Книга: Эра млекопитающих: Из тени динозавров к мировому господству
Назад: 6 Модернизация млекопитающих
Дальше: 8 Млекопитающие и климатические изменения
7
Экстремальные млекопитающие
Deinotherium
Туристы нашли кости самого большого животного в истории?25 июля 2831 г.
«Нью-Майами» (информационное агентство). Одна семья, путешествуя по пустыне Флорида, наткнулось на гигантские окаменелые кости, принадлежавшие, вероятно, одному из самых больших животных в истории Земли.
По оценкам вызванных на место находки палеонтологов, это было животное длиной более 30 м, а весило оно, вероятно, свыше 100 т.
«Поразительно, – говорит профессор Лола Брикер из Института изучения климата и окружающей среды Нью-Майами. – Никогда не видела таких великанов. Это идет вразрез со всеми представлениями о возможных размерах животных».
Разбросанные кости занимают площадь больше футбольного поля. Они принадлежат пока еще безымянному существу с длинным цилиндрическим телом, напоминающим подводную лодку.
«Если мои расчеты верны, оно было как минимум в два раза больше любого современного животного», – сказала журналистам Брикер, стоявшая рядом с одним из ребер гиганта, которое возвышалось над ее головой сантиметров на тридцать.
Ученые сообщают, что нашли фрагменты передних конечностей животного, похожих на рыбьи плавники, но никаких признаков задних пока не обнаружено. Его массивная голова с дугообразными беззубыми челюстями сохранилась отлично.
«Вся семья вчетвером действительно поместилась у него в пасти», – сказала Брикер, имея в виду облетевшую мир фотографию, которая была сделана туристами, когда они нашли кости, и которую многие ученые мужи сочли подделкой.
Чем этот зверь питался и как он передвигался, остается загадкой. Ученые говорят, что кости были обнаружены в отложениях глинистого сланца, образовавшихся на морском дне около 5000 лет назад, когда Флорида была зеленым субтропическим полуостровом, разделявшим Мексиканский залив и Атлантический океан.
«Я уверена, что оно было водным, и по форме слуховых косточек могу сказать, что это было млекопитающее, но это все, что нам известно на данный момент», – заявила Брикер.
Она планирует выкопать кости и собрать скелет в лаборатории, но опасается, что ей не хватит ресурсов.
«Мне понадобится как минимум двадцать человек и, вероятно, шесть месяцев, чтобы извлечь все кости, а затем – достаточно большое помещение для изучения в моем университете», – объясняла она в надежде на отклик какого-нибудь богатого благотворителя, который профинансировал бы ее исследование.
Эти затраты оправданны, утверждает она, так как подобные находки поощряют интерес детей к природе.
«Вы можете вообразить себе такого зверя живьем? Представить, что его сородичи живут рядом с вами? Это поражает воображение», – говорит она.
Прежде чем извлекать кости, Брикер надеется уточнить видовую принадлежность колосса.
«У моих коллег это может вызвать насмешки, но в сагах тысячелетней давности упоминаются левиафаны, обитавшие в морях. Они назывались китами. Есть рассказы о том, что синий кит достигал 30-метровой длины. Мы всегда считали их мифическими, но, может быть, мы ошибались».
Этот шуточный пресс-релиз, конечно, выдуман и несколько пародиен. Хочется надеяться, что Флорида не превратится в пустыню, мы не утратим связи со своей письменной историей, а киты не вымрут! Но представьте себе, что они исчезнут и будущие поколения найдут их окаменелые кости. Наверняка люди будут испытывать такой же восторг, какой испытываем мы по поводу колоссальных динозавров – как великанов прошедших эпох, которых нам не довелось увидеть во плоти.
Многие из нас – честно говоря, и я тоже – часто не осознают тот факт, что и в наши дни, на одной с нами планете существует немало животных-великанов. Многие из них – млекопитающие. Синий кит – самое экстремальное из этих экстремальных млекопитающих. Это не просто крупнейшее из ныне живущих млекопитающих, а самое крупное из реально существовавших животных, точка. Никто никогда не находил ископаемых остатков животных крупнее его, а значит, синий кит – рекордсмен всех времен, чемпион тяжелого веса в истории мироздания.
Это простое, но содержательное высказывание, которое необходимо повторить: самое большое животное в истории живет именно сейчас. Из миллиардов видов, живших на протяжении миллиардов лет истории Земли, мы – в числе тех немногих, кому выпала счастливая возможность сказать это. Разве не замечательно, что мы с синим китом дышим одним воздухом, плаваем в одних морях, видим одни и те же звезды?
Пока вы читаете эти строки, синие киты бороздят океаны – все океаны, так как их ареал практически всемирный, за исключением самой северной оконечности Арктики. Наиболее крупные взрослые особи чуть более 30 м в длину и 100–110 т весом, что на добрых 20 т больше максимального взлетного веса «Боинга-737» и, вероятно, на 30–40 т больше самого огромного динозавра. Самки синего кита рожают трехтонных детенышей величиной с моторный катер, и те за первые полгода вскармливания достигают веса в 15 т. Взрослые киты способны нырнуть на глубину до 315 м, задержать дыхание на час с лишним и вынырнуть, выпустив из дыхала фонтан воды высотой в два этажа. В свою растяжимую пасть они могут разом втянуть всю воду из небольшого плавательного бассейна – и проделывают такое по нескольку раз в день, чтобы собрать 2 т криля (мелких рачков вроде креветок), из которого извлекают энергию для своего метаболизма. Они умны и общительны, а их низкочастотные вокализации – самые мощные звуки в царстве животных, способные разноситься в пучине на полторы тысячи километров.

Но не все так радужно. По оценкам специалистов, 99 % популяции синих китов было уничтожено китобоями за прошедшие два столетия. От сообщества, некогда насчитывавшего сотни тысяч, в данный момент осталось самое большее несколько десятков тысяч особей. Пусть это звучит банально, но давайте воздадим им должное, пока не поздно, и давайте сделаем все возможное, чтобы сберечь и защитить их, пока у нас еще есть шанс и пока синий кит не разделил участь бронтозавра.

Синий кит, самое большое животное из когда-либо живших на Земле. Скелет в Лондонском музее естественной истории (на предыдущей странице); Трэвис Парк, специалист по ископаемым китам, позирует рядом с черепом.
Фотографии Яна Беранека и Трэвиса Парка соответственно
Говоря о синих китах, сложно избежать гипербол. Это относится и к другим экстремальным млекопитающим, будь то другие киты, или гигантские наземные виды вроде слонов, или более мелкие млекопитающие, перестроившие свой организм ради удивительных достижений, например летучие мыши – единственные млекопитающие, способные к машущему полету, представители одного из всего лишь трех классов позвоночных, которые вообще это сумели (остальные два – птеродактили и птицы). Все эти экстремальные млекопитающие – слоны, киты и летучие мыши – начали свой путь в эоцене и только в ходе длительной эволюции достигли нынешних высот.
Слоны – крупнейшие из современных наземных млекопитающих и вообще наземных животных. Самый крупный вид – африканский саванный слон, достигающий 3 м в холке, что соответствует высоте баскетбольной корзины. Самцы весят 5–7 т, вдвое больше пикапа «Форд F-150». Если бы один саванный слон уселся на конец доски качелей, понадобилось бы сто человек, чтобы его уравновесить. Конечно, слоны меньше синих китов, но им приходится иметь дело с ограничением, которого не знают синие киты, – гравитацией. Синий кит может пассивно плавать, используя поддерживающую силу воды, но слоны опираются на все четыре конечности, когда они поднимаются с земли, передвигаются и спариваются.
Ныне сохранилось лишь три вида слонов, рассеянные по Черной Африке, Индии и Юго-Восточной Азии. Это всего лишь жалкие остатки некогда процветавшего семейства, распространенного на многих континентах, – в него входили такие виды, как мамонты и мастодонты, и некоторые представители весили вдвое больше африканского саванного слона, а значит, были самыми большими наземными млекопитающими в истории (ну, вероятно, как мы увидим дальше). Однако до своего всемирного распространения и последующего упадка слоны десятки миллионов лет обитали исключительно в Африке – как одна из ветвей великого разнообразия афротериев.
Афротерии, как мы узнали в предыдущей главе, – одно из четырех основных подразделений родословной плацентарных млекопитающих. Как и ксенартры – группа, куда входят ленивцы и броненосцы, – афротерии отделились от ствола родословного древа у самых его корней, то есть они являются одной из самых примитивных групп плацентарных. Другое их сходство с ксенартрами состоит в том, что ранняя история афротериев в палеоцене и эоцене протекала на одном континенте, изолированном от остального мира. Ленивцы и броненосцы, как и южноамериканские копытные вместе с саблезубыми сумчатыми, жили, разумеется, на островном Южно-Американском континенте, а слоны и другие афротерии, как следует из их названия, – в Африке.
