными или лабильными полями, которые перекрывают локальные поля» (стр. 285). Здесь, в моторной коре обнаружены явления, которые еще надо найти в зрительной коре (Теперь эти факты уже получены (Spinelli, Pribram and Bridgman, 1971).).
Таблица XIII-1

Сравнивались в простом рандомизированном плане антидромные латентные периоды реакции 88 корковых клеток, сгруппированных по типам рецептивных полей -Ответы группы «лабильных» клеток отличались от ответов обеих групп ‹Фикси1 рованных» клеток более чем на 1% not-критерию в предположении равенства групповых дисперсии.
s. d.- стандартное отклонение
S. Е.- средняя ошибка
См. описание сходства роли лабильных клеток и чувствительных клеток узко-яаправленного действия в корковых структурах Вернера (рис, VH-7), основанное/на -концепции Т-О-Т-Е (Welt et. al. 1967).
Остается сделать еще только один шаг в анализе работы моторной коры, а именно указать на то, что уже прекрасно выполнено Родиеком в отношении сетчатки: выяснить, какая именно трансформация точно отражает взаимоотношения между корой и периферией.
Итак, контроль над поведением со стороны нервной системы достигается главным образом посредством воздействия на рецеп-торные функции. На рефлекторном уровне чувствительность рецепторов к грузу возбуждает и направляет приспособительный контрпроцесс в сервомеханизме. Сумма таких приспособительных процессов и составляет фоновое тонически устойчивое состояние, на котором возникают новые приспособительные действия. Приспособления широкого диапазона, например изменения позы, контролируются экстрапирамидной системой мозга, в которую входят базальные ганглии и передние структуры мозжечка, тогда как более дискретные движения, такие, как печатание на пишущей машинке, как игра на фортепиано, регулируются быстродействующей неомозжечковой системой осуществляющей экстраполяционные расчеты. Точный механизм этого центрального контроля за движениями пока еще не изучен, однако то, что мы знаем, достаточно, чтобы утверждать, что в этом процессе участвуют явления моделирования периферических сервомеханизмов и что такое моделирование осуществляется посредством механизмов коррекции. Наконец, предлагаемая концепция радикально изменяет наши представления о функциях прецентральной моторной коры. Эта часть мозга, как было показано, является сенсорной по отношению к действиям. Образ конечного результата формируется и непрерывно -мгновенно видоизменяется благодаря нейронным голографическим процессам, так же как и перцептивный Образ. Однако Образ ожидаемого результата создается на основе уже известных из прошлого опыта антиципации тех внешних сил и их изменений, которые требуются для выполнения задания. Эти поля сил,, оказывающие влияние на мышечные рецепторы, становятся параметрами сервомеханизма и непосредственно (через таламус) или опосредствованно (через базальные ганглии и мозжечок) связываются с моторной корой, где они сопоставляются с быстродействующими расчетами мозжечка по предвосхищению результата и последующего звена действия. Когда ход действия становится предсказуем на основании тенденций предшествующих успешных предсказаний, может быть сформирован Образ конечного результата для управления окончательной фазой двигательной активности.
Эта модель, которая вытекает из результатов приведенных исследований, существенно отличается от представлений о моторной коре как о клавиатуре, к которой стекается активность остальной части мозга, чтобы разыгрывать мелодию движения. Эта модель, точнее, представление о том, как совершить движение, кодируется в моторной коре и превращается в эффективные действия, акты – внешние выражения мозговых процессов.
В следующих главах мы рассмотрим вопрос о том, как эти внешние представительства движений модифицируются в последовательности действий. Последующая часть книги посвящен» исследованию действий, связанных с процессом коммуникации.
РЕЗЮМЕ
Из ряда мозговых процессов, участвующих в организации поведения, один привлекает особое внимание. Этот процесс включает в себя действие моторного механизма коры больших полушарий и коры мозжечка, формирующего Образ конечного результата с помощью быстродействующих расчетов. Посредством этого-механизма осуществляется экстраполяция последствий непосредственно предшествующих движению изменений во внешнем поле, которые воздействуют на мышечные рецепторы. В этом Образе-конечного результата кодируются, следовательно, внешние явления (например, силы), а не наборы мышечных сокращений. Образ-конечного результата регулирует поведение так же, как термостат регулирует температуру: на диске прибора закодированы пе включение и выключение котла, а лишь пределы допустимо»' температуры.
Глава XIV

