Книга: Эра млекопитающих: Из тени динозавров к мировому господству
Назад: 1 Предки млекопитающих
Дальше: 3 Млекопитающие и динозавры
2
Как стать млекопитающим
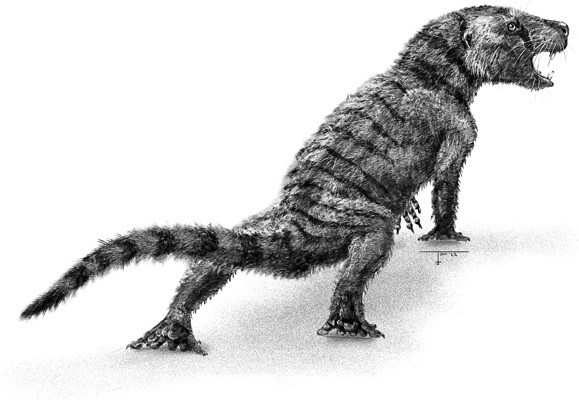
Thrinaxodon
Вдалеке прогремел гром, хлынул дождь, и зверек высунул голову из норы. Он повел носом и выставил по ветру усики. Пора выйти и надо поспешить.
Несколько месяцев назад, когда это существо размером с лисицу – тринаксодон (Thrinaxodon) – выкопало себе нору, земля была выжженной. Дождь не шел месяцами. Река почти пересохла, а от папоротников и плаунов, обрамлявших ее берега, осталась увядшая шелуха. По долине носились пылевые смерчи, засыпая стада пузатых травоядных, отчаянно пытавшихся отыскать последние съедобные листья и корешки. Иные так и оставались торчать из песчаных дюн мумифицированными трупами, чьи острые клыки придавали еще более зловещий вид этой антиутопической картине.
Уже было ясно, что пищи нет. Не видно насекомых, не пахнет вкусными земноводными, лишь иссохшая падаль. Поэтому у пушистика не было выбора – оставалось только зарыться, затаиться на время и сберегать драгоценную энергию, пока условия не улучшатся.
Теперь, словно по мановению руки, долину затопили муссонные дожди. Река вышла из берегов, и вода заливала норку, потихоньку заполняя грушевидную камеру, в которой прежде спал тринаксодон. Снаружи зеленые побеги уже пробивались сквозь ил, который вздувшаяся река разносила по дюнам, погребая жуткие мумии. Похоже, надо было начинать с чистого листа. Жизнь возвращалась, а засушливые месяцы стали далеким воспоминанием. Но в этом биполярном мире дожди долго не продержатся, и тринаксодону следовало использовать все возможности.
Для начала ему требовалось поесть, чтобы вновь запустить метаболизм. Тринаксодон поглощал пищу жадно. Прожорливость позволяла ему быстро расти и накапливать запасы энергии, необходимые, чтобы затаиться в норе и поддерживать температуру тела на стабильно высоком уровне на протяжении многомесячного оцепенения. После периода голодовки он хотел есть еще больше, чем обычно, и мечтал вонзить свои острые пильчатые зубы в панцири насекомых или в склизкую кожу мелких амфибий, кучковавшихся у реки.
Набив живот, можно было перейти к следующей важной задаче – найти себе пару. Наш тринаксодон родился меньше года назад, под конец прошлого сезона дождей. Несколько недель он жался к матери, братьям и сестрам, поедая насекомых и осваивая топографию речной долины, прежде чем начал вести самостоятельную жизнь – нашел удобный участок илистой поймы и вырыл норку, чтобы укрываться в ней, свернувшись калачиком, когда жара становится нестерпимой. Теперь, с возвращением дождей, у него будет, скорее всего, лишь один шанс спариться – единственная попытка за короткую странную жизнь, которая состоит из рождения, спячки и финальной лихорадки обжорства и размножения.
Но, по крайней мере, здесь хватало потенциальных товарищей – пойма по обе стороны реки была усеяна норками тринаксодонов. Их подземные спальни выходили на поверхность в виде небольших отверстий, напоминающих лунные кратеры. Повсюду наш тринаксодон видел, как его соплеменники высовывают головы из норок и поводят носами, как дождь стекает по их мохнатым мордочкам, как топорщатся усики, стараясь уловить, что происходит. Все размышляли над одним и тем же вопросом: вылезать или нет?
Наш тринаксодон сделал выбор. Он вывинтился из норы, поджав лапки к туловищу, чтобы протиснуться. Он ступил на илистую поверхность, вязкую и липкую, и растопырился всеми четырьмя лапами для опоры. Взглянув, как его норка заполняется водой, тринаксодон побежал прочь, в неизвестное будущее. Пища и товарищи могли подождать, а могли и нет. Как бы там ни было, все это скоро закончится.
Тринаксодон, о котором идет речь, не знал, что ему выпало жить в интересное время. Естественно, у него не хватало интеллекта, чтобы осмыслить свое место в истории живого, эволюции, планеты Земля. Но ведь и люди, живущие в интересные времена, обычно не осознают этого – они слишком заняты настоящим моментом, их волнует, что они будут есть завтра, волнуют собственная семья и множество других вещей. Мы часто не осознаем, что живем в эпоху потрясений, пока все не уляжется и мы не сможем оглянуться назад. Эти тринаксодоны, как окажется впоследствии, переживали величайший катаклизм в истории Земли – кратковременный период в несколько десятков или сотен тысяч лет, который начался с катастрофического вымирания, запечатлел неустойчивое восстановление и помог выковать класс млекопитающих из древнего племени терапсид.