Десятки миллионов лет Африка оставалась изолированной крепостью. После того как около 100 млн лет назад, в меловой период, она откололась от Гондваны – южной половины бывшей Пангеи, – она существовала самостоятельно. К западу от нее располагался расширявшийся Атлантический океан, который изредка пересекали на плотах обезьяны и грызуны. К юго-востоку лежал Индийский океан, омывавший Антарктиду и Австралию; по нему ускоренно двигался Индийский субконтинент, который в эоцене столкнется с Азией. А к северу от Африканского континента лежало теплое экваториальное море Тетис, отделявшее северные материки от южных. Море Тетис не было непреодолимым барьером; северным животным то и дело удавалось перебраться на юг по архипелагу европейских островов, доходивших до североафриканского побережья, но это было трудное путешествие. Долгая ночь африканской изоляции окончилась 20 млн лет назад, когда Аравия сомкнулась с Евразией, и от моря Тетис остался только западный рукав, который мы теперь зовем Средиземным морем.
Слоны – одни из множества афротериев. Наряду с ними в семейный альбом входят водные ламантины, маленькие даманы с копытцами, питающиеся муравьями трубкозубы, подземные златокроты, тенреки Мадагаскара (и близлежащих островов), прыгунчики, которых зовут также слоновыми землеройками – они похожи на мелких грызунов со слоновьими хоботками. Если этот набор вам кажется странным, то вы не одиноки. Не так много есть очевидных анатомических признаков – если таковые вообще существуют, – объединяющих афротериев. Может быть, предполагают некоторые ученые, у них есть один общий характерный зубной бугорок или какие-то нюансы развития позвоночника. Возможно. Однако основные данные в пользу единства афротериев – генетические, и они убедительны. Установление родства слонов, ламантинов, даманов и тенреков стало одним из самых поразительных достижений первых генетических родословных построений на рубеже 1990-х и 2000-х гг. Оно выдержало проверку временем, и его подтвердили более совершенные полногеномные анализы. К досаде анатомов старой школы, генетический тест оказался убедительным, и надотряд афротериев существует реально.
Современные афротерии так непохожи друг на друга потому, что диверсифицировались очень давно – они произошли от примитивного общего предка и заполнили разнообразные экологические ниши Африканского островного континента. Этот предок, вероятно, был «архаическим» плацентарным величиной с собаку и похожим на кондиляртр из Нью-Мексико, причем, вероятно, пришел он с севера по островам моря Тетис в самом конце мелового периода или в начале палеоцена, вскоре после падения астероида. С исчезновением динозавров внезапно открылось множество вакансий в пищевой цепи, и афротерии воспользовались шансом. Они адаптировались к наземным лесным и луговым нишам, к жизни на деревьях и на побережье моря Тетис.
Параллельный процесс независимо шел на севере, где диверсифицировалась еще одна из четырех основных групп плацентарных – лавразиатерии. На севере возникли непарнокопытные и парнокопытные, в Африке – даманы. На севере появились питающиеся муравьями панголины и роющие кроты, в Африке – трубкозубы и златокроты. На севере были землеройки и ежи, а в Африке – прыгунчики (они же слоновые землеройки) и тенреки, которых иногда называют «мадагаскарскими ежами». И как мы вскоре увидим, киты в ходе эволюции появились на севере, в то время как в Африке аналогичную трансформацию из бегающих в плавающих претерпели сиреновые (отряд, куда входят ламантины). Это еще один красивый пример конвергентной эволюции. Примитивные плацентарные оказались изолированными на разных территориях в мире, внезапно освободившемся от динозавров. Они не имели контакта друг с другом – не могли встречаться и обмениваться генами, – однако в ходе их диверсификации эволюция пошла сходными путями и приспособила афротериев и лавразиатериев к сходным нишам.
С позиции сегодняшнего времени трудно оценить диверсификацию афротериев. Во многом она оказалась перекрыта более поздней иммиграцией животных с севера – предков зебр и антилоп, львов и гиен – после того, как Африка соединилась с Евразией. Но в палеоцене, а особенно в эоцене и последующем олигоцене Африка была царством афротериев. Там было на что посмотреть.
Даманы упоминаются в Библии под названием «горных мышей», которые «народ слабый, но ставят домы свои на скале», однако нам, выходцам с северных континентов, простительно не знать об их существовании. Ныне существует пять видов, обитающих только в Африке и на Ближнем Востоке: либо это коренастые, похожие на сурков травоядные, которые карабкаются по скалам на своих копытцах, либо древолазы с подушечками-присосками на лапках. Однако в эоцене и олигоцене в Африке обитали десятки видов даманов, от совсем мелких до титанохиракса (Titanohyrax), который был величиной с носорога и весил 1,3 т. Одни были всеядными, другие обладали складчатыми зубами, которыми жевали растения, третьи специализировались на семенах и орехах. Антилохиракс (Antilohyrax), бегающий по лесу на длинных ногах, был точь-в-точь антилопа, в то время как другие раскапывали землю длинным рылом, как свиньи. Даманы занимали ниши, которые впоследствии займут антилопы, бородавочники, бегемоты и носороги. В сущности, это были дамановые аналоги знакомых нам копытных современной Африки.
Тем временем на побережье стали осваиваться первые сиреновые. Вначале они были наземными животными с мощными лапами, которыми можно было также загребать по-собачьи на мелководье. В эоцене эволюция полностью перестроила их для водного образа жизни: их передние лапы превратились в ласты, а задние исчезли – на смену им пришел широкий веслообразный хвост. Двигая им вверх-вниз, можно было, пусть и неуклюже, плавать, преодолевая течение. Сиреновые стали первыми афротериями, распространившимися по всему миру. Они мигрировали вдоль побережья моря Тетис и вдали от него; их ископаемые остатки обнаруживаются повсюду, от Северной Каролины до Венгрии и Пакистана. Но, как и в случае с даманами, от их былого величия мало что осталось. Ныне существует лишь три вида ламантинов – на Карибах, в Западной Африке и на Амазонке – и один вид дюгоней, обитающий в Индийском океане и на юго-западе Тихого. Они стали трогательными символами природы, нуждающейся в защите: их вялый образ жизни – медленно плавающих, питающихся водной растительностью – делает их легкими жертвами рыболовных сетей и столкновений с судами.
Если мы не убережем их, ламантины и дюгони могут вымереть. Если это случится, они пополнят скорбный список легиона других афротериев, некогда разнообразных и господствовавших, но затем исчезнувших. Самый примечательный из них – арсиноитерий (Arsinoitherium), один из тех вымерших млекопитающих, что составляют пантеон фантастических зверей. Он напоминал носорога на стероидах и обладал двумя огромными рогами, торчавшими на лбу, при этом каждый был значительно выше остальной части головы, как прическа Мардж Симпсон. В отличие от рогов носорога, непарнокопытного лавразиатерия, рога арсиноитерия были не роговыми, а костными, причем они были полыми и торчали вперед под таким углом, что явно заслоняли животному обзор при ходьбе. Может быть, арсиноитерию это не особо мешало, так как столь массивному травоядному вряд ли угрожало много хищников. Трудно представить себе для этих нелепых рогов какое-то иное назначение, кроме как суперкомпенсаторный способ привлечения партнеров и запугивания соперников.

Чудовищный вымерший афротерий Arsinoitherium.
Фотография Арама Дуляна
Затем, конечно, слоны. Образцовые афротерии, и эта группа еще больше, чем даманы, ламантины и арсиноитерий, подходит под определение «жалкие остатки былого величия».
Как и все великаны, слоны когда-то были маленькими и невзрачными. Последовательность переходных ископаемых форм из фосфатных карьеров Марокко, которую изучали французский палеонтолог Эммануэль Гербрант и его марокканские коллеги, показывает, как слоны увеличились в размерах. Древнейший из этих ископаемых слонов, эритерий (Eritherium), жил в середине палеоцена, около 60 млн лет назад. Жалкое зрелище: при росте 20 см и весе 5 кг его зашугала бы любая комнатная собачка, а современный слон расплющил бы в блин, наступив ногой. Однако его коренные зубы уже начинали обнаруживать признаки слоновых: поперечные гребни, соединяющие бугорки. Эти так называемые лофы проходят от щечной стороны к язычной и придают жевательной поверхности зуба гофрированную форму, идеально подходящую для измельчения растений.
Палеоцен сменился эоценом, период глобального потепления ПЭТМ начался и закончился, а тем временем карликовые марокканские слоники подросли, и зубные лофы у них стали заметнее. Сначала появился фосфатерий (Phosphatherium), втрое крупнее эритерия и с полноценными рифлеными зубами. Потом дауитерий (Daouitherium), первый по-настоящему крупный представитель слоновых, – он достиг веса в 200 кг. Когда 55 млн лет назад он прогуливался по лесам – тогда еще не превратившимся в саванны и степи, – он был на тот момент самым крупным африканским млекопитающим в истории. У обоих видов резцы выдавались вперед – предвестие будущих бивней, которые появятся на следующей стадии эволюции у таких видов, как алжирский нумидотерий (Numidotherium), достигавший в высоту метра и весивший 300 кг. Как раз на этом этапе слоны приобрели свой высокий лоб, а ноздри у них сместились назад ради крепления маленького хоботка, как у тапира. Еще немного поколдовав, эволюция вырастила из хоботка хобот. Пусть это выглядело забавно, но именно хобот все изменил: он позволил слонам добывать пищу и воду, не двигая всем телом, что дало им возможности роста до огромных размеров.
