ЧАСТЬ ВТОРАЯ ОРГАНИЗАЦИЯ ПСИХОЛОГИЧЕСКИХ ПРОЦЕССОВ
«Мозг – это машина, создающая модели подобные себе».
Крейк
Глава VI
ОБРАЗЫ
Главная проблема нейропсихологического исследования состоит в том, чтобы определить, в каком отношении и до какой степени мозговые процессы и психологические функции скоординированы между собой. В первой части книги мы стремились разработать связный язык, связную систему терминов на основе экспериментов и наблюдений за функционированием простейших механизмов организации мозга. Во второй части мы обратимся к изучению психологических функций путем экспериментального анализа вербальных отчетов о субъективных переживаниях.
На протяжении последней половины XIX в. субъективный опыт редко был предметом научного изучения. Вместо этого внимание исследователей было сосредоточено на инструментальном или вербальном поведении, как таковом. Такой подход был, в общем, весьма успешным для количественного описания факторов. внешней среды, оказывающих влияние на поведение. Но он был менее пригоден для изучения параметров самого организма, наряду с факторами внешней среды определяющими его поведение. При таких обстоятельствах особенно возрастает ценность экспериментов, в которых помимо параметров внешней среды регистрируются и некоторые физиологические показатели. Важно,. однако, отметить, что нельзя обойтись без такого поведенческого-подхода, хотя многие клинические неврологи и физиологи мозга игнорировали анализ соответствующей окружающей среды, некритично отождествляя наблюдаемые ими физиологические и психологические процессы. Таким образом, психологические процессы стали изучаться, с одной стороны, бихевиористами и, с другой – физиологами. Первые бихевиористы провозгласили, что невозможно дать определение субъективных состояний и поэтому научный психологический язык должен полностью исключить-термины, описывающие наши внутренние переживания. В то же иромя исследователи, имевшие медицинскую практику, потеряли всякий интерес к таким психологическим функциям, как волевое действие, аффективное состояние или воображение, из-за того, "что о последних судили на основании не поддающихся контролю субъективных ответов при отсутствии точного определения понятий, которыми пользовались психологи, что очень затрудняло понимание того, о чем, собственно говоря, шла речь.
В 60-е годы XX в. поведенческая психология оценила основную идею гештальтпсихологии о том, что субъективно переживаемое сознание является столь важной составной частью биологического и социального мира и что ею нельзя игнорировать при изучении поведения.
Таким образом, «респектабельные» психологи начали изучение таких проблем, как познание, мышление и внимание. А к концу 60-х годов стала открыто и без особого риска обсуждаться на встречах психологов проблема формирования образов, составляющая предмет настоящей главы.
Это расширение круга проблем психологического исследования произошло, разумеется, в результате того, что была сделана попытка строго определить те операции, которые делают возможным научное описание субъективных процессов. Мой собственный метод (рис. VI-1) состоит в том, что сначала я пытаюсь описать основные категории, относящиеся к свойствам организма и окружающей среды, не прибегая к поведению, затем я изучаю взаимодействие между этими категориями (которые являются независимыми переменными эксперимента) и рассматриваю поведение как Зависимую переменную. На основе данных, полученных в таком эксперименте, я делаю заключение о психологических функциях и устанавливаю их сходство (или отличие) с субъективными переживаниями, о которых сужу по словесному отчету испытуемых. И только тогда, когда удается обнаружить соответствие между ними, я перехожу на язык, типичный для субъективной психологии (Pribram, 1962, 1970).
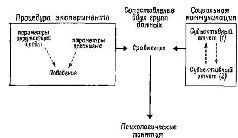
Рис. VI-1. Схема взаимоотношений мозга и поведения.
Это обращение к субъективным терминам не случайно. Во-первых, множество клинических неврологических данных получено на основе анализа вербальных отчетов о субъективных переживаниях при повреждении мозга или его электрическом раздражении. Во-вторых, я понял, что бихевиористский жаргон (которым я пользовался в своих рассуждениях) изобилует противоречиями, которые не смогут получить разъяснения до тех пор,, пока я не признаю правомерность субъективного метода. Иными словами, я должен был обратиться к терминам известной книги Гилберта Райла (1949) «Призраки в машине» («Ghosts in the Machine»). Образы и чувства – это призраки, которые населяют мой собственный субъективный мир, как и субъективный мир моих пациентов. Они наши постоянные спутники, поэтому я хотел бы больше знать о них. И хотя эти призраки заключены в машине, которая называется мозгом, мы не можем пока точно определить их. Если же мы будем их игнорировать, то окажемся лишь перед бездушной поведенческой машиной. Меня же интересуют призраки – психологические функции, а не сама эта машина-мозг и не осуществляемая ею регуляция поведения.
В ходе работы над этой книгой мне стало ясно, что определенного успеха можно было бы достигнуть, рассматривая взаимоотношения между мозгом, поведением и психологическими функциями именно в свете двухпроцессного механизма функционирования мозга. Может быть, такие образования, как образы и чувства, зависят от каких-то состояний центральной нервной системы? Мы попытаемся дать ответ на этот вопрос во второй части книги, а в третьей части дополнить его и определить, может ли поведенческий акт осуществляться за счет нейронных операций, которые в свою очередь определяются этими состояниями.
ГИПОТЕЗА ОСОЗНАНИЯ
Большинство неврологических и поведенческих исследований, посвященных проблеме образа, было сосредоточено на разработке методики для определения того, каким образом протекает восприятие тех или иных структур. Как осуществляется этот процесс восприятия конфигураций сигналов? Этим вопросом интересовались не только психологи и неврологи, но и инженеры. Техническое моделирование «восприятия» структуры, безусловно, важно для таких проблем, как разработка машинного перевода и создание распознающих машин. В настоящее время устройства, реагирующие на конфигурацию сигналов, приобрели важное значение для конструирования роботов и автоматов, которые могут заменять человека во многих узкоспециализированных видах деятельности. Даже в элементарных автоматических устройствах важное значение приобретает создание соответствующего представления об окружающей среде, в которой должен действовать робот, а также сохранение этого представления в памяти. Такая машин» (рис. VI-3), если она имеет «карту» местности, в которой он» должна действовать, может решать более сложные проблемы, чем простое выполнение задач в весьма ограниченных и строго детерминированных условиях. Устройства, «воспринимающие» такие структуры, тем или иным способом создают некоторое пространственно закодированное представление, карту своего окружения,, сформированную на основе своего опыта. Таким образом, они могут «научиться» по-разному отвечать на специфические структуры раздражителей, они могут «узнавать» знакомую структуру и отличать ее от новой.

Рис. VI-2. А – Speculatrix – черепаха Грея Уолтера находит путь домой; Б – схема Speculatrix (Грей Уолтер, 1966).
Конечно, автомат может быть сконструирован таким образом, что ему не нужно будет обращаться к «карте»: черепаха Грея Уолтера (1966; см. рис. VI-2) и гомеостат Росса Эшби (1962) -два примера первых искусственных устройств из множества механизмов, обладающих способностью двигаться, избегать препятствий и выбирать правильный путь. Эти первые искусственные меха-пизмы имели весьма ограниченные возможности, пока они не были снабжены другим устройством, которое может реагировать на-структуру сигнала.
Психологи в период расцвета бихевиоризма пытались объяснять поведение организмов и особенно процесс научения, не прибегая к понятию образа. Однако очень скоро стало ясно, что бе» такого построения, как «карта» (Tolman, 1932), объяснительные-возможности бихевиористских наблюдений, хотя они, разумеется, и имелись, весьма ограниченны. Не так давно вновь было признано значение концепции образа: сторонники психологии познавательных процессов, анализируя процесс научения, опосредствованного речью, столкнулись с разнообразием процессов формирования образов, потребовавших неврологического обоснования. Почему же психологи колебались в отношении того, признать ли образ в качестве основного и эффективного объяснительного-принципа? Это было иызвано двумя достаточно вескими причинами: 1) до недавнего времени было немного строгих экспериментальных данных (если они вообще были), говорящих о том, как происходит формирование образа, и 2) существовали большие трудности в понимании неврологического механизма, который со-вдавал бы и сохранял бы – в соответствующей форме – пространственно закодированное представление о жизненном опыте организма.
Причины таких колебаний существовали недолго. Недавние исследования поведения дали обоснование проблеме образа, а неврологические исследования, как и открытия в области изучения информационных процессов, сделали понятным механизм, вызы-пающий этот неуловимый процесс формирования таких «признаков». Изложению этих данных и посвящена настоящая и несколько последующих глав книги.
Любая создаваемая нами модель процессов восприятия должна учитывать два обстоятельства: значение образа, включающего в себя часть субъективного опыта человека, а также тот факт, что существуют такие влияния на поведение, которые мы не осознаем. Инструментальное поведение и его осознание часто противоположны друг другу: чем эффективнее выполняемые действия, тем меньше мы их осознаем. Шеррингтон выразил этот антагонизм в следующем кратком тезисе: «Между рефлекторным действием и сознанием существует, по-видимому, настоящая противоположность. Рефлекторное действие и сознание как бы взаимно исключают друг друга – чем больше рефлекс является рефлексом, тем меньше он осознается». Таким образом, если исследование носит исключительно бихевиористский характер, возникает опасность игнорирования некоторых проблем. Здесь я хотел бы подробно остановиться на рассмотрении именно этих часто игнорируемых проблем. (Проблемы различения, распознавания той или иной структуры и подобная категоризация выходят за пределы этой и следующих двух глав. Однако то, что говорится здесь, относится к значительно большему кругу проблем, которые обсуждаются в гл. XIV и XVII.)

P и с. VI- 3. A – робот Wanderer, созданный в Стэнфордском исследовательском институте; Б – Wanderer-самоуправляющийся робот, изучающий незнакомую ему комнату, подразделяя ее на воображаемые зоны и регистрируя в каждой зоне то, что он воспринял. Затем этот образ становится познавательной «картой мира» робота (Block and Ginsburg, 1970).
Реципрокные отношения между поведением и его осознанием, по-видимому, лучше всего можно проиллюстрировать на примере психологических процессов навыка и привыкания. Когда организм повторно оказывается в одной и той же ситуации, находится в неизменной среде, происходит следующее: если он должен постоянно выполнять сходное задание в этой неизменной ситуации, задача начинает выполняться им фактически автоматически, то есть организм действует все более эффективно. Организм научился выполнять задание, и у него выработались определенные навыки. Одновременно субъект обнаруживает привыкание: у него больше не возникает ориентировочная реакция, он уже не замечает тех явлений, которые постоянно повторяются в его окружении при выполнении задания. Его вербальные интроспективные отчеты, отсутствие движений головой и глазами в направлении стимула, такие электрофизиологические индикаторы, как кожногальваническая реакция, плетизмограмма и ЭЭГ, свидетельствуют об исчезновении ориентировочной реакции при повторении однообразных воздействий в неизменной ситуации. Однако, как было отмечено, привыкание свидетельствует не о потере чувствительности в какой-то части нервной системы, а, скорее, о создании некой нервной модели окружающей среды – представления, ожидания, иначе говоря, одного из видов механизма памяти, с которым непрерывно сопоставляются входные воздействия. Таким образом, нервная система благодаря входным воздействиям непрерывно настраивается, подготавливается к обработке новых входных воздействий.
Привычные действия организма также являются результатом нервной активности. В случае, когда имеет место ожидание, входной процесс при повторении стимуляции, по-видимому, ослабевает; когда же речь идет о выполнении задания, эффективность выходного процесса, вероятно, усиливается. Поэтому возникает вопрос: чем отличаются эти два вида нервной активности, которые формируют, с одной стороны, осознание, а с другой – навык и привыкание, находящиеся в реципрокных отношениях?
Нервные импульсы и медленные потенциалы являются двумя видами процессов, которые могут функционировать реципрокно. Естественно было бы предположить, что, чем эффективнее осуществляется процесс превращения структуры афферентных возбуждений в эфферентные, тем менее долговечна композиция, сформированная микроструктурой медленных потенциалов. При возникновении навыка и привыкания поведение становится рефлекторным – при этом более или менее устойчивые композиции узора медленных потенциалов соответствуют осознанию. Из этого следует вывод, что сами по сзбе динамические структуры нервных импульсов и поведение, которое ими вызывается, непосредственно не осознаются. Так, даже процесс речи остается неосознаваемым в тот момент, когда слова произносятся. Таким образом, я придерживаюсь традиционной точки зрения: из всех явлений, происходящих в нашем мозгу, мы осознаем лишь некоторые.
Короче говоря, нервные импульсы, достигающие соединений, генерируют микроструктуру медленных потенциалов. Узор этой микроструктуры взаимодействует с тем, который уже имеется в нервной системе благодаря ее свойству спонтанной активности и предшествующему «опыту». Это взаимодействие усиливается тормозными процессами, и весь процесс порождает эффекты, сходные с интерференцией динамических структур, возникающей вследствие взаимодействия одновременно появляющихся волновых фронтов. Таким образом, микроструктуры медленных потенциалов действуют как аналоговые кросс-корреляционные устройства, создавая новые рисунки, которые запускают динамические структуры выходных нервных импульсов. Вполне возможно, что быстрые изменения в сфере осознания отражают длительность корреляционного процесса.
Какие факты подтверждают тезис, что электрическая активность соединений в центральной системе связана с процессом сознания? И, Камийя (1968) и другие (Galbraith et al., 1970; Engst-rom, London and Hart, 1971), используя методику инструментального обусловливания, показали, что людей можно легко научить различать, продуцирует их мозг или нет волны определенной формы, следующие с частотой примерно 10 раз з секунду, то есть так называемый альфа-ритм, даже если они сталкиваются при этом с трудностями в обозначении различный состояний сознания, которые они ощущают. Испытуемые, которые были способны определить «состояние альфа-ритма», утверждали, что оно характеризуется как состояние приятной расслабленности. Сейчас проводится много подобных экспериментов с тем, чтобы найти пути сокращения продолжительности процесса обучения, получившего широкое распространение в дзен-буддизме, у йогов и у психотерапевтов па Западе, цель которых – определение и достижение состояния удовольствия (рис. VI-4).
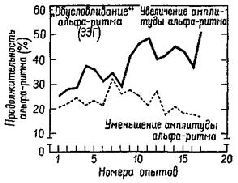
Рис. VI-4. График результатов, полученных в экспериментах (каждый длительностью в одну минуту), в которых испытуемые должны были сначала произвольно генерировать альфа-ритм большой амплитуды (сплошная линия), затем вызывать его депрессию (пунктир). (Kamiya, 1970).
Более специфичными являются некоторые из недавних экспериментов Б. Либета (1966),в которых исследовался хорошо известный феномен. С тех пор как Г. Фритч (1880) и Э. Гитциг (1969) показали, что электрическое раздражение определенных участков мозга человека вызывает движение, нейрохирурги исследовали нею поверхность мозга, чтобы определить, какие реакции возникают в ответ на раздражение. Например, О. Форстер (1936) создал карту областей постцентральной извилины, раздражение которых вызывало ощущение в той или другой части тела (рис. VI-5).
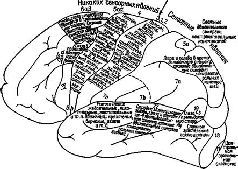
Рис. VI-5. Суммарные данные, полученные О. Форстером в результате электрического раздражения различных точек коры головного мозга человека (1926) (Бернштейн, 1966).
Так, ощущения покалывания или определенного положения тела в пространстве могут быть вызваны и при отсутствии каких-либо заметных изменений в самом теле, которые обычно переживаются пациентом как ощущения. Либет показал, что ощущение, вызываемое стимуляцией, возникает не сразу: проходит минимум полсекунды и максимум 5 мин, прежде чем пациент начинает что-либо ощущать. Видимо, электрическая стимуляция должна создать некоторое состояние в мозговой ткани и, только когда это состояние возникает, пациент начинает что-то чувствовать (рис. VI-6).