Тринаксодон относится к цинодонтам, жившим около 251 млн лет назад, в самом начале триасового периода. Цинодонты входили в так называемую «стволовую линию» млекопитающих. Они были представителями группы терапсид, наряду с клыкастыми дицинодонтами (мумиями из нашей истории), бодучими диноцефалами и саблезубыми горгонопсами. Терапсиды произошли от пеликозавров, те, в свою очередь, произошли от тех «чешуйчатых зверюшек», которые в каменноугольный период разделились на линии синапсид и диапсид, чью родословную можно проследить до тетрапод, развившихся из рыб, которые выбрались на сушу и обзавелись амниотическими яйцами.
Все это мы узнали из предыдущей главы. Но жизнь гораздо древнее: рыбы происходят от первых позвоночных, ловких пловцов, которые начали укреплять свое тело костными образованиями в эпоху бурных эволюционных перемен, получившую название «кембрийского взрыва», – от 540 до 520 млн лет назад. Как раз в это время большинство ныне известных групп морских животных изобрели собственные скелеты и начали процветать – моллюски вроде мидий и гребешков, иглокожие вроде морских ежей и морских звезд, членистоногие вроде креветок и крабов. Мягкотелые предки этих животных жили в эдиакарский период, начавшийся около 600 млн лет назад, от которого остались призрачные отпечатки их бесформенных тел в песчанике. Они были первыми животными и развились из эукариотических клеток, которые смогли собраться в более крупные, более сложные многоклеточные организмы. Это произошло около 2 млрд лет назад, через 2 млрд лет после того, как возникла первая одноклеточная бактерия, а это случилось только через полмиллиарда лет после того, как Земля сформировалась из облака газа и пыли.
Жизнь – эволюционный спектакль длиной в 4 млрд лет, который сегодня, конечно, все еще продолжается. За все это время самым опасным моментом, когда жизнь оказалась на грани полного исчезновения и Земля могла стать безжизненной планетой, был переход между пермским и триасовым периодами, 252–251 млн лет назад. Это было незадолго до того, как наш тринаксодон укрылся в норе – во время мучительной фазы восстановления после катастрофы – в нынешнем южноафриканском регионе Кару.
Пермско-триасовое вымирание было крупнейшим из всех массовых вымираний и погубило около 90 % всех видов, а может быть, и больше. В отличие от большинства других массовых смертей в ископаемой летописи, этот сюжет не детективный. Убийца известен – вулканы, а точнее, мегавулканы, питаемые горячей областью магмы в глубине земной мантии под нынешней Сибирью. Тогда эта область находилась на северной окраине суперконтинента Пангея. Ничего подобного этим извержениям человечеству на своей памяти, к счастью, видеть не доводилось. Их масштаб был просто неправдоподобен. На протяжении сотен тысяч лет лава изливалась из гигантских трещин в земле – обширной сети вулканических разломов, каждый из которых достигал нескольких километров в длину и истекал лавой, словно землю располосовали гигантским мачете. Огненные взрывы сменялись затуханием, и в конце концов миллионы квадратных километров на севере Пангеи покрылись корой базальта – застывших лавовых отложений. Даже сейчас, после 250 млн лет эрозии, этот базальт покрывает около 2 млн кв. км, что примерно равно площади всей Западной Европы.
Эти вулканы нарушили покой терапсидового мира, эпохи, когда древние предки млекопитающих шествовали по тверди Пангеи. Их было великое множество видов, удивительных форм и размеров, с бивнями, клювами, бодучими куполообразными головами и кусачими клыками, поедающих столько всякой всячины и заполняющих столько разнообразных ниш – от рыкающих хищников до ценителей зелени. С точки зрения конца пермского периода незадолго до извержения первых вулканов могло казаться, что терапсиды будут господствовать и дальше, но этому было не суждено сбыться.
В позднепермский период многие терапсиды жили на территории нынешней России, неподалеку от вулканов. Горгонопсы вонзали свои сабли в дицинодонтов, были там и цинодонты, таившиеся в рощах семенных папоротников. Они стали непосредственными жертвами извержений, и многих из них, вероятно, в буквальном смысле поглотила лава, как в дешевых фильмах-катастрофах. Но они оказались не единственными погибшими – вулканы были гораздо смертоноснее, чем казались с виду. Вместе с лавой на поверхность вырвались безмолвные убийцы – ядовитые газы, такие как углекислый газ и метан. Они проникли в атмосферу и распространились по всей планете. Эти газы парниковые; они удерживают в атмосфере тепло, поглощая излучение и отражая его обратно на Землю. Они вызвали неудержимое глобальное потепление. За несколько десятков тысяч лет температуры выросли на 5–8 ℃, аналогично тому, что происходит в наши дни, – только в реальности тогдашнее потепление шло медленнее, чем нынешнее (этот факт должен заставить задуматься любого читателя). Однако этого было более чем достаточно для закисления океанов, которые остались без кислорода, что вызвало повсеместную гибель раковинных беспозвоночных и другой морской фауны.
