РЕЗЮМЕ
Выполнение действия состоит из двух фаз: 1) продолжение процесса научения, изменяющего компетентность организма, и 2) продолжение действия для достижения результата. Теория, объясняющая выполнение действия через результат, основана на экспериментах с угашением актов поведения, в частности на опытах с привыканием к наркотикам и опытах, показавших, что частота ответа предопределяет взаимоотношение звеньев в системе «средства – цель» в ситуации подкрепления.
ЧАСТЬ ЧЕТВЕРТАЯ. СТРУКТУРА КОММУНИКАТИВНОГО ПРОЦЕССА
«Границы моего языка определяют границы моего мира».
Витгенштейн
Глава XVII
ЗНАКИ
Наиболее впечатляющие достижения человека связаны с теми видами поведения, коюрые обеспечивают его коммуникативную деятельность. В четвертой части этой книги рассматривается структура коммуникации у приматов исходя из двухпроцессного механизма функционирования мозга, неврологической организации восприятия и чувств, а также регулирующего воздействия нервной системы на поведение. В гл. XVI указывалось, что коммуникация предполагает иерархическую взаимосвязь сенсорных процессов органов чувств и ствола мозга с моторными механизмами. В последующих главах эта взаимосвязь будет рассматриваться детально. Можно выделить два вида коммуникативных актов исходя из того, зависит ли смысл действия от ситуации, в которой оно происходит. Коммуникативные акты, не зависящие от ситуации, мы будем называть «знаками», и их неврологическая организация рассматривается в данной главе. Коммуникативные акты, зависящие от ситуации, мы будем обозначать термином «символы», механизмы мозга, ответственные за их образование, детально описываются в гл. XVIII. Части мозга, участвующие в образовании «знаков», в значительной степени отличаются от тех, которые участвуют в образовании «символов». Однако у человека развивается взаимосвязь более высокого порядка. Лингвистические знаки употребляются химво-лически в связной речи, а лингвистические символы используются в процессах мышления. Таким образом, в главе XIX ставится проблема, каким образом обе части мозга (части, связанные с формированием знаков, и части, связанные с формированием символов) объединяются в процессе речи и мышления.
Во всех этих главах мы будем исходить из того, в какой мере эти процессы зависят от ситуапии. По существу, предполагается, что знаки не зависят от контекста, они являются результатом воздействия ассоциативной коры (рис. XVII-1, таблица – XVII-1) на входные системы. Роза остается розой независимо от того, находится ли она в саду, на обеденном столе или в мусорном ведре.
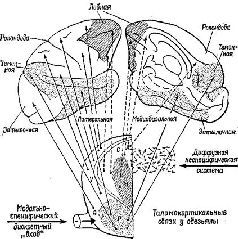
Рис. XVII-1. Схематическое изображение проекционных путей от дорсальной части таламуса к коре больших полушарий у обезьяны. Нижняя часть рисунка – схема таламуса; вертикальная линия соответствует средней линии таламуса. Верхняя часть рисунка изображает латеральную и мо-диобазальную поверхности больших полушарий мозга. Заштрихованная' часть на схеме таламуса показывает границу между внутренней частью, которая получает неспецифическую диффузную афферентацию, и наружной, частью, к которой приходят модально-специфические раздельные проекционные пути. Обозначенные точками и линиями части таламуса представляют собой ассоциативные системы: медиальные ядра внутренней части таламуса и их проекции к передней лобной коре; группа задних ядер-в наружной части таламуса и их проекции к теменно-височно-затылочной коре. Подробности см. в табл. XVII-1 (Pribram, 1958)
Значение символа, напротив, определяется контекстом. Таким образом, широко распространенный символ мира в других контекстах может быть символом победы, приветствия и т. д. Существует определенное сходство между символами и другими зависящими от контекста типами поведения, например теми, которые определяют взаимоотношения людей, так как и в символическом поведении, и в регулировании влечений, эмоций и интересов, и в распознавании правильного и ошибочного участвуют одни и те же части мозга (фронтолимбическая область переднего мозга, включающая лобную кору (рис. XVII-1, табл. XVII-1) и уже известные нам амигдалу и гиппокамп).
Таблица XVII-1


Классификация составлена на основе следующих работ: Электрофизиология: Magoun, 1950; Starzl et al., 1951; Работы, выполненные методом серебрения: Morin et al., 1951; Сравнительная гистоморфология: Каррегв et al., 1936; Hose and "Woolsey, 1949; Ретроградная дегенерация таламуса после удаления норы (на обезьянах): Walker, 1938; Chow, 1950; Chow and Pribram, 1956; Pribram et al., 1953; Цитоархитектоника и стрихниниая нейроно-графия: Von Bonin and Bailey, 1947; Bailey et al., 1950; Mac Lean and Pribram, 1953; Pribram and Mac Lean, 1953.
Пользуясь в этих главах терминами знак и символ, мы исходим из обычного их употребления в философии (например, Ауег, 1946; Morris, 1946; Langer, 1951) и в липгвистике (например, Jakobson, 1956). Поскольку употребление этих терминов связано с анализом человеческой речи, лингвистические символы в большинстве случаев рассматриваются как более высокий класс по сравнению с лингвистическими знаками, и наоборот. С неврологической точки зрения я рассматриваю знаки и символы как классы, не зависящие друг от друга и объединяющиеся только в связной речи и мышлении.
ЧТО ТАКОЕ «ЗНАК»
Когда я начал проводить опыты на обезьянах, я обычно применял в качестве подкрепления правильного выбора, ведущего к решению проблемы, очищенный арахис. При подготовке к серии опытов приходилось тратить много времени на очистку орехов. Внезапно я понял, что нет оснований считать, что обезьяны не смогут сами чистить орехи, и действительно оказалось, что они справляются с этим, не задерживая процедуры эксперимента. Меня интересовала выработка дифференцировок, и я понял, что обучение правильно реягировать на скорлупу ореха было в некотором смысле выработкой более примитивной формы дифферен-цировки, что скорлупа являлась постоянным индикатором, знаком лакомства, имевшегося внутри, совершенно таким же, как знак «плюс», нарисованный на крышке коробки, в которой находился земляной орех. Конечно, нарисованный знак был более отвлеченным, абстрактным (и произвольно выбранным), он не столь органически связан со съедобным орехом. Графический знак лишь условно вызывает соответствующие формы поведения, но тем не менее сходство такого знака со скорлупой арахиса было налицо.
Эти ранние эксперименты убедили меня в том, что выработка дифференцировки представляет собой изучение способности организма раскрывать смысл знаков. Этой формулировке, однако, не хватало той строгости, которую принято связывать с результатами эксперимента. Ведь «знак», который четко указывает на что-нибудь независимо от контекста, представляет собой нечто такое, что можно либо связывать с данной ситуацией, либо не связывать с нею. Но могут ли животные действительно пользо-ватьея знаками? Не является ли человеческий язык необходимой предпосылкой к пользованию знаками? Проследив истоки моей неуверенности, я понял, что она является следствием того, что во всех экспериментах я придумывал знак и требовал, чтобы обезьяна просто отвечала на него, нажимая на клавиши или рычаг, смещая крышку или давая какой-нибудь другой ответ. Знаки входят в процесс коммуникации, и, предполагая это, я еще не мог быть уверен, играют ли эти знаки для обезьяны такую же коммуникативную роль или нет. Так и возникла неясность.
Эта неясность была устранена сразу благодаря удивительным результатам, полученным Робертом и Беатрис Гарднер (1969), работавшими с молодой самкой шимпанзе по кличке Уошо. Поскольку попытки научить шимпанзе речи не удались, они решили попытаться научить ее общаться при помощи рук, а не голосовых связок. Они остановили выбор на американской системе знаков, применяемой глухонемыми, считая эту систему идеальным средством, поскольку она легко поддается изучению благодаря своей наглядности, ведь в этой системе при помощи кистей рук и пальцев создается некоторое подобие предмета или действия, обозначаемого этим знаком. Уошо, которой в то время, когда писалась эта книга, было около трех с половиной лет, заучила примерно 150 таких знаков (табл. XVII-2). Гарднеры и Уошо легко общались друг с другом, используя этот метод. Фактически Уошо даже изобреланесколько новых знаков, которые обычно употребляются людьми, обучающимися этой системе.
Мы еще несколько раз вернемся в следующих трех главах к этому имеющему большое значение эксперименту с Уошо. Здесь же нам нужно обсудить только один основной факт, то, что приматы могут создавать знаки и сообщать при помощи знаков, независимых от контекста, об устойчивых признаках ситуации, которые они могут таким образом дифференцировать и узнавать. А о механизмах мозга, участвующих в выработке диф-ференцировок и узнавания, нам известно очень много.
Таблица XVII-2






1 Легко видеть, что «знаки» обезьян, о которых здесь идет речь, являются лишь сокращенными действиями и не могут рассматриваться как подлинные знаки, используемые в языке человека и имеющие общественно-историческое происхождение. – Прим. ред.
ВЫРАБОТКА ДИФФЕРЕНЦИРОВОК И УЗНАВАНИЕ
Некоторые аспекты распознавания образов обсуждались в гл. VI, VII и VIII. Участие механизмов памяти описывалось как образование нейронного экрана, включающего ряд фильтров и голографических микроструктур, посредством которых и на которые проецируется вводимая информация для создания Образа. В данной главе мы займемся детальнее некоторыми из факторов, влияющих на этот зкран во время обучения и опознавания.
В гл. VI, VII и VIII процесс формирования Образа рассматривался как сравнительно пассивный механизм, который приводится в действие поступающей информацией. Однако ранее мы останавливались и на некоторых активных аспектах этого процесса. Например, в гл. V была показана необходимость опережающей связи и обратного влияния движения на восприятие, без которого невозможно сохранение стабильности и константности образа. Материал, приведенный в гл. XVI, свидетельствует о том, что весь механизм восприятия участвует в образовании значения и что этот процесс носит весьма активный характер.
Образ в нашем понимании является -целостным и топографическим. Однако, когда мы в действительности смотрим на окружающий мир или слушаем его, мы избираем тот или иной аспект, мы каким-то образом концентрируем внимание, выбираем, ограничиваем наше поле зрения, особенно если мы активно запяты рассматриванием, слушанием и т. д.