И они росли. К началу олигоцена, около 34 млн лет назад, виды наподобие палеомастодона (Palaeomastodon) переросли современных африканских лесных слонов (самый мелкий из ныне живущих видов), достигнув веса в две с половиной тонны. Верхние бивни палеомастодона были обращены вниз, а нижние торчали из челюсти горизонтально, выдаваясь за верхние и образуя причудливый прикус. Это была лишь одна из разновидностей форм бивней, которые разовьются у слоновых в олигоцене (34–23 млн лет назад) и следующем за ним миоцене (23–5 млн лет назад). Одни виды обладали лопатообразными нижними бивнями, другие – комплектом длинных верхних и нижних бивней, торчавших изо рта, словно гигантские пинцеты, а у вида под названием «динотерий» (Deinotherium) верхние бивни исчезли, зато нижние загнулись вниз и стали походить на открывалку для бутылки. Это разнообразие форм, вероятно, имело двойное назначение: оно позволяло различным видам специализироваться на разных растениях – например, загнутые бивни динотерия могли служить для того, чтобы пригибать ветки деревьев, – и использовалось для демонстрации силы или привлекательности перед стадом.
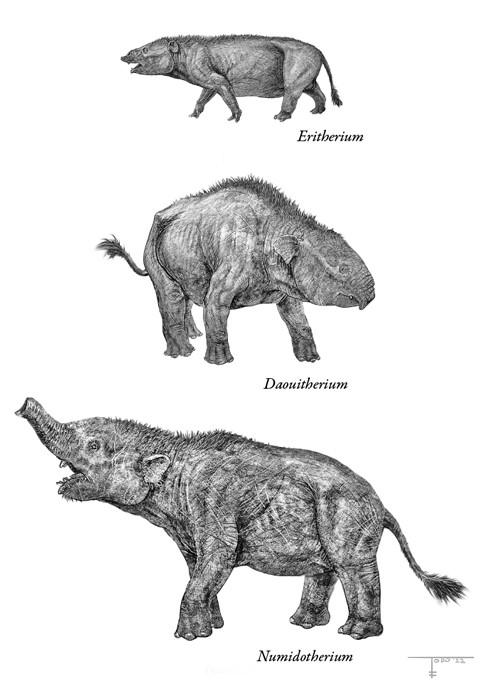
Последовательность эволюции слоновых.
Рисунок Тодда Маршалла
Некоторые из миоценовых слонов стали настоящими гигантами и превзошли по размерам все современные виды, а также распространились за пределы Африки – как раз тогда Аравия столкнулась с Евразией. Динотерий достигал четырех метров в холке и весил до 14 т, то есть был вдвое тяжелее африканского лесного слона. Но даже динотерий не был самым большим из тогдашних слонов. Эта честь принадлежит более позднему виду – палеолоксодону (Palaeoloxodon), известному по обломкам костей невероятных размеров – они указывают на пятиметровый рост и вес около 22 т. Если эти цифры верны – оценки размеров живого тела по отдельным костям всегда ненадежны, то палеолоксодон был самым огромным млекопитающим, когда-либо жившим на суше. Он потягался бы за первенство с млекопитающим, которое учебники называют самым большим в истории планеты, – безрогим носорогом индрикотерием (Paraceratherium), жившим на границе эоцена и олигоцена. Считается, что индрикотерий достигал 4,8 м в высоту и весил 17 т.

Вымершие слоновые: Palaeomastodon (вверху) и Deinotherium (внизу).
Фотографии Египетского геологического музея и Alexxx 1979 соответственно
В конечном итоге неважно, кому принадлежит первенство: и слоны вроде палеолоксодона, и носороги вроде индрикотерия были великанами. Они, вероятно, были в целом аналогичного размера, что говорит о более общей тенденции в эволюции млекопитающих: на эоцен-олигоценовом рубеже 34 млн лет назад наземные млекопитающие достигли максимальных размеров в своей истории, и с тех пор за первенство борются носороги и различные группы слоновых. Следовательно, существует некий предел общего размера тел наземных млекопитающих, что, вероятно, диктуется сочетанием факторов. Первый – питание: самые массивные млекопитающие всегда травоядные и обычно в десять раз крупнее хищников, обитающих рядом с ними. Чтобы стать сказочным гигантом, нужно иметь стабильный источник калорий, и лучший выход – налегать на растения, которые гораздо доступнее, чем мясо. Второй связан с температурой: крупным животным грозит перегрев, и чем больше они вырастают, тем острее проблема. Вероятно, размеры палеолоксодона и индрикотерия приближались к функциональному пределу. Став еще крупнее, млекопитающие, возможно, не смогли бы получать достаточно пищи или достаточно быстро избавляться от излишков тепла. Могут быть и другие факторы ограничения. Но если мы найдем хотя бы одну кость еще более крупного наземного млекопитающего, нам придется пересмотреть эти соображения.
Вы можете задаться другим вопросом: почему наземные млекопитающие не доросли до размеров динозавров? Хотя палеолоксодон и индрикотерий были огромными, они не достигали даже половины массы типичных завропод. Эту головоломку разрешить непросто, но я предполагаю, что она связана с дыханием. Легкие млекопитающих работают в две фазы, расширяясь и сжимаясь: вдох – выдох. Мы ощущаем это всякий раз, когда при дыхании грудь у нас вздымается и опускается. Птицы устроены иначе: воздух у них проходит сквозь легкие только в одном направлении. Это чудо механики действует благодаря воздушным мешкам, которые соединяются с легкими и гонят сквозь них воздух в строгой последовательности. Когда птица вдыхает, часть богатого кислородом воздуха проходит напрямую через легкие, а остаток закачивается в воздушные мешки. Затем, когда воздушные мешки сжимаются, все еще сохраняющий в них кислород воздух проходит через легкие на выдохе, то есть птицы – как и гигантские динозавры с тем же строением легких – усваивают кислород и на вдохе, и на выдохе. Это означает, что динозавр с каждым вдохом получал больше кислорода, чем млекопитающее того же размера. Более того, воздушные мешки располагаются по всему телу, даже в костях, – они одновременно действуют как система кондиционирования воздуха и облегчают скелет. В итоге получается, что крупные динозавры эффективнее дышали, им было проще охлаждаться, а их скелеты были легче и маневреннее. Вот почему, я полагаю, ни одно наземное млекопитающее не сумело приблизиться к их титаническим размерам.
Хотя они и не достигают динозавровых габаритов, современные африканские и индийские слоны – по любым объективным меркам – все-таки очень большие. Все их строение тела приспособлено к таким размерам. Их ноги, словно греческие колонны, служат опорами для массивного туловища. Смешные уши-лопухи – охладительные панели, помогающие избавляться от избытка тепла. Традиционный набор зубов млекопитающего у слонов упростился до комплекта из удлиненных бивней (резцов) и коренных зубов размером с башмак, таких больших, что единовременно в каждой челюсти помещаются лишь один-два. Поэтому слонам потребовался совершенно новый способ роста зубов – так называемое серийное замещение. Их челюсть работает как лента конвейера: новые коренные зубы прорезаются позади и постепенно смещаются вперед. Изнашиваясь при жевании, они выпадают спереди, и на смену им приходят новые зубы сзади. Все ради того, чтобы съедать много растений – по центнеру ежедневно – для прокорма такого гиганта. Размер имеет значение: прожорливые слоны способны выворачивать с корнями деревья и обращать саванны в степи, а в поисках воды они выкапывают колодцы, которые становятся центрами жизнеобеспечения новых мини-экосистем.
Однако не стоит принимать слонов за тупоголовых голиафов. Самое замечательное в них то, что они сочетают силу с умом. Их мозг велик в абсолютном выражении, что неудивительно для огромного животного. Но он велик и в относительном выражении: объем слоновьего мозга в сравнении с телом попадает в диапазон приматов. Наряду с приматами и китообразными слоны входят в Клуб Знатоков – млекопитающих с самым большим размером мозга относительно массы их тела, способных на многие интеллектуальные достижения. У слонов исключительно развитая долгосрочная память, они способны изготавливать орудия с помощью хобота, демонстрировать сложное общественное поведение и умение решать задачи, а также распознавать себя в зеркале. Они общаются друг с другом на дальних расстояниях, используя либо низкочастотные инфразвуковые вокализации, либо сейсмическую коммуникацию: они разговаривают, устраивая миниатюрные землетрясения! Возможно, полагают биологи, слоны проявляют даже некую форму эмпатии, они беспокоятся за больных или умирающих собратьев по стаду и проявляют интерес к костям своих предков и родичей.
При всех своих умениях, однако, кое-чего слоны не могут. Хотя Дамбо, герой голубого экрана, летал по небу, взмахивая ушами, настоящие слоны Африки и Азии явно неспособны к полету. Как и любое другое млекопитающее в истории планеты, за исключением одной группы – рукокрылых.
Когда я писал диссертацию в Нью-Йорке, я работал в Американском музее естественной истории, на западной стороне Центрального парка. Я приходил туда ради динозавров: в этом музее хранится одно из крупнейших в мире собраний, включая ряд знаменитых скелетов тираннозавров. По пути в динозавровые запасники, в коридорах с высокими сводами и деревянными шкафами вдоль стен, я не раз встречал Нэнси Симмонс.
В то время я был мало знаком с Нэнси и, наверное, за без малого пять лет своей работы в музее разговаривал с ней всего пару раз. Сейчас я об этом жалею, ведь теперь, когда мои интересы все больше смещаются от динозавров к млекопитающим, она стала одним из моих научных кумиров.