На суше было не лучше, и самые наглядные свидетельства случившегося – кто погиб, кто выжил, как быстро наступило восстановление – находятся в Кару. С извержением вулканов и разогревом атмосферы климат Кару в начале триаса становился все жарче и суше. Времена года стали более выраженными, как и суточные колебания температур. По сути, территория Кару превратилась в пустыню, напоминающую современную, но с одним заметным исключением: временами на нее обрушивались муссоны, проносившиеся через всю Пангею. Богатые флорой пермские леса, в которых преобладали семенной папоротник глоссоптерис (Glossopteris) и вечнозеленые голосеменные, исчезли, когда растения постигло второе и последнее в их истории массовое вымирание – после кризиса каменноугольных лесов, имевшего место на 50 млн лет раньше. Их вытеснили папоротники и плауны – более низкорослые родичи древовидных лепидодендронов (Lepidodendron) из каменноугольных болот, – которые быстро прорастали из спор (а не из семян), что позволяло им легче переносить резкую сезонность и перепады количества осадков. С изменением растительности широкие, извилистые речные системы пермского периода уступили место более быстрым многорукавным рекам триасового. В отсутствие крупных деревьев, укрепляющих своими корнями берега, реки разливались по земле в дождливый сезон, но почти полностью пересыхали в сухие месяцы.
Этот экологический каскад катастрофически отразился на фауне, обитавшей в Кару, особенно на терапсидах. До вымирания они были процветающим сообществом: в основании пищевой пирамиды располагалось несколько растительноядных видов дицинодонтов, на которых охотились хищники среднего размера – биармозуховые, еще одна группа терапсид с заметными выступами и рожками на головах, – и высшие хищники горгонопсы. Немногочисленные цинодонты, такие как харассогнат (Charassognathus) – древнейший известный представитель этой группы, величиной с белку, – питались насекомыми, занимая нишу мелких позвоночных наряду с множеством рептилий и амфибий, из которых одни тоже были насекомоядными, другие – рыбоядными. Но по мере изменения климата леса отступали, и исчезло от 70 до 90 % наземной растительности. В результате вся экосистема посыпалась как карточный домик. Пищевые цепочки в начале триаса упростились – свелись всего к нескольким видам травоядных и хищников – и на протяжении следующих 5 млн лет переживали циклы подъемов и спадов, пока наконец не стабилизировались, когда извержения вулканов прекратились и температура нормализовалась.
Будь вы на месте терапсид, вам пришлось бы выбирать себе судьбу из трех вариантов. Первый – вымирание, что и произошло с горгонопсами: в триасе уже некому будет пугать добычу клыками-саблями и широко разинутой пастью. Второй – выживание, но при этом вырождение. Это случилось с дицинодонтами, которые пережили катастрофу и снова диверсифицировались, но так и не повторили своего пермского успеха; в конце концов их убогому существованию положило конец следующее массовое вымирание, триасово-юрское. И третий – выжить и занять господствующее положение. По этому пути пошли цинодонты, и чем больше испытаний они преодолевали: вулканизм, глобальное потепление, засуху, муссоны, гибель лесов, обвал экосистемы и тяготы восстановительного периода в 5 млн лет, тем сильнее становились. На протяжении всех оставшихся 50 млн лет триасового периода они продолжали диверсифицироваться, породив огромное разнообразие видов – крупных, мелких, мясоедов, вегетарианцев. Одна из этих линий цинодонтов приведет к млекопитающим, постепенно накапливая все больше «маммальных» признаков.
Почему цинодонты – наряду с некоторыми из своих родичей, дицинодонтов, – сумели выжить? Я хорошо помню, когда узнал ответ. Это было в 2013 г. в Лос-Анджелесе, на ежегодной конференции Общества палеонтологии позвоночных. Я недавно защитился и начал преподавать в Эдинбурге, а диссертацию представил на секции конференции, посвященной конкурсу имени Альфреда Шервуда Ромера. Конкурс назвали в честь легендарного гарвардского палеонтолога, руководившего той самой экспедицией в Новую Шотландию, которая обнаружила пеньки с дуплами, полными древних синапсид, – о них рассказывалось в предыдущей главе. Премия имени Ромера – главная награда для аспирантов в моей области, и я надеялся впечатлить экспертов своей работой о происхождении птиц от динозавров. Увы, премию я не получил, но досталась она более чем достойному коллеге – Адаму Хаттенлокеру. Он выступал через несколько докладов после моего и покорил аудиторию своим объяснением, почему цинодонты пережили пермско-триасовое вымирание. Когда он вернулся на свое место, я уже смирился со своей судьбой. Млекопитающие (в данном случае их предки) снова переиграли динозавров.
Адам рассказывал о любопытном эволюционном феномене, имевшем место во время вымирания и после него, – «эффекте лилипутов». Как явствует из названия, отсылающего к населенному маленькими человечками острову из «Путешествий Гулливера», речь идет об уменьшении размеров тела животных, переживающих массовое вымирание и преуспевающих впоследствии. Это происходит не всегда, но произошло в случае с цинодонтами и их близкими родичами, и это сыграло важную роль для их выживания. Адам собрал огромную базу данных по ископаемым терапсидам Кару и обнаружил заметное сокращение как максимального, так и среднего размера древнейших триасовых терапсид по сравнению с их самыми поздними пермскими предшественниками. Различие было обусловлено более интенсивным вымиранием крупных видов при извержениях вулканов и повышении температуры. Большие размеры, похоже, в эпоху нестабильности стали помехой. Таким образом, у цинодонтов, которые были мельче большинства других терапсид, оказалось больше шансов пережить хаос.