Следовательно, заучивание и опознавание знака требуют чего-то большего, чем просто формирование Образа. Становится необходимым внимательный выбор, избирательность, почти так же, как при осуществлении, двигательного навыка. Факты говорят о том, что контроль за формированием образа осуществляется посредством тех проводящих путей и структур мозга, которые принято считать двигательными по их функции. Читатель не должен считать это чем-то парадоксальным, поскольку ему уже известно, что двигательная функция, то есть поведение, в значительной мере осуществляется посредством контроля за работой периферических рецепторов.
В этой главе мы утверждаем, что знаки создаются тогда, когда наши действия влияют на воспринимаемые Образы. Знак – это действие, представляющее воспринимаемый Образ. Выработка дифференцировок, распознавание образов, избирательное внимание – все это требует участия нервных механизмов выбора, а этот выбор и запускает в ход действия, которые в свою очередь изменяют созданный Образ.
Это положение является итогом серии экспериментов на обезьянах, которые опрокинули представления о том, каким образом функционируют так называемые ассоциативные зоны мозга приматов. Т. Мейнерт (1867-1868) и П. Флексиг (1896) в конце XIX в. приписывали ассоциативную функцию всем тем частям коры головного мозга, которые не связаны более или менее непосредственно с периферическими рецепторами и эффекторами. Эти авторы исходили как из фактического материала, указывавшего, что все части коры чрезвычайно тесно связаны друг с другом, так и из теории, основанной на господствовавшей тогда эмпирической доктрине ассоциации идей. Ассоцианистскнй подход к работе мозга все еще сохраняет свое значение при изучении языка и мышления человека. Что же касается выработки дифференцировок, распознавания образов и т. п., то прежний эмпирический подход к работе мозга, согласно которому ассоциативная кора всего лишь «ассоциирует», интегрирует информацию, получаемую от различных первичных сенсорных зон, оказался несостоятельным.
В частности, множество экспериментов, проведенных на обезьянах, показало, что выработка дифференцировок и распознавание образов возможны и после удаления больших участков коры, окружающей первичные проекционные зоны (Pribram, Spinelli and Reitz, 1969, рис. XVII-2). Однако удаление более ограниченных участков, расположенных на некотором расстоянии от первичных зон, вызывает серьезные нарушения как при заучивании «знаков», так и при опознавании (Mishkin and Pribram, 1954, рис. XVII-3). Простое рассечение внутри'корковых проводящих путей, соединяющих эти зоны с первичными, нарушений не вызывает. Напротив, рассечение проводящих путей, соединяющих кору с подкорковыми структурами, вызывает столь же серьезное нарушение, как и удаление самой корковой ткани (Pribram, Blehert and Spinelli, 1966, рис. XVII-4).
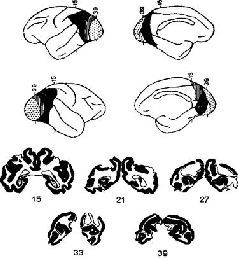
Рис. XVII-2. Схема двусторонних престриарных операций, после которых обезьяна все еще может осуществлять зрительное различение (Pribram, Spinelli and Reitz, 1969).
Р ис. XVII-З. Схема расположения первичных проекционных и ассоциативных полей в корковой зрительной системе обезьяны-резус. Обратите внимание на область, находящуюся между ними, которая была удалена в эксперименте, изображенном на рис. XVII-2.
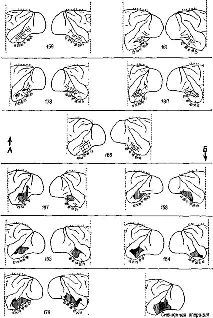
Рис. XVII- 4. A – схема операций по рассечению мозга (тонкие линии); Б – схема операций по подрезке коры снизу (черным цветом обозначена поверхностная подрезка, штриховкой – глубокая подрезка) (Pribram, 1966),
Для исследователей, стоящих на позициях ассоцианизма, эти: результаты кажутся еще одним из тех парадоксов, которые так стимулировали исследования в области нервных механизмов поведения на протяжении последних полутора столетий. Каким образом какой-то участок коры мозга может осуществлять ассоциативную роль, влияя на информацию, вводимую в другие, первичные зоны мозга, если разъединение этих зон не оказывает никакого действия?
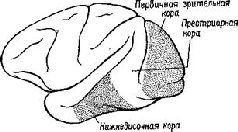
Эта загадка была частично разрешена, когда было установлено, что результаты разрушения мозга неоднозначны. Локализация функций различных сенсорных модальностей была обнаружена и внутри так называемой ассоциативной коры. Один участок в теменной доле мозга оказывает исключительное влияние на различение кожно-кинестетических сигналов; другой участок в передней части височной доли имеет отношение только к вкусовым ощущениям. Средневисочные участки коры специализированы исключительно на слухе, а участок в нижней части височной доли имеет отношение только к зрению (рис. XVII-5). Более того, повреждения в зтой так называемой ассоциативной коре не вызывают никаких нарушений взаимодействия между раздражителями разных модальностей (Pribram, 1969a).
Таким образом, проблема сводится к определению функций этих специфических сенсорных зон. Экспериментальные исследования для разрешения зтой проблемы проводились в основном на зрительных процессах и поэтому касались коры нижней части височной области. Однако и в отношении слуховых и соматосен-сорных процессов было сделано достаточно для того, чтобы прийти к выводу, что результаты, полученные при изучении зрения, применимы и к другим частям ассоциативной коры, обслуживающим иные сенсорные модальности.
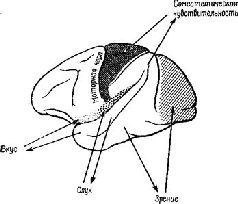
Рис. XVII-5. Схема латеральной поверхности мозга обезьяны, на которой изображены модально-специфические первичные сенсорные зоны и специфические сенсорные подразделения ассоциативной коры.
Прежде всего возник вопрос, вызывают ли повреждения нижневисочной коры мозга обезьян нарушение всех форм зрительных навыков. Скоро удалось установить, что такое повреждение не нарушало слежения за зрительным объектом, например за летящим комаром. Позднее это было подтверждено посредством фотографирования отражения наблюдаемых объектов от роговицы (Bagshaw, Mackworth and Pribram, 1970).
Нарушения возникали лишь в том случае, когда перед обезьянами с повреждениями мозга стояла задача выбора или дифференцировки. Такие нарушения были обнаружены при самых различных формах зрительного выбора независимо от того относился ли он к цвету, форме, контуру или яркости (Mishkin and Pribram, 1954). Единственным общим в этих задачах было то, что все они были зрительными и во всех предусматривался выбор, а также то, что массивность нарушения поведения была пропорциональна трудности, которую испытывали при обучении нормальные обезьяны.
Эти результаты сразу же заставили исследователей предположить, что затруднения, наблюдаемые у обезьян с повреждениями мозга, выражаются скорее в их неспособности заучить (и запомнить) задачу, чем в неспособности воспринять основные ключевые признаки, которые ведут к ее разрешению. Для проверки этой гипотезы было проведено исследование того, как обезьяны с удалением нижневисочной коры и с частичным удалением первичной зрительной зоны решают различные зрительные задачи. В общем результаты подтвердили гипотезу, что научение и восприятие могут быть разграничены путем повреждений разных частей коры. Так, повреждение первичной зрительной коры сказывалось на решении задач на восприятие: обезьяна не могла подтянуть орех, привязанный к шнурку, находящемуся среди нескольких других, к которым не была привязана приманка; разрушение нижневисочных отделов коры не вызывало таких нарушений. Противоположная картина наблюдалась при выполнении заданий на научение (Wilson, Mishkin, 1959, рис. XVII-6).
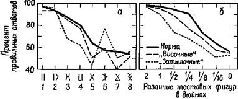
Рис. XVII- 6. A – средние результаты выполнения структурированного серийного теста у трех групп обезьян (контрольная группа; обезьяны с поражением первичной коры; обезьяны с поражением нижновисочной зрительной коры); В – средние результаты выполнения задания на различение размера у трех групп обезьян (Wilson, Mishkin, 1959).
Следует, однако, обратить внимание на то, что здесь сказано «в общем». Расхождение между влияниями, оказываемыми этими двумя типами повреждений на восприятие и научение, не было ни полным, ни вполне определенным. Результатом повреждения первичной коры было некоторое замедление процесса научения, что можно было ожидать, так как обезьяны приспосабливались к сниженной остроте зрения и большим скотомам в зрительном поле. Более загадочным было то, что у обезьян с повреждениями нижневисочной коры наблюдалось некоторое снижение остроты зрения, например в отношении размера (Mishkin, Hall, 1955) н критической частоты мельканий (Mishkin and Weiskrantz, 1959). Поскольку острота зрения проверялась методом выработки дифференцировок, эта загадка легко объяснялась тем, что был нарушен механизм выбора, а в результате этого нарушалось как научение, так и запоминание. Однако проверка на людях, у которых по той или иной причине была удалена височная доля, поставила под сомнение такое объяснение (Goldman et al., 1968), Изменения критической частоты мельканий наблюдались, даже когда использовался словесный отчет о слиянии (метод определения порогов при повышении и снижении частоты мельканий). Читателя не должны слишком удивлять эти результаты, как не должны были они удивлять нас, исследователей. Распознавание образов, дифференцировка знаков требуют участия как процесса формирования Образа, так и памяти. Знакомое лицо стало знакомым благодаря опыту, но оно воспринимается целостно и непосредственно.
КАК МОЗГ КОНТРОЛИРУЕТ ПОСТУПАЮЩУЮ ИНФОРМАЦИЮ
Мы были озадачены тем, что вначале не могли представить себе механизм, который частично – но не полностью – разделял бы восприятие и научение. В классическом понимании функций коры вводимая информация воспринимается посредством первичной коры; к тому, что уже заучено, добавляется затем сумма других видов информации и ассоциаций, хранимых в ассоциативной коре. У. Пенфилд (1969) как-то назвал кору височной доли «корой памяти», так как электрическая стимуляция этой части мозга у эпилептиков вызывает воспроизведение картин пережитого. Однако эта классическая точка зрения не согласуется с парадоксальными фактами, обнаруженными в ходе экспериментов с рассечением коры.