Рис. VI-6. Схема взаимоотношений между последовательностью прямоугольных импульсов (пороговой интенсивности и длительностью 0,5 мсек), приложенных к постцентральной извилине, и амплитудой прямых ответов коры (ПОК), записанных вблизи области раздражения. Третья линия показывает, что субъективные ощущения не возникают примерно в течение первых 0,5 сек стимуляции. Интенсивность появившегося после этого периода ощущения сохраняется на постоянном уровне, несмотря на то, что стимуляция продолжается (Libet, 1966).
ВОСПРИЯТИЕ
Сформулированная в этой главе гипотеза имеет во многих отношениях решающее значение для последовательно развиваемой здесь точки зрения на функции нервной системы. Но она не бесспорна. В первой части книги был дан обзор логических операций, которые выполняются нервной системой. Здесь я хочу показать, каким образом эти операции делают возможным осознание организмом того, что происходит в данный момент и в данном месте, иначе говоря, как формируется восприятие настоящего.
Проблема состоит в следующем: если вы посмотрите на своего друга, а затем на его соседа, то вы сразу же понимаете, чем они отличаются друг от друга. Далее происходит небольшая интерференция – если только она действительно происходит – между тем, что вы видели в первый момент, и тем, что было воспринято в следующий. При слуховой форме коммуникации такое непрерывное быстрое узнавание – музыкальных фраз, фонетических сочетаний речи и т. д. – обычное явление. Традиционный взгляд на функции нервной системы сталкивается со значительными трудностями при объяснении непосредственности, точности и видимой многомерности мимолетных восприятий. Здесь, должно быть, действует единый процесс. Но каким образом он действует?
Д. Хебб детально рассмотрел эту проблему в первых трех главах своего классического труда «Организация поведения» (1949). Он пишет: «Следует решить, зависит ли восприятие 1) от возбуждения специфических клеток или 2) от структуры возбуждений, местоположение которой не имеет значения». Хебб сделал следующий вывод: «Восприятие зависит от возбуждения специфических клеток в какой-то точке центральной нервной системы».
Когда накопились нейрофизиологические данные, особенно полученные в микрозлектродных исследованиях Юнга (1961), Маунткасла (1957); Матураны, Леттвипа, Мак-Каллока и Питтса (1960); Хьюбела и Визела (1962), точка зрения Хебба получила подтверждение. Исследования, проведенные с помощью микроэлектродов, показали существование нейронов, которые отвечают только на то или иное свойство стимула, например на направление движения, наклон линии и т. д. Я думаю, что сегодня большинство нейрофизиологов согласны с мнением Хебба о том, что единица восприятия соответствует нейронной единице (рис. VI-7).
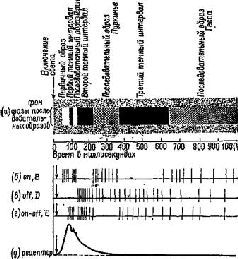
Рис. VI-7. Корреляция субъективных зрительных последовательных образов человека и разрядов нейронов в сетчатке и зрительной коре кошки после кратковременной вспышки света (300 лк) (взято из: Grusser and Grutzner, 1958, с изменениями): а – схсма сменяющих друг друга фаз последовательных образов (по: Froh-lich, 1929). Время в миллисекундах, б-г – схематическое изображение ответов on, off и on – off нейронов сетчатки и различных типов (В, D, Е) кортикальных нейронов; д – ответ рецептора наружного плексиформного слоя сетчатки при внутриклеточной регистрации. Вспышка света указана стрелкой, направленной вниз. В б-д стрелка сдвинута вправо, потому что в экспериментах Фрелиха латентный период ощущения на 20-40 мсек длиннее латентного периода ответов у кортикальных нейронов кошки. Затененная область, окружающая последовательные образы, означает фоновое освещение и фон собственного света сетчатки. В схеме объединены сетчаточные и кортикальные ответы, хотя последние характеризуются более низкой частотой и большей периодичностью разрядов. Пауза в первичной активации кортикальных нейронов В совпадает с начальным разрядом нейронов Е. При продолжительном освещении и то и другое соответствует «черной полосе» Шарпантье (Jung, 1961), а при кратковременной вспышке – вероятно, первому темному интервалу. Когда Грюссер л Грюцнер описали в реакции on – off нейронов паузу между 200 и 450 мсек, они соответствующим образом внесли поправки в свою схему (Jung, 1969).
Гипотеза, предложенная в первой части этой главы, говорит о том, что выбор, сделанный Хеббом, был единственно возможным и что возникновение структур возбуждения должно зависеть, конечно, от возбуждения специфических клеток, но что эти динамические структуры становятся в какой-то степени независимыми от клеточных единиц, так как определяются анатомией соединений, синаптическими и дендритными микроструктурами мозга. При соответствующих условиях эти узоры, образуемые с помощью синаптических систем, служат неврологическими эквивалентами единиц восприятия. Я согласен с предположением Хебба о том, что то, что мы осознаем, во многом зависит от прошлого опыта организма, но я буду отличать это явление от того непосредственного психологического настоящего, принимающего форму Образов, на котором основана предлагаемая гештальтпсихологией целостная концепция восприятия. Хебб смешал историческое развитие процесса узнавания с тем, что составляет содержание Образов. Чтобы составить программу, которая позволит компьютеру произвести вычисления, требуются многие часы работы, сама же машина выполнит эти вычисления за микросекунды. Необходимо много времени, чтобы научиться узнавать незнакомые структуры, но даже младенцы в возрасте нескольких недель (см. гл. VIII) могут правильно определять относительный размер фигур, находящихся на различном расстоянии (константность величины), и дифференцировать фигуры в различном положении (константность формы). Научение – это только часть проблемы узнавания. Гештальтисты в значительной мере были правы в своем нативизме. Существуют врожденные нервные механизмы формирования Образов, но гештальтисты были неправы, когда утверждали, что это единственный механизм восприятия. Однако сейчас маятник качнулся далеко в другую сторону и существует опасность, что целый ряд интересных явлений выпадет из поля зрения исключительно из-за интереса к проблеме, каким образом мы учимся распознавать.
Когда Хебб писал свою книгу, существовали две основные точки зрения на функционирование нервной системы. Одна из них была хорошо обоснована, другая – нет. Первая касалась вопросов генерации нервных импульсов и их передачи через «инапсы. Другая имела отношение к электрическим полям, создаваемым потенциалами мозга. В. Кёлер построил свою гешталь-тистскую концепцию на основе этих неироэлектрических полей и потерпел неудачу, доказывая их повсеместное существование десять лет спустя после опубликования Хеббом своих взглядов.
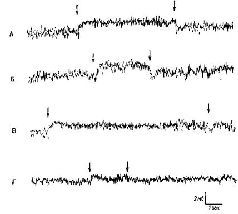
Рис. VI-8. Сдвиг постоянного потенциала (ПП) в слуховой коре в ответ на звуковую стимуляцию. «Активный электрод» – на твердой оболочке мозга над средней эктосильвиевой извилиной; другой электрод – на фронтальной надкостнице. Отклонение вверх означает негативность на твердой оболочке относительно кости. Л – сдвиг ПП в ответ на белый шум; Б – сдвиг ПП в ответ на тон 4000 гц; В – сдвиг ПП, возникающий в ответ на белый шум, возвращается к нулевой линии до прекращения звуковой стимуляции; Г – ответ на звуковые щелчки (50 в сек) и его возвращение к нулевой линии до прекращения стимуляции (Gumnit, 1960).
Я был счастлив, что смог принять участие в этих исследованиях. Экспериментальное наступление оказалось в целом успешным (КбЫег, 1958), и исследователи в моей лаборатории, а также в других лабораториях, регистрируя и вызывая в мозгу электрические поля постоянного тока, установили -их корреляции с функциями мозга и поведением (Gumnit, 1961; Stamm, 1961; Morrell, 1961). Всякий раз при стимуляции организма через тот или другой сенсорный вход (рис. VI-8) в соответствующих ограничейных областях мозга возникал сдвиг постоянного потенциала. Прямое раздражение мозга постоянным током могло задержать или ускорить научение в зависимости от полярности прикладываемого потенциала. Но я не согласился с Кёлером, когда он стал настаивать на связи между электрическими полями постоянного тока и восприятием. Позднее, когда я закончил эксперименты, в которых я накладывал на поверхность коры мозга алюминиевую пасту, мы снова смогли прийти с ним к согласию. Эксперименты показали, что различение структуры сигналов не нарушается, не-смотря на заметные изменения постоянного потенциала и ЭЭГ (рис. VI-9, VI-10).
А Б
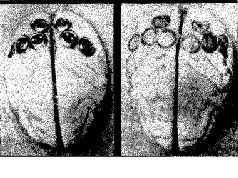
Рис. VI- 9. A – фотография мозга обезьяны со вживленными з него серебряными дисками, содержащими пасту; Б – фотография того же самого мозга после снятия дисков. В данном случав вживлепие было произведено в теменную кору. Другие эксперименты были проведены на затылочной, височной и лобной коре тем же способом или методом множественных точечных инъекций в кору алюминиевой пасты (Stamm and Warren, 1961).
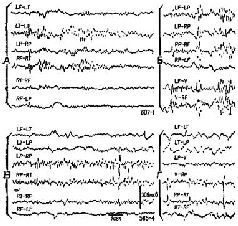
Рис. VI-10. Образец записи ЭЭГ у двух обезьян со вживленными дисками, как показано на рис. VI-9. А и Б – участки записи ЭЭГ до операции, В и Г – спустя 5 месяцев после вживления дисков с алюминиевой пастой. Биполярное отведение: L – левое, R – правое полушарие, F – лобная кора, Т – височная, Р – задняя темен-лая, V – макушка. Калибровка, как показано (Stamm and Warren. 1961).
Кёлер никогда не признавал экспериментов, проведенных Лешли (Lashley, Chow and Semmes, 1951), в которых для опровержения его теории и разрушения нейроэлектрических полей использовалась золотая фольга. Не признавал он также и экспериментов Сперри с перекрестом нервных волокон (Sperry, Miner and Meyers, 1955), в которых использовались изолирующие полоски слюды. Но когда он познакомился с результатами эксперимента со вживлением дисков с алюминиевой пастой, он воскликнул: «Это опровергает не только мою теорию поля постоянного тока, но и всякую другую современную неврологическую теорию восприятия!»
Позвольте мне коротко остановиться на тех данных, которые накопились в ходе этой дискуссии, чтобы рассеять такой мрачный взгляд на неврологию восприятия. Как было подробно показано в первой части книги, нервный импульс не является единственной важной электрической характеристикой нервной ткани. Другой характеристикой является микроструктура медленных потенциалов. Хотя последние близки к кёлеровским полям постоянного тока, они существенно отличаются от них тем, что они не диффузны, а строго локализованы на соединениях между нейронами или на дендритах и могут даже иметь вид миниатюрных спайков, которые чаще всего при распространении ослабевают. Проведение нервного импульса в любом месте нервной системы ведет к образованию на соединениях микроструктуры медленных потенциалов. Когда нервные импульсы достигают синапсов, генерируются постсинаптические потенциалы. Они никогда не бывают единичными, а образуют динамические структуры афферентных воздействий. Когда постсинаптические потенциалы возникают в дендритных полях мозга, они часто оказываются недостаточно большими, чтобы немедленно вызвать разряд нервного импульса. Поэтому динамические структуры постсинаптических потенциалов образуют узор, который имеет сходство с фронтом волны. Но этот рисунок медленных потенциалов не является каким-то таинственным фоном, миражем, наложенным на известную нервную функцию. Он представляет собой микроструктуру, состоящую из классических нейронных медленных потенциалов, возникающих в результате прихода нервных импульсов, ожидающих выхода через аксон.
Таким образом, структуры, образуемые на входе и выходе и принявшие форму микроструктуры медленных потенциалов, становятся источником третьей силы в нервной клетке. Все это говорит против концепции «плавающего» поля как возможного нейрофизиологического механизма, обусловливающего целостный характер восприятия. Потребность в этом третьем источнике была обнаружена до его открытия. Лешли глубоко волновала следующая проблема:
«Здесь мы сталкиваемся с такой дилеммой. Нервные импульсы проводятся по определенным, строго ограниченным путям, по сенсорным и моторным нервам и в центральной нервной системе от клетки к клетке и через-определенные межклеточные связи. Однако кажется, что все поведение детерминировано массой возбуждения, формой, отношением или соотношением возбуждения внутри общих полей активности безотносительно к отдельным нервным клеткам. Важна динамическая структура возбуждения, а не отдельный элемент. Какой тип нервной организации способен отвечать на такую структуру возбуждения при отсутствии специфических, ограниченных путей, обеспечивающих его проведение? Эта проблема имеет отношение к деятельности всей нервной системы, и необходимо сформулировать какую-нибудь гипотезу, чтобы дать направление дальнейшему исследованию» (Lashley, 1942, стр. 306).
Таким образом, Лешли предположил, что модель такой динамической структуры, возникающей в результате интерференции, была бы более адекватным объяснением этого феномена, чем любая из противоположных точек зрения. Однако он не имел ясного представления о такой модели, о том, каким образом может работать этот механизм. Он никогда не говорил о том, что «волны», генерируемые в результате прихода нервных импульсов, образованы хорошо известными классическими нейрофизиологическими процессами: синаптическими и дендритными потенциалами. Таким образом, он никогда не приводил аргументов в пользу существования микроструктуры соединений, в какой-то мере независимой от проведения нервных импульсов, то есть идеи, которая развивается на страницах данной книги. Лешли слишком связывал волновые формы с циркуляцией возбуждения по нейронам, которую он считал недостаточной, и в то же время разграничивал их, когда нужно было объяснять гибкость приспособления. Таким образом, он совсем не развил своей догадки. А ведь даже самая проницательная догадка, чтобы стать очевидной, нуждается в доказательстве. Прервем же на время наше изложение и приведем цитату Лешли, которая дает возможность проследить за ходом его мысли.
«Анатомические работы Лоренте де Но открыли в коре систему перекрестных связей, которые позволяют возбуждению распространяться по поверхности коры в любом направлении. Многие соседние нейроны способны к взаимному возбуждению, и вся система организована как сеть с петлями различной длины и сложности. Последние способны передавать импульсы от одной клетки коры к другой, или повторно возбуждать те точки мозга, которые первыми испытали воздействие стимуляции, посредством систем с возвратной циркуляцией с различными характеристиками.
Заключение о функциональных свойствах такой структуры организации может быть сделано с известной уверенностью. Возбуждение, возникающее в какой-либо точке, должно распространяться от нее по всей системе, поскольку угашение (возникающее в результате активации колец реверберации) будет возникать только после прохождения первоначального импульса. Если бы все системы были однородными, то возбуждение продуцировало бы серию радиально бегущих волн, так как первые волны возбуждения сопровождались бы волной угашения, за которой снова следовало бы возбуждение либо от последующих залпов, прибывающих по сенсорным путям, либо вследствие повторного возбуждения, возникающего на дальней границе зоны угашения. Длительность волн была бы одинаковой, ибо она» зависит от скорости проведения и рефрактерных периодов элементов всей системы. При наличии нескольких или многих точек возбуждения будут возникать интерферирующие динамические структуры.