Почему маленький размер стал преимуществом? Во-первых, мелким животным легче спрятаться и переждать в норах неблагоприятную погоду, скачки температуры и пыльные бури. А норы они рыли. Отложения Кару выше слоя, связанного с вымиранием, – сланцы, образовавшиеся в пойме реки, вперемежку с мумиями, засыпанными нанесенной ветрами пылью, – изобилуют ископаемыми норами, в которых иногда находят скелеты. Это и есть скелеты тринаксодонов – зверьков из рассказа, с которого началась эта глава. Самое удивительное из этих норных захоронений содержит тринаксодона, лежащего бок о бок с небольшой раненой амфибией. У маленького родича саламандр были повреждены ребра, но они заживали, пока амфибия лежала рядом со свернувшимся в калачик спящим тринаксодоном. Так как нора тесная, а тринаксодон был зубастым хищником, странно, что другое животное могло просто лежать и выздоравливать там незамеченным. Единственное удовлетворительное объяснение – что тринаксодон находился в спячке, длившейся, возможно, неделями и даже месяцами, чтобы сберечь энергию и пережить сухой сезон.
Во-вторых, маленький размер был связан с рядом других аспектов развития и метаболизма. Дженнифер Бота, работавшая вместе с Адамом и другими коллегами, описала их в важной работе 2016 г. Древнейшие триасовые цинодонты, включая тринаксодона, росли быстро, начинали размножаться в раннем возрасте и жили очень недолго, вероятно всего года два. Откуда это известно? Ответ дает костная гистология – исследование тонких срезов костей под микроскопом. У пермских терапсид линии роста на костях обычно множественные, и это означает, что взрослых размеров они достигали за много лет. Однако у раннетриасовых цинодонтов таких отметок меньше, причем у тринаксодона их, как правило, нет вообще. Они, должно быть, росли с головокружительной скоростью и, возможно, достигали зрелости, размножались и умирали за один год. По сути, они компенсировали раннюю смерть размножением в юном возрасте. Так как ни один тринаксодон не доживал до старости, эта стратегия роста поддерживала воспроизводство вида. Чем быстрее они росли и чем раньше начинали размножаться, тем выше у них были шансы успешно дожить до брачного сезона и гарантированно передать свои гены следующему поколению в этом суровом, нестабильном мире.
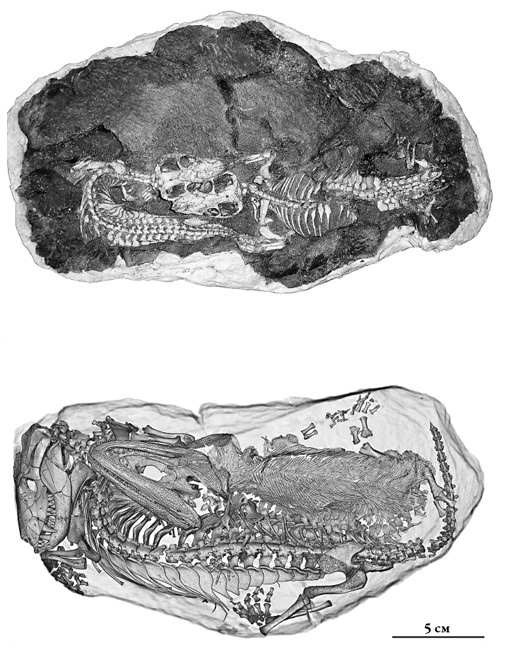
Скелеты тринаксодонов (вверху) и компьютерная томограмма тринаксодона, окаменевшего в норе рядом с амфибией (внизу).
Фотография Кристиана Каммерера и иллюстрация из Fernandez et al., 2013, PLoS ONE, соответственно
У тринаксодона и других цинодонтов, похоже, был мощный набор козырей, которые помогли им избежать вымирания. Скелеты тринаксодонов десятками начинают появляться 30 м выше горизонта вымирания, то есть они распространились по бассейну Кару за какие-то несколько десятков тысяч лет. Возможно, они произошли от пермских предков, которые редко встречались в экосистемах Кару до вымирания или, что вероятнее, мигрировали из тропических регионов Пангеи, где суровый пермский климат подготовил их к выживанию в засуху. В раннетриасовых отложениях так много скелетов тринаксодонов, что они, наряду со столь же многочисленными дицинодонтами, называемыми листрозаврами (Lystrosaurus), считаются «видами, приспособленными к бедствиям» (disaster species). Они были особенно хорошо приспособлены к тяжелым парниковым условиям мира после вымирания, оказавшимся невыносимыми для большинства других видов. По-видимому, в этих условиях они отлично себя чувствовали и процветали. Тринаксодоны и листрозавры, таким образом, были крысами и тараканами подземок раннего триаса.
Однако сравнение с современными вредителями несправедливо по отношению к тринаксодону. Он был настоящим героем, одним из немногих отважных животных, которым удалось пережить мрачную ночь самого страшного побоища доисторических времен и благодаря которым линия млекопитающих не прервалась задолго до того, как млекопитающие получили шанс развиваться.