Как указывалось во введении к этой главе, можно полностью разрешить этот парадокс, если серьезно рассмотреть гипотезу о том, что влияние нижневисочной коры на зрительные процессы обусловлено не столько тем, что она получает зрительную информацию отпервичной коры, сколько тем, что она воздействует через кортикофугальные связи напроцессы переработки зрительной информации, происходящие в подкорковых структурах. Какие же факты делают вероятным существование такого коркового контроля подкорковых механизмов зрения? И как можно представить себе его функционирование на основании уже известных нейропсихологических данных?
Вспомним, что подрезка нижневисочной коры, прерывающая связи между этой зоной и подкорковыми уровнями, вызывает такое же нарушение функции, как и удаление самой этой зоны. Куда же ведут эти связи?
Путь нервных волокон, идущих от нижневисочной коры, был прослежен при помощи анатомических (Whitlock and Nauta, 1954) и физиологических (Reitz and Pribram, 1969, рис. XVII-7) методов. Некоторые из этих волокон достигают таламуса, откуда афферентные связи идут к височной коре. Но значительно большее число нервных волокон доходит до подкорковых структур, обычно считающихся двигательными по своей функции, – ба-зальных ганглиев, таких, как амигдала и скорлупа, – и таких глазодвигательных структур, как верхнее двухолмие ствола мозга (рис. XVII-8).
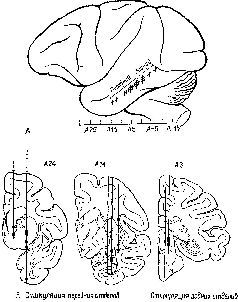
Рис. XVII-7. А – боковая проекция мозга с указанием участков стимуляции, обнаруживающих подкорковые связи нижневисочной коры; В – заштрихованные участки обозначают области, где возникает ответ при стимуляции нижневисочной коры. Обратите особое внимание на ответы в скорлупе и верхнем двухолмии (Rcitz, Pribram, 1969).
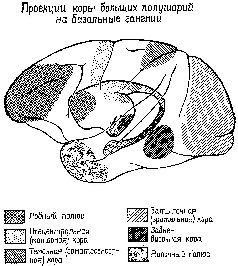
Рис. XVII-8. Мозг внутри мозга: схема проекций коры больших полушарий на хвостатое ядро и скорлупу. На этой схеме не показано перекрытие зон (на основании данных Kemp, Powell, 1970).
Влияние электрической стимуляции нижневисочной коры может быть зарегистрировано на различных уровнях проводящих путей. Изменения электрической активности, вызванные вспышками света, можно получить и путем раздражения височной коры. Эти изменения наблюдались в первичной коре, в латеральном коленчатом теле, таламусе и даже в зрительном нерве. Связи, идущие от базальных ганглиев и двухолмия к зонам таламуса и сетчатки, не были полностью исследованы, но было сделано достаточно, чтобы установить существование таких связей, хотя в отношении контроля мозга над функциями сетчатки (и, следовательно, зрительного нерва) пока имеются только электрофизиологические данные (Spinelli and Pribram, 1966, 1967, рис. XVIII-9) и нет еще гистологических.
Более полно были прослежены проводящие пути от височной коры к периферии в слуховой системе. И снова двухолмие (в данном случае слуховое) представляет собой важное место переключений; от него проводящий путь нисходит к кохлеарному ядру (либо непосредственно, либо через верхнюю оливу, либо обоими путями; Nobel, Dewson, 1966).
Таким образом, снимается вопрос о том, существуют ли связи, посредством которых так называемая ассоциативная кора может оказывать влияние на поступающую информацию. То, что этот контроль тесно связан с двигательными механизмами, согласуется с данными нейропсихологии, говорящими о том, что височная кора играет некоторую роль в тех случаях, когда следует произвести активный выбор. Но каким образом этот выбор влияет на классификацию поступающей информации, на создание знаков и придание им значения?
ВНИМАНИЕ
Одной из самых поразительных особенностей Образов является их богатство. Кк правило, все это богатство нельзя обнаружить сразу, его нужно выявлять постепенно, признак за признаком. Выделяется то цвет, то форма, то структура воспринимаемых объектов. Однако это ограничение процесса внимания не ослабляет непосредственность и калейдоскопическую природу Образа – пожалуй, и то и другое возрастает при надлежащем внимании. Каким же образом?
Ответ нужно, конечно, искать в непрерывном взаимодействии между процессом формирования Образа и вниманием. Для осуществления этого имеется соответствующий нервный механизм. Известно, что процессы, возникающие в так называемых ассоциативных зонах, воздействуют на функции воспринимающих систем. Экспериментальные данные говорят о том, что посредством этих путей осуществляется контроль за направленностью внимания со стороны подкорковых, а возможно, и корковых фильтров, или экранов, в пределах входных систем.
Эти данпые получены в экспериментах с применением методики оценки скорости восстановления возбудимости во входных каналах (рис. XI-6 и XI-7). Когда какое-то довольно краткое изменение в окружающей среде, например вспышка света или звук щелчка, вызывает ответ в сенсорной системе, этот ответ занимает ограниченный отрезок времени. Если прежде чем система «восстановилась» после своей реакции на первоначальное изменение, появится вспышка или щелчок, ответ на второй сигнал будет испытывать на себе влияние первого. Продолжительность времени реакции в ответ на возбуждение имеет поэтому значение при определении способа обработки доходящего до мозга сигнала. В наших экспериментах нам удалось показать, что электрическая стимуляция (или даже удаление) так называемой ассоциативной коры может изменить продолжительность этого времени.
Участками, где, вероятнее всего, обнаруживается такое влияние, являются, конечно, те, где осуществляются тормозные взаимодействия нейронов, иначе говоря – «на входе». Эти взаимодействия образуют фильтры, через которые и на которые передается вводимая информация. Например, путем увеличения латерального торможения может быть усилен сенсорный контраст и замедлено восстановление системы (поскольку считается, что латеральное торможение реципрокно влияет на затухание; см. рис. III-10).
Замедление восстановления возбудимости в первичной зрительной системе действительно наблюдается при стимуляции нижневисочной коры (Spinelli and Pribram, 1966, рис- XI-7). Мы испытали большое удовлетворение, получив этот результат, потому что он указывал, посредством какого нервного механизма так называемая ассоциативная кора осуществляет контроль над первичными рецепторными системами. Мы хотели провести дальнейшее исследование этого механизма, но, как часто случается в лабораторной работе, обнаружили, что обладаем недостаточными знаниями переменных, входящих в состав изучаемого явления. Когда мы пытались повторить опыт, нам не удавалось получить надежный результат. Так, как эта проблема имела большое значение, мы упорно продолжали работу и нашли путь определения условий, необходимых для получения искомого эффекта.
Поскольку нам нужен был более стабильный индикатор возбудимости, мы временно отказались от многократных предъявлений вспышки света. Мы полагали, что сетчатка является нестабильной зоной и что электрическая стимуляция более центрального участка зрительной системы посредством вживленного электрода даст более надежные результаты. Возможно, однако, "что за достижение стабильности нам пришлось бы заплатить деной утраты возможности оказывать влияние на возбудимость системы. И действительно, электрическая стимуляция нижневисочной коры не смогла оказать влияние на возбудимость, определяемую корковым электродом.
Эту дилемму разрешил наш сотрудник Л. Гербрандт посредством простого наблюдения. Он показал, что амплитуда ответов на электрическую стимуляцию зрительной системы зависит от уровня внимания обезьяны во время опыта. Когда обезьяну сажали в закрытый ящик, реакция была слабой. Когда же ящик открывали и обезьяна получала возможность оглядываться по сторонам, реакция усиливалась. Более того, раздражение нижне-височной коры могло превращать слабый ответ, полученный в закрытом ящике, в сильный, но оно не оказывало влияния на «ильный ответ. И наконец, используя величину зтого вызванного при помощи электрической стимуляции ответа в качестве контрольного, Гербрандт мог предсказать, повлияет ли раздражение лижневисочной коры в условиях закрытого ящика на функцию восстановления возбудимости зрительной системы или нет.
В случаях, когда обезьяна была внимательна, ранее полученные нами .результаты не наблюдались. Когда же обезьяна начинала «скучать», клевать носом и т. д., влияние электрического раздражения на функцию восстановления было очевидным. В наших первоначальных экспериментах мы по заведенному порядку проделывали длительные ежедневные процедуры: парные вспышки, парные щелчки, комбинации щелчок – вспышка и вспышка – щелчок, вспышки по определенному шаблону и т. д., причем все это предъявлялось в определенном порядке, изо дня в день, из недели в неделю. Не только обезьяны, но и мы со Спинелли, проводившие эти опыты, разочаровались в этом рутинном порядке опытов. Спинелли и я поочередно следили за двумя своими обезьянами, чтобы они не заснули. Мы наблюдали за обезьяной через смотровое отверстие и, если она начинала дремать, тихонько постукивали по стенке. Если начинал дремать исследователь, мы с той же целью слегка сдвигали табурет, на котором он сидел Короче говоря, наши результаты объяснялись тем, что обезьяны не были внимательны. Впоследствии группы исследователей, изучавшие только цикл восстановления возбудимости у обезьян, и работавшие с обезьянами в новой для них ситуации, Kirja их явно интересовала процедура опыта, получали иные результаты. Только тогда, когда из-за повторения ситуация наскучивала и нам и обезьяне, вновь появлялись интересовавшие нас эффекты (Gerbrandt, Spinelli and Pribram, 1970, рис. XVII-9).
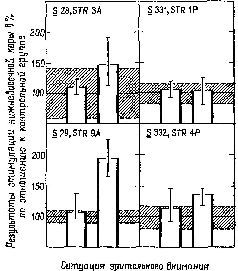
Рис. XVII-9. Влияние стимуляции нижневисочной коры на величину ответов, возникающих в зрительной коре при раздражении латерального коленчатого тела. Отметьте, что только в ситуации закрытого ящика (правая часть рис.) у трех обезьян из четырех имеется эффект, который превосходит дисперсию (затемненные участки), наблюдающуюся при отсутствии стимуляции (Gerbrandt, Spinelli and Pribram, 1970).