Нэнси тоже палеонтолог, хотя и по совместительству. Ее научная карьера началась в аспирантуре, когда она стала одним из ведущих мировых специалистов по многобугорчатым, тем самым млекопитающим с торчащими зубами, которые жевали цветочки под ногами у тираннозавров в меловой период. Затем она неожиданно сменила курс и стала ведущим мировым специалистом по рукокрылым. За несколько месяцев до того, как я приступил к своей диссертации, она опубликовала сенсационное открытие, удостоившееся заметки в Nature, – это был онихониктерис (Onychonycteris), древнейшая и самая примитивная в мире летучая мышь из раннего эоцена, жившая около 52,5 года назад. Но в основном Нэнси занята не описанием древних костей – она пробирается сквозь джунгли Юго-Восточной Азии и неотропиков в поисках ныне живущих рукокрылых, собирает образцы крови и других тканей летучих мышей, чтобы добыть ДНК и построить их родословные деревья.
Летучие мыши в представлении не нуждаются. Нравятся ли они вам или пугают вас, они общеизвестны как единственные млекопитающие, способные к настоящему полету. Некоторые другие млекопитающие умеют парить или скользить в воздухе на кожных перепонках, например летяги и шерстокрылы (их неправильно зовут летучими лемурами, но они не приматы) или вымершие харамииды юрского и мелового периодов вроде той засекреченной окаменелости, которую я видел в китайской мастерской несколькими главами раньше. Но среди млекопитающих только рукокрылые освоили машущий полет: активно взмахивая крыльями, они создают подъемную силу и тягу, необходимую, чтобы передвигаться в воздухе.
Машущий полет – непростое дело, вот почему эволюция сумела создать его всего три раза за всю историю позвоночных. Каждый случай был отдельным экспериментом по подъему в воздух. Птеродактили удлинили безымянный палец для крепления гигантского кожистого паруса. Динозавровые предки птиц удлинили переднюю конечность целиком и отрастили на ней крыло из перьев. Рукокрылые, наоборот, удлинили большинство пальцев, и получилась рука-крыло, которой они обязаны своим названием. Крыло летучей мыши – хитроумная конструкция: натянутая между пальцами кожа тонкая и гибкая, взмах происходит при сокращении больших мышц, прикрепленных к грудине. Это обеспечивает мышам скорость полета – по некоторым измерениям, до 160 км/ч – и способность плавно огибать препятствия, что весьма кстати для животных, ведущих преимущественно ночной образ жизни.
Полет – сверхспособность рукокрылых. Он дает доступ к средам обитания и видам пищи, недоступным для наземных млекопитающих, и, безусловно, исчерпывающе объясняет нынешнее изобилие и разнообразие летучих мышей. Каждый пятый вид ныне живущих млекопитающих относится к рукокрылым – в общей сложности около 1400, и по разнообразию их опережают лишь грызуны. Виды рукокрылых не только многочисленны, но еще и прекрасно уживаются вместе: в тропиках одну и ту же экосистему может населять свыше сотни видов. А некоторые из этих видов отличаются огромной численностью. Одна из причин, почему летучие мыши навевают ужас, – их склонность жить густонаселенными колониями, где гнездится бессчетное множество особей, которые висят вниз головой, уцепившись лапками, в пещерах или под мостами, сгрудившись так, что издали они кажутся грандиозным ковром и их животную природу выдает только вонь и обилие капающего помета. Я навсегда запомню свой визит в одну из таких колоний под мостом Конгресс-Авеню в Остине, штат Техас, где полтора миллиона летучих мышей разом вылетает в сумерках, плавно взмахивая крыльями, на вечернюю охоту за насекомыми. Это было биологическое представление, какого я больше никогда не видал. Но именно из-за своего коммунального образа жизни рукокрылые сильно подвержены инфекционным заболеваниям, а их колонии печально известны как инкубаторы вирусов, которые затем передаются людям.

Ископаемая летучая мышь Onychonycteris, описанная Нэнси Симмонс.
Фотография Мэтью Диллона
Как летучие мыши обрели свою сверхспособность летать? Удивительно, но мы очень мало знаем об эволюции их крыльев и полета. Из ДНК-теста на отцовство нам известно, что рукокрылые – представители северной группы лавразиатериев и на родословном древе близки хищным (кошкам и собакам) и копытным (парнокопытным и непарнокопытным). Излишне пояснять, что летучая мышь совершенно не похожа на собаку или лошадь, а значит, должна была существовать переходная последовательность вымерших видов, развивавшихся от наземного млекопитающего с ходячими конечностями к рукокрылому летуну. Беда в том, что у нас не хватает окаменелостей, запечатлевших эту эволюционную трансформацию. Скелеты первых известных летучих мышей появляются с эоцена, как открытый Нэнси Симмонс онихониктерис, и это уже настоящие летучие мыши. У них узнаваемые очертания эмблемы Бэтмена: маленькая головка, компактное туловище, коротенький хвостик и широкие крылья, растянутые на передних конечностях. Это были почти невесомые зверьки, чьи хрупкие скелеты могли сохраниться только при исключительном стечении обстоятельств. Онихониктерис, например, был погребен в спокойных глубинах озера в штате Вайоминг, и так же происходило захоронение летучих мышей из более молодых эоценовых сланцев немецкого озера Мессель. Предки этих, к счастью, сохранившихся эоценовых рукокрылых, вероятно, были еще мельче и еще больше подвержены разрушению после смерти, поэтому нам не повезло обнаружить их ископаемые остатки… пока.
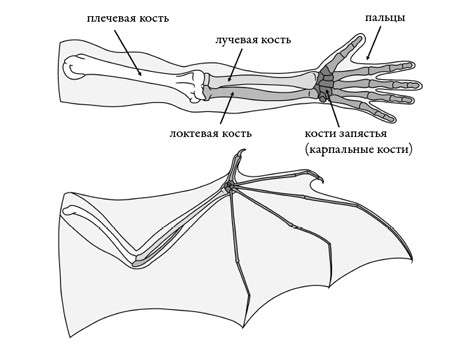
Крыло летучей мыши и рука человека.
Рисунок Сары Шелли
Хотя мы все еще не располагаем ископаемыми образцами реальных предков летучих мышей, впервые отрастивших крылья и поднявшихся в воздух, по находкам вроде онихониктериса и других ранних рукокрылых можно сделать кое-какие выводы. Описывая онихониктериса, Нэнси и ее коллеги отметили, что он намного примитивнее современных видов. Конечно, он обладал руками-крыльями и большим гребнем на грудине для крепления махательных мышц, следовательно, умел летать. Но крылья у него были непривычной формы – короткие и толстые, непохожие на более широкие и изящные крылья большинства современных летучих мышей. А значит, онихониктерис был не столь маневренным, и ему требовалось лететь очень быстро, чтобы обеспечивать достаточную подъемную силу. Полет у него, вероятно, тоже был странным – он то махал крыльями, то парил, трепыхаясь, словно пьяная бабочка. Были у него и другие анатомические странности. Что любопытно, если не смотреть на крылья, то по пропорциям тела онихониктерис напоминал типичное планирующее животное вроде летяги. Кроме того, на всех пальцах у него имелись острые загнутые коготки – тогда как у современных рукокрылых на пальцах крыльев когтей нет, – а значит, он был ловким древолазом, цеплявшимся за ветки всеми четырьмя конечностями.
Если сложить все эти подсказки вместе, они указывают на то, что рукокрылые произошли от древесного планирующего предка, который перешел к машущему полету, удлинив пальцы передних лап и обратив их в крылья. Новообретенное крыло совершило переворот: по сравнению с кожными перепонками планирующих животных оно обладало вдвое большей площадью для обеспечения подъемной силы и тяги и таким образом давало возможность более длительного и точного полета. И все же первые активные полеты неминуемо не могли не быть неуклюжими. Аэродинамическая эффективность онихониктериса была ниже, чем у современных рукокрылых, – он тратил больше усилий, чтобы обеспечить подъемную силу, и не умел так ловко облетать ветки и другие препятствия. Но Рубикон был перейден, мыши взлетели, и с этого момента естественный отбор непрерывно совершенствовал их полет, наращивая ширину крыльев, дающих подъемную силу, увеличивая дальность полета и маневренность.
Это правдоподобная версия, и она убедительна в свете того, что нам известно сейчас, но все-таки нам не хватает ископаемых переходных видов – заполняющих промежутки между наземным образом жизни, планирующим полетом и машущим, – чтобы доказать ее.
Какими бы путями рукокрылые ни приобрели свою рукокрылость и способность к машущему полету, очевидно, что стоило им полететь, как они быстро распространились по всему миру. К концу раннего эоцена, всего через несколько миллионов лет после периода ПЭТМ, рукокрылые оставляют свои окаменелости в Северной Америке (тот же онихониктерис), Европе и Африке. Ископаемые остатки появляются даже в Австралии, уже тогда изолированной на краю света, и в Индии, еще не соединившейся с Азией. Рукокрылые стали первой космополитической группой плацентарных млекопитающих – первыми, кто преодолел оковы географии, до сих пор диктовавшие пути эволюции палеоценовых и эоценовых видов. Почему, не секрет. Рукокрылые умели летать, а значит, без труда могли пересекать водные барьеры, которые затрудняли или делали вовсе невозможным расселение наземных млекопитающих. Они, как инвазивная орда, распространялись по земному шару с такой скоростью, что достигли Южной Америки и Африки – двух крупнейших островных континентов – раньше, чем там успели возникнуть эндемичные летающие формы. К сожалению, именно по этой причине, по-видимому, не существует экзотических рукокрылых сумчатых или рукокрылых афротериев – все рукокрылые принадлежат к лавразиатериям.