Чтобы представить себе более простую картину, отвлечемся на время! от тех явлений, которые возникают в результате влияний от колец обратной связи; в этом случае происходило бы нечто аналогичное явлению распространения волн по поверхности жидкой среды. В такой системе интерференция волн создает структуру из гребней и впадин, которая характерна-для всякого пространственного распределения источников волнового движения и остается примерно одинаковой на всей поверхности. Можно предположить, что и в самой коре имеет место процесс, сходный в какой-то-степени с формированием структур возбуждения. Пространственное распределение в коре импульсов, пришедших к ней из сетчатки, не воспроизводит структуру возбуждения сетчатки, но создает условия для появления другой и специфической структуры стоячих волн, которая повторяется по» всей функционирующей области. Непосредственным препятствием этому является то, что возбуждение одной части поля может создавать в другой части состояние рефрактерности для приходящих от остальных частей-мозга импульсов и таким образом блокировать формирование единой динамической структуры. Однако если транскортикальные пути колец реверберации имеют случайную длину, как это, видимо, и есть на самом деле,. то в любой области они не все будут приходить в рефрактерное состояние одновременно и блокирования не будет» (Lashley, 1942, р. 313-314).
В последующих главах мы рассмотрим наиболее современную-и обоснованную точку зрения на «третью силу» в нейрофизиологии восприятия, представление о которой основано на признании существования двух типов микроструктур: нейронной микроструктуры и микроструктуры соединений.
РЕЗЮМЕ
Первые бихевиористы решительно отказывались от изучения субъективного опыта. В результате этого произошло значительное ограничение предмета психологии. Концепция двухпроцесс-ного механизма функционирования мозга устраняет это ограничение, вооружая психологов объективными методами и понятийным аппаратом механизмов соединений, с помощью которых можно изучать перцептивные образы, а также эмоции и мотивы.
Глава VII
ПРИЗНАКИ
НЕКОТОРЫЕ НЕЙРОАНАТОМИЧЕСКИЕ ПРОБЛЕМЫ ВОСПРИЯТИЯ
К числу главных факторов, препятствующих пониманию неврологии процесса восприятия, относится одно из тех противоречий, один из тех парадоксов, с которыми мы неоднократно встретимся в этих главах, пытаясь связать функции нервной системы с психологическими процессами. В данном случае парадокс касается неироанатомическои организации системы входа и тех нарушений в поведении, которые возникают в результате разрушения этой организации (рис. VII-1).
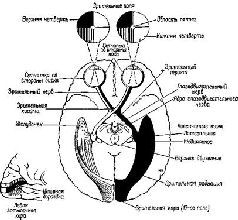
Рис. VII-1. Схема зрительных путей, как они выглядят на базальной поверхности мозга. Свет от верхней половины зрительного поля падает на нижнюю половину сетчатки. Свет от височной половины зрительного поля падает на сетчатку носовой стороны, тогда как свет от зрительного поля со стороны носа – на височную половину сетчатки (Truex and Carpenter, 1969).
В сущности, анатомическая организация систем- входа является единственной организацией, в которой сохраняется топологическое соответствие между рецепторной поверхностью и корой головного мозга. Это означает, что расположение точек на сетчатке будет проецироваться на кору в виде сходного расположения точек. Таким образом могут быть построены карты ретино-кортикального соответствия. Такие карты строятся на основе данных, полученных либо методом дегенерации нервных волокон после небольших разрушений в рецепторных системах (Browner, 1934; Browner and Zeeman, 1926), либо в результате точечной электрической или световой стимуляции сетчатки при одновременной регистрации электрических ответов от участков ‹коры, в которых они могут быть вызваны (Talbot and Marshall, 1941).
Подобные карты были пелучены для кожной чувствительности. По этому же принципу устроен и кохлеарно-кортикальный слуховой механизм (Rose and Woolsey, 1949).
Такая рецепторно-кортикальная организация не сопровождается прямым однозначным соответствием посредством единой нервной нити. Скорее всего, столь же сложная организация, которую мы уже анализировали на уровне сетчатки, повторяется как на каждом уровне механизма рецепторной системы, так и в коре. Фактически наблюдается общее сокращение числа клеток в ганглиозном слое сетчатки по сравнению с рецепторным (у человека это уменьшение оценивается примерно как 100 или 50 к одному). При переходе от слоя ганглиозных клеток к латеральному коленчатому телу таламуса число зрительных элементов, по существу, остается постоянным. При переходе от таламуса к коре возникает обратная картина: одиночная .клетка латерального коленчатого тела может контактировать с 5000 кортикальных нейронов каждый из которых находится в контакте еще примерно с 4000 других нейронов через их дендритные связи. Благодаря такому механизму тормозных взаимодействий при стимуляции у обезьяны двух точек сетчатки в фовеа, несмотря на некоторое перекрытие зон возбуждения, возникает их раздельное восприятие, причем таким образом, что двум угловым минутам на сетчатке соответствует расстояние в 1 мм на поверхности коры (Talbot and Mar-chall, 1941, p. 134). Казалось бы, что такое строение нервной системы обеспечивает проекцию некоторого «образа» от рецеп-торпой поверхности на поверхность коры во многом так же, как проецируется изображение в кинокамере. Парадокс проявляется, когда в результате болезни или хирургической операции повреждаются системы входа. Правда, как и ожидалось, в некоторых случаях может быть обнаружено выпадение части поля зрения (скотома) в месте, которое может быть предсказано в соответствии с анатомической организацией мозга (рис. VII-2).
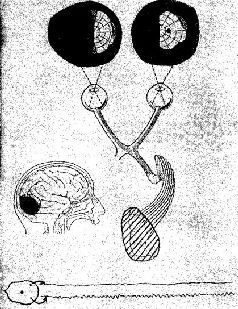
Рис VII-2. Карта зрительного поля, анатомическая схема зрительной системы 20-летней девушки с разрушением в правой затылочной долей ЭЭГ обеих зрительных долей (Case, 1942).
Однако при сохранении хотя бы малой части входного механизма это выпадение часто не замечается даже в том случае, если глаза удерживаются в неподвижности. При этом распознавание структуры сохраняется и во многих отношениях не отличается от нормального. Люди с громадными скотомами либо совсем не знают о них, либо могут быстро научиться легко их игнорировать. Животное, у которого было удалено или разрушено от 80 до 98% нервных связей входного механизма, способно решать задачи, требующие различения структур, отличающихся только деталями. Лешли (1929) удалял у крыс 80-90% стриарной коры, не нарушая их способность различать зрительные структуры. Р. Галамбос перерезал у кошек 98% волокон зрительного тракта, и животные еще могли хорошо выполнять задания, требовавшие различения весьма сходных фигур (Galam-bos, Norton and Frommer, 1967). Недавно Чау (1970), также экспериментируя с кошками, разъединял более 3/4 волокон зрительного тракта и удалял более 3/4 зрительной коры; в таких условиях едва ли оставалась неповрежденной любая из проекционных систем, характеризующихся точечным («точка в точку») представительством. Хотя зрительное различение структур после такого значительного разрушения вначале оказалось нарушенным, животное снова научилось выполнять задание, на что ему потребовалось примерно такое же число проб, как и до операции.
В моих экспериментах, как связанных с клинической нейрохирургией, так и проведенных в лаборатории (например, Wilson and Mishkin, 1959), удаление мозговой ткани было локальным и ограничивалось корой, так что оно не вызывало массивного разрушения белого вещества, благодаря чему у пациента или подопытного животного сохранялась в значительной мере способность к восприятию. После временно возникающей скотомы, длящейся несколько недель, можно было обнаружить лишь очень незначительные нарушения зрительной функции.
Как уже отмечалось, исследователи пытались использовать и многие другие методы для разрушения предполагаемой ими организации входных систем мозга, но безуспешно: Р. Сперри и его группа (1955) хирургически производили перекрестное рассечение области, воспринимающей сенсорные сигналы, и даже помещали полоски слюды в места рассечения, чтобы электрически изолировать маленькие квадратики ткани друг от друга. Лешли, Чау и Семмез (1951) пытались накоротко замкнуть электрическую активность мозга, накладывая полоски золотой фольги на воспринимающие поверхности. Я создавал в воспринимающей области коры множество эпилептогенных очагов посредством инъекции микроскопического количества алюминиевой пасты (Kraft, Obrist and Pribram, 1960; Stamm and Pribram, 1961; Stamm and Warren, 1961). Хотя такое множество очагов заметно задерживало научение различению зрительных структур, оно не препятствовало его проявлению, если научение уже закончилось или происходило после того, как были произведены многочисленные разрушения. Эти результаты показывают, что эффект входного сенсорного воздействия на мозговую ткань, получаемая на входе информация распределяются по всей входной системе.
Запись электрической активности мозга также свидетельствует о том, что в мозгу происходит распределение информации. Э. Р. Джон (John, Herrington and Sutton, 1967), например, при изучении зрительной системы использовал технику «меченых стимулов», предъявляя кошкам стимулы, которые различались не только своей геометрической структурой, но и частотой образующих эти стимулы мельканий. Эта различная частота мельканий отражалась на электрической активности мозга в ответ на предъявление стимула (или, если это мелькание было очень быстрым, то возникала субгармоника этой частоты). Таким образом, кодируемое различие частоты может оставить «след» в мозгу. Эта техника дала ряд интересных результатов, но для нас здесь важнее всего то, что тщательный анализ меченых волновых форм (вычисление позволяет обнаружить различие в волновых формах, возникающих в различных участках мозга) показывает, что одинаковые меченые волновые формы возникают во многих структурах мозга одновременно.
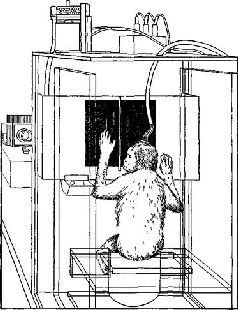
Рис VII-3 Рисунок, показывающий выполнение обезьяной задания на зрительное различение На находящейся перед обезьяной полупрозрачной панели она видит либо круг, либо вертикальные почосы, которые освещаются вспышками света в течение микросекунды Если обезьяна нажимала на правую половину панели при появлении круга и на левую – при появлении полос, то она шпучала вознаграждение – арахис, который падал в ящик под ее левой рукой Когда у нее вырабатывался навык в выполнении этого задания, электроды в зрительной коре обезьяны регистриро-1али волны определенной формы в ответ на структуру световою сигнала В начале экспериментов форма волн показывает, видит ли обезьяна круг или полосы По существу, они заранее сообщают, на какую половину панели будет нажимать обезьяна и будет ли ответ правильным или ошибочным (Pribram, 1969).

Рис VII-4 Результаты эксперимента на зрительное различение у обезьяны, условия его показаны на рис VII-3 Для характеристики реакции биотоков мозга на действие стимула в течение 3 дней производилось усреднение участков ЭЭГ, регистрируемых непосредственно после появления на панели круга или полос Для характеристики ответов животного использовалась запись ЭЭГ, непосредственно предшествующая тому моменту, когда обезьяна отвечала на стимул нажатием либо на левую, либо на правую половину панели Запись ЭЭГ производилась также во время подкрепления (арахисом), если обезьяна правильно отвечала на стимул, и в отсутствие подкрепления, еои она ошибалась
То, что видит обезьяна – полосы или круг, – находит отражение в различии волновых форм (второй впадины W – волны), возникающих на стимулы Только после того, как она научается правильному выпот-нению этого задания, появляются различия в волновых формах ЭЭГ, которые непосредственно предшествуют моменту нажатия левой ити правой половины панели Эти различия, по-видимому, не зависят ог того, видит ли обезьяна круг или полосы Таким образом, вотаовые формы в ЭЭГ отражают ее намерение нажать на определенную половину панели и ничего не говорят о том, будет ли ее ответ правильным или ошибочным Однако имеется третий вид различия в волновых формах, записываемых во время подкрепления Оно показывает, было ли полечено подкрепление медленный сдвиг нулевой линии, следующий за двигательной реакцией, указывает, что животное ожидает награды, а появление в ЭЭГ волн частотой 25-30 гц свидетельствует оегоразоча ровании (Pribram, 1969)
Однако другая группа экспериментов, проведенных в моей лаборатории (Pribram, Spinelli and Kamback, 1967, рис. VII-З, VII-4), показывает, что если научение уже имело место, то информация в мозгу не распределяется по всей системе. Были использованы очень маленькие электроды. Обезьяны были обучены по-разному реагировать на различные геометрические стимулы. В отличие от экспериментов Джона стимулы освещались очень короткой вспышкой света В зрительной коре было обнаружено несколько различных типов волновых форм электрической активности. Один тип был получен, когда форма волны определялась относительно момента предъявления стимула. Этот тип вызванного ответа обнаружил четкую зависимость от характеристики стимула. Два других типа были получены, когда форма волны определялась по отношению к началу ответа животного. Один тип волновой формы зависел от того, получила обезьяна подкрепляющий пищевой шарик за правильный ответ или не получила, когда ее ответ был ошибочным Другой тип волновой формы возникал непосредственно перед двигательной реакцией животного. Эта форма волны коррелировала с определенным типом двигательного ответа (нажатием на правую или левую панель); она следовала за стимулом, но не зависела от него и от полученного подкрепления. Важным для нас является тот факт, что все эти типы характерных волновых форм появляются локально в зрительной коре. Один тип волновой формы записан от одних электродов, другой от других. Распределение этих волновых форм происходило не в соответствии с какой-либо структурой возбуждения. Однако со дня на день, от недели к неделе наблюдалось полное постоянство в записях, получаемых от каждого отдельного электрода. Какой бы процесс кодирования ни происходил, он стабилизировался по мере того, как проводилась запись.
Результаты этих экспериментов не согласуются с представлением о том, что образ, подобно фотографии, проецируется на поверхность коры. Они показывают, что каждая сенсорная система функционирует, имея большой резерв. Поскольку системы с разрушением и без него обнаруживают лишь незначительные различия в поведении, этот резерв, должно быть, распределен по всей системе – сохраняемая в ней информация, необходимая для осуществления различения, дублируется одновременно многими ее участками. Таким образом, вполне вероятно, что задержка в научении, возникающая при наличии эпилептогенных очагов, вызванных наложением на поверхность мозга алюминиевой пасты, обусловлена помехами, которые вносятся этими очагами в процесс дублирования хранения информации (рис. VII-5).
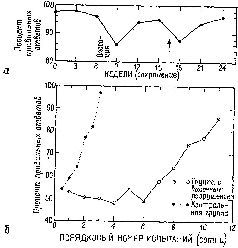
Рис. VII-5. а – сохранение зрительного различения, которое проверялось каждые 3 недели в экспериментах, аналогичных тем, которые были показаны на рис. VI-9 и VI-10. Вживление диска с пастой алюминия отмечено словом «операция»; появление первых эпилептоидиых разрядов указано стрелкой; 6 – кривые научения, полученные при выполнении задания на различение зрительных структур (каждая точка – медиана данных группы животных и каждой сотни испытаний). Белые кружки показывают выполнение задания ооезьянами с эпилептогенными разрушениями в коре височной доли, черные кружки – выполнение задания контрольной группой ооезьян (Stamm and Pribram, 1961).
Вопросы, поднятые этими наблюдениями, следует сопоставить с другим рядом проблем: каким образом предметы кажутся нам достаточно стабильными, так что мы можем узнавать их независимо от угла зрения и расстояния до предмета? Каким образом мы узнаем предмет, несмотря на то, что светом, идущим от предмета, возбуждается только часть сетчатки, а следовательно, и мозга? Константность восприятия предметов и величины существует у детей уже в возрасте нескольких недель. Таким образом, простое объяснение феномена константности восприятия в терминах научения ставится под сомнение. Какого же рода механизм мог бы обеспечить одновременно существующую гибкость восприятия и константность узнавания?