Эти эксперименты, вместе взятые, показывают, что результаты электрической стимуляции нижневисочной коры и результаты, получаемые, когда обезьяна «внимательна», сходны и что эти два процесса характеризуются высоким уровнем конвергенции на каком-то конечном механизме. Поэтому логично предположить, что процесс внимания предполагает влияние, оказываемое нижневисочной ассоциативной корой на механизм входа. Посредством этого влияния внимание может изменять временные параметры тормозных взаимодействий в фильтрах, а следовательно, и характеристику Образа, вызванного любым данным сигналом.
Таким образом, влияние височной доли на систему входа представляет собой частный случай и результат какого-то более общего процесса, посредством которого происходит усиление-определенных операций. Каким же образом тогда этот механизм: влияет на дифференцировку, на процесс узнавания?
ФИЛЬТРЫ ПРИЗНАКОВ И МЕХАНИЗМЫ ОПОЗНАНИЯ
Многочисленные исследования показали, что узнавание предполагает идентификацию признаков, различающих объекты. На этом основывается имитация этого процесса на ЭВМ при составлении программ. Открытие нейронов, чувствительных к определенным признакам, то есть способных избирательно реагировать на линии, наклонные под разным углом, движение, цвет и т. д., стало поэтому считаться решением проблемы узнавания. Не умаляя значения этого открытия, мы показали в предыдущих главах, что нельзя объяснить все проблемы восприятия наличием механизма детекции признаков. Позвольте мне развить эту точку зрения, высказав предположение, что проблема узнавания, завися-щая от памяти, также останется неразрешенной, если мы будем основываться только на детекции признаков. Поэтому я разграничу детекцию признаков и их опознание. Детекторы признаков являются необходимым звеном нервного аппарата и не могут быть радикально модифицированы в результате накопления опыта, если они должны выполнять задачу выделения признаков.
Поэтому детекторы признаков – это стабильные, «встроенные», врожденные элементы входных систем, которые проводят предварительную обработку сигналов, до того как они подвергнутся дальнейшей обработке. Напротив, механизм идентификации признаков, на котором основывается узнавание, должен по необходимости быть гибким и поддающимся модификациям в процессе приобретения опыта. Однако в силу непосредственности узнавания при предварительной обработке доходящей до мозга информации происходит одновременно как выделение признаков, так и процесс их опознания. Это участие двух разных процессов и привело к смешению этих двух механизмов и к предположению, что детекторы признаков целиком выполняют эту работу.
Детекторы признаков воспринимаемых объектов играют довольно-таки ограниченную роль. Полагают, что они наряду с другими механизмами обеспечивают необходимый уровень, фон, на котором возникают другие, более подвижные конфигурации нервных событий. Они представляют собой постоянные элементы, «основу», на которой возникает узор микроструктурных изменений, чувствительных к влиянию приобретаемого опыта. В любой данный момент эта «основа» подвергает обработке воздействующие на нее нервные сигналы, то есть подготавливает их к переключению на последующие клеточные элементы. Процесс обработки не оказывает влияния на саму «основу», но добавляет к ней еще один новый узор – еще одну нить.
Ряд таких механизмов, посредством которых опыт может оказывать влияние на нервную систему, был детально описан в гл. II и XIV. Эти механизмы, по-видимому, могут настраивать, изменять действие тех элементов, которые составляют экран. Небольшие изменения времени восстановления возбудимости нервных элементов, вызываемые, например, стимуляцией нижневисочной коры, могли бы, если бы они стали постоянными, изменять характеристики ответов элементов системы, пока они не будут «настроены» особым образом для восприятия только данной конфигурации пейронной микроструктуры. Спинелли ( 1970, см . рис. VII-8) имитировал в программе ЭВМ подобную систему, названную им «Оккам». Эта программа может «узнавать» форму волны, если она применялась несколько раз, и может опознавать данную структуру даже по ее части. Кроме того, эта программа может различать десятки форм волны.
Можно, пожалуй, считать, что нервный механизм, посредством которого осуществляется опознание, действует сходным образом. Обычно организованные элементы фильтра, колонки кортикальных клеток более или менее связаны между собой посредством элементов, чувствительных к направлению. Напомним, что в гл. VII высказывалось предположение, что чувствительность к направлению рецептивных полей корковых клеток является определяющим фактором для соседних клеток, превращая таким образом кортикальную колонку в структуру,, включающую определенные координаты, или реестр. Латеральное-торможение строит этот реестр путем разделения организованных в определенную систему элементов. Каждый модуль этой системы, каждый реестр можно представить себе как диполь,, поляризующийся при помощи входных сигналов. Многое говорит в пользу существования электрических диполей в коре (см. Elul, 1964; Barrett, 1969). Электрический диполь мог бы образоваться из тех изменений в молекулярной структуре, которые обсуждались в гл. II и VIII. Эти структурные изменения можно-измерить при помощи ядерной магнитно-резонансной спектроскопии и изучения дифракции рентгеновских лучей, поскольку каждая структура резонирует на своей частоте. Известно, что макромолекулы типа липопротеинов и глюкоидов, образующие-синаггхические и дендритные мембраны в микроструктурах мозга,, подвержены таким изменениям. Всякий раз, когда нервный сигнал проходит через такую мембрану, эти изменения имеют тенденцию-стабилизироваться, упорядочивая молекулярные структуры, пока длится сигнал и в течение некоторого ограниченного времени после него. Если эти временно стабилизирующиеся изменения достаточно" обширны, они будут вызывать электрическую поляризацию микроструктур. Эта поляризация возрастет, когда вследствие латерального торможения влияние каждого диполя на соседний становится минимальным. Без таких тормозных взаимодействий действие диполей имело бы тенденцию к ослаблению, подобно тому, как полюса магнитов, поставленные параллельно, имеют тенденцию нарушать упорядочивание. Поэтому в результате влияния раздражения височной коры на латеральное торможение действие-диполей при входном сигнале возрастает (см. рис. XVII-10), причем это возрастание обеспечивает постоянство связей между входным сигналом и организованными элементами системы. Эти связи составляют мелкие корковые функциональные единицы, обеспечивая тем самым более легкую адаптацию каждого элемента к своему входному сигналу. Как указывал Росс Эшби (1960, рис. XVI1-10), полностью взаимосвязанная система ультрастабильна, что затрудняет адаптацию. Модификация может происходить только путем «разделения системы на части» при постоянстве связей.
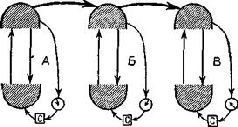
Рис. XVII-10. Схема установления устойчивого состояния в гомеостате Эшби. Верхние полукруги изображают вариации стимулов; нижние – вариации нервных ответов. С – выключатель селектора, стрелка-монитор. Обратите внимание на сходство этой схемы со схемой Т -О -Т -Е (Ashby, 1960).
Такая система диполей, структур нейронных реестров имеег определенные преимущества, потому что она представляет собой, параллельно обрабатывающий механизм, связанный с содержанием. Элементы такой системы действуют одновременно, а непоследовательно; место хранения информации теряет свое значение. Предварительная обработка и модификация, основанные на опыте, происходят бок о бок и практически одновременно. Короче говоря, некоторые части механизма предварительной обработки изменяются в процессе этой обработки. Нервный механизм самоорганизуется.
Подведем итог. В реальной жизни нелегко разграничить перцептивный процесс формирования Образа и познавательный процесс его опознания. Мы опознаем то, что ощущаем как значительное, почти в тот же момент, когда воспринимаем это посредством своих органов чувств. Но для целей научного анализа, особенно если нас интересует механизм, важно расчленить, как мы и сделали в этих главах, различные участвующие в этом процессы. Поскольку в итоге мы выясняем, что нервные механизмы этих процессов в действительности тесно взаимосвязаны, мы приходим к более полному пониманию того, каким образом возникает единство психологического процесса. Когда это единство ясно выражено, знак опознается или актуализируется, но эта актуализация (как и все действия) должна осуществляться последовательно. Любое мгновенное формирование знака поэтому не является полным – знак извлекает значение только из целостного Образа и только тогда может заменить его.
ЗНАЧИМОСТЬ
Нам осталось обсудить последний вопрос: как некоторые аспекты Образов обретают смысл, становятся значимыми? В экспериментах по выработке дифференцировок у обезьян какое-либо изображение получает смысл, становится знаком в результате поведения самой обезьяны. При выборе используется нервный механизм, необходимый для совершения действия. Тот факт, что проводящие пути от нижневисочной области коры, которая оказывает влияние на зрительное внимание, проходят через двигательные отделы мозга, обеспечивает структурную базу для взаимодеиствия двигательных и сенсорных процессов и для влияния-Образов результата на Образы восприятия. Поэтому значение данных сигналов создается посредством действия. Именно, этот активный процесс образования знака порождает значение: он является результатом перцептивного научения посредством подкрепления. Оказывается, что значимость сигналов зависит от механизма, который привлекает к ним внимание, от подкреплений и альтернатив (см. рис. XVII-11). Обезьяны, лишенные нижневисочной коры, выбирают из ограниченного числа альтернатив (при меньшей неопределенности); зрительный выбор является результатом четкого различения воздействующих агентов (Pribram, 19606, см . рис. XIX-11, XIX-12, Butter, 1968, рис. XVII-12). Если животное внимательно или если нижневисочная кора подвергается электрической стимуляции, происходит замедление восстановления сенсорных систем (рис. XI-5, XI-6). Такое замедление уменьшает избыточность поступающей информации. Таким образом, данный ряд сигналов начинает входить в более мелкие части нейронных голограмм. Когда для реконструкции Образов используются маленькие части физической голограммы, глубина поля возрастает и большая часть Образа попадает в фокус, хотя и ценой некоторых небольших потерь в деталях. Работа этого механизма весьма напоминает укорочение фокусного расстояния транслокаторного фотообъектива – при этом в фокусированное поле зрения попадает большая площадь. Были получены прямые доказательства наличия такого механизма у человека. Измерения рецептивных полей отдельных элементов в зрительной коре человека показывают, что константность восприятия величины достигается здесь при помощи процесса, поддерживающего постоянный угол относительно точки фиксации взора в геометрическом центре рецептивного поля (Marg and Adams, 1970). Так, когда я хочу рассмотреть большую часть какой-либо сцены, я укорачиваю фокусное расстояние. Парадоксально, что, когда затем меня начинает интересовать тот или иной аспект этой сцены, соответствующие признаки воспринимаемого выделяются и становятся свободными от влияния побочных деталей Когда же, напротив, я намереваюсь уточнить какой-то ограниченный сегмент данной сцены и исключить все остальное, я удлиняю фокусное расстояние. Сегмент, находящийся в фокусе, лишается, таким образом, контекста, с которым первоначально был связан, вследствие чего этот сегмент может получать разные значения в зависимости от цели или установки наблюдателя.