Распространяясь по планете – в наши дни рукокрылые заселили все континенты, кроме Антарктиды, и практически каждый клочок земли, не покрытый ледниками, – они при этом диверсифицировались по размерам, форме крыльев, способам полета, питанию и экологическим нишам. Около 48 млн лет назад, в среднем эоцене, сотни летучих мышей падали в ядовитый ил озера Мессель, и их изумительно сохранившиеся отпечатки демонстрируют немалое разнообразие. Известно семь видов, и форма их крыльев и окаменевшее содержимое желудков свидетельствуют о том, какой разный образ жизни они вели. Одни, с узкими крыльями, высоко парили в открытом небе над озером, другие – ширококрылые лихачи – носились среди густых ветвей среднего яруса леса, третьи, с переходными формами крыльев, летали в прогалинах между деревьями. Одни виды питались мотыльками и прочими летающими насекомыми; другие собирали с веток жуков и менее подвижных букашек.
Все рукокрылые Месселя, по-видимому, обладали еще одной сверхспособностью большинства современных видов – эхолокацией. Это мощный биологический сонар, но совершенно не сравнимый ни с чем в нашем собственном сенсорном репертуаре. Летучие мыши испускают высокочастотный писк горлом или щелкают языком, прислушиваются к эху и создают у себя в мозгу «картину» звукового ландшафта. Таким образом они получают возможность «видеть» в темноте – это шестое чувство, помогающее обнаружить затаившихся хищников, съедобных насекомых и ветки деревьев, чтобы не врезаться их. Летучим мышам, использующим при эхолокации горло, необходимы две вещи: во-первых, большая спиральная улитка в ухе, чтобы слышать эхо; во-вторых, надежное соединение между гортанью и ухом, которое обеспечивает так называемая шиловидно-гиалиновая кость, окружающая барабанное кольцо, – таким образом, нервная система может сопоставлять издаваемый писк с входящим эхом, а кроме того, эта кость обеспечивает крепление мощных гортанных мышц, необходимых, чтобы пищать. Эти характерные анатомические признаки наблюдаются на ископаемых остатках, поэтому нам известно, что летучие мыши из Месселя обладали способностью к эхолокации, а онихониктерис нет. Следовательно, эхолокация появилась позже, чем полет.
Как и в случае с полетом, первые опыты эхолокации были скромными. У некоторых летучих мышей из Месселя улитка была ненамного больше, чем у видов, неспособных к эхолокации, и, вероятно, их сонар работал с низким разрешением – он использовался для приблизительной ориентации и избегания крупных препятствий. Со временем эта система получила тонкую настройку путем естественного отбора: приставка «Денди» 1990-х, с примитивной графикой и ограниченными возможностями, превратилась в современную игровую консоль. Увеличенная улитка позволила более прогрессивным видам рукокрылых четче «видеть» в темноте и не просто избегать препятствий, а активно использовать свой сонар для охоты на насекомых в полете. Ныне рукокрылые – одни из самых умелых воздушных охотников, способные услышать шелест крылышек насекомого в кромешной тьме, точно определить его местонахождение и схватить его на лету. Эхолокация стала судьбой летучих мышей, их билетом в ночное небо, а птицы, которые еще раньше произошли от динозавров, за исключением немногих видов, так и не приобрели эхолокацию – не смогли завоевать ночные ниши.
Не все рукокрылые, способные к эхолокации, используют ее для поиска насекомых; некоторые с ее помощью разыскивают другие виды пищи. Самые одиозные из них – это летучие мыши, которых боятся больше всего: вампиры, действительно пьющие кровь. Три вида вампиров, обитающие в Центральной и Южной Америке, – единственные млекопитающие, промышляющие исключительно кровью. Этот необычный рацион называется гематофагией. Трудно представить себе более жуткое создание природы, чем эти мыши: испустив пару эхоимпульсов, они находят своих спящих жертв, а их мозг настроен на обнаружение ритмов дыхания. Они бесшумно подлетают к ничего не подозревающей жертве – разумеется, в потемках, – приземляются рядом с ней, медленно подползают на четвереньках, определяют с помощью теплового сенсора в носу место на коже, где близко проходят кровеносные сосуды, а затем вонзают свои острые зубы и слизывают языком текущую кровь. Попойка продолжается около получаса, причем они стараются не выпускать слишком много крови, чтобы хозяин остался жив и обеспечил следующую кормежку. Обычно жертвами становятся птицы, коровы и лошади, но зафиксированы и случаи нападения вампиров на людей. Как будто они и так недостаточно страшные, вампиры селятся колониями, в которых сотни и тысячи особей свисают с потолков пещер в дневное время, а ночью вылетают кормиться. За год колония в сто особей выпивает объем крови как у 25 коров.
Встречались ли среди эоценовых рукокрылых кровососы? Мы не уверены, и это один из множества вопросов эволюции этих животных, которые еще остаются открытыми. Если вы молодой палеонтолог и хотите прославиться, попробуйте отыскать кости рукокрылых палеоцена или самого начала эоцена. И если у вас получится, я хотел бы предложить вам назвать какое-нибудь из них в честь Нэнси, как наша команда в Нью-Мексико официально назвала «первобытного бобра» – открытого Кариссой Рэймонд в начале этой главы – Kimbetopsalis simmonsae, в знак признания ранних исследований Нэнси по многобугорчатым, до того, как она переключилась на летучих мышей. Пока переходные окаменелости не покажут, как летучие мыши перешли от наземного образа жизни к планированию и полету, все еще остается немало тайн. Но есть еще одна группа млекопитающих, претерпевшая другую, но столь же удивительную трансформацию, и в этом случае ископаемые остатки пошагово демонстрируют нам, как это происходило.
Пирамиды Гизы высятся уже 4000 лет, опаляемые солнцем Сахары и обдуваемые ветрами. Как и многие египетские памятники, они построены из прочного материала – известняка, чья родословная намного древнее фараонов. Эти твердые, богатые кальцием отложения образовались в теплых спокойных водах моря Тетис более 40 млн лет назад, в эоцене. Существа, жившие в этом забытом мире – задолго до того, как Северная Африка стала вначале сушей, затем пустыней, а затем и колыбелью человеческой цивилизации, – оставили свои окаменелые отпечатки. Сами блоки пирамид в Гизе изобилуют частицами водорослей, раковинками микроскопического планктона и окаменелостями моллюсков. А примерно в 200 км к юго-западу, возле пальмового Файюмского оазиса, орошаемого Нилом, из эоценовых слоев выступают куда более крупные ископаемые остатки.
В переводе с арабского Вади-аль-Хитан означает «Долина китов». Это не метафора. Пустынная почва усыпана китовыми скелетами, словно эоценовое морское дно вывернулось на сушу и окаменело. Эта сцена вызывает ощущение диссонанса, неправдоподобия: тысячи китов на песке, под палящим солнцем, в двух сотнях километров от ближайшего моря, где сейчас плавают киты. Киты без воды кажутся столь же невообразимыми, как если бы они оказались на Луне среди кратеров. Многие скелеты в отличном состоянии, учитывая их древность: огромные кости сохранились полностью сочлененными, зубастые черепа соединены с плавно изогнутыми позвоночниками, по бокам торчат ребра. Если обойти змеиные извивы туловища, можно увидеть отходящие от плеч уплощенные ласты, затем спинные позвонки перейдут в хвостовые, а затем в месте, где хвост сужается, покажутся маленькие косточки, отделенные от остального скелета.
Таз и нога.
Странно. Ни у одного из современных китов нет задних ног, да они им и не нужны: они используют передние ласты, чтобы рулить, а двулопастный хвост – чтобы плавать.
Скелеты из Вади-аль-Хитан – принадлежащие 15-метровому базилозавру (Basilosaurus), его меньшему собрату дорудону (Dorudon) и ряду других видов – это не обычные киты. Они напоминают о временах, когда киты ходили. Хотя сами виды из Вади-аль-Хитан обитали в океане, они еще сохраняли ноги, унаследованные от наземных предков, которые за 10 млн лет в эоцене перебрались в воду, видоизменили свое тело, превратившись из длинноногих бегунов в обтекаемые подводные лодки, и уже никогда не возвращались на твердую почву. При этом они продолжали увеличиваться в размерах и все больше приспосабливались к морю, постепенно теряя части тела, характерные для наземных млекопитающих, и приобретая те, что необходимы для полностью водного образа жизни.

Ископаемые скелеты китов в египетской пустыне, долина Вади-аль-Хитан.