Рис. VII-6. Кортикальная клетка активируется только при освещении определенной области левого (противоположного) глаза. Эта область имеет размеры примерно 5 X 5° и расположена на 10 выше и левее центра. Нейрон лучше всего отвечает на предъявление черного горизонтального прямоугольника, который может быть без изменения эффекта параллельно смещен в пределах рецептивного поля на 1/3 X 6° (А- В). Наклонное положение прямоугольника делает его неэффективным (Г – Д). Отметка предъявления стимула показана горизонтальной линией над каждой записью нейронной активности (Hubel and Wiesel, 196).
Как факты восприятия структур при наличии скотомы, так и факты константности восприятия говорят о том, что существует, должно быть, какой-то эффективный нейрологический механизм пространственного распределения информации на входе мозга. Для объяснения фактов восприятия недостаточно изоморфизма, простого («точка в точку») соответствия между стимулом и картиной возбуждения в коре, обусловленной анатомией системы. Когда в результате удаления коры происходит выпадение 80% зрительного поля, узнавание осуществляется благодаря оставшейся части зрительного поля; когда в зрительной коре производится множество разрушений, то сохранившиеся части функционируют столь успешно, что при различении испыты-ваются лишь незначительные трудности; рассматриваем ли мы предмет той или другой частью сетчатки, под тем или иным углом зрения, мы всегда можем узнать его. Восприятие отличается от обычного фотографического изображения – оторвите 98 или даже 80% У фотографии и попытайтесь идентифицировать ее: в большинстве случаев это невозможно.
ПРОЦЕСС ВЫДЕЛЕНИЯ ПРИЗНАКОВ
Один из способов решить эту дилемму – это отказаться от представления о том, что нейронный механизм входа является в каком-либо отношении устройством, формирующим образ, предположить, что узнавание структур есть результат операции выделения признаков, имеющей место на самом входе и возникающей с помощью отдельных нейронов или их небольших групп, и что восприятие зрительных структур осуществляется посредством иерархической системы таких «детекторов признаков». В пользу такого представления говорит то, что внутри частей входных каналов происходит конвергенция сигналов.
Можно получить карты зрительных рецептивных полей для нейронов, находящихся на разных уровнях входной системы.
В коре мозга было обнаружено два типа таких нейронов. Примерно у половины из них эти карты имеют форму более или менее округлых полей, аналогичных рецептивным полям ганг-лиозных клеток сетчатки, но в отличие от последних не имеющие четко выраженной окружающей зоны (Jung, 1961; Spinelli, Pribram and Bridgeman, 1971). Другая половина кортикальных нейронов со значительно большей вероятностью активируется входными системами, имеющими не округлые, а удлиненные формы. Чтобы вызвать максимальный эффект, эти удлиненные формы должны быть определенным образом ориентированы в отношении глаза, на который они воздействуют; разные нейроны отвечают на различную длину и ориентацию линий, границ и даже углов (Hubel and Wiesel, 1962; рис. VII-6).
Известное объяснение этих наблюдений состоит в том, что нейроны, чувствительные к воздействию линии или границы, активируются только тогда, когда возбуждается большая часть или все линейное множество нейронов, чувствительных к воздействию таких пространственных признаков (рис. VII-7) и связанных с рецепторами линии и края. Высказано предположение, что благодаря дальнейшей конвергенции активность детекторов линии и края может комбинироваться и создавать клетки, реагирующие на более сложные структуры. Это предположение подтверждается тем, что найдены нейроны, особенно чувствительные к таким сложным структурам, как углы и короткие участки линий.
Таким образом, объяснение восприятия на основе механизма анализа признаков становится крайне соблазнительным. Электрофизиологический анализ рецептивных полей клеток внутри вход-пых каналов показывает, что выделение признака в какой-то степени происходит внутри этих каналов. Далее, есть основание предполагать, что выделение основных признаков, таких, как цвет (De Valois, 1960; De Valois and Jacobs, 1968), контур и направление, осуществляется врожденными, встроенными в организм детекторами. Этот тип механизма, осуществляющего детекцию основных признаков, дополняется механизмом, обеспечивающим выбор признака, который зависит от прошлого опыта организма. Возникает вопрос, изменяет ли прошлый опыт сами эти детекторы признаков или в результате этого опыта в деятельность вовлекаются дополнительные нейроны, вызывающие изменения в нейронной популяции, изучаемой экспериментатором. Однако очевидно, что в зрелом организме процесс выделения признаков не может быть ограничен работой встроенных в организм детекторов. Должен происходить также анализ признаков посредством нейронов памяти. Поэтому обратимся теперь к логике той архитектоники нейронов, которая присуща анализаторам признаков, чтобы определить, что можно и что нельзя объяснить с этих позиций в самом восприятии.
СТРУКТУРА АНАЛИЗАТОРОВ ПРИЗНАКОВ
Софистическое утверждение Г. Вернера (1971), основанное отчасти на его собственных обширных исследованиях и работах В. Маунткасла (1957), Д. Хьюбела и Т. Визела и отчасти на концепции Т-О-Т-Е, относится к процессу анализа признаков в соместетической чувствительности и может быть с небольшими изменениями отнесено также и к другим входным системам. В коре мозга он различает базальную структуру – колонку, каждый нейрон которой обладает рецептивным полем, образующим единицу представительства входа. Эти нейроны имеют, как правило, идентичные или по крайней мере сходные рецептивные поля и, таким образом, образуют одинаковый уровень представительства. Эти колонки в свою очередь образуют более сложные структуры благодаря нейронам с дирекционпой чувствительностью, которые выполняют функцию указателей для согласования деятельности колонок (рис. VII-7).
Рис. VII-7. Схематическое представление организации соматосенсорной коры в терминах рецептивных полей отдельных клрток. А – экспериментальные данные, Б – обобщение результатов, касающихся кортикальных колонок, полученных в нескольких микроэлрктродных проходках у одной, или более обезьян; В – представление об организации коры, основанное на совокупности всех данных, в терминах концепции Т – О – Т – Е, демонстрирующее проекционное отношение между организацией коры и поверхностью тела. Структура коры сравнима с кодированием информации при последовательном изложении инструкции в форме единиц Т – О – Т – Е, из которых составляются программы для компьютеров. Ср с рис. VII-8 (см. Werner, 1970).
Рис. VII-8 ОККАМ Спинелли. Объяснение на стр. 148, а – входные нейроны: е – выходные нейроны, т – тестирующие нейроны. Ср. с рис. VII-9 (Pribram, 19696).
Эти указатели в зависимости от того, на какой ответ они настроены, определяют тот или иной тип взаимоотношений в электрической активности колонок; если имеются нейроны-указатели, которые чувствительны более чем к одной ориентации сигнала, то группы колонок оказываются связанными таким образом, что формируют «кольцевые структуры». Вернер сравнивает такую структуру коры с последовательным изложением инструкций, из которых составляются программы для электронно-вычислительных машин. Каждый лист программы содержит пункты, которые указывают на переходы к другим листам. Таким образом, посредством такого простого устройства могут быть запрограммированы сложные взаимодействия, последовательность операций. В терминах концепции Т-О-Т-Е наличие у колонок коры рецептивных полей со статическими свойствами создает условия для осуществления фазы проб в работе сервомеханизма; свойства колонок, свидетельствующие об их чувствительности к направлениям, составляют основу операционной фазы и фазы выхода. Сшшелли (1970) фактически составил программу (названную «Оккам»), моделирующую работу анализатора признаков, построенного па оспове такой кортикальной структуры. Предъявляя структуры нервных импульсов или волновых форм, можно настроить эту программу таким образом, чтобы она реагировала при повторении волн определенной формы (рис. VII-8). Несколько упрощепно точка зрения Вернера и Спинелли на анализатор признаков заключается в следующем: колонка коры состоит из входных и операционных нейронов, интернейронов и тестовых клеток. Входной сигнал, доходящий до нейронной единицы колонки, которая имеет свое рецептивное поле, распределяется по интернейронам. Последние в свою очередь связаны с операционными нейронами. Интерпейроны способны к настройке, то есть они адаптируются, обнаруживают привыкание и обладают памятью. Таким образом, каждый интернейрон действует как сумматор в электронно-вычислительной машине, который усредняет динамические структуры, вызванные поступающими на вход сигналами. Только при повторении таких динамических структур возбуждения в результате суммации возникает устойчивая структура, неповторяющиеся динамические структуры просто повышают нулевую линию и среднюю величину выходного сигнала. Таким образом, операционный нейрон, чувствительный только кдинамическим структурам возбуждения, активируется лишь при повторении структуры входных сигналов. В целом процесс достигает своей завершенности благодаря посылке выходного сигнала от операционного нейрона к входному через тестирующий нейрон, который сличает структуру нейронной активности, вызванную входным сигналом, со структурой возбуждения, возникающей в операционных нейронах. Когда обе динамические структуры совпадают, тестирующая клетка продуцирует выходной сигнал, в противном случае при их несовпадении продолжается процесс настройки. Таким образом, каждая кортикальная колонка создает энгр:шму в соответствии со своей специфической чувствительностью к данной структуре нейронной активности, которая выступает в виде «описания» либо последовательности межспайковых интервалов, либо в волновой форме, которая являет собой огибающую узора спайковой нейронной активности.
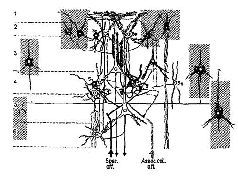
Рис. VII-9. Полусхематическая зарисовка наиболее важных типов нейронов неокор-текса и их связей. Обратите особое внимание на связи пирамидных клеток (выходных нейронов) со звездчатыми (тестирующими) интернейронами (Si, S 3 ), имеющими вход от специфических афферентов (Szentagothai, 1969).
Считается, что каждая колонка связана с другими через горизонтальные клетки и их базальные дендриты, которые ответственны за тормозные взаимодействия (рис. VII-9). Всякий раз, когда эти горизонтальные клетки активируются несимметрично как это происходит, когда они возбуждаются через входы с ди-рекционной чувствительностью, создаваемые при этом временные структуры отдельных колонок образуют единую структуру. Следовательно, эти обширные структуры, зависящие скорее от гиперполяризации, чем от проведения нервных импульсов, обусловлены микроструктурой соединений. -Они и создают временно существующие состояния нейронов. Но более подробно мы остановимся на этом в следующей главе.
Сейчас мы располагаем надежным доказательством того, что так называемые ассоциативные зоны коры осуществляют некоторый контроль над входными системами, во многих отношениях подобный тому, который имеет место при изменении кривизны хрусталика глаза. Такая функция контроля могла бы менять число (и, по-видимому, сложность) кортикальных колонок, которые могут участвовать в создании такой динамической структуры (гл. XVII).
Таким образом, можно считать, что входные системы представляют собой некий фильтр для входных сигналов, экран, непрерывно настраивающийся на входные воздействия. Одной из характерных особенностей этого фильтра является то, что он создает самоадаптирующуюся систему, параметры адаптации которой находятся под контролем его собственной прошлой истории и тех операций, которые были осуществлены в нем посредством других нейронных механизмов. Другой особенностью фильтра как системы, которая обусловлена взаимосвязями между логическими элементами, является его способность к конструированию Образа. Но здесь мы забегаем вперед. Рассмотрим сначала проблему, почему необходим процесс формирования Образа вообще и в чем состоит ограниченность концепции анализа признаков при объяснении восприятия.
ГРАНИЦЫ ПРИМЕНЕНИЯ ГИПОТЕЗЫ АНАЛИЗА ПРИЗНАКОВ
Почему же неправильно рассматривать перцептивные процессы как безобразный анализ признаков? По трем причинам. Во-первых, анализируемые признаки не являются какими-то особыми, отличными от других. Во-вторых, богатство феномена восприятия не объясняется теми детекторами признаков, которые открыты. И в-третьих, изменения, которые имеют место в системах входа в ходе развития организма, позволяют различать эффекты, возникающие в детекторах признаков, изученные с помощью микроэлектродов, и эффекты, продуцируемые в системе, анализирующей эти признаки, что было изучено в поведенческих опытах с различением сигналов.
Рассмотрим прежде всего проблему различимости признаков. И. Рок (1970) со всей проницательностью сформулировал эту проблему следующим образом:
«… Те, кто утверждал бы, что ответ нейронных детекторов может объяснить, почему вещи выглядят такими, какими они являются на самом деле, возможно, не знают тех фактов, касающихся восприятия, на которые гештальтисгы обращали особое внимание. Например, известно, что воспринимаемая нами величина, не просто зависит от величины образа на сетчатке, воспринимаемая форма не является непосредственной функцией формы соответствующего изображения, а воспринимаемая скорость – непосредственной функцией скорости перемещения изображения на сетчатке и т. д. Напротив, несмотря на значительные изменения находящегося в отдалении от нас стимула, возникающие при изменении позиции наблюдателя, предметный мир сохраняет удивительную константность. Эти факты нельзя отрицать, и их нельзя оставить без внимания на том основании, что вещи в этих изменяющихся условиях не выглядят константными, а мы знаем, что они константны. Против этой формулировки имеются достаточно веские возражения.
Рассмотрим детально некоторые факты, касающиеся восприятия движения. Начнем с того, что существует константность положения, а именно-известно, что, когда мы перемещаемся или двигаем глазами, перемещается и образ всей картины на сетчатке, но сама картина не кажется движущейся. Случается также обратное, а именно неподвижный образ на сетчатке вызывает восприятие движения при движении наших глаз или головы, как это наблюдается при слежении глазами за движущимся объектом или при рассматривании последовательного образа во время движения тлаз. Известно, далее, стробоскопическое движение, когда сменяющиесянеподвижные изображения вызывают ощущение движения. Известен также феномениндуцированного движения предмета, изображение которого на сетчатке неподвижно. Этот феномен возникает вследствие перемещения окружающих объектов. Так, например, Луна кажется движущейся на фоне перемещающихся облаков.
Можно принять гипотезу, что действительной сенсорной основой восприятия движения является стимуляция детекторов движения, но тогда необходимо предположить, что значимость такой стимуляции определяется другой информацией. Например, можно сказать, что движение вообще не воспринимается, если данным о смещении изображения на сетчатке противостоит информация о том, что оно возникает в результате собственного произвольного движения организма, как это предположил фон Хольст, -а до него Гельмгольц. Или можно считать, что в случаях индуцированного движения детекторы предоставляют решающую информацию о том, что что-то движется, но по неизвестным еще причинам движение приписывается неподвижному объекту.
Откровенно говоря, я не считаю это достаточно убедительным. Это не объясняет, почему я вижу свой последовательный образ движущимся, когда я двигаю глазами даже в совсем темной комнате, где не видно неподвижных предметов, изображения которых могли бы перемещаться на сетчатке. В случае же стробоскопического движения, если наши глаза удерживаются в стационарном положении, то источником стимуляции детекторов движения мог бы быть только возникающий в этих условиях нистагм глаза. Но это не могло бы объяснить движения, которое возникает между двумя раздельно освещаемыми объектами. Эта гипотеза не может объяснить и индуцированного движения, потому что предмет, который вызывает ощущение движения, может двигаться так медленно, что его движение будет ниже порога, в то время как только оно и воспринимается. Далее, очевидно, что в данном случае основой восприятия движения является изменение локализации предмета, который кажется движущимся относительно местонахождения предмета, который вызывает это движение, а не стимуляция детекторов перемещения изображения.