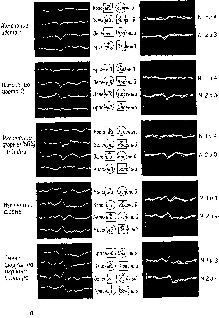
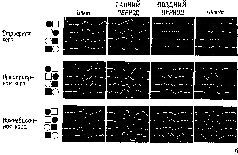
Рис. XVII-11. Результаты экспериментов по изучению функций нижневисочной коры электрофизиологическим методом. Эксперимент сходен с тем, который описан на рис. VII-3 и VII-4. Обезьяне предъявляют зрительные стимулы на экране, и она отвечает нажатием либо на правую, либо на левую часть панели, чтобы получить вознаграждение; одновременно производится запись электрических ответов мозга 'с помощью небольшого компьютера общего типа («РДР-8»). В этом эксперименте вспышки света освещали окрашенные (красные и зеленые) квадраты и круги. Обезьяны в случайном порядке, в зависимости от подкрепления обращали внимание и отвечали либо на форму (круги или квадраты), либо на цвет стимула (красный или зеленый). Как и в предыдущем эксперименте, было обнаружено, что стимул, ответ и подкрепление закодированы в первичной зрительной коре. Кроме того, этот эксперимент показал, что связь между параметром стимула (форма или цвет) и ответом обнаруживается прежде всего в нижневисочной коре. Этот факт представлен на рисунке б, третья линия, где усредненные электрофизиологические записи (250 мсек до и 250 мсек после вызванного ответа) указывают на четкое разлячие в форме волн в зависимости от того, что подкрепляется: форма или цвет. Обратите внимание, что это различие возникает, несмотря на то что образ на сетчатке, образуемый вспышками света, как при ответе на форму, так и при ответе на цвет, идентичен. Когда обезьяна «перетренируется», подкрепление вызывает направленную ассоциацию между признаком стимула и ответом, которая начинает кодироваться в первичной зрительной коре, как это видно из рисунка (Rothblat, Johnston and Pribram).
Это сужение фокуса в зависимости от контекста представляет собой другой аспект проблемы значимости. В гл. XVIII мы рассмотрим этот аспект – зависимость значения от символических процессов. Символы, как и знаки, порождают значение.
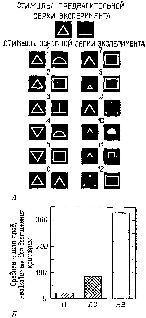
Рис. XVII-12. Результаты эксперимента, выясняющего влияние поражения нижневисочной коры обезьяны на специфику процесса обучения реагировать на треугольник (верхняя фигура) по сравнению с неоперированными животными. А – сигналы, применявшиеся в эксперименте; В – результаты; Н – норма; ЛС – обезьяна с удалением латеральной стриарной коры (первичная зрительная кора); НВ -обезьяна с удалением нижневисочной коры (Butter, 1968).
РЕЗЮМЕ
Своими достижениями человек в большой степени обязан своим уникальным коммуникативным способностям. Эти способности развиваются, когда в восприятие и переживание включаются моторные механизмы мозга. Возникающие в результате этого кодирующие процессы создают знаки и символы. Знаки создаются и опознаются тогда, когда моторные механизмы действуют на структуру нейрональных соединений, структуру, образуемую поступающей информацией, а символы – когда моторные механизмы воздействуют на сенсорные процессы в центральной части ствола мозга (Различение символов и знаков, предлагаемое автором, нуждается, безусловно, в дальнейшем уточнении. – Прим. ред.). Нервная организация знаков и символических процессов различна: знаки становятся коммуникативными действиями, остающимися неизменными в большом диапазоне контекстов; символические коммуникации почти полностью зависят от контекста, в котором встречаются символы. Знак свободен от контекста благодаря функции проводящих путей от ассоциативной коры мозга, которые влияют через моторные зоны на соединительный механизм сенсорного входа, описанный в гл. VII и VIII. Эта кортикофугальная регуляция входного сигнала соответствует процессу избирательного внимания.
Глава XVIII
СИМВОЛЫ
ПОБУЖДАЮЩИЕ ЗНАКИ
Символы – это побудители к действию. Они обретают смысл на основе их применения в прошлом и состояния организма, использующего их, в настоящем. Этим они отличаются от знаков. Символы как обозначения зависят от контекста, в котором они появляются. Знак
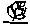
– это роза, и более ничего. Символ – свастика имеет различное значение для еврея и для индуса.
Цель данной главы – показать, что символы создаются, когда действия влияют на чувства, на интересы. Фактический материал касается функций лобной коры приматов, той части мозга, которая на протяжении примерно десятилетия, в 40-е и 50-е годы, подвергалась изоляции посредством лоботомии (лейкотомии) в целях излечения больных, страдающих психическими расстройствами. Хотя терапевтическая эффективность этой процедуры остается сомнительной, ее применение вызвало многочисленные нейропсихологические исследования, которые лишь теперь начинают становиться такой отраслью знаний, которая поддается точной интерпретации.
ИССЛЕДОВАНИЕ ОТСРОЧЕННЫХ РЕАКЦИЙ
Это началось вскоре после первой мировой войны в психологических лабораториях Чикагского университета. Функционализм в области психологии, который был тогда новостью, поставил много методических вопросов. Один из них задал Уолтер Хантер (1913), который был в то время аспирантом: «Возможно ли, что у детей и животных действительно появляются «представления»? И можно ли это доказать?» Он разработал метод, при котором на глазах у ребенка какое-то лакомство, например кусок шоколада, прятали в разных местах при последовательных пробах. При каждой пробе либо ребенка помещали в другое место, либо между ним и местом, где спрятано лакомство, ставили ширму. Несколько минут (или даже часов) спустя ребенка допускали к месту, где было спрятано лакомство, причем это место каждый раз менялось и, таким образом, в противоположность тому, что происходит при различении знаков, теряло всякую определенность. Если ребенок найдет лакомство, значит, у него сохранилось «представление» о спрятанном лакомстве и месте, где оно спрятано, и это представление сохраняется в течение интервала между тем моментом, когда оно было спрятано, и моментом, когда оно было обнаружено. Оказалось, что как дети, так и большинство животных хорошо справлялись с этой задачей, что доказывало их способность к припоминанию.
В 30-х годах К. Джекобсен в Йельском университете заинтересовался разработкой методики для определения нарушений, вызванных повреждением лобной коры мозга у приматов. Клинические исследования и лабораторные наблюдения над обезьянами наводили на мысль, что повреждение лобных долей мозга вызывает нарушение каких-то мыслительных процессов. Поэтому Джекобсен счел логичным воспользоваться для изучения последствий повреждения лобной доли вышеупомянутым тестом, который стал для ученых показателем способности к припоминанию.
К этому времени было создано несколько вариантов этого теста. В первом варианте приманку на глазах у животного прятали в один из двух одинаковых ящиков, затем закрывали крышку и на несколько секунд (обычно 5-15) ставили ширму между животным и ящиками, после чего убирали ширму и предлагали животному выбрать один из ящиков. Иногда ширма бывала прозрачной, иногда непрозрачной. Только опыты при непрозрачной ширме являлись настоящим тестом на отсроченную реакцию. В одном из вариантов теста применялся «косвенный метод»: какой-то вспомогательный признак, например цветной предмет, на некоторое время показанный над местом, где было спрятано лакомство, обозначал, в каком ящике по прошествии определенного интервала времени можно найти подкрепление. После интервала животному разрешали выбрать один из ящиков, который теперь уже не воспринимался непосредственно. Другой модификацией была методика «отсроченного чередования», в которой сигнал, указывающий место, не применялся вовсе. Подкрепление просто перекладывалось в каждой пробе из одного ящика в другой {рис. XVIII-1).
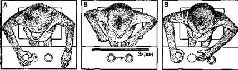
Рис. XVIII-1. Опыт с реакцией выбора: чтобы дать правильный ответ и получить в награду орех, обезьяна должна запомнить, какую чашку она поднимала в последний раз. Обезьяна получает вознаграждение, если она запоминает, что чашки надо поднимать по принципу простого чередования: левую, правую, левую и т. д. После каждой пробы между обезьяной и чашками выдвигался экран, который оставался в таком положении в течение некоторого периода времени, варьировавшего от нескольких секунд до нескольких минут. На рисунке В обезьяна забыла изменить свою реакцию. Эксперименты показывают, что поражение лобных и лимбических отделов мозга влияет на способность обезьяны выполнять такого рода задания. Однако при небольшом изменении задания (см. рис. XVIII-6) обезьяна с указанными поражениями мозга выполняет его так же успешно, как и неопе-рированные контрольные животные (Pribram, 1969).
Подход Джекобсена (1928, 1936) к изучению лобной коры оказался удачным: резекция лобного полюса нарушала нормальное выполнение всех вариантов задачи на отсроченную реакцию. Следовательно, от лобных долей каким-то образом зависит способность организма припоминать недавние события.