Тринаксодон не очень походил на героя. Максимум полметра в длину, вероятно, обладавший вибриссами и по крайней мере частично покрытый шерстью, тринаксодон бóльшую часть времени проводил не на виду, укрывшись в норе. Наружу он выходил кормиться – он любил насекомых и мелкую дичь. Как и у его предков-терапсид, у него был набор резцов, клыков и щечных зубов. Отличался он тем – и за это получил свое название, означающее «трехзубчатые зубы», – что коренные зубы имели форму мультяшной горной гряды с центральным пиком, по бокам которого торчали пики поменьше. Эти три острых зубца, которые называются выступами, идеально подходили для того, чтобы прогрызать панцири насекомых и разрывать мясо. Фестончатые зубы тринаксодона менялись на протяжении всей его короткой жизни, как у рептилии или амфибии.
Но во многих отношениях тринаксодон был заметно ближе к млекопитающим, чем его терапсидные предки. При ходьбе он держал тело еще прямее, а живот выше над землей в положении полуприседа. Это позволяло ему быстрее бегать, меньше уставать и удобнее устраиваться в норе. Его позвоночный столб состоял не из одинаковых позвонков, а из отделов, одни из которых несли ребра, а другие нет, что обеспечивало ему большую гибкость и возможность сворачиваться калачиком во время спячки. Мышцы, смыкающие челюсти, были массивными и крепились к костному выступу на своде черепа, называемому сагиттальным гребнем. Пока тринаксодон стремительно взрослел, рос и его гребень, усиливая мощь его укуса. Встречаются окаменелости, сохранившие нескольких тринаксодонов вместе, из чего следует, что они были общественными животными и собирались в группы. В некоторых случаях образцы запечатлевают взрослых особей в компании особей помельче и помоложе – явное свидетельство заботы о потомстве.
Осталось отметить еще одну важную вещь о тринаксодоне. Места его обитания не ограничивались бассейном Кару. Его ископаемые остатки находят также в Антарктиде, и это признак того, что он настолько хорошо приспособился к ужасному миру раннего триаса, что распространился по всей Пангее. Земля теперь собралась в единый материк, окруженный обширным океаном, и по мере того, как вулканы остывали, а экосистемы оправлялись от вымирания, цинодонтам предстояло стать главными игроками на этой новой арене.
Как и многие другие писатели, Вальтер Кюне создал некоторые из своих лучших творений в тюрьме. То, как он – палеонтолог, увлеченный строением микроскопических зубов, – попал туда, заслуживает отдельного рассказа.
Кюне родился в 1911 г. в Берлине в семье учителя черчения. Он изучал палеонтологию в Университете Фридриха-Вильгельма в Берлине, а затем в Университете Галле и был известен двумя увлечениями: интересом к средневековым церковным колоколам, о которых написал восторженную статью в туристический журнал, и своими прокоммунистическими симпатиями. Последнее значительно больше волновало представителей нацистских властей, которые как раз начинали свой террор. Молодой Кюне был приговорен к девятимесячному тюремному заключению – это был его первый опыт за решеткой, – и в 1938 г. ему пришлось эмигрировать в Великобританию.
Как мог политический беженец без средств прокормить себя и молодую супругу в чужой стране? Естественно, собирая окаменелости. Кюне слышал, что в середине XIX в. в пещере близ Холуэлла, деревеньки в несколько сотен жителей, обнаружили горстку зубов триасовых млекопитающих. Это открытие должно было сыграть важнейшую роль, но осталось без внимания. Похоже, среди палеонтологов нашлось не слишком много желающих месяцами просеивать горы гравия в надежде отыскать малюсенькие зубки. Тем не менее куратор Британского музея посоветовал ему не утруждаться. «Все английские местонахождения ископаемых уже известны, – видимо, сказал куратор. – Бессмысленно ожидать новых крупных открытий».
Кюне это не отпугнуло. Он отчаянно нуждался в деньгах и горел интересом к млекопитающим. Кроме того, у него были козыри в рукаве. Еще в студенчестве он обнаружил чутье на окаменелости, и вдобавок у него было одно важнейшее качество – терпение. Он отправился в Холуэлл, где с энтузиазмом собрал, промыл и просмотрел более 2 т глиняной засыпки из пещеры. Задачу облегчила помощь его жены Шарлотты, хотя о ее интересе к крошечным зубам (или об отсутствии такового) история умалчивает. Трудолюбие четы Кюне было вознаграждено: они нашли два предкоренных зуба. Вальтер тут же направился в Кембриджский университет и показал их палеонтологу Рексу Паррингтону, которого находка так впечатлила, что он внес Вальтера в платежную ведомость. Отныне за каждый зуб млекопитающего ему полагалось £5.
Полные надежд, Вальтер и Шарлотта взялись за обследование других пещер и ущелий на юге Британии. Вскоре, в августе 1939 г., они открыли новые окаменелости в Мендип-Хиллс, пасторальных холмах к югу от Бристоля, в сельских окрестностях графства Сомерсет. Они собрали десятки разрозненных зубов и костей, принадлежавших очень близкому к млекопитающим цинодонту олигокифусу (Oligokyphus), описанному десятилетиями раньше по какой-то паре зубов из Германии. Затем они двинулись дальше, вперед, к следующему великому открытию. В сентябре Вальтер с киркой и геологической картой отправился на атлантическое побережье, где принялся дотошно исследовать известняковые скалы. Неизвестно, был ли он в курсе, что покинутое им отечество только что вторглось в Польшу.