Фотографии Ахмеда Мосада (вверху) и Мохаммеда али Муссы (внизу)
Это образцовый пример крупного эволюционного изменения, вошедший во все учебники биологии, – превращение одного типа организма в другой, который выглядит и ведет себя совершенно иначе, с перестройкой тела в соответствии с новым образом жизни. Это не гипотеза; мы располагаем реальной последовательностью ископаемых скелетов, включая базилозавра и дорудона, демонстрирующих поэтапные метаморфозы китов. Если кто-нибудь попытается утверждать, что «переходных форм» или «недостающих звеньев» в палеонтологической летописи не существует, расскажите ему о ходячих китах.
Прежде чем углубиться в историю китов, давайте отбросим лишнее и отметим очевидное. Кит похож на рыбу. Принять его за гигантскую рыбину простительно – я уже не первый год учился в школе, когда наконец уразумел, что киты – млекопитающие. Они похожи на рыб потому, что конвергентная эволюция приспособила их строение к сходному образу жизни – плаванию, питанию и размножению в воде. То есть киты не похожи на других млекопитающих потому, что во время эоценового перехода от наземного к водному образу жизни многие черты и повадки стереотипного «млекопитающего» у них исчезли или перепрофилировались. Среди млекопитающих киты самые немаммальные – и все же они млекопитающие. При ближайшем рассмотрении это видно. У них одна кость нижней челюсти и три слуховые косточки, а эти анатомические признаки как раз и определяют млекопитающих; у них есть молочные железы, и они выкармливают потомство молоком; и хотя у них гладкая кожа, рудименты волос сохраняются – в виде вибрисс вокруг пасти (у некоторых видов они присутствуют только в детстве). Более того, киты – плацентарные млекопитающие: они рожают крупных – нередко очень крупных – и хорошо развитых детенышей, и плод получает питание от плаценты. Если вы не верите, доказательством тому служит их пупок – место, где прикрепляется пуповина, отходящая от плаценты, в утробе матери.
Итак, киты – млекопитающие, но какие именно? Иными словами, от каких наземных млекопитающих они произошли? Это озадачивало мыслителей на протяжении тысячелетий. Аристотель признавал, что киты – не рыбы, но в ту пору, задолго до появления эволюционной теории, он и помыслить не мог о том, что они произошли от других видов. Дарвин, которому изредка случалось давать маху, допустил, что киты могли произойти от медведей, которые плавали с открытым ртом, собирая насекомых с поверхности воды, – идея столь нелепая, что из более поздних изданий «Происхождения видов» он ее убрал. Джордж Гейлорд Симпсон, создавая свое знаменитое родословное древо млекопитающих в 1945 г., замял вопрос, выделив китам особую ветвь, далеко отстоящую от других групп. Наконец, во второй половине XX в. в палеонтологической летописи материализовались многообещающие кандидаты в предки китов – «архаические» плацентарные палеоцена, а именно мезонихиды, чьи толстые острые зубы напоминали зубы ископаемых китов. Эта гипотеза, впрочем, была неубедительна, так как родственные связи, выстроенные на основании сходства зубов, часто бывают ненадежными из-за конвергентной эволюции.
Настоящий ответ появился в конце XX столетия. Загадку разрешили одновременно палеонтологи и генетики, причем те и другие на этот раз – в отличие от множества других случаев, которые уже упоминались выше, – пришли к одному и тому же выводу. Киты – парнокопытные.
Вначале анализ ДНК показал, что киты попадают на родословном древе в один кластер с коровами, овцами, гиппопотамами, верблюдами, оленями, свиньями и другими парнокопытными травоядными. Затем это подтвердили ископаемые. В 2001 г. у нескольких скелетов примитивных ходячих китов эоцена обнаружили самый характерный признак парнокопытных: двойной блоковый сустав таранной кости, то есть кость щиколотки у них имеет по желобку на обоих концах. Как мы знаем из предыдущей главы, это уникальное строение щиколотки парнокопытные приобрели еще в начале эоцена, на пике периода ПЭТМ, когда они только появились, это была адаптация к быстрому бегу, чтобы не подворачивались щиколотки. Ни у одного другого млекопитающего, включая таких быстрых бегунов, как лошади и собаки, этой адаптации нет. Конечно, нет ее и у современных китов, у которых щиколотки исчезли без следа. У древних китов, находившихся в процессе перехода к водному образу жизни и уже почти утративших ноги, она оставалась только потому, что те унаследовали ее от своих парнокопытных предков. Как и наш аппендикс, подобное строение щиколотки некогда было функционально, а затем функция была утрачена, но особенность сохранилась. Для науки – удача, ведь, в отличие от ДНК, таранную кость можно пощупать руками и она немедленно убедила даже самых скептичных палеонтологов, что киты относятся к парнокопытным.
Здесь возникает следующий вопрос: от каких именно парнокопытных произошли киты? Отчасти ответ дает генетический анализ: ближайшие современные родственники китов – гиппопотамы. Но гиппопотамы не очень-то похожи на китов – вы можете себе представить, как выглядел общий предок синего кита и речного гиппопотама? К тому же древнейшие гиппопотамы жили в миоцене, спустя миллионы лет после того, как базилозавры и дорудоны уже бороздили моря в эоцене. Это значит, что гиппопотамы – не предки китов, а их двоюродные братья. Их истинными предками были переходные виды в ископаемой летописи, куда входят базилозавр и дорудон. Здесь палеонтологам раздолье: только окаменелости, а не ДНК могут рассказать о том, как киты перебрались в воду. Это рассказ о том, как Бэмби стал Моби Диком.
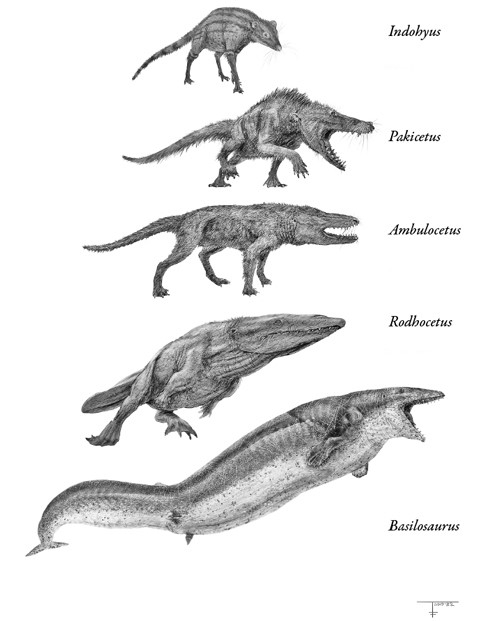
Последовательность эволюции китов.
Иллюстрации Тодда Маршалла
Эта история начинается за 10 млн лет до египетских китов, на другом берегу моря Тетис, к востоку. Индия тогда еще была островом, хотя период ее изоляции подходил к концу. Она стремительно дрейфовала через экваториальные воды на север, где ее ждало столкновение с Азией – первый акт «закрытия» моря Тетис. Примерно 50–53 млн лет назад – плюс-минус – между двумя массивами суши оставалась лишь узкая полоска тропических вод. Вскоре они сложатся, и эта область образует Гималаи – шов между двумя неподвижными блоками земной коры. Но на протяжении нескольких миллионов лет это была тихая заводь, зона освещенного солнцем мелководного морского шельфа, куда стекали реки из Индии. Это непритязательное местечко стало полигоном для одного из величайших экспериментов в эволюционной истории.
В числе многих млекопитающих, заброшенных судьбой на остров Индостан, было парнокопытное ростом с енота – индохиус (Indohyus). Это был грациозный скакунок с собачьей мордочкой и туловищем олененка, гарцевавший по лесу на длинных тонких ножках. Образ жизни он вел скромный – жевал листики и прятался от хищников. За ним мало кто мог угнаться: он удирал большими скачками, используя возможности двойного блокового сустава. Впрочем, иногда ему угрожали более ловкие хищники, не пешие, а летучие, – птицы. Но крохотное копытное млекопитающее владело одним трюком: оно могло, подобно современному африканскому оленьку, прыгнуть в реку или озеро и затаиться под водой. Этот зверек не был чемпионом по плаванию, воду он использовал как убежище, а заодно мог там пожевать водные растения, дожидаясь, пока злодей улетит. Надо признаться, эта картина – вымысел, но основанный на ископаемых находках, которые не только проливают свет на образ жизни индохиуса, но и показывают, как это тщедушное создание, хрупкое телосложение которого предельно далеко от титанических пропорций синего кита, стало китовым предком.
Первые находки индохиуса были сделаны в Кашмире – пограничном регионе Гималаев, за владение которым спорят Индия, Пакистан и Китай, – индийским геологом А. Ранга Рао и описаны в 1971 г. Они были скудные – несколько зубов и обломок челюсти, – и Рао до самой смерти понятия не имел, какое животное он нашел. Его вдова, однако, проявила упорство. Она сохранила ящики с породой из кашмирских раскопок, многие из которых оставались неоткрытыми, и переслала их голландско-американскому палеонтологу Хансу Тевиссену. Поначалу Тевиссен тоже не придал им значения… пока его лаборант случайно не расколол череп, заключенный в одном из камней. Тевиссен не поверил своим глазам: полая слуховая булла, окружающая три косточки среднего уха, загибалась раковиной, и внутренняя стенка ее была утолщена и изогнута. Возможно, какая-то косточка, примостившаяся в задней части черепа, никому, кроме анатома, ни о чем не сказала бы, но она имела огромное значение в двух отношениях.