Здесь для нас представляет особый интерес косвенная форма задачи на отсроченную реакцию. Использование этого косвенного метода (выделяющего место) привела некоторых из сотрудников Джекобсена к разработке других задач для исследования способности пользоваться памятными знаками. Наиболее известен прибор под названием «Шимпомат». Автомат выдает фишки, такие, как при игре в покер, и вся «игра» проводится с фишками, которые только позднее и в другом автомате могут быть «обменены» на арахис. Нормальные шимпанзе без затруднений пользуются фишками, этими знаками памяти, но шимпанзе, у которых произведена резекция лобных долей, совершенно не в состоянии оживить следы, ведущие от условного знака к подкреплению (Jacob-sen, Wolfe, Jackson, 1935; рис. XVIII-2).
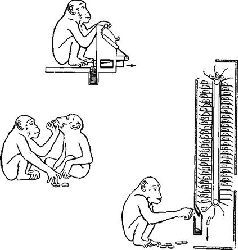
Рис. XVIII-2. Рисунок, показывающий, как шимпанзе научилась «работать» за деньги-фишки (вверху) и опускать заработанные жетоны в автомат, продающий бананы и орехи (справа внизу). Шимпанзе работают, откладывают, копят фишки (миниатюрная экономическая система) и даже крадут их друг у друга (слзва внизу).
Этот метод обучения получил недавно дальнейшее развитие в другом экспериментальном исследовании, в котором была предпринята попытка научить шимпанзе общению. В гл. XVII описывалась способность обезьяны Уошо пользоваться знаками. Д. Примак (1970) в Калифорнийском университете в Санта Барбара обучал шимпанзе Сару совсем иным методом, чем тот, которым пользовались Гарднеры. Примак применил методы выработки оперантного условного рефлекса для установления того, какой сложности систему условных знаков можно использовать для регуляции поведением Сары. «Шимпомат» уже показал, что шимпанзе будут «работать» за условные знаки. Шимпанзе Примака продемонстрировала не только возможность поведения, зависящего от таких знаков, но также и то, что животное может давать надлежащий ответ и при серийной организации условных знаков (рис. XVIII-3).

Рис. XVIII-3. Несколько примеров наборов символов, использованных Сарой для общения со своим экспериментатором (Preinack, 1970).
Во всех этих экспериментах суть проблем заключается в том, что условный знак не вызывает единообразного ответа. В зависимости от ситуации (контекста, в котором появляется такой знак) он должен быть понят, перенесен в другое место, опущен в автомат или вручен кому-нибудь, обменен на другой условный знак или на вознаграждение. Либо же, как в первоначальном опыте с отсроченными ответами, знак символизирует вознаграждение, которое должно появиться то в одном, то в другом месте.
Для описания этих зависящих от контекста побудительных знаков я буду пользоваться термином «символы». (Это различение терминов согласуется с тем, которое проводит Н. Хомский, 1963, и применяется в данном случае, чтобы указать на то, что истоки правил, определяющих человеческую речь, нужно искать в том, что здесь называется «значимыми» и «символическими» процессами.) Как указывалось в гл. XVII, теперь в нашем распоряжении имеется множество фактов, указывающих на то, что третичные отделы коры, лежащие между классическими сенсорными проекционными зонами в задних отделах мозга, участвуют в различении свободных от контекста знаков. Далее излагаются факты, показывающие, что лобная кора, лежащая кпереди от двигательных зон, участвует в символических процессах, зависящих от контекста.
Джекобсен полагал, что процессы, изучавшиеся при помощи этих процедур, должны быть связаны с кратковременной памятью и что эти процессы можно отнести только за счет функции лобных долей мозга. Более поздние исследования показали, что эта гипотеза подтвердилась при двух исключениях. Первое исключение состоит в том, что повреждения хвостатого ядра в области базальных ганглиев (части моторной системы мозга) также-вносят нарушения в выполнение отсроченных реакций. Здесь мы вновь находим доказательство участия двигательных механизмов в осуществлении высших корковых процессов. Второе исключение касается выполнения задач, связанных с отсроченным чередованием; хотя выполнение их и не предполагает использование памятных знаков, повреждение лобных долей нарушает такой вид поведения. Выполнение такой з?дачи страдает также при удалении всех частей лимбической системы (Pribram, Wilson and Connors, 1962).
Таким образом, имеются и поведенческие и анатомически» основания для объединения лобных полюсов мозга с лимбиче-скими образованиями (Pribram, 1958, см . рис. XVII-1 и табл-XVII-1). Разрушение этих систем не сказывается на дифферен-цировке знаков, но нарушает выполнение таких задач, как отсроченное чередование (Pribram et al., 1952, 1966; Pribram, Wilson and Concors, 1962), переделка дифференцировки (Pribram, Douglas, Pribram, 1969), поведение приближения или избегания («пассивное» избегание) (McCleary, 1961). При выполнении всех этих задач возникает конфликт в тенденциях ответа, конфликт между установками. Правильный ответ зависит от контекста (то есть от состояния), а контекст изменяется в результате задачи, стоящей перед данным организмом. Таким образом, для получения правильного ответа должна возникнуть внутренняя установка (то есть определенное состояние готовности мозга). Создание такой установки зависит от механизма памяти, включающего систему хранения и воспроизведения установок, возникших в результате периодически повторяющихся функций. (Формальные свойства систем памяти подобного типа были детально описаны Квиллианом, 1967.) Связь лимбических систем по принципу замкнутой петли всегда подчеркивалась анатомами, а это делает эту систему идеальным механизмом, определяющим зависимость действия от контекста (Pribram, 1961; Pribram and Kruger, 1954).
Кроме того стоит отметить, что социально обусловленное эмоциональное поведение в очень большой степени зависит от контекста воспринимаемых воздействий. Это заставляет предположить, что значение лимбических формаций для эмоционального поведения обусловлено не только анатомической связью с гипо-таламическими и мезенцефалическими структурами, но также ее собственной организацией по принципу замкнутой петли, поддерживающей раз возникшее возбуждение. Остается доказать (хотя некоторые предварительные данные уже имеются; Fox, et al., 1967; Pribram, 19676), что префронтальная кора оказывает влияние на сигналы лимбической системы примерно так же, как задняя кора участвует в предварительной переработке сенсорных сигналов.
Таким образом, связь лобной коры с двигательными механизмами, с одной стороны, и с лимбическими образованиями – с другой, предполагает участие лобной доли в процессе, посредством которого осуществляется связь действия, с одной стороны, с потребностями и эмоциями – с другой. Поэтому способность организма пользоваться побуждающими знаками, или символами, столь зависящая от лобной коры, возникает, по-видимому, в результате этого взаимодействия.
ЭМОЦИИ И ПАМЯТЬ
Это предположение подтверждается многими фактами. Возможно, наиболее убедительным из них является еще один ряд нейропсихологических парадоксов. Как указывалось в начале этой главы, лоботомия, или лейкотомия, у человека применялась в качестве лечебной процедуры. Применение лоботомии на человеке непосредственно связано с уже описанными экспериментами на обезьянах. Мы видели, что хотя после резекции лобных долей животные оказывались неспособными к выполнению отсроченных реакций, они не проявляли никаких признаков беспокойства в связи с этим. Казалось, что может быть проще, чем ампутировать у больного орган, ответственный за беспокойство? (См. рис. XVIII-4).
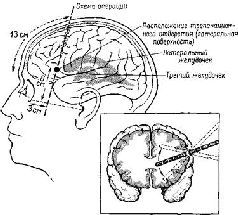
Рис. XVIII-4. Схема операции фронтальной лоботомии, показывающая место введения электрода на латеральной и фронтальной проекциях мозга.
Несмотря на то что такая операция была в известной мере эффективной, исследователи столкнулись с парадоксом, обнаружив, что фактически у больных, перенесших лоботомию, сохранялась способность к решению задач. Пытаясь проанализировать этот парадокс, я провел эксперименты, показавшие, что лобото-мированные обезьяны (а также человек) в действительности проявляли некоторое беспокойство при фрустрации, но это беспокойство было более кратковременным, чем у контрольных испытуемых (Pribram and Fulton, 1954). Этот эксперимент заключается в том, чтобы научить животное правильно дифференцировать сигналы в 100% случаев, после чего время от времени вводились тестовые пробы, не сопровождавшиеся подкреплением. В результате этого возникало беспокойство, отрицательно сказывавшееся на выполнении дальнейших проб, следовавших с равными интервалами. Определялось время, необходимое для достижения установленного критерия (100% правильных ответов). Хотя беспокойство оказывало столь же пагубное влияние после лоботомии, как и до нее, это нарушение продолжалось не так долго. Результаты этого эксперимента привели к предположению, что лобная кора обычно увеличивает устойчивость эффекта неудачи, причем это предположение согласуется с представлением -о важной роли лобной доли приматов в процессах кратковременной памяти. Однако обычные тесты на кратковременную память у людей, перенесших лоботомию, например опыты с припоминанием недавно прочитанных номеров телефонов, не смогли обнаружить никакого отклонения от нормы, хотя болезненные аффективные реакции иа неудачу уменьшились.
Но это не исчерпывало наблюдавшихся явлений. В целях излечения некоторых форм эпилепсии нейрохирурги начали вторгаться также и в височную долю человеческого мозга. Казалось-бы, результаты экспериментов с животными свидетельствуют о том, что операции в лимбических областях височной доли приводят к радикальному изменению аффектов. Однако ничего подобного после такой операции у человека не произошло. Вместо этого после резекции можно было наблюдать специфические нарушения памяти. Непосредственная память, как показали тесты с цифрами, оставалась неповрежденной. Так, больной мог припомнить весьма детально свои впечатления перед операцией в сохранял ранее приобретенные перцептивные и двигательные навыки. Однако его способность регистрировать текущие события в долговременной памяти оказалась нарушенной. Стоило прервать серию опытов и возобновить ее через несколько минут или часов, и больной не только не мог припомнить данные им ответы, но даже не имел представления о том, что ему задавали какие-то вопросы или что он вообще принимал участие в эксперименте (Milner, 1958; рис. XVIII-5).
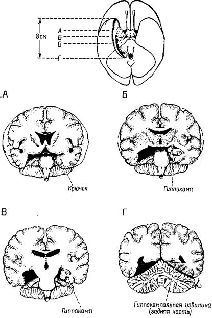
Рис XVIII 5 Схемы поперечных срезов мозга человека, показывающие расположение двусторонней резекции медиальных отделов височной доли, включающей миндалевидное ядро и гиппокамп и приводящей к значительным нарушениям мнемических процессов (Milner, 1959).