Рассмотрим теперь следующую проблему – проблему восприятия ориентации предмета. Является ли основой воспринимаемой ориентации линии ориентация ее изображения на сетчатке? Очевидно, нет, поскольку при повороте головы вертикальные и горизонтальные линии на стене продолжают оставаться для нас вертикальными и горизонтальными. Здесь мы «нова сталкиваемся с константностью, в данном случае с константностью восприятия направления, несмотря на изменения в ориентации изображений на сетчатке. Это происходит в значительной степени даже тогда, когда мы рассматриваем в темноте одну светящуюся линию, наклонив или откинув голову в другую сторону. Следовательно, обоснованно заключение, что, прежде чем сделать вывод о значении для восприятия ориентации данного сетчаточного образа, необходимо принять во внимание информацию о собственной позиции наблюдателя.
Далее, как бы мы поняли открытие, сделанное Хьюбелом и Визелом, о существовании в зрительной коре детекторов ориентации? Очевидно, что они не могут быть знаками ориентации объектов в окружающей среде. Можно согласиться, что они являются знаками, указывающими на ориентацию объектов относительно организма, другими словами, детекторами эгоцентрической ориентации, и что они создают условия, необходимые для того, чтобы организм мог определить ориентацию объектов в окружающей среде, когда он дополнительно получает информацию и об ориентации самого ' наблюдателя. Например, можно сказать, что, когда изображение линии на сетчатке будет иметь вертикальное положение, оно всегда будет казаться эгоцентрически вертикальным, то есть параллельным продольной оси головы, и линия будет восприниматься вертикальной в поле зрения наблюдателя, если он занимает вертикальное положение, и горизонтальной, если наблюдатель повернут на 90°.
Хотя такое объяснение кажется мне правдоподобным, оно вызывает серьезные трудности из-за существования некоторых дополнительных фактов. Если наблюдатель сохраняет вертикальное положение, то вертикальная линия, воспринимаемая им на фоне изображения комнаты, при наклоне последнего будет выглядеть наклоненной в направлении, противоположном наклону изображения комнаты. Таким образом, в данном случае вертикальное изображение на сетчатке не будет более соответствовать вертикально расположенному предмету, несмотря на то, что наблюдатель находится в вертикальном положении. Более того, линия не будет уже казаться эгоцентрически вертикальной линией. К этим фактам можно было бы добавить данные о том, что наблюдатели будут адаптироваться к повернутым с помощью призматических стекол изображениям, так что вертикальное изображение на сетчатке не будет больше ни знаком вертикального объекта, когда наблюдатель находится в вертикальном положении, ни фактически знаком того, что объект ориентирован в вертикальном направлении относительно наблюдателя.
Имеют ли какое-либо отношение детекторы ориентации в коре к восприятию формы? Можно предположить, что восприятие формы сводится к восприятию ориентации линий, из которых строится форма. Таким образом, предмет воспринимается как квадрат тогда, когда одновременно стимулируются два детектора вертикальных и два детектора горизонтальных линий. Хорошо известно, что фигуры выглядят весьма по-разному, когда меняется их ориентация. На первый взгляд сам этот факт может казаться доказательством того, что детекторам ориентации принадлежит существенная роль в восприятии формы, потому что в этом процессе принимают; участие совершенно различные волокна. Можно сказать, что это является основой для понимания явления изменения внешнего вида предмета. Однако в действительности, вообще говоря, объяснением изменения внешнего вида предмета служит неизменение ориентации изображения на сетчатке. В проведенных мною и моими сотрудниками экспериментах неоднократно-было показано, что, если положение фигуры в отношении ее окружения не меняется, но она рассматривается наблюдателем в наклонной позе, фигура невыглядит измененной, или, выражаясь более точно, распознавание не ухудшается. Могло ли бы это быть, если за это явление были бы ответственны совершенно различные детекторы? Напротив, если фигура наклонена так же, как и наблюдатель, и ориентация изображения на сетчатке, таким образом, совсем не изменилась, фигура тем не менее выглядит совершенно иной и иоэтому часто не узнается. Можно провести простой опыт: создайте последовательный образ квадрата, держа голову в вертикальном положении, а затем, закрыв глаза и повернув голову на 45°, посмотрите на него, он будет подобен ромбу. Могло ли бы это быть, если за это явленяе были бы ответственны одни и те же детекторы?
Однако эти факты приобретают смысл, если считать, что решающим-для ориентации является восприятие определенной части фигуры как верх, а другой – как низ и т. д. Если мы будем правильно оценивать одну и ту же часть фигуры в качестве верха, как мы это обычно делаем, приняв наклонное положение, то мы увидим, что форма предмета не меняется. Однако, если мы за верх фигуры принимаем другую ее часть, как это бывает при наклоне фигуры относительно окружения, то мы увидим, что ре форма очень изменилась. Этот процесс, по-видимому, не имеет ничего-общего со специфическими детекторами ориентации.
Далее, роль этих детекторов, возможно, состоит в том, что они служат для различения ориентации одного образа по отношению к другому. В прошлом мы ошибочно считали, что такое различение осуществляется благодаря различной пространственной ориентации проекционных кортикальных «образов». Иными словами, нейронные детекторы, вероятно, являются механизмами, посредством которых воспринимается определенная информация о стимуле, в данном случае информация о том, что сохраняется или изменяется пространственная ориентация. Конечно, такая информация необходима для восприятия в качестве исходной, но ответ волокон детекторов никоим образом нельзя рассматривать как нейронные корреляты единиц восприятия» (1970, р. 2-5).
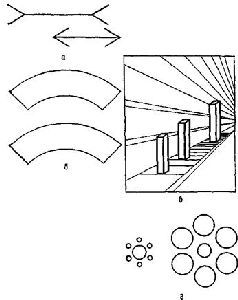
Рис. VII-10. Некоторое нарушение константности величины (иллюзии) при тождественности сетчаточных образов. а – классическая иллюзия Мюллера – Лайера, когда две одинаковые линии кажутся различными из-за стрелок на их концах, которые направлены в противоположные стороны; б – нижняя фигура кажется больше верхней, хотя обе имеют один и тот же размер; в – три колонки имеют одинаковую высоту; г – два центральных круга равны. (Kretch and Crutchfield, 1962).
В остальной части статьи Рока анализируется еще ряд интересных экспериментов, которые трудно объяснить с позиции детекторов признаков (см. также рис. VII-10, VII-11).

Рис. VII-11. Два примера, иллюстрирующие влияние окружающего фона на восприятие формы, а – фигура, лежащая на штриховом фоне, имеет форму точного круга; б -дуги, находящиеся внутри двух параллельных кривых, идентичны. Эти иллюзии трудно объяснить с позиции механизма детекции признаков (Kretch and Crutchfield, 1962).
Второй недостаток точки зрения, рассматривающей восприятие как безобразный процессвыделения признаков, состоит в том, что по сравнению с богатством явлений сознания процесс распознавания структуры, осуществляемый методом классификации, довольно беден деталями, которыми он оперирует. Механизм распознавания зрительной структуры, который осуществляется путем избирательного выделения признаков, может приводить к ошибкам. Каждый из нас ждал кого-нибудь, кто опаздывал на свидание, назначенное в многолюдном месте, и ошибочно принимал посторонних за ожидаемое лицо, так как они имели сходные с ним черты. Мы узнаем А, Л и а как букву А; мы хорошо знаем о различиях почерков. Восприятие, по-видимому, обеспечивает в одно и то же время не только богатство нашего опыта и «сильное предпочтение сохранять постоянную организацию», но также и тенденцию к «максимальному упрощению», как показали опыты гештальтпсихологов. Имеются данные, свидетельствующие о том, что богатство восприятия связано со сложностью конфигураций окружающей среды, которые существуют независимо от организма и возбуждают его рецепторные поверхности (см. Gibson, 1966), последние приводят в действие некоторый механизм мозга с соответствующей емкостью, позволяющей вместить это богатство. Данные, представленные Роком и другими, показывают, что только механизм выделения признаков обусловливает тенденцию к упрощению, создает схему отношений. Однако даже здесь начинает играть роль другой механизм – более сложный «познавательный» процесс.
Третий недостаток объяснения фактов восприятия с позиции выделения признаков выступает в экспериментах, в которых новорожденные в течение различных периодов времени подвергались сенсорной депривации. Как уже говорилось в гл. VI и VIII, младенцы в возрасте нескольких недель обнаруживают способность к восприятию константности величины и формы. У животных способность к различению развивается также рано; при изучении мозга детенышей животных были обнаружены детекторы признаков, которые уже достигли своего полного развития. Однако при ограничении потока сенсорных воздействий эта простая корреляция между поведением и функциями мозга нарушается. Так, когда кошки и крысы содержатся в полной темноте, их способность различать яркость и контур остается неизмененной, но способность различать более сложные зрительные структуры значительно ухудшается. Хотя ориентация рецептивных полей кортикальных нейронов у таких животных еще различима, она имеет менее острую настройку, и утомление клеток развивается быстрее, чем у животных, содержащихся в нормальных условиях. Но когда сенсорная депривация ограничена депри-вацией одного глаза в течение определенного времени, как зрительная ориентация, так и способность к различению оказываются резко нарушенными, если животное пользуется только одним этим глазом. Это связано с тем, что стимуляция глаза, подвергнутого депривации, может привести в активное состояние лишь небольшое число нейронов, которые вряд ли могут сколько-нибудь избирательно отвечать на определенным образом ориентированную линию или движение. Эта реакция возникает, даже несмотря на то, что в коре присутствует обычный для нее комплект клеток с их нормальной реактивностью, как это было обнаружено при стимуляции не подвергавшегося депривации глаза (см. рис. VII-12). Таким образом, когда имеет место нарушение в привычном способе возбуждения, происходит какое-то подавление реактивности и это подавление переходит в нарушение функции исключительно в результате неупотребления нервных структур (как у животного, подвергшегося полной сенсорной депривации). Если монокулярное подавление реактивности коры объясняется некоторым изменением в связях, то ответственным за этот эффект подавления должен быть какой-то процесс, отличный от простого выделения признаков.
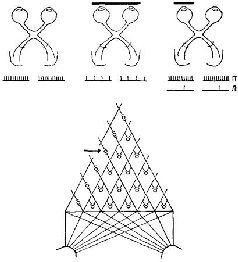
Рис. VII-12. Схема, иллюстрирующая результаты экспериментов со зрительной депривацией одного или двух глаз у новорожденных. Следует отметить, что односторонняя депривация ведет к значительно большему подавлению активности нейронов зрительной коры, чем двусторонняя. Это обстоятельство создает известные трудности (см. текст) при объяснении некоторых экспериментальных фактов с позиции простого механизма детекции признаков, поскольку нейроны имеют связи с обоими глазами (нижняя схема) и продолжают нормально отвечать на стимуляцию глаза, не подвергшегося сенсорной депривации.
Л. Ганц в недавнем обзоре (1971) этих и других сходных данных доказал это. Основываясь на результатах экспериментов с полной депривацией животных, он различает два типа заданий. Ориентация по яркости и контуру требует только, чтобы организм отвечал на языке реакций, свойственных большинству его детекторов признаков. Различение более сложных структур, например двух треугольников, один из которых перевернут, не может осуществляться на этой основе. Мы нуждаемся в каком-то дополнительном, более гибком мозговом механизме, который Ганц называет «избирательным вниманием». Этот механизм активно подавляет доминирующую реакцию детекторов признаков, реакцию, которая, в сущности, не имеет отношения к адекватному решению задачи на различение.
Следовательно, мы снова сталкиваемся с необходимостью иметь дополнительный механизм, помимо выделения признаков. Поэтому мы снова приходим к мысли о той, что, возможно, системы входа организованы таким образом, что нейронные сигналы координируются с каким-то психологическим процессом формирования образов. Проблема состоит в том, чтобы определить нейро-логическую природу этой организации. На основе рассмотренных в начале этой главы данных становится очевидно, что эта организация не может быть просто мозаикой точек, которые генерируются в рецепторах и без изменения проводятся до коры, где они формируют образ, подобно фотографии. Столь же очевидно, что следует принять во внимание наличие нейронов, чувствительных к линиям, и других детекторов признаков, которые создают в мозгу нейронную основу для их анализа.
РЕЗЮМЕ
Почти полное удаление мозговой ткани независимо от ео л№-кализации в пределах систем входа оказывает лишь незначительное влияние на процесс распознавания. Следовательно, логика работы нейронов, свойственная любому конкретному восприятию, распространяется, по-видимому, на всю систему. Были получены также прямые данные, касающиеся распределения памяти в мозгу. Логику этой памяти создают как встроенные детекторы элементарных признаков (таких, как линии и углы), так и анализаторы для более сложных преобразований особых признаков (таких, как волновые формы), которые перестраиваются в ходе жизненного опыта организма.
Глава VIII
ГОЛОГРАММЫ
ГИПОТЕЗА
Вернемся на время к нашей общей модели. Модель предполагает два основных процесса: пространственно организованные-состояния и операции, выполняемые на фоне этих состояний с помощью импульсной передачи между нейронами. В первой части книги было описано, каким образом эти основные свойства нейронных групп могут комбинироваться в логические операции, усиливающие аналитические и контрольные (серво) функции" нервной системы. Учитывая их значение, а также то, что сейчаа они игнорируются в нейропсихологической и нейрофизиологической литературе, я подчеркнул, что построение имеющих определенную структуру топологических, то есть пространственных, представительств в нервной системе является одной из форм, которые могут принимать состояния мозга. Я предположил, что-взаимодействие динамических структур возбуждения, падающих на рецепторные поверхности, после их передачи по параллельным путям кодируется благодаря горизонтальным связям в активность медленных потенциалов групп нейронов и образует временные микроструктуры, рисунки которых зависят скорее от функциональной организации нейронных соединений, чем от нейронов, как таковых.
В гл. VII были подробно рассмотрены факты, свидетельствующие о существовании детекции признаков и механизма анализа, а также приведены аргументы в пользу необходимости особой нейронной организации, помимо той, которая представлена анализаторами признаков. В гл. VIII формулируется гипотеза о том, что такой организацией может быть механизм микроструктуры медленных потенциалов соединительных аппаратов. Эта гипотеза основывается на предположении, что нейронное отображение входных воздействий не является фотографическим и создается не только посредством имеющейся системы фильтров, выделяющих признаки, но и с помощью особого класса преобразований, которые обладают значительным формальным сходством с процессом отражения оптического образа, открытым математиками и инженерами. Этот оптический процесс, названный голографией, основан на использовании явления интерференции структур. Он обладает множеством удивительных свойств, из которых первостепенное значение имеет его способность к распределению и сохранению большого количества информации. Именно эти свойства дают возможность разрешить противоречие между потребностью в функциональной лабильности, быстром темпе изменений и уже рассмотренными анатомическими особенностями в организации нервных систем приемл информации.
Прежде чем точно сформулировать проблему, полезно посвятить несколько параграфов разъяснению нашего общего подхода. Обработка оптической информации с помощью голографии описывается математически в понятиях механической волны. В физической оптике для описания распространения света в экспериментальных условиях могут быть использованы как квантовые, так и волновые уравнения. Физика не интересует то, является ли свет квантом или волной, распространяется ли он в виде частиц или электромагнитных волн, или в форме того и другого. Он заботится о том, чтобы как можно полнее количественно описать результаты своих наблюдений, и соответственно этому выбирает инструменты для своего описания. Некоторые наблюдения могут быть хорошо описаны как статистическая вероятность квантовых событий, другие легче описываются с помощью волновой теории.
Особенно трудно привести в соответствие с квантовым подходом одно наблюдение. Когда свет очень низкой интенсивности проходит через решетку, то есть когда предполагается, что через каждую отдельную щель пройдет самое большее несколько квантов света, уравнение, описывающее световой поток по другую сторону решетки, для объяснения наблюдаемых эффектов должно учитывать некоторое взаимодействие, которое, по-видимому, имеет место между световыми потоками, проходящими через соседние щели (рис. VIII-1).