Британским солдатам, патрулировавшим побережье, безусловно, было известно о начале мировой войны. Им показалось странным, что по английскому побережью ходит немец с картами, и они его арестовали. Так Вальтер во второй раз очутился в заточении, в лагере для интернированных на острове Мэн – маленькой точке в Ирландском море между Великобританией и Ирландией. Лагерь станет его домом с 1941 по 1944 г.
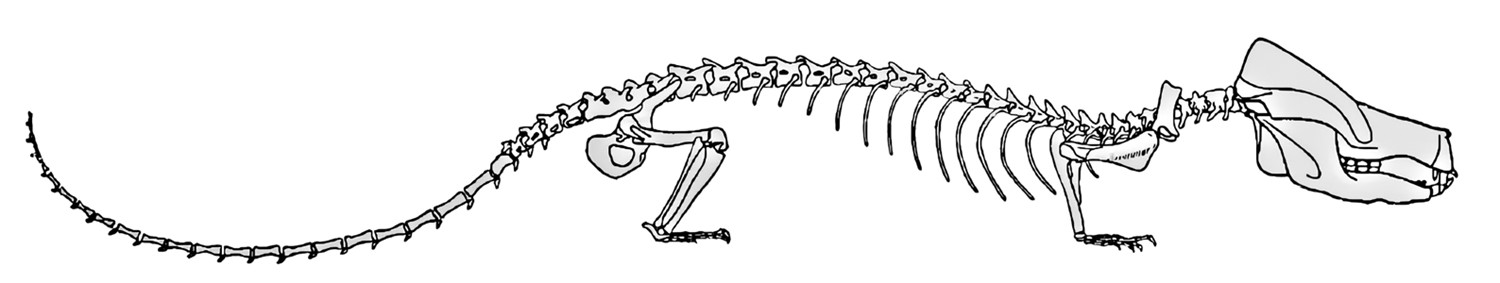
Oligokyphus.
Иллюстрация из монографии Вальтера Кюне 1956 г. (по Kühne, 1956)
Единственное, что скрашивало его жизнь, – это то, что к тому времени Вальтер снискал себе репутацию среди английского научного истеблишмента. Уже вскоре после бомбежек Лондона кураторы и научные сотрудники Британского музея – когда-то отговаривавшие увлеченного немца от поисков британских окаменелостей – организовали новые экспедиции в пещеры Мендип, которые принесли множество новых открытий костей и зубов. Две тысячи находок переслали в лагерь для интернированных.
Как говорил Вальтер, «в распоряжении у меня было достаточно времени». Он разбирал находки, складывал кости и собрал бóльшую часть скелета олигокифуса. Скрупулезное описание ископаемых остатков не давало ему скучать, и к тому времени, когда на исходе войны его освободили, он начал записывать свои выводы, что положило начало его будущей монографии 1956 г. об олигокифусе. Она до сих пор считается одной из основополагающих работ о тех цинодонтах, которые находились на грани превращения в млекопитающих.
Олигокифус – размером и формой напоминающий таксу – не был млекопитающим и даже прямым предком млекопитающих. Скорее их очень близким родственником, вроде двоюродного брата. Он был представителем подгруппы прогрессивных растительноядных цинодонтов – тритилодонтид, соседней с млекопитающими на родословном древе. Как и положено близким родственникам, тритилодонтиды и первые млекопитающие были очень похожи и по внешнему облику, и по поведению. Например, и у тех и у других конечности располагались прямо под туловищем, так что они ходили полностью на лапах, в отличие от других цинодонтов, которые все еще передвигались несколько распластавшись. И тритилодонтиды, и млекопитающие были результатом волны диверсификации цинодонтов, последовавшей в конце триаса, около 220 млн лет назад, то есть 30 млн лет спустя после того, как тринаксодон пережил пермско-триасовое массовое вымирание и – до поры до времени – уберег линию млекопитающих в момент ее наибольшей уязвимости. За эти 30 млн лет многое изменилось: «стволовая линия» млекопитающих продолжала накапливать «маммальные» признаки – вроде выпрямления лап, – пока цинодонты лавировали в лабиринте с суровым климатом и еще более суровой конкуренцией.
Самое большое изменение состояло, в сущности, в малом. Уменьшение размеров уже помогло цинодонтам в конце пермского периода, и они продолжали в том же духе. На протяжении триаса млекопитающие становились все более миниатюрными. Представленная поначалу видами размером с ласку – вроде тринаксодона, – к концу триаса эта линия обратилась в разнообразных вредителей в основном размером от мыши до крысы. Из этого правила были исключения, так как боковые ветви родословного древа иногда давали более крупные виды, подобные олигокифусу и его собратьям-тритилодонтидам, нуждавшиеся в более объемистом кишечнике для переваривания растительной пищи. Но в целом эволюция триасовых цинодонтов шла по пути миниатюризации.