Если бы не этот двойной парадокс, можно было бы легко пренебречь противоречивыми результатами хирургических операций на лобной и височной долях в отношении памяти и эмоций, объясняя их видовыми различиями. Конечно, эти различия представляют собой очевидный фактор, но чем объясняется это удивительное сближение функций памяти и эмоций? Может быть, нарушение функции в основном одинаково у приматов и у человека, но то, что отличает человека, например его владение речью, приводит к существенным различиям между людьми и животными, перенесшими операцию? Возможно, поражение лобной доли сказывается у обезьян во время решения задач, а у человека в тех социальных взаимодействиях, которые протекают без участия речи? Так ли это при лимбических нарушениях?
Что касается лобных долей, то на эти вопросы можно дать утвердительный ответ. Сохранная способность больного пользоваться речью позволяет ему решать задачи в опытах с отсроченными реакциями. Однако задачи, слегка усложненные по сравнению с простой отсроченной реакцией, которые, как указывалось, не в состоянии были решить обезьяны, не могли выполнить и люди, перенесшие лоботомию (даже через десять лет после операции), если только эти задачи не сопровождались словесными инструкциями (Poppen, Pribram and Robinson, 1965). Так, больные, которым давалась задача на отсроченную реакцию, говорят,., что они словесно кодируют местоположение спрятанного объекта и таким образом могут припомнить это место после операции, даже если операция происходит после того, как был спрятан объект.
Не так легко разрешается парадокс, касающийся лимбическизг повреждений. Прежде всего мы не вполне уверены в том, что при" резекциях височной доли у человека повреждаются только лим-бические структуры. Тяжелая потеря памяти, наблюдаемая у больных, может быть в известной степени воспроизведена у обезьян, когда повреждение захватывает нижнюю латеральную поверхность височной доли, то есть ту часть, которая, как указывалось в предыдущей главе, участвует в опознании и узнавании знаков (Weiskrantz, 1967). Такие обезьяны не в состоянии выработать дифференцировку, возможно, только вследствие разрушения ассоциативной коры височной доли. Когда же резекция ограничивается медиальной частью височной доли, где расположены лимбические структуры (амигдала и гиппокамп), выработка дифференцировки, по крайней мере у обезьян, не нарушается (Pribram, Douglas, and Pribram, 1969).
КРАТКОВРЕМЕННАЯ ПАМЯТЬ – КОДИРУЮЩИЙ МЕХАНИЗМ, ОБЕСПЕЧИВАЮЩИЙ СОХРАНЕНИЕ КОНТЕКСТА
Предположим, что дело обстоит иначе и рассмотрим гипотезу,, согласно которой фактически существует какая-то единая функция мозга, лежащая в основе как эмоциональных, так и мнести-ческих процессов, причем на эту функцию оказывают влияние лимбические и лобные повреждения. Мы уже видели, что лобные повреждения не препятствуют возникновению эмоционального поведения, лишь сокращая его продолжительность в ситуации фрустрации. Задача, связанная с отсроченной реакцией, также требует длительного сохранения ответа. Может быть, лобная (и лимбическая) резекция просто ускоряет стирание следов памяти? Я подверг эту гипотезу проверке в следующем эксперименте (Pribram, Tubbs, 1967; рис. XVIII-6). Обычно пробы при отсроченном чередовании делаются через равные интервалы (правый ящик – 5 сек, левый ящик – 5 сек; правый ящик – 5 сек, левый ящик – 5 сек и т. д.). Вместо этого я использовал спаренные пробы – правый ящик – 5 сек – левый ящик – 15 сек; правый ящик – 5 сек – левый ящик – 15 сек и т. д. Почти сразу же-обезьяны с лобными повреждениями начали действовать правильно, несмотря на то, что вводились большие (15-секундные) интервалы. Таким образом, гипотеза о более быстром стирании; следов памяти не подтвердилась.
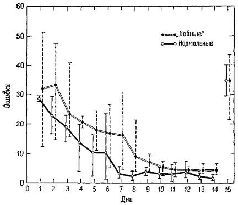
Рис. XVIII-6 Модификация задачи па чередование, упоминавшейся ранее (см. рис. XVIII-1), с которой легко могли справиться обезьяны с частичным удалением лобной коры. Обезьяны с поврежденным мозгом были не способны решить стандартную задачу на чередование, даже если интервал между пробами составлял только 5 сек. Тогда задача была изменена таким образом, что интервалы между пробами составили следующую последовательность: правый стимул – интервал 5 сек, левый стимул – интервал 15 сек, правый стимул – интервал 5 сек, левый стимул – интервал 15 сек и т. д. Как только были внесены эти изменения, обезьяны с удаленными лобными долями стали выполнять задания так же хорошо, как и нормальные животные, что и видно на данном рисунке. Верхняя линия – кривая выполнения задания группой «лобных» обезьян; нижняя линия – группой нормальных обезьян. За ошибки принималось число дневных проб, необходимых для достижения 40 правильных ответов. Вертикальные линии обозначают разброс ошибок, допускаемых различными обезьянами. Данные за 15-й день показывают результаты опыта, когда все пробы снова были разделены равными интервалами по 5 сек (Pribram, Tubbs, 1967).
Идея провести этот эксперимент возникла в связи с любимым примером У. Мак-Каллока, который он приводит в качестве доказательства важности кодирования. Когда слова сливаются вместе, как во фразах:
угривтине
нетихвглине
смолавели
вдубенет,
трудно, а иногда и невозможно найти в них смысл. Но когда вводятся нужные паузы, кодирующие вереницу букв в отдельные куски, сразу же различаются слова: «Угри в тине» и т. д. Задача с чередованием может казаться обезьяне с лобным повреждением слитой в одно целое, ответ интерферирует с предыдущим и последующим. Введение группировки устраняет эту трудность. Организация событий в группы – простой способ обеспечить контекст, необходимый для основных форм кодирования, а также и для более сложного программирования.
Возможно, что у человека эта склонность к группировке и организации, к созданию и сохранению контекста, в пределах которого происходят какие-то события, может возрасти до того, что она перейдет границы нормы и явится причиной навязчивости и невроза. Уменьшение этой склонности путем лоботомии на ограниченном участке могло бы в этих случаях привести к успеху! Возможно также, что изменение длительности реакции беспокойства у обезьян, вызванное лоботомией, объясняется тем же самым изменением способности сохранять один и тот же контекст (см. Ornstein, 1969). После операции следы событий сохраняются очень недолго и полностью зависят от контекста воспринимаемых воздействий. Для описания этого аспекта поведения больных, перенесших лоботомию, в клинике употребляется выражение «привязанность к стимулу».
Таким образом, эти наблюдения и экспериментальные данные наводят на мысль, что лобная кора участвует в обеспечении и сохранении контекста, временной организации событий. В гл. Хи XV указывалось, что лимбические формации принимают участие в контроле над состоянием мозга. В известном смысле действие лобной коры на лимбический контроль состояний можно сравнить с действием задней «ассоциативной» коры на специфические сенсорные функции. Лобная кора участвует в организации пусковых Образов, создавая зависящие от контекста символы, тогда как задняя кора участвует в организации последовательности перцептивных Образов, обеспечивая сохранение свободных от контекста знаков. В обоих случаях имеет место влияние на Образы. Но это влияние не будет иметь успеха до тех пор, пока не будет предпринято некое действие, некое высшее выражение Образов.
Однако нервные структуры, принимающие участие в организации знаков, отличаются от тех, которые участвуют в символических процессах. Перцептивные и двигательные навыкиостанутся сохранными, несмотря на повреждения лобной и лимбической систем. Напротив, операция кодирования при группировке илвс расчленении событий, необходимая для решения задачи на отсроченное чередование, зависит именно от этих систем.
Какие же нервные механизмы принимают в этом участие?
САМОРЕГУЛЯЦИЯ ПОВЕДЕНИЯ
Пока еще на этот вопрос нельзя дать исчерпывающего ответа. Но некоторые указания относительно такого механизма можно получить, анализируя взаимоотношения между лобной корой и лимбической системой, а также процесс решения задач с отсроченным чередованием и с отсроченной реакцией. Ключ к правильному выполнению теста с чередованием (который сопровождается коррекцией, то есть повторяется до тех пор, пока не будет дан правильный отчет) можно найти в недавно завершенном опыте. Обезьяна должна была составить простой стратегический план, который учитывает контекст, созданный непосредственно предшествовавшим опытом. В этом случае ей очень помогает стратегия «перемещение – ответ». Однако следует отметить, что правильность поведения в любой момент зависит не от событий, происходящих в этот момент в окружающей среде, а от контекста, -созданного памятью (кратковременной). В гл. XV мы выяснили, что амигдала и гиппокамп, входящие в лимбическую систему, регистрируют и оценивают поведение. Поэтому не удивительно, что при повреждении этих структур организм неспособен выполнить задачу, в которой правильность ответа зависит от того, зарегистрирована ли (и оценена ли) предыдущая задача. Чтобы создать стратегический план, связанный с контекстом, контекст должен быть сформирован и сохранен.
Однако дефекты выполнения задач на чередование после лобной резекции можно объяснить по-другому. Контекст устанавливается, но организм, очевидно, не знает, как им пользоваться. Мы столкнулись с этой проблемой при объяснении результатов удаления задней «ассоциативной» коры. В этом случае (Bagshaw, Mackworth, Pribram, 1971) запись движений глаз показала, что -обезьяны нормально рассматривали объект и, очевидно, хорошо видели сенсорные сигналы, но не могли действовать на основе того, что они воспринимали.