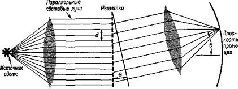
Рис. VIII-1. Дифракционная решетка (Murray and Cobb, 1970.
Трудно, хотя и не невозможно, объяснить это взаимодействие с точки зрения квантовой теории: каким образом квант света может влиять на соседние, отделенные от него решеткой? Кажется, будто каждый квант оказывает силовое (возникает искушение сказать «магнетическое») действие на соседний. Этот феномен взаимодействия был широко изучен и получил названиесуперпозиции. Математические уравнения, удовлетворительно объясняющие явление суперпозиции, представляют собой систему линейных уравнений, названных интегралами свертки, которые обычно используются для описания взаимодействия волновых процессов. Описание одной волновой формы свертывается вместе с описанием другой. При объяснении суперпозиции не возникает серьезных теоретических проблем, если считать, что ровный фронт волн света, встречаясь с решеткой, разбивается ею на небольшие, взаимодействующие между собой «вихри». Можно считать, что такие вихри и вызывают эффект интерференции и объясняют суперпозицию – «взаимодействие по соседству».
Голографическая гипотеза функционирования мозга в процессе восприятия принимает форму суперпозиции. Фактически выбор способа объяснения того, как изменяются электрические потенциалы мозга – в статистических терминах или в волновых,- зависит от наблюдений, которые должны быть описаны. Следует выбрать тот способ, который может дать наиболее полное количественное описание наблюдений и в то же время быть приемлемым для концепции. В гл. III описано возникновение латерального, или периферического, торможения в рецепторах и на разных уровнях центральной нервной системы. Латеральное торможение обусловливает эффект взаимодействия между процессами, протекающими в соседних нейронах. Если это пространственное взаимодействие адекватно описывается интегралами свертки, которые характеризуют физическое явление суперпозиции, то мы можем рассматривать взаимодействие в нервной системе на основе интерференционных эффектов. Правомочность такого выбора подтверждается теми фактами, что по крайней мере в сетчатке взаимодействие между соседними горизонтальными клетками обусловлено исключительно тормозными взаимодействиями посредством медленных потенциалов гиперполяризации, а не деполяризации, которая приводит к возникновению нервных импульсов.
Как и в гл. III, рецепторные явления служат миниатюрными моделями на этот раз «нейронного голографического» процесса.
Мы уже отмечали, что возбуждение одной единицы зрительного нерва оказывает влияние на частоту разрядов соседних единиц. Мы отметили также, что рецептивное поле отдельной единицы образуется в результате такого пространственного взаимодействия между соседними элементами. В зрительном нерве эти рецептивные поля обычно состоят из более или менее округлого центрального пятна, которое реагирует либо возрастанием (on – Центральный нейрон), либо снижением (off – центральный нейрон) частоты своих спонтанных разрядов, и из окружающей зоны, которая характеризуется активностью, противоположной по знаку активности центра. Глава VII была посвящена проблеме: .какой тип кода, какой тип организации создается в результате действия многих таких единиц? Настоящая глава посвящена другому кругу проблем, а именно: что кодируется в одном рецептивном поле, то есть какие черты оптического изображения определяют форму рецептивных полей?
Ответ на этот вопрос следует из проделанного Родие-ком (1965) детального математического анализа количественных отношений, существующих между специфическим образом движущимися зрительными стимулами (вспышками) и соответствующими им формами рецептивного поля. Кривые полученных ответов (каждая из которых представляет собой вертикальный срез через трехмерное рецептивное поле, рис. VIII-2) имеют особую форму. При одновременном предъявлении двух или более стимулов кривая ответов достаточно хорошо соответствует кривой, которая возникает в результате суперпозиции кривых ответов на раздельные предъявления этих стимулов. Таким образом, Родиек делает зывод, что, исходя «из структур ответа на небольшое световое пятно, возникающих при засвечивании разных участков рецептивного поля клетки, можно получить структуру ответа этой клетки на фигуру любой формы, движущуюся в любом направлении, с любой скоростью, через любую часть рецептивного поля». Это открытие позволило воспроизвести рецептивное поле клетки, используя только движущиеся пятна света, а также сделало возможным с помощью компьютера произвести запись, показанную на рис. VIII-7. Таким образом, форма рецептивного поля оказывается сверткой первой производной от формы зрительного стимула.
Как уже отмечалось, интегралы свертки, устанавливающие соотношения между событиями, происходящими в соседних областях (пространственная суперпозиция), описывают основной го-лографический процесс в системах, перерабатывающих оптическую информацию (Gabor, 1949, 1951). Эффекты интерференции описываются также уравнениями преобразований Фурье, Френеля и Лапласа. Определение же того, какое преобразование больше подходит для описания действия системы в данных условиях, является эмпирической проблемой.

Рис VIII-2 Карта рецептивного зрительного поля одиночной ганглиозной клетки сетчатки, построенная на основе записей активности ее аксона при точечном световом раздражении различных участков зрительного поля Карта представляет собой семейство контурных линий, так как ганглиозная клетка интегрирует ответы соседних нейронов, с которыми она связана, см. рис Ш-4 Высота контурной линии в любой ее точке дает представление о числе нейронов, которые одновременно активируются при предъявлении точечного раздражения в данном пункте рецептивного поля Максимум нейронных разрядов возникает, когда расположение светового пятна соответствует позиции центрального пика. В математическом смысле каждая контурная линия представляет собой «интеграл свертки» первой производной от формы светового раздражителя Взаимодействие многих таких интегрэлов свертки может вызвать в зрительной системе и в любом друюм месте мозга интерференцию динамических структур, подобную топографической. Сохранение таких структур могло бы быть основой для системы распределенной памяти (Rodieck, cm Pribram, 1969).
В зрительной системе – и имеются достаточные основания благодаря работам Г. фон Бекеши (1960, 1967, рис. VIII-3), чтобы предполагать, что также и в слуховой, соматической и вкусовой системах, – эти виды преобразований с большой точностью описывают взаимодействие между структурами энергетических воздействий, возбуждающими рецепторные поверхности, жспонтанными изменениями потенциалов в рецепторных единицах. В системах, обрабатывающих оптическую информацию, такие преобразования обычно имеют отношение к анализу формы волны (то есть эффектов интерференции), поэтому нет особых причин полностью игнорировать волновой подход и при объяснении эффекта суперпозиции, возникающего в нейронных системах. Преимущество такого подхода состоит в том, что физический процесс, основывающийся на эффектах интерференции, обладает многими свойствами нервного процесса восприятия и, таким образом, облегчает понимание мозгового механизма формирования образов. Рассмотрим теперь физическую голограмму.
Рис. VIII-3. Схема эксперимента, показывающего существование полос Маха для кожной чувствительности В эксперименте использовалось вибрационное точечное раздражение, обозначенное на схеме как стимул. Ощущение, вызываемое вибрационным раздражением, приложенным к предплечью, обозначено на схеме как интенсивность ощущения. Сходный, но несколько ослабленный эффект можно получить в результате прямого давления (с различной интенсивностью) невибрирующего стимула (Bekesy, 1967).
ФИЗИЧЕСКАЯ ГОЛОГРАММА
Большинству из нас известно, каким образом возникает изображение в обычных оптических системах. Фотоаппарат регистрирует на фотопленке, помещенной в плоскости изображения, копию пространственного распределения интенсивности света, отраженного от предметов, попавших в поле объектива. Каждая точка на фотопленке сохраняет информацию, полученную от соответствующей точки зрительного поля, и, таким образом, зарегистрированная картина выглядит подобно картине зрительного поля. Недавно-были проведены исследования свойств записей, сделанных на фотопленке, которая находится в оптической системе где-то передплоскостью изображения (то есть в фокальной плоскости). При соответствующей экспозиции такой фотопленки с помощью источника когерентного света она становится оптическим фильтром, в котором информация о каждой точке зрительного поля распределяется и хранится по всему фильтру (рис. VIII-4).
Рис VIII-4. Схематическое представление метода, используемого для создания оптического топографического фильтра (van Heerden, 1968).
Такие фильтры обладают многими удивительными свойствами. Как мы знаем, когда фотопленка внутри камеры находится не точно в плоскости оптического изображения, образ становится расплывчатым, его границы менее четки, контраст менее выражен. В оптическом фильтре информация распределяется таким образом, что зафиксированное на пленке изображение вообще не является подобием зрительного изображения. Оптический фильтр представляет собой запись волновых узоров, излучаемых объектом или отражаемых от него. «Такую запись можно рассматривать как «замороженный» волновой узор, который остается «замороженным» до тех пор, пока не приходит время восстановить процесс, после чего волны «считываются» с регистрирующего посредника» (Leith and Upatniecks, 1965). Так, когда оптический фильтр освещается источником когерентного света, он восстанавливает волновые фронты света, которые имели место при экспозиции объекта. В результате при рассматривании фильтра становится видно действительное изображение зрительного поля. Этот зрительный образ, по-видимому, точно совпадает с картиной, имевшей место при экспозиции, он является ее полным трехмерным изображением (рис. VIII-5). По существу, вся информация, описывающая зрительное поле, из которой может быть восстановлено изображение, содержится в этом фильтре.
Рис. VIII-5. Обычная фотография объекта и фотография реального изображения того же самого объекта, сделанная голографическим методом (справа), обе с помощью лазерного источника света. Обратите внимание,, что при современной технике голографическое изображение достигает высокого уровня совершенства (Stroke, 1969).
«Когда наблюдатель при рассмотрении фильтра меняет свое положение, перспектива изображения тоже меняется, как если бы наблюдатель рассматривал реальную сцену. Обнаружены эффекты параллакса между близко и далеко расположенными объектами: если один предмет находится на переднем плане и заслоняет какой-то другой предмет, то наблюдатель может, двигая головой, заглянуть за заслоняющий предмет и: таким образом увидеть то, что за ним скрывается… Короче говоря, восстановленная голограмма обладает всеми визуальными свойствами реальной сцены, и мы не знаем ни одного зрительного теста, который можно было бы использовать, чтобы отличить их друг от друга (Leith and Upatnieeks,. 1965, p. 30).
Еще до того, как было продемонстрировано практически использование оптических фильтров в восстановлении изображений, Д. Габор (1949, 1951) математически описал другой способ получения изображения с помощью фотографии. Габор стремился увеличить разрешающую способность электронной микрофотографии. Он предположил возможность интерференции когерентной фоновой волны с волнами, которые преломляются тканью. (Этой цели может служить также отражение волн от темного объекта.) Возникающий в результате интерференции узор сохранял бы и амплитудную и фазовую (взаимодействие по соседству) информацию, которая затем на втором этапе могла бы быть использована для восстановления при освещении источником когерентного света изображения естественной ткани. Габор назвал свою технику «голографией», а сфотографированную картину- «голограммой», потому что она содержала всю информацию, необходимую для восстановления целостного образа.

P и с. VIII-6. А – схема, иллюстрирующая метод получения голограмм по Габору; Б – фотография аппаратуры (Stroke, 19G9).
Голограммы Габора можно построить двумя способами. Волна разделяется с помощью прибора, расщепляющего лучи (то есть полупрозрачного посеребренного зеркала) таким образом, что-одна часть служит в качестве опорной волпы, а другая отражается от объекта, который должен быть сфотографирован (рис. VIII-6 и VIII-7). Затем можно использовать один опорный луч для восстановления изображения. Либо каждая часть расщепленного-луча может быть отражена от различных объектов. Когда при восстановлении изображения один из объектов используется в качестве опорного, другой появляется как образ его «призрака» (фантомное изображение). В этом случае голограмму можно использовать как механизм ассоциативного хранения информации. Постепенно становится очевидным формальное сходство междуголограммами преломления и отражения Габора и различными типами оптических фильтров. Их сходство состоит в том, что-закодированная в каждой из них информация является линейным преобразованием узора световых волн не только по интенсивности света, как в обычной фотографии, но и по их пространственному взаимодействию (пространственной фазе). Наиболее-изученными являются голограммы, у которых эти фазовые отношения могут быть математически выражены посредством преобразований Фурье. Эти голограммы представляют собой особую форму интеграла свертки, который обладает тем свойством, что одно и то же уравнение осуществляет прямую и обратную операции свертки. Таким образом, процесс, выражаемый пространственным преобразованием Фурье, может осуществлять кодирование и последующее декодирование просто в результате возвращения к первоначальному состоянию на некой второй стадии.
Все голограммы в целом (рис. VIII-8) обладают рядом интересных свойств, которые делают их потенциально важными для понимания функционирования мозга. Первое – ив данный момент наиболее важное для нас – состоит в том, что информация о каждой точке объекта распределяется по всей голограмме и тем самым делает регистрацию ее устойчивой к разрушениям. Любая малая часть голограммы содержит информацию о всем объекте-оригинале и, следовательно, может восстановить ее. Когда куски голограммы становятся еще меньше, несколько уменьшается ее-разрешающая способность. Когда же для восстановления изображения используются большие части голограммы, уменьшается глубина поля восстановленной картины, то есть сужается зона фокуса. Таким образом, для конкретных целей может быть установлена оптимальная величина голограммы (Leith and Upatniecks, 1965).
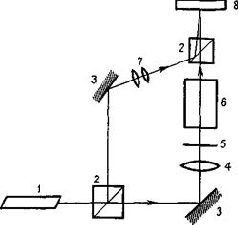
Рис. VIII-7. Топографический микроскоп, дающий возможность видеть изображение предмета на различной глубине. 1 – лазер; 2 – призма, разделяющая лучи; 3 – зеркало; 4 – конденсорная линза; 5 – объект; 6 – микроскоп; 7 – линзы микроскопа; 8 – голограмма (Stroke, 1969).
Голограмма обладает фантастической способностью к эффективному (то есть восстановимому) хранению информации. Информация, включенная в соответствующую систему восстановления, может быть непосредственно размещена и точно восстановлена. Плотность хранения информации ограничена только длиной волны когерентного света (чем короче длина волны, тем больше емкость памяти) и зернистостью используемой фотопленки. Более того, одновременно может сохраняться множество различных узоров, особенно когда голограмма наносится на твердый предмет. Каждое изображение хранится по всему твердому телу, хотя каждое из них воспроизводится независимо от других. Как описывают Лейт и Упатниекс (1965),
«…на одну пластинку может быть последовательно наложено несколько изображений, а затем каждое из них может быть восстановлено в отдельности, не испытывая помех со стороны других. Это достигается благодаря использованию различных пространственно-частотных носителей информации для каждого изображения … Решетки, несущие информацию, могут быть различной частоты … и возникает еще одна степень свободы – угловая» (р. 31).
Обычно в одном кубическом сантиметре голограммы хранится несколько десятков миллиардов бит (единиц измерения) информации. Как заметил П. ван Хирден, если бы мы на протяжении своей жизни каждую секунду запоминали один -бит информации, то для выполнения этой задачи мозг должен был бы каждую секунду совершать около 3 X 10 10 элементарных двоичных операций (нервных импульсов). «Если бы так обстояло дело, то это (прежде всего) было бы невозможно… Однако, столкнувшись с таким парадоксом, постепенно начинаешь понимать… что-оптическое хранение информации и ее обработка могут предоставить в наше распоряжение способ осуществить эту «невозможную» операцию… » (1968, р. 28-29).

Рис. VIII-8. Отпечаток реальной голограммы. Нельзя видеть самого изображения, хотя, несмотря на различие рисунка в разных частях голограммы, по существу, он может быть восстановлен от каждой ее части.