Почему цинодонты уменьшались? Хотя бы потому, что они были не одни в дивном новом мире триасового периода. Пермско-триасовую катастрофу пережили и другие животные, и когда Пангея пошла на поправку, они стали конкурировать за место под солнцем. Из этого горнила эволюции вышли не только млекопитающие, но и многие другие привычные нам животные, которые и поныне сосуществуют с млекопитающими: черепахи, ящерицы и крокодилы. Кроме того, по всему суперконтиненту распространялось нечто еще более страшное. Оно росло в размерах и множило разнообразие видов, происходивших от скромного предка величиной с кошку, пережившего вулканизм.
Динозавры.
Их отцы-основатели боролись за первенство со своими родичами-крокодилами, причем те и другие становились все крупнее. К концу триасового периода уже существовали девятиметровые длинношеие динозавры весом в 10 т, наподобие лессемзавра (Lessemsaurus) – примитивного родича колоссальных завропод типа бронтозавра, – и множество зубастых хищников, охотившихся на них. Чем больше становились динозавры, тем меньше – предки млекопитающих. Это стало началом повторяющегося сюжета с переплетающимися судьбами млекопитающих и динозавров.
Одновременно с уменьшением в размерах многие цинодонты из линии млекопитающих, вероятно, перешли на ночной образ жизни. Выползать в ночную тишь для поисков пищи и общения с себе подобными – удачная тактика, чтобы не попасть в зубы или под ноги динозаврам, которые были, по-видимому, дневными животными. Переход в ночную нишу не вызвал особых затруднений, так как, судя по всему, многие предшественники из синапсидной линии, ведущей к млекопитающим, – пермских пеликозавров и терапсид – уже экспериментировали с таким образом жизни. Однако темнота имела свои последствия. Предки млекопитающих фактически отказались от острого зрения, обменяв его на обоняние, осязание и слух. Большинство млекопитающих не различает цвета, поэтому их шерсть обычно окрашена в унылые серо-коричневые тона. Зачем одеваться ярко – как это делает большинство современных дневных птиц и рептилий, обладающих хорошим зрением, – если партнеры или соперники все равно не видят цвета? Нам это может показаться странным, у нас-то зрение цветное! Но мы представляем собой редкое исключение среди млекопитающих – один из немногих современных видов, восприимчивых к цвету, наряду с нашими ближайшими родственниками из числа приматов. Когда матадор размахивает перед быком красным плащом, быку он кажется черным.
Если бы избегать динозавров было недостаточно причин, малый размер мог дать и другие преимущества. Пангея была единым массивом суши, но жить на ней было небезопасно. Там было жарко, на полюсах не было ледников, а бóльшая часть внутренних областей представляла собой бескрайнюю пустыню. Вдоль экватора проходили мощные воздушные течения, создававшие так называемые мегамуссоны – как явствует из их гиперболического названия, это были гигантские аналоги современных тропических штормов. Хотя теоретически протомлекопитающее могло пройти от полюса до полюса, это было бы бессмысленное путешествие. Мегамуссоны способствовали разделению Пангеи на климатические провинции с разным уровнем осадков и скоростью ветров, с разными температурами. Экваториальные регионы, адски жаркие и влажные, с обеих сторон окружали непроходимые пустыни. Средние широты, впрочем, были немного прохладнее и гораздо более влажными, чем пустыни. Именно там в основном и жили быстро эволюционировавшие животные Пангеи. Небольшой размер мог быть стратегией выживания в этом опасном мире. Чем ты меньше, тем легче спрятаться, зарыться, переждать в спячке мегамуссонные дожди и погибель, которую они несут.
Независимо от причин сокращения размеров цинодонтов, этот процесс глубоко изменил всю их биологию и эволюционную траекторию. С уменьшением размеров менялись и многие аспекты их роста, метаболизма, питания и способов добывания пищи. Они уже имели повышенные температуру тела и уровень метаболизма, унаследованные от терапсидных предков, но теперь у них развилась полноценная теплокровность. Они и так обладали сильными челюстными мышцами и сильным укусом – доставшимися от еще более древних предков-пеликозавров, – но теперь у них появилась возможность быстро поглощать большие объемы пищи и в то же время дышать, что позволяло есть на ходу.
Это было продолжением «взаимосвязанного прогресса», описанного Томом Кемпом применительно к пермским терапсидам. Многое менялось согласованно, и трудно разобраться, что было первичным. Вероятно, малый размер предполагал необходимость более высокой температуры тела, чтобы противостоять внезапным климатическим изменениям, или требовал более эффективных способов сбора и переваривания маленьких порций пищи. Может быть, теплокровность предписывала цинодонтам больше есть, чтобы хватало энергии, или наоборот – сначала изменилось строение челюстей и мышц, позволяя больше есть и обеспечивая таким образом избыток энергии для развития теплокровности. Ответ неизвестен. Зато мы знаем, что маленький размер, теплокровный метаболизм и более мощный, эффективный укус появились вместе как часть пакетной сделки.
Откуда это известно? Подсказку дает удивительное богатство ископаемой летописи триасовых цинодонтов – непрерывный ряд от тринаксодона до олигокифуса и млекопитающих.