У больных отчетливо проявляется эта неспособность регулировать поведение, когда смысл его зависит от контекста, в котором протекает их деятельность. Клинические наблюдения изобилуют примерами неадекватного поведения. Возможно, наиболее показательным из них является наблюдение, проведенное над одним из больных Г.-Л. Тёйбера (1964). Этот больной регулярно посещал лабораторию по четвергам. Когда один из четвергов совпал с праздником, он, как обычно, отправился в лабораторию, говоря себе на каждом шагу: «Сегодня праздник, мне не нужно идти в -лабораторию, там никого нет». Специальный контекст «праздника», хотя и вербализованный, не оказывал влияния на его поведение. Только когда он дошел до лаборатории и не обнаружил там никого, он вернулся домой, однако так и не сделав вывода о происшедшем.
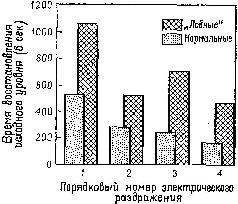
Рис. XVIII-7. График, показывающий время восстановления исходного уровня сопротивления кожи после электрического раздражения; средние данные по группам испытуемых (Grueninger et al., 1965).
Мы уже много говорили по поводу задачи на чередование. Нарушение, проявляющееся в классической прямой и косвенной формах задач на отсроченную реакцию после резекции лобных долей мозга, объясняет нам еще кое-что относительно взаимоотношений между восприимчивостью к контексту и кратковременной памятью. В этом случае правильность выполнения задания зависит не от контекста, установленного в результате какого-то предшествовавшего действия, а от временного появления сигнала, указывающего на начало пробы. Множество экспериментов показало, что правильная регистрация мозгом этого на мгновение появляющегося знака, способствующая его усвоению путем активной реакции на сигнал, является решающим условием верного выполнения задания (Pribram, 1969b). Что же включает в себя правильная регистрация? В нейрофизиологическом отношении, если нет некоторых изменений в электрической активности мозга, это приведет к ошибке. Именно такие изменения указывают на появление ориентировочной реакции и образуют основу для последующего привыкания. И действительно ориентировочная реакция резко изменяется под влиянием поражений лобных долей как у человека, так и обезьяны (Лурия, Pribram, Хом-ская, 1964).
Теперь уже хорошо известно, что имеется ряд индикаторов ориентировочной реакции (Е. Н. Соколов). К ним относится поворот глаз к стимулу, изменение частоты сердцебиений и дыхания, КГР, изменение кровенаполнения -периферических и центральных сосудов (измеряемое при помощи пальцевой и височной плетизмографии) и активация в ЭЭГ. После резекции лобных долей мозга (рис. XVIII-7), так же как после амигдалэктомии, остаются только поведенческие и электроэнцефалографические индикаторы (хотя продолжительность активации в ЭЭГ сокращается). Вместе с тем повторение сигнала в этих случаях уже не вызывает привыкания и поведенческие реакции продолжают сохраняться. Как уже подробно говорилось, это привело к предположению, что ориентировочная реакция состоит по крайней мере из двух отдельных процессов: из компонента настораживания, поиска и отбора стимулов и из их регистрации в- сознании и памяти. Лобная кора и амигдала оказывают влияние на этот процесс регистрации. Когда регистрация нарушается, организм не только утрачивает способность припоминать события, но и становится в гораздо большей степени подверженным ретроактивному и проактивному торможению, обусловленному интерферирующими воздействиями. Люди и обезьяны с лобными повреждениями чувствительны к интерференции, влияющей на процессы кратковременной памяти (Crueninger and Pribram, 1969; рис. XVIII-8). Данные электрофизиологии говорят о том, что лобная кора в норме контролирует интерференцию посредством кортикофугаль-ных влияний на афферентные системы. Электростимуляция лобной коры вызывает результаты, противоположные эффектам стимуляции задней ассоциативной коры (рис. XVIII-9). Раздражение лобных отделов мозга увеличивает избыточность каналов (см. рис. XI-6 и Х1-7; каналы имеют тенденцию в любой момент действовать все вместе. Таким образом, возможность интерференции между следами последовательных сигналов сокращается. Б проводившихся экспериментах длительность вызванного изменения равнялась миллисекундам; ретро- и проактивная интерференция в поведенческих ситуациях занимает значительно большее время. Тем не менее приведенные электрофизиологические данные наводят на мысль, что, по-видимому, в поведенческих ситуациях, в которых наблюдаются эффекты интерференции, лобная кора обеспечивает более длительное сохранение возбуждения, чем нам удавалось достигнуть в эксперименте.
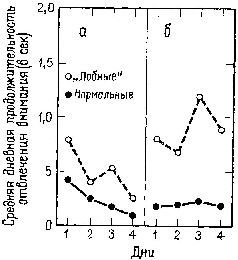
Рис. XVIII-8. График, показывающий среднюю дневную продолжительность отвлечения внимания (средний латентный период отвлечения внимания на пробу минус медиана латентного периода): а – для условий, когда стимул изменяется, а место его остается постоянным; б – для условий, когда изменяется место стимула, а сам ой постоянен (Grueninger and Pribram, 1969).

Рис. XVIII-9. Карты зрительных рецептивных полей, показывающие, как изменяется информация, поступающая по первичным зрительным путям, при стимуляции различных отделов мозга. Карта а – нормальный ответ клетки латерального коленчатого тела при движении источника света по экрану; карта в показывает, как уменьшается рецептивное поле при стимуляции нижневисочной коры; карта б свидетельствует о расширении рецептивного поля, вызванном стимуляцией лобной коры; карта г – конечная контрольная запись, сделанная через 55 мин после записи a (Spinelli and Pribram, 1967).
Приведенный анализ опытов с отсроченным чередованием и отсроченными реакциями позволяет предположить, что повреждение лобных долей вызывает нарушение тех процессов мозга, существенным элементом которых является кодирование изменений состояний. Такие процессы свойственны работе кратковременной памяти, предполагающей скорее решения, зависящие от контекста, чем хорошо упроченные действия, не зависящие от контекста, и нарушения их отражаются как на решении задач, так и на эмоциональном поведении. Можно предположить, что подобный дефект сказывается на решении задач потому, что организм не в состоянии регулировать свое поведение на основе событий, вносящих беспокойство и сигнализирующих об изменениях в контексте. Этот дефект проявляется в эмоциональном поведении: организм не справился с контролем, регистрацией и оценкой изменений, непрерывно усложняющих контекст и таким образом увеличивающих беспокойство в настоящем. При повреждении височной доли утрата способности к восприятию контекста проявляется в том, что поток событий не сохраняется в следах и, таким образом, протекает только в настоящем, которое не имеет ни прошлого, ни будущего. Организм становится автоматом, находящимся во власти своих мгновенных состояний, которые он не может регулировать.
ПРИПОМИНАНИЕ И СИМВОЛИЧЕСКИЙ ПРОЦЕСС
Остается проанализировать связь между кратковременной памятью и символическими процессами. Рассмотрим сначала память. Обычно считают, что процесс сохранения следов происходит примерно так: событие регистрируется в кратковременной памяти, где оно циркулирует в течение некоторого времени, как след в нервных цепях, а затем переводится в более долговременное хранение в молекулярной форме. Этот процесс переноса называется консолидацией следа памяти. Если рассматривать только процессы хранения следов, эту картину можно считать относительно верной. Мы уже отмечали, что привыкание зависит, вероятно, от замкнутой системы с обратной связью, которая уменьшает нервную активность. Ряд таких замкнутых контуров может успешно создать буферный механизм хранения следов в пределах сенсорных систем. Мы также отмечали, что подкрепление может продлить некоторые виды нервной активности, вызывая химические изменения и даже рост глиальной и нервной ткани. Изменения в этих основных механизмах могли бы легко объяснить данные, полученные при экспериментальной проверке гипотезы консолидации.
Но анализ процессов памяти только с точки зрения хранения следов не объясняет другие факты, связанные с запоминанием, – узнавание и припоминание. Узнавание предполагает наложение входного сигнала на след, хранящийся в долговременной памяти. Такое наложение во многих случаях может происходить непосредственно, без вмешательства кратковременной памяти, а это значит, что узнавание осуществляется благодаря параллельной обработке информации механизмом памяти, который может восстанавливать опознанный образ даже на основе информации, лишь частично воспроизводящей хранящийся сигнал. Узнавание, однако, не всегда бывает таким непосредственным. В более сложных ситуациях для узнавания может потребоваться некоторое время и оно может зависеть от более медленных процессов выравнивания кортикальных диполей и других процессов. Этот тип узнавания соответствует большинству классических нейрофизиологических точек зрения на процесс памяти.
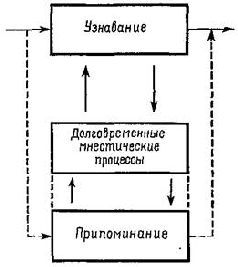
Рис. XVIII-10. Узнавание и припоминание в терминах концепции Т – О – Т – Е. Обратите внимание, что припоминание есть процесс предвосхищения. Ср. с рис. V-10.
Иной характер имеет процесс припоминания (рис. XVIII-10). Стимулирующее событие запускает механизм кратковременной памяти, который отыскивает в памяти наиболее подходящий материал. В случае узнавания настоящего поиска не происходит – подходящее отыскивается немедленно или очень быстро посредством кросс-корреляции между одновременно возникающими и интерферирующими фронтами волн. Поэтому узнавание «порт-ретно» по отношению к стимулирующему событию и независимо от контекста, в котором оно происходит. Напротив, при припоминании в поиск сходного образца вовлекаются некоторые произвольные «ассоциативные» контекстные связи. Согласно модели, описанной в гл. XVII, лобная стимуляция усиливает торможение побочных связей и, таким образом, повышает связь между модулями памяти. Вовлекаются большие сегменты голографического механизма, фокусное расстояние увеличивается настолько, что внимание сосредоточивается на той или иной детали, то есть концентрируется. Объем и локализация этих усилившихся связей между модулями памяти зависит от предшествующей истории этих модулей, от того, как их следы изменялись непосредственно предшествовавшим опытом. Эта установка выравнивания диполей служит, вероятно, топографическим эталоном, который, будучи активирован неспецифическим входом, запускает посредством опережающего процесса (рис. XVIII-10) реконструкцию «призрака» Образа, ранее ассоциировавшегося с этим эталоном. Возможно, что здесь кроется ключ к пониманию роли лобной коры в процессе припоминания и ее связи с символическими процессами.