В заключение следует сказать, что оптические системы неединственные системы, которые могут быть объектом топографического процесса. В настоящее время математически описаны и составлены программы для компьютера, «моделирующие» процесс хранения оптической информации. В одной такой программе-интенсивность входного воздействия кодируется размером диска; пространственные фазовые отношения представлены угловой регулировкой щели внутри этого диска (рис. VIII-9).
Рис. VIII-9. Пример неоптической голограммы. Схема показывает три способа конструирования ячейки, с помощью которой могут восстанавливаться голограммы, и пример голограммы Фраунхо-фера (метод А). Ширина и высота щели, как и величина ячейки, – регулируемые параметры (Brown and Lohmann, 1966; см Pribram, 19696).
Таким образом, голограммы не зависят от физического присутствия «волн», хотя они наиболее полно описываются уравнениями волновой механики. Эта независимость голографии от-наличия физической волны является важным соображением при подходе к проблеме нейронного голографического процесса. Существует достаточно серьезное сомнение в том, что «волны мозга» в том виде, в каком они в настоящее время регистрируются, могут представлять собой субстрат какого-либо узора интерференции, адекватного для переработки информации, хотя они могут указывать на то, что такой процесс имеет место. Конечно, длина регистрируемых волн значительно больше, чем длина волн света, и, следовательно, они могут быть носителями малого количества информации – даже в форме пространственно интерферирующих голографических узоров. Таким образом, предлагаемая далее гипотеза является развитием идеи, высказанной в предыдущих главах о той роли, которую играют в функционировании мозга микроструктуры, образованные медленными потенциалами соединений. Эти микроструктуры могут быть описаны либо в статистических понятиях квантовой теории, либо на языке интегралов свертки и преобразований Фурье, применяемых для описания механической волны. Микроструктуры не меняют своих существенных характеристик из-за того, что мы выбираем тот или другой способ описания. Каждый язык, каждая форма описания имеет свои преимущества. Для объяснения проблем восприятия, особенно проблем формирования образа и фантастической способности узнавания, голографическое описание не имеет себе равных. Поэтому почему же не попытаться применить голографическое описание к процессам мозга?
НЕЙРОННЫЙ ГОЛОГРАФИЧЕСКИЙ ПРОЦЕСС
Сущность голографической концепции состоит в том, что образы восстанавливаются, когда их представительства в виде систем с распределенной информацией соответствующим образом приво-. дятся в активное состояние. Эти представительства действуют как фильтры или экраны. Фактически, как мы уже отметили, представление о голографической процессе возникает еще при рассмотрении оптических фильтров. В этой связи голография понимается как мгновенная аналоговая кросс-корреляция, осуществляемая в результате сопоставления фильтров. Корреляция в мозгу может иметь место на различных уровнях. На более периферических уровнях возникает корреляция между последовательными конфигурациями, порождаемыми возбуждением рецепторов: остаточные явления, сохранившиеся после адаптации, действующей посредством механизма затухания, создают регистр буферной памяти, которая обновляется текущими входными воздействиями. На более центрально расположенных станциях корреляция влечет за собой более сложное взаимодействие: в любой момент времени входное воздействие коррелирует не только с конфигурацией возбуждения, существующего в любом пункте, но также со структурами возбуждения, прибывающими от других уровней системы. Пример этого вида сложности показан в экспериментах, описанных в гл. VII, где конфигурация изменения потенциалов в зрительной коре определялась не только зрительными стимулами, за которыми наблюдала обезьяна, но также условиями подкрепления и «намерением» осуществить тот или иной тип ответа.
Согласно голографической гипотезе, механизм этих корреляций не является следствием ни некоего разобщенного «динамического поля», ни даже изолированных, расщепленных волновых структур. Действительно, рассмотрим еще раз структуру более или менее постоянных организаций кортикальных колонок (или в других участках нервной системы других клеточных ансамблей), возникающих в результате прихода импульсов на нервные синапсы, которые активируют тормозные взаимодействия горизонтальной клетки. При конвергенции таких приходящих динамических структур, исходящих по крайней мере от двух источников, появляются интерференционные картины. Предположим, что оти интерферирующие динамические структуры, образованные классическими постсинаптическими потенциалами, имеют отношение к сознанию. Предположим также, что анализ, проведенный в начале этой главы, правилен и что эта микроструктура медленных потенциалов точно описывается теми уравнениями, которые описывают и голографический процесс, также строящийся на основе интерференционных узоров. Из этого следует вывод, что информация о входных воздействиях распределяется на всем протяжении нейронной системы точно так же, как она распределяется по всему узору физической голограммы. Однако в настоящее время мы еще плохо понимаем, как происходит такой процесс распределения. Следующий обмен мнениями по этой проблеме, проведенный в журнале «Nature», привлекает внимание к некоторым логическим аргументам.
Рис. VIII-10. Построение коррелограммы (верхний рисунок) и восстановление структуры (нижний рисунок). Д – диффузный источник света; Л – линзы; В – плоскость, в которой возникает коррелограмма между А и В. Сплошная линия показывает пути прохождения пересекающихся лучей при построении коррелограммы, пунктирная – непересекающихся (Willshaw, Buneman and Longuet-Higgins, 1969).
«Нашим отправным пунктом является наблюдение Габора, состоящее в том, что любая физическая система, которая может осуществлять корреляцию (или сплетать) пары узоров, способна имитировать работу голографа Фурье. Такая система, которая могла бы быть собрана в любой школьной фазической лаборатории, показана на рис. VIII-10. Такой аппарат предназначен для получения «коррэлограмм» между парами узоров, выполненных в виде булавочных отверстий, а затем для использования коррелограммы и одного из узоров для восстановления его пары.
Однако возникает небольшое затруднение при попытке понять, каким образом принцип работы дискретного коррелографа с его способностью к восстановлению мог бы быть реализован в нервной ткани. Мы не будем подробно останавливаться на этом пункте. Скажем только, что внимание в нему было привлечено д-ром Ф. X. С. Криком, у которого такое представление вызвало критические замечания. Но это ведет нас к дальнейшему усовершенствованию нашей модели (рис. VIII-11) … В такой форме наша модель ассоциативной памяти, теряя способность узнавать смещенные узоры, перестает быть коррелографом, но объем ее информации сейчас потенциально гораздо больше, чем прежде…
Рис. VIII-11. Ассоциативная сеть, описанная в тексте. Обратите внимание, что узлы на пересечениях могут представлять собой синаптические области (Willshaw, Buneman and Longuet-Higgins, 1969).
Итак, мы пытались вычленить в голографии те свойства, которые дают возможность представить ее как модель ассоциативной памяти, и обнаружили, что работу голографа можно имитировать и существенно улучшить посредством дискретных нелинейных моделей, а именно с помощью коррелографа и ассоциативной сети… Вполне возможно, что в мозгу нет системы, точно соответствующей принципу, на котором работает голограф, ш количественным отношениям, которые, как мы показали, должны соблюдаться, чтобы такая система работала с высокой эффективностью. (Willshaw, Buneman and Longuet-Higgins, 1969, p. 960-962).
Уиллшоу, Бунеман, Лонге-Хиггинс предложили неголографическую модель ассоциативной памяти мозга (Willshaw, Buneman and Longuet-Higgins, 1969, p. 960). Они также критикуют высказанное мной (van Heer-ец, 1963, p. 393) и Прибрамом (1966, 1969) предположение о том, будто мозг организован по голографическому принципу. Они говорят: «Каким образом мог бы мозг с достаточной точностью подвергать входные сигналы анализу Фурье…»
В книге, посвященной этой проблеме (van Heerden, 1968), я рассмотрел, каким образом мозг физически мог бы очень хорошо работать в качестве трехмерной голограммы. Если мы имеем трехмерную сеть нейронов, в которой каждый нейрон связан с несколькими соседними, и если нейрон определенного слоя, воспринимая сигнал, будет посылать его нескольким нейронам соседнего слоя, то сигналы в этой сети будут проводиться подобно тому, как волна распространяется в упругой среде. Более того, если способность нейронов к проведению воспринятых сигналов может постоянно возрастать благодаря частому их повторению, то сеть должна действовать как трехмерная голограмма, у которой почти все нейроны, входящие в сеть, обладают способностью к запоминанию.
Для моделирования узнавания нам необходимо сочетание двухмерной голограммы с ее способностью обеспечить быстрый поиск с трехмерной голограммой – с ее большим объемом памяти, – что может быть легко достигнуто (van Heerden, 1968). Однако, чтобы объяснить удивительную емкость человеческой памяти, этого еще недостаточно. Мы можем узнать человека, с которым долгое время не встречались, на любом расстоянии и при различном угле зрения. Память, зафиксированная в голограмме, не способна осуществить эту операцию. Необходимая пластичность может быть достигнута с помощью оптических средств: например, хрусталик может выполнять исследовательскую функцию, сопоставляя размеры воспринимаемого образа и образа, сохраненного в памяти. По-видимому, естественно предположить, что нейронная сеть обладает такой пластичностью. Она может реализовываться благодаря изменяющим свою протяженность полям, аналогичным тем, которые используются в электронной оптике, и электрическим или химическим путем вызывать различные изменения в скорости проведения сигнала через сеть нейронов. Это могло бы повлечь за собой изменение фокального расстояния, или вращение образа, или небольшие его искажения для достижения ясного, четкого распознавания сигнала в плоскости проекции.
Хотя голографический принцип вполне естествен для нейронной сети, не исключена возможность, что в действительности в мозгу реализуется другая модель, такая, как, например, коррелограмма Уиллшоу, Бунемана и Лонге-Хиггинса. Однако следует прежде всего показать приемлемость такой модели. Их модель в той оптической форме, которую они предла гают, видимо, обладает малым объемом памяти из-за дифракции, имеющей место в любом волновом ноле (а это важно!). С другой стороны, в предложенной ими сетевой модели они получают такой же объем памяти, что и в топографической, но, по-видимому, в ущерб пластичности системы, необходимой для распознавания смещающихся, различных по величине или слегка искаженных образов. Следует рассмотреть еще один аспект, а именно: способность трехмерных голограмм запоминать временные отношения сигналов (van Heerden, 1963). Любая модель должна объяснить еще один аспект обработки информации в мозгу – распознавание речи, способность говорить, или бежать, или управлять автомобилем.
П. Дж. ван Хирден
Ван Хирден обсудил некоторые различия, существующие между голо-графической моделью памяти (1963) и парой неголографических моделей, которые мы предложили в прошлом году (Willshaw, Buneman and Longuet-Higgins, 1969).
Действительно, коррелограф в своей оптической форме имеет несколько ограниченные возможности из-за дифракции и не может быть принят в качестве физической модели памяти; но мы и не предполагали этого. Однако мы считали, что его логика, которую легко уловить, может быть реализована в нервной системе. Например, можно построить ассоциативную сеть, функционирующую как коррелограф, путем «связывания» определенных ее переключателей. Однако нет данных, что такое связывание имеет место в нервной системе, поэтому мы предложили рассматривать ассоциативную сеть как наиболее вероятную модель. Ее можно было бы легко реализовать, как мы уже говорили, посредством системы нейронов, имеющих свои пороги и изменчивые синапсы. Известно, что оба эти свойства характеризуют периферическую нервную систему (Eccles, 1957, 1964} и, вероятно, также центральную (Burns, Bliss and Uttley, 1968).
Хотя нет еще окончательных нейрофизиологических данных, подтверждающих нашу теорию и опровергающих теорию ван Хирдена, способность отдельных участков нервной системы проводить волны в соответствии с принципом Гюйгенса трудно, по-видимому, согласовать с наблюдениями, свидетельствующими о нелинейности некоторых нейронных ответов, а существование стабильного периодического источника возбуждения нужно еще доказать. Мы также считаем, что преимуществом любой модели мозга является способность модифицировать как синапсы, так и состояние клеток. В коре головного мозга отношение синапсов к нервным клеткам, по-видимому, порядка 10 4 – 10 5 ; таким образом, информация, которая могла бы сохраняться в синапсах, соответственно значительно больше (Cragg, 1967). Что же касается удивительной пластичности механизма восприятия у человека, то мы считаем, что ни модель ван Хирдена, ни наша в их современной форме не в состоянии объяснить этой пластичности».
Д. Уиллшоу
X. Лонге-Хиггинс
О. Бунеман
«Я согласен с утверждением Уиллшоу, Бунемана и Лонге-Хиггинса в ответ на мое сообщение (van Heerden, 1970), что предложенная ими ассоциативная сеть (Willshaw, Buneman and Longuet-Higgins, 1969), так же-как голограмма, выполняет специфическую функцию. Однако в их сети не представлены две наиболее удивительные способности человеческой памяти. Первая – это наша способность узнавать, знакомого человека среди более чем сотни людей, которые могут находиться в нашем поле зрения. Мы можем внезапно ощутить вспышку узнавания, эту абсолютную уверенность, что «это именно он и никто другой»; она не является просто субъективным чувством, а вызывается, очевидно, крайне надежной и быстрой обработкой информации в мозгу. Эта функция распознавания выполняется также двухмерной голограммой в оптическом устройстве, когда в плоскости его проекции появляется яркая светящаяся точка. Яркость и четкость изображения световой точки является научно обоснованной мерой уровня распознавания.
Второй способностью нашей памяти является способность поел© узнавания данного лица быстро воспроизвести значительное количество информации, которой мы о нем располагаем. В оптическом устройстве сигнал от двухмерной голограммы, свидетельствующий об распознавании образа, является инструкцией для трехмерной голограммы осуществить полное воспроизведение соответствующей информации.
Верно, что научные усилия объяснить возможности человеческого интеллекта на основе теории деятельности мозга находятся еще на начальной стадии. Однако мало сомнений в том, что мы располагаем основой для такой теории. Такой основой является теория информации, та саман теория, которая используется в радио, телевидении, радарной технике и фотографии. В теории информации распознавание, или сообщение о количественной степени сходства двух вещей, описывается корреляциопной функцией двух временных функций, или двух образов. Сложное вычисление корреляционной функции может быть описано математически как операция фильтрации, но первоначально, разумеется, должен быть произведен расчет фильтра, который требуется для осуществления этой операции фильтрации и с которым будут сопоставляться сигналы. Тот факт, что голограмма, подобно нейронной сети с постулированными нами простыми свойствами, осуществляет свою функцию фильтра с 50%-ной эффективностью, обусловлен тем, что по счастливой случайности – или, может быть, это заложено в самой природе вещей – распространяющееся волновое ноле автоматически выполняет это трудоемкое вычисление, отвечая требованиям теории».
П. ван Хирден
Существование нейронного голографического или сходного с ним процесса не означает, конечно, что входная информация волей-неволей распределяется по всей глубине и поверхности мозга. Информация распределяется только в тех ограниченных областях, где входные воздействия действительно вызывают устойчивые узоры синаптических микроструктур. Более того, для объяснения любого эффекта, развивающегося вслед за специфическим входным воздействием, следует привлечь более локализованные механизмы памяти, описанные в первой части. Однако информация иногда может быть введена в участки, которые распределены по нейронному пространству, и тогда она становится рассеянной. Восстановление того, что более длительное время хранится в памяти, зависит главным образом от повторения данной структуры, которая первоначально вызвала этот процесс сохранения, или ее существенных частей. Эта способность «адресоваться» прямо к содержанию информации безотносительно к ее локализации, которая столь легко достигается в голографическом процессе, устраняет необходимость иметь в мозгу специальные пути или пункты для хранения информации.