Скелеты ископаемых китов, иллюстрирующие переход от наземного к водному образу жизни. Сверху вниз: Pakicetus, Ambulocetus, Basilosaurus, Dorudon. Масштаб 1 м относится только к двум нижним изображениям.
Фотографии Кевина Гертина, Notafly, а также из статьи Voss et al., 2019, PLoS ONE
Во-первых, генеалогически эта необычная слуховая булла связывала индохиуса с китами. Почти у всех млекопитающих булла хрупкая, иногда тонкая, как яичная скорлупа, и похожа на пузырь. Но не у китов, у которых она в форме раковины и твердая, как камень. Это такая же визитная карточка китов, как двойной блок таранной кости у парнокопытных. При отсутствии генетических данных, обусловленном тем, что невозможно получить ДНК столь древнего животного, это самое надежное доказательство семейного родства, какое только может дать анатомия.
Во-вторых, слуховая булла кое-что говорит нам об образе жизни индохиуса. У китов ее необычное строение имеет свою причину. Вспомним, что у всех млекопитающих эти полые кости – по одной с каждой стороны головы – играют роль шумопоглощающих наушников, которые изолируют среднее ухо во время передачи звука в улитку, а затем в мозг. Китам нужно слышать под водой, а это труднее, чем в воздухе. Поэтому им нужен слуховой аппарат, а утолщенная скорлупа буллы, как и ее изгиб, усиливает способность улавливать звук. Следовательно, если таким слуховым аппаратом обладал индохиус, он тоже должен был хорошо слышать под водой. Это согласуется с другими странностями находок Рао. Кости ног индохиуса были чрезвычайно плотными, с толстыми стенками и маленькими мозговыми полостями – характерный признак водных животных, которым нужен балласт, чтобы уменьшить плавучесть и оставаться под водой. А пропорции его тела – маленькая головка, крупное туловище, тоненькие ножки, длинные кисти и стопы – удивительно напоминали современного оленька, что наводит на мысль, что он вел сходный образ жизни: кормился по берегам рек и ручьев и нырял в воду в случае опасности.
Если сложить все это вместе, станет ясно, что индохиус был наземным млекопитающим, экспериментировавшим с водной средой. Это был первый шаг его длительного эволюционного пути, который отнюдь не был предопределен. Природа не задавалась целью создать китов. Эволюция так не работает: она не планирует загодя, а действует в настоящий момент, приспосабливая организмы к непосредственным задачам, с которыми они сталкиваются. Когда индохиус мчался в воду, он просто хотел спастись или искал пищу. Он и понятия не имел, что его потомки станут морскими левиафанами. Но Рубикон был перейден, и как только парнокопытные малютки окунули свои копытца в воду, естественный отбор был готов превратить их в более умелых пловцов.
Следующая задача состояла в том, чтобы сделать что-то с ногами. Ножки-спички с копытцами, какие были у индохиуса, позволяли плавать лишь кое-как. Если вы хоть раз смотрели одно из этих вирусных видео с оленем, свалившимся в бассейн, вы понимаете, о чем речь. Ответом эволюции стал пакицет (Pakicetus), следующее звено перехода от наземной жизни к водной. Его описал научный консультант Тевиссена Фил Гингрич, уже знакомый нам из предыдущей главы как специалист по периоду глобального потепления ПЭТМ. Не менее известен Гингрич как специалист по ранним китообразным, и пакицет – его главное открытие. Это был зверь размером с крупную собаку, с длинной мордой и волчьим оскалом – признак того, что от вегетарианского рациона он перешел к мясному. Но больше всего обращают на себя внимание ноги: они толще, чем у индохиуса, и лапы начинают походить на ласты аквалангиста. Пакицет все еще был по большей части пешеходом. Он жил в двух мирах – благодаря таранной кости с двойным блоком он хорошо передвигался по суше и мог ходить по мелководью пресноводных ручьев, впадавших в море Тетис. Но он также умел грести лапами и немного вилять туловищем, так что получался многофункциональный вездеход-амфибия, умевший передвигаться всевозможными способами.
И все же пакицета нельзя было назвать грациозным пловцом, и он вел двойную жизнь между сушей и рекой. Следующее звено в эволюционной цепи – амбулоцет (Ambulocetus), описанный Тевиссеном. С Индийского субконтинента он делал вылазки в соленые прибрежные воды моря Тетис. Размером с крупного морского льва, он явно был лучше приспособлен к водному образу жизни, чем пакицет, – туловище длиннее и более обтекаемой формы, ноги короче, а лапы шире и похожи на весла. Прибрежные течения он мог преодолевать двумя способами – старым и новым: либо грести лапами, как его наземные предки, либо волнообразно изгибать позвоночник вверх-вниз, чему он научился в воде. У него также появилось еще одно слуховое приспособление – жировая подушка в нижней челюсти, прилегающая к кости буллы, чтобы улавливать подводные вибрации и передавать их в ухо. Амбулоцет – как и современные киты – буквально слышал посредством челюсти. Эта эволюционная хитрость позволила обойти такую проблему, как неэффективность барабанной перепонки наземных млекопитающих под водой, в чем мы можем убедиться при попытке поболтать друг с другом в бассейне. Животному с плавательными и сенсорными способностями амбулоцета на суше было особо нечего делать. Хотя, вероятно, он все еще умел ходить – как минимум ковылять, – по образу жизни он скорее напоминал крокодила: это был засадный хищник, предпочитавший таиться на мелководье и хватать заостренными зубами рыбу.
На этой стадии эволюция взялась за животное, которое выглядело как миниатюрный олень, и сотворила достаточно крупное ластоногое плавающее млекопитающее, умеющее хорошо слышать под водой. Оно порвало связь с сушей и пресноводной средой и теперь плавало, извивалось и подстерегало добычу на прибрежном мелководье океана, омывающего Индийский субконтинент. Пути назад не было. Суша осталась в зеркале заднего вида. Впереди был океан – целиком и полностью. Не только прибрежные воды острова, но и открытые просторы и темные пучины, покрывающие 70 % поверхности Земли.
Первыми китами, распространившимися по всей планете, стали протоцетиды, типичным представителем которых был родхоцет (Rodhocetus) – еще один вид, описанный Гингричем. Они все еще оставались ходячими китами, но лишь номинально. Их лапы, особенно задние, были до смешного велики – но не потому, что они хорошо бегали. Нет, это было приспособление для плавания, и хотя протоцетиды могли передвигаться по суше, они делали это так же неуклюже, как аквалангист, вздумавший пробежаться в ластах. Вероятно, по образу жизни они напоминали тюленей – бóльшую часть времени проводили в воде, но время от времени выбирались на скалы, чтобы погреться на солнышке, спариться, родить и выкормить детенышей. Больше на суше им было делать нечего. Практически расставшись с наземной родиной предков, они направились в открытый океан. Уже 40 млн лет назад – к моменту столкновения Индии с Азией – протоцетиды населяли моря у побережий Азии, Африки, Европы и Северной Америки, а вид под названием «перегоцет» (Peregocetus) добрался до тихоокеанского побережья Южной Америки. Протоцетиды и мигрировавшие одновременно с ними афротерии в лице ламантинов стали второй волной плацентарных космополитических видов вслед за рукокрылыми. Если рукокрылые преодолели географические барьеры по воздуху, то протоцетиды – по воде, загребая своими широкими перепончатыми лапами.
А потом ходячие киты перестали ходить. Это ознаменовало их окончательное расставание с сушей. Египетские скелеты из Вади-аль-Хитан запечатлели этот решающий момент в эволюции китообразных, когда они перешли к полностью водному образу жизни и впервые стали напоминать знакомых нам китов.
У базилозавра есть несколько отличий от протоцетид. Во-первых, гигантские размеры: он достигал 17 м в длину и весил более 5 т, что на порядок больше типичных протоцетид. Чтобы перемещать свою грандиозную тушу в воде, базилозавр обзавелся новым плавательным приспособлением – хвостовым плавником, который поддерживался широкими позвонками. Волнообразные движения плавника вверх-вниз создавали тягу. Эти гибкие движения стали возможны лишь потому, что тазовый пояс и задние конечности все дальше усыхали, отделяясь от позвоночника и давая хвосту новую степень свободы. Ноги базилозавра являли собой жалкое зрелище – они были меньше человеческих, притом что корпус был длиннее многих яхт. Они все еще выступали из туловища, но вскоре киты утратят все внешние признаки их присутствия. Сохранятся лишь внутренние редуцированные косточки таза и конечностей, заслужившие пощаду на том основании, что к ним крепятся генитальные мышцы. Передние лапы базилозавра, однако, оставались выраженными – они приобрели вид плоских широких рулевых весел, – но явно не могли служить опорой пятитонному телу на суше. Шея укоротилась, слившись с туловищем в одно торпедообразное целое; в околоушных костях появились полости, чтобы регулировать давление при нырянии, а ноздри начали смещаться назад, превращаясь в дыхала. Все эти приспособления помогли базилозавру стать первоклассным пловцом. И не каким-нибудь, а грозным: он был высшим хищником, поедавшим других китов, что доказывают кости дорудона, найденные в области желудка одного из скелетов.
К концу эоцена ходячие киты исчезли совсем. Переход завершился: наземное животное стало облигатным пловцом, уже неспособным вернуться на землю – ни ради спасения от хищников, ни ради кормежки, размножения или сна. Отныне киты будут заниматься всем этим в воде. Эволюция, однако, на этом не закончилась. Она никогда не заканчивается. Примерно на границе эоцена и олигоцена, 34 млн лет назад, началась следующая фаза истории китообразных: настала пора превратить китов в самых совершенных водных животных, какими они только могли стать.
В это время киты разделились на две линии, которые привели к двум версиям современных китов: зубатым и усатым. Каждая из них обладает собственным набором анатомических признаков и форм поведения, в высшей степени приспособленных к подводной жизни. Окаменелости обеих групп начинают появляться на границе эоцена и олигоцена. С этого времени киты современного типа выходят за границы расселения протоцетид и базилозавра, распространяясь в самые высокие широты и холодные океаны. Теперь они освоили и прибрежные воды, и открытое море, и арктические, и тропические условия, и мелководье, и глубины, отважились даже забраться в пресные воды – речные дельфины (да, дельфины – тоже китообразные) и другие виды вернулись в среду обитания индохиуса и пакицета. Китовая империя стала глобальной во всех отношениях.
Среди зубатых китов – официально известных как Odontoceti – ныне числятся кашалоты, косатки, нарвалы, дельфины и морские свиньи. Это свирепые хищники, стоящие на вершине пищевых цепей в океане, и в их распоряжении три главных оружия.
Первое – их острые зубы, видоизменившиеся так, что они уже не похожи на зубы млекопитающего. Исчезли все сложные бугорки и гребни; исчезла последовательность резцов, клыков, предкоренных и коренных зубов; молочные зубы перестали меняться на постоянные; утрачена способность жевать. Все зубы превратились в конусообразные колышки, которыми зубатые киты просто откусывают мясо рыб или других китов и затем проглатывают. Некоторые виды почти не пользуются зубами, когда едят, и лениво заглатывают добычу целиком.
Второе оружие, как ни странно, роднит китообразных с рукокрылыми, хотя явно развилось у тех и других независимо друг от друга – эхолокация. Зубатые киты издают высокочастотные щелчки и свист, выпуская воздух через звуковые губы – мясистое уплотнение носовых проходов сразу под дыхалом. Жировая подушка на лбу – так называемая акустическая линза – фокусирует звуки, которые затем отражаются обратно и улавливаются специализированной улиткой уха. Если летучие мыши применяют эхолокацию для ловли насекомых или поиска жертвы, у которой можно попить крови, то зубатые киты пользуются своим сонаром, чтобы обнаруживать стаи рыб или кальмаров в темных мутных глубинах океана. Это настолько чувствительный механизм, что зубатым китам больше не нужно обоняние – и, по сути, они его утратили.

Черепа зубатого и усатого китов.
Рисунок Сары Шелли
Наконец, у зубатых китов невероятно большой мозг. У кашалота самый крупный мозг среди всех животных на Земле, возможно, за всю ее историю. Он весит 10 кг – в пять раз больше человеческого мозга и больше мозга любого слона. По соотношению размеров мозга и тела, которое служит примерным относительным показателем интеллекта, он занимает второе место после человека, опережая всех остальных животных. Кашалоты достаточно умны, чтобы перехитрить свою добычу, они используют орудия и узнают себя в зеркале.
Все эти хищные способности развились задолго до появления современных кашалотов и их родичей, их начало прослеживается эоцен-олигоценовой грание. Древнейшие ископаемые зубатые киты, как, например, котилокара (Cotylocara) и эховенатор (Echovenator) из Южной Каролины, уже обладали коническими зубами, большим мозгом и особенностями строения черепа, связанными со способностью издавать и улавливать высокочастотные звуки, – в частности, чашеобразным углублением для лобной линзы и управляющих ею мышц, а также улиткой с расширенным основанием. Значит, зубатые киты приобрели крупный мозг и эхолокацию, когда они отделились от усатых китов и быстро выработали собственный уникальный стиль охоты и мышления. С начала олигоцена и до наших дней они совершенствовали свои способности к эхолокации, наращивая лицевые мышцы, чтобы издавать звуки, и акустическую линзу, чтобы отражать их.
Иные из древних зубатых китов посрамили бы современных – даже кашалотов. Миоценовый мелвиллов левиафан (Livyatan melvillei), получивший название в честь автора «Моби Дика», бороздил Тихий океан у берегов Южной Америки 12 млн лет назад. Это был один из крупнейших хищников в истории Земли, длиной до 18 м, из которых 3 м приходилось на череп, внутри которого мог свободно разместиться человек, хотя вряд ли кому-то там понравилось бы. Его пасть раскрывалась так широко, что он легко откусил бы голову крупнейшему из когда-либо живших наземных хищников – тираннозавру. В довершение всего его тридцатисантиметровые зубы были толще железнодорожного костыля и были отлично приспособлены к тому, чтобы крушить кости добычи – ею служили другие киты, усатые. Мелвиллов левиафан, словно придуманный для низкопробного фильма ужасов, обитал в тех же водах, что и прославленная суперакула мегалодон (Megalodon), причем акула, несомненно, кита остерегалась.
Но, как бы ни были велики мелвиллов левиафан и его родичи, им далеко до размеров крупнейших усатых китов. Усатые киты, официально именуемые мистицетами (Mysticeti), включают синих китов, гладких китов, полосатиков и горбатых китов. Их скелеты выглядят карикатурно – с раздутыми черепами и тонкими беззубыми челюстями, выступающими далеко наружу, словно обод мусорного бака или баскетбольного кольца. На этих огромных челюстных костях не найти ни следа зубов, потому что усатые киты утратили предковые зубы-колышки. Их заменило приспособление, из-за которого они и получили свое название, – китовый ус, ряд тесно расположенных пластин из кератина (вещества, из которого состоят наши ногти), свисающих с нёба. С его помощью усатые киты делают то, что не умеют другие млекопитающие, – кормятся, отфильтровывая из воды мелкую живность. Представление, которое некоторые виды устраивают во время кормежки, поистине впечатляет: опустив нижнюю челюсть, кит распахивает зияющую пасть, заглатывает чудовищный объем морской воды, а затем с помощью языка и глоточной мускулатуры выдавливает воду изо рта сквозь ус, добывая в один прием тысячи (если не больше) планктонных организмов. Забавно, что крупнейшее животное в истории питается микроскопической добычей – но поглощает ее с аппетитом Гаргантюа. Ему не требуется эхолокация для поиска пищи и нет необходимости задействовать свой большой мозг для охоты. Все, что нужно, – это лавировать в толще воды и время от времени разевать рот.
Ископаемая летопись показывает, что эволюция усатых китов не была прямолинейной. У древнейших из них, эоценовых мистакодонов (Mystacodon) из Перу и льяноцет (Llanocetus) из Антарктиды, сохранялись зубы, причем одни напоминали конические колышки зубатых, а другие обладали сложным веером бугорков, расходившихся от центрального выступа. У них не было уса, и они не могли питаться как фильтраторы, но тем не менее уже стали намного крупнее других современных им китообразных. В частности, льяноцет достигал по меньшей мере 8 м в длину и был одним из самых больших китов, в миоцене этот рекорд не побили другие усатые и зубатые киты вроде мелвиллова левиафана. Следовательно, китовый ус и питание путем фильтрации появились уже после отделения линии усатых китов от зубатых, и не в них заключался секрет больших размеров усатых китов – по крайней мере, поначалу. Напротив, палеонтологические данные показывают, что первые зубастые усатые киты были кусачими, затем они утратили зубы и начали кормиться путем всасывания и уже после этого на беззубых челюстях появился ус, а с ним и новая способность к фильтрации. Как и в случае с переходом китовых предков от ходьбы к плаванию, переход от зубов к китовому усу и от кусания к фильтрации был постепенным процессом, проходившим в несколько этапов.
Но после того как появился этот способ питания, он дал усатым китам возможность еще больше увеличиться в размерах. В отличие от зубатых китов, постоянно зависящих от «урожая» кальмаров, рыбы и других китов, которыми они питаются, усатые киты располагают практически неисчерпаемыми запасами планктона, который они могут поглощать без особых энергозатрат. Они могут пассивно дрейфовать, объедаясь на шведском столе морепродуктов, особенно во время сезонных цветений планктона или в зонах апвеллинга, где поднимающиеся из глубин океана питательные вещества привлекают рои криля. Синие киты – крупнейшие из ныне живущих и когда-либо живших китообразных – относятся к усатым. Они – кульминация долгосрочной тенденции, начавшейся на эоцен-олигоценовой границе и не ослабевающей доныне, ко все большему гигантизму китообразных. Иную картину являет собой эволюция наземных млекопитающих – как мы убедились в предыдущей главе, слоны и носороги достигли максимальных размеров на эоцен-олигоценовой границе и с тех пор не увеличивались.
Значит ли это, что когда-нибудь появятся левиафаны еще крупнее синего кита? Могут ли эти экстремальные млекопитающие стать еще более экстремальными? Но это вполне можно сделать ставку… если только синие киты и их усатые родичи переживут нынешние бурные изменения климата и окружающей среды, не вымрут и найдут достаточно планктона для пропитания в океанах будущего.