Какими возможностями располагает нервная система для формирования устойчивых микроструктур соединений? Объяснить хранение информации в течение разного времени можно лишь с помощью какого-то более постоянного свойства протоплазмы. Сильное взаимодействие во времени возникает между входными сигналами, разделенными интервалом в несколько часов (как в случае эффекта Мак-Каллока, когда экспозиция набора цветовых полос оказывает влияние на последующее восприятие цвета) или у некоторых индивидов в несколько дней (как у редко встречающихся людей с настоящей эйдетической способностью). И конечно, для объяснения узнавания и воспроизведения следует принять во внимание более продолжительные процессы взаимодействия. Структурные изменения в таких макромолекулах, как липиды или протеины, и даже длительное анизотропное упорядочение макромолекулярных структур дают основание для следующих рассуждений. Сходные по конфигурации и последовательно возникающие микроструктуры соединений могут порождать остаточный кумулятивный эффект путем упорядочения вначале случайных цепей макромолекул или волокон либо путем увеличения их возбудимости, вследствие чего эта область с большой легкостью отвечает на повторение того же самого возбуждения. Ранние результаты экспериментов, проведенных на культуре ткани сетчатки, которая была исследована с помощью электронного микроскопа, показывают, что такие изменения в молекулярной структуре могут возникать при возбуждении (Sjostrand, 1969). Сходные, хотя и не подтвержденные еще, предположения были сделаны Л. Уайтом (1954) и У. Халстэдом (Katz and Halstead, 1950). Уайт предполагает, что
«…это кумулятивное упорядочение, сохраняющееся в течение более или менее длительного времени у некоторых протеиновых цепей, обнаруживается по всему специфическому объему кортикальной цитоплазмы и представляет собой один из способов процесса роста структур нервной ткани, которые определяются не наследственностью, а активностью. Этот процесс предполагает не рост дифференцированной ткани, а упорядочение молекулярной структуры обширной массы цитоплазмы. Здесь мы имеем дело с дифференциацией направлений специфического вектора, который в особых кортикальных слоях, возможно, ориентирован параллельно поверхности коры. Шаблоны памяти – не единственные локализованные молекулярные структуры. Существуют также обширные компоненты системы, создающей долговременное упорядочение молекулярной структуры, которые ориентированы под различными углами друг к другу… (Однако) упорядочение будет соответствовать только статистически доминирующей структуре активности или простейшему принципу последовательного изменения динамических структур типичной активности. Более того, эта тенденция выбирать доминирующую структуру будет подкрепляться тем, что простейшие, самые общие динамические структуры будут наиболее стабильными, так как их части будут взаимно поддерживать друг друга. Случайным образом организованные протеиновые структуры могут, следовательно, действовать как структурный фильтр, получая стабильный оттиск сперва только от наиболее простого, наиболее унифицированного и статистически доминирующего компонента во всех динамических структурах активное ги данной общей формы… (Затем) развитие процесса модификации структуры идет в направлении от сильно упрощенной к менее упрощенной и более точной записи. Этот процесс развития иерархически организованной модификации соответствует «прогрессивной индивидуализации» поведенческих форм в ходе онтогенеза (Coghill) и может служить ключом к пониманию самокоординирующейся способности кортикального процесса» (Whyte, 1954, р. 162-163).
Конечно, здесь отмечено не все, что свидетельствует о необходимости регистрации волновых форм. Структурные изменения в макромолекулах обладают способностью к восстановлению. Вероятно, более устойчивая запись требует таких механизмов, как настройка «усредняющих циркуляции» в кортикальных колонках и рост, вызванный изменениями в мембранной проницаемости вследствие и в зависимости от этих макромолекулярных изменений. «Фильтр», или «экран», топографических узоров состоит нетолько из решетки макромолекул мембраны, образующих синаптически-дендритную сеть, но также из облегчения всех тенденций формировать образ и создавать выходные динамические структуры нервных импульсов.
ПОДТВЕРЖДЕНИЕ ГИПОТЕЗЫ
Одна группа данных в пользу голографической гипотезы идет от исследований развития зрительного восприятия у новорожденных. Вопреки широко распространенным представлениям (например, Хебб) константность величины и формы не возникает исключительно из комбинаций следов прошлых и, по-видимому, более элементарных восприятий (например, восприятие длины линий, углов). Предположение Хебба основывалось на результатах экспериментов и клинических наблюдений, когда испытуемые находились в условиях сенсорной депривации. Он считал, что для соответствующего развития рецепторной функции необходим опыт. Однако электрофизиологические эксперименты (воспроизведенные на рисунке VII-12) показывают, что сетчаточные и нейронные механизмы, необходимые для детекции признаков, уже хороша развиты у млекопитающих (кошек) к моменту рождения. Поэтому эффект, наблюдающийся при ограничении сенсорных воздействий, по-видимому, является следствием либо атрофии, возникающей в результате бездействия синапсов, либоактивного подавления сенсорного механизма посредством вызванного ненормального функционирования. Как уже отмечалось, у человека возникает аналогичная ситуация. Бауер (1966) проводил эксперименты с шестинедельными младенцами, и эти эксперименты показали, что у них уже наблюдается константность величины ж в значительной степени константность формы. В этих экспериментах предметы помещались таким образом, что их действительная величина, расстояние до них и сетчаточный образ (который вычисляется исходя из действительной величины рассматриваемой фигуры) могли образовывать пары в различных сочетаниях. Использовалось полное и незаконченное изображение фигур-Плоское двухмерное изображение предметов заменяло трехмерные предметы. При обучении младенца различению экспериментатор в качестве подкрепления использовал игру в прятки. Затем производилась проверка, которая состояла в выявлении у младенца заученного ответа при замене одного стимула пары другим. Результаты экспериментов показали, что константность возникает у младенца только тогда, когда он располагает сведениями о расстоянии или когда один элемент фигуры перемещается,, тогда как остальные остаются на прежнем месте. Таким образом, даже у младенцев зрительная система представляет собой врожденную организацию, регистрирующую «такой вид информации, который порождается двигательным и бинокулярным параллаксом». Какой же тип встроенного механизма мог бы регулировать параллакс лучше, чем голографический процесс, который сам конструируется посредством параллактического процесса (эффекта интерференции)?
Прямым доказательством того, что в формировании образов участвует параллактический кортикальный механизм, подобный голограмме Фурье, являются результаты экспериментов, проверенных Кемпбеллом и его сотрудниками (Blakemore and Campbell, 1969; Campbell et al. 1968; Campbell, Cooper and Enroth-Cugell, 1969; Campbell and Robson, 1968). Эти исследования показали, что кора головного мозга может настраиваться на восприятие пространственной информации разного диапазона. Эксперименты -были проведены на кошках и на людях. Предъявлялись решетки с различной контрастностью и изучалось влияние их вращения на зрительные вызванные потенциалы (у человека) и на динамические структуры разрядов, возникающих в нейронах зрительной коры (у кошки). Нейроны отвечали на ограниченную полосу пространственной частоты (в четыре октавы), а длительное рассматривание вызывало подавление контрастной чувствительности за пределами соседних частот (рис. VIII-12). Взаимоотношение между этими экспериментальными результатами и голограммой Фурье лучше всего сформулировано самими авторами.
Рис. VIII-12. Эффект пространственной адаптации на вызванные потенциалы у человека. Синусоидально-волновой узор решетки (12 кол/градус) сдвигался вокруг своей оси на 180° со скоростью 8 раз в сек. Вызванный потенциал зрительной коры суммировался 200 раз на специальном аппарате для получения записей, показанных в рамках слева. Для всех записей стимул был одинаковым. Каждая кривая показывает потенциал для двух фазовых сдвигов пространственной решетки. Первая запись получена при рассмотрении решетки низкой контрастности (верхняя часть справа расположенной панели), составляющей 10 0 ' 7 порогов контрастности для этой пространственной частоты. Затем испытуемый в течение 30 сек рассматривал решетку высокой контрастности (на 1,5 единицы выше порога), после чего немедленно производились записи вызванного потенциала в ответ на повторное предъявление решетки с тем же низкич контрастом. Эта вторая кривая имеет гораздо меньшую амплитуду, чем первая. После 5-минутного периода восстановления предъявление решетки низкой контрастности вновь вызывает запись (3-я рамка), очень сходную с первоначальной. Последняя запись вызванного потенциала на предъявление решетки низкой контрастности после 60 сек экспозиции решетки высокой контрастности не содержит ответа. Отсутствие в записи вызванного потенциала сопровождается попышением субъективного порога (Blakemore and Campbell, 1969).
«В этом исследовании мы намеренно использовали простейший зрительный стимул… Решетка с синусоидально-волновым узором проста потому, что она содержит только одну пространственную частоту, которая представлена в одном меридиане. Наиболее сложным стимулом, исходя из частотного анализа Фурье, является одиночный, резко очерченный круг света, так как он содержит очень широкую полосу пространственных частот и они ориентированы в разных направлениях. Частотная «ширина полосы» индивидуальных пространственных механизмов, выявленная с помощью адаптации, достаточно узка (примерно 1 октава для половины амплитуды). Следовательно, любая сложная световая структура, воздействующая на сетчатку и содержащая широкий спектр компонентов- Фурье, будет активировать многие механизмы. Мы склонны предположить, что структура ответов, получаемых от группы подобных механизмов, может служить для кодирования пространственного содержания определенного сетчаточиого образа и, следовательно, может каким-то неизвестным способом приводить к его идентификации.
Преимущество системы, основанной на частотном анализе, возможно, состоит в том, что она упрощает узнавание знакомых объектов, которые предъявляются непривычно увеличенными. Рассмотрим случай с ребенком, который только что научился различать буквы алфавита, и допустим, что его просят узнать буквы, имеющие разный масштаб увеличения. Он делает это без труда, хотя никогда ранее не видел букв такого размера. Известно, что если мы находимся так близко к предмету, что не можем воспринять его целиком, то он не может быть быстро идентифицирован: «мы не можем видеть за деревьями леса». Сатерленд (1968) дал хороший обзор литературы, посвященной константности величины, и пришел к заключению, что «многие виды обладают способностью воспринимать форму как ту же самую независимо от изменения ее величины, во всяком случав в значительных пределах, и что эта способность врожденная».
Должно быть, существует ограниченный диапазон пространственных величин, которыми зрительная система может управлять легко и быстро. Если она анализирует распределение пространственных частот в объекте лю системе независимых каналов, охватывающих диапазон ее действия, а затем использует соотношение этих частот, чтобы идентифицировать объект, то воспроизведение абсолютной величины объекта было бы излишним для опознания образа, так как отношения внутри гармонического состава не зависят от абсолютной величины. Только этот гармонический состав, должно быть, и фиксируется в системе памяти, и это требует значительно меньшего объема памяти, чем в том случае, если бы распознавание каждого знакомого объекта основывалось на раздельном обучении распознаванию его при каждом изменении его размера. Это явление генерализации величины и, следовательно, расстояния значительно облегчило бы процесс обучения распознаванию образов в нашем естественном окружении. Эта система могла бы быть аналогичной слуховой системе, которая может идентифицировать музыкальные интервалы (отношение частот) независимо от их положения на звуковом спектре…
Такой механизм анализа пространственных частот было бы трудно допустить, если бы он должен был действовать одновременно в двух измерениях. Возможно, имеет большое значение то, что зрительная система также проводит входной сигнал через ряд отдельных избирательно ориентированных каналов, каждый из которых может затем анализировать содержание пространственной частоты объекта в пределах узкого диапазона пространственной ориентации… Хотя такая организация вела бы к дальнейшей экономии объема памяти, она имела бы свой недостаток: ни один из каналов был бы не ограничен в распознавании знакомых объектов, лишь при условии их предъявления при ранее заученной ориентации.
Вот доказательство.
Мы не в состоянии описать явление генерализации ориентации так же хорошо, как мы можем это сделать в отношении размера объекта» (Blake-more and Campbell, 1968, p. 257-259).
Другая линия доказательств основана на том опыте, который мы все имеем. Можно вспомнить и представить себе множество мельчайших подробностей, когда мы оказываемся в соответствующей обстановке – например, приехав в район, где мы жили много лет назад, мы вспоминаем магазины, расположение дверей, мебель в гостиной, которые еще несколько часов назад казались забытыми навсегда. Какой механизм может действовать лучше, чем ассоциативное припоминание, которое обеспечивается голографическим процессом?
Конечно, побудительные импульсы, извлекающие образы из памяти, исходят не от рецепторов. Как я уже отмечал и буду подробно говорить об этом в гл. XVII и XVIII, возбуждение так называемых ассоциативных зон мозга может вызывать во входных каналах динамические структуры возбуждения, аналогичные тем, которые вызываются сенсорными раздражениями. Однако 'Образы, которые предположительно возникают при таком возбуждении, как правило, легко отличить от образов, вызываемых возбуждением рецепторов. Лишь в таких особых условиях, как временная сенсорная депривация, дифференциация внутреннего и" внешнего механизмов формирования образов нарушается, вызывая появление различных иллюзий и галлюцинаций. Однако сам факт, что образы такого типа возникают и формирование их имеет сходство с процессом восприятия, свидетельствует о том,, что восприятие само по себе в значительной степени является реконструктивным. Какой иной процесс, кроме процесса реконструкции образа посредством голографии, может выполнить эту функцию?
Главной зоной реконструкции зрительных образов является стриарная область затылочной коры. Известно, что люди, подвергшиеся двустороннему удалению затылочной доли, полностью лишены зрительных образов (Konorski, 1967). Периферическое разрушение не вызывает такого эффекта в слуховой модальности. Прекрасным примером этого может служить Бетховен. Несмотря на периферическое нарушение слуха, он сохранил достаточное воображение, чтобы написать Девятую симфонию и поздние квартеты.
Другие данные, согласующиеся с голографической гипотезой,, получены в экспериментах с электрической стимуляцией проекционной области зрительной коры человека (Brindley and Lewin, 1968). Такая стимуляция, если в ней закодирована только интенсивность и не закодированы фазовые отношения, вызывает появление пятен света, выглядящих подобно «звездам в небе» и не похожих на линии или углы. Когда такие пятна возникают в стороне от точки фиксации взора, они становятся слегка удлиненными подобно «зернам риса». Наиболее периферические восприятия выглядят подобно «облаку» и имеют «величину горошин, которые рассматриваются со значительного расстояния». Эти образы появляются на постоянных участках зрительного поля. Однако в результате стимуляции через несколько-отдельных электродов у испытуемого можно вызвать видение-простых зрительных структур. При произвольных движениях глаз воспринимаемые пятна света движутся вместе с глазами; при рефлекторных движениях, вызываемых вестибулярной стимуляцией, воспринимаемые пятна света остаются фиксированными в пространстве. Иногда после очень сильной стимуляции зрительные образы сохраняются в течение одной-двух мипут. Все-эти удивительные наблюдения предполагают существование некоторого механизма, помимо механизма детекции признаков, необходимого для построения весьма сложных зрительных образов,, воспринимаемых нами в повседневной жизни.
Еще одна интересная линия доказательств в пользу существования нейронного топографического процесса связана с исследованиями частичных поражений затылочной коры. В соответствии. с моделью простого детектора не следовало бы ожидать каких-либо нарушений вне области скотомы, полученной в результате локальных поражений коры. Но тщательно проведенные исследования Г.-Л. Тейбера и В. С. Баттерсби (Teuber, Battersby and Bender, 1960) показали, что во всей сохранившейся части поля зрения быстрее теряется четкость контуров фигур; снижаются пороги слияния мельканий и нарушается восприятие реального кажущегося движения.
Все эти феномены зависят, вероятно, от тормозных взаимодействий, которые представляют собой фильтр, создаваемый эффектами интерференции нейронного голографического процесса. В таком фильтре при большом повреждении следовало бы ожидать скорее изменения качества образов, формируемых оставшейся частью фильтра, чем нарушения самого процесса их формирования.