Для начала разберемся с физиологией и метаболизмом. Мы уже касались этого предмета в предыдущей главе, но он заслуживает более подробного пояснения. Термин «теплокровность» – удобное краткое обозначение целого спектра сложных способов регуляции температуры тела. Он не означает, что у теплокровных животных кровь теплая, а у холоднокровных холодная. На самом деле, если поставить термометр среднестатистическому теплокровному млекопитающему и среднестатистической холоднокровной ящерице, показатели, скорее всего, получатся близкими. Может быть, у ящерицы он окажется даже выше, особенно в солнечный день. Дело в том, что холоднокровные животные согревают свои тела теплом из окружающей среды, а значит, они зависимы от изменений погоды, сезона, даже температурных перепадов дня и ночи, солнца и тени. Теплокровные животные – по-научному эндотермы – освободились от этого недостатка. Они производят собственное тепло – обычно благодаря повышенному содержанию в клетках митохондрий, вырабатывающих энергию, – и таким образом способны поддерживать температуру тела выше окружающей среды. Мы можем испытывать это всякий раз, когда выходим зимой на мороз и не превращаемся в ледышку.
По сути, теплокровные животные имеют собственную внутреннюю печь, которая всегда включена и всегда горячая. Это позволяет повысить метаболизм, быстрее расти, вести более активный образ жизни, быть выносливее и спортивнее. Млекопитающие способны поддерживать скорость бега в восемь раз бóльшую, чем ящерицы, например, и обследовать в поисках пищи более обширные территории. Но за эти сверхспособности приходится платить. У теплокровных животных интенсивнее метаболизм в состоянии покоя, а значит, в покое они сжигают больше калорий, чем холоднокровные. И конечно, они затрачивают намного больше калорий в активном состоянии – когда бегают, прыгают, охотятся, удирают от хищников, лазают по деревьям, роют норы и делают многое другое, что упрощается благодаря теплокровности. Поэтому им нужно потреблять больше калорий, чем холоднокровным животным такого же размера, и вдыхать больше кислорода.
Впрочем, не все животные относятся либо к теплокровным, либо к холоднокровным. Существуют промежуточные варианты. Современные млекопитающие и птицы полностью теплокровны, что подразумевает три особенности. Температура их тела контролируется изнутри, она высокая и постоянная, независимо от условий внешней среды. Подобная система эволюционировала не единовременно, а постепенно – через ряд переходных стадий, на которых их предки совершенствовали как выработку тепла, так и регуляцию температуры своего «отопления». В случае млекопитающих этот процесс начался с терапсид пермского периода, вынужденных в своих высоких широтах приспосабливаться к климату с выраженными сезонами. Их быстрый рост и усовершенствованная регуляция температуры тела, по-видимому, сыграли решающую роль в выживании – в частности, цинодонтов вроде тринаксодона – во время пермско-триасового события. В триасе эти цинодонты продолжили двигаться в сторону полноценной теплокровности.
Существует множество свидетельств в пользу того, что в ходе триасового периода цинодонты постепенно превращались в полностью теплокровных. Фиброламеллярной костной ткани – с хаотически организованной текстурой, указывающей на быстрый рост, – в линии, ведущей к млекопитающим, становилось все больше. Костные клетки и канальцы кровеносных сосудов внутри костей в ходе эволюции цинодонтов становились меньше, что говорит об уменьшении эритроцитов – еще одна особенность млекопитающих, благодаря которой эти клетки быстрее и лучше усваивают кислород. В одном интересном исследовании для измерения изотопного состава кислорода в организмах различных пермских и триасовых видов измельчили их кости и зубы. Соотношение двух наиболее стабильных изотопов – более легкого и распространенного 16O к более тяжелому и редкому 18O, отличающемуся количеством нейтронов, – зависит от температуры, при которой росли кости и зубы. По существу, это соотношение кислорода являет собой палеотермометр, и его данные однозначны: у триасовых цинодонтов температура тела была более высокой и постоянной, чем у сосуществовавших с ними других животных, включая остальных терапсид.
Как цинодонты оплачивали счета за отопление? Так же как любое теплокровное животное – потребляя много кислорода и много калорий. На пути к млекопитающим цинодонты приобрели многие анатомические особенности, позволившие им увеличить потребление того и другого.
В первую очередь цинодонты решили нелегкую проблему так называемого ограничения Карриера, которое одолевает рептилий и амфибий. Передвигаясь враскоряку, они изгибают тело из стороны в сторону, вправо-влево. Из-за этого бокового изгибания одно легкое растягивается, а другое сдавливается попеременно, поэтому животное не может одновременно дышать и бегать, что серьезно ограничивает их скорость и подвижность. Как мы уже знаем, у цинодонтов не только туловище приняло более приподнятое положение, но, кроме того, появились костные выросты на позвонках, мешающие позвоночнику колыхаться из стороны в сторону. Эти модификации скелета полностью изменили их способ ходьбы: теперь их конечности двигались вперед-назад, а не из стороны в сторону, а позвоночник у них изгибался вверх-вниз (как у скачущей газели), а не вправо-влево (как у ползущей змеи). Теперь они могли при ходьбе дышать без затруднений.
Позвоночный столб подвергся еще одному изменению: в то время как у амфибий и рептилий он примерно одинаков по всей длине, у цинодонтов он разделился на отделы со специализированными функциями. Туловищный отдел позвоночника разделен на два: грудной с ребрами и поясничный без ребер. Это явный признак наличия диафрагмы – мощной мышцы, накачивающей воздух в легкие.
Изменился и череп. У цинодонтов появилось вторичное нёбо – твердый свод ротовой полости, отделяющий ее от носовой. Воздух получил собственный отдельный проход в легкие, и цинодонты смогли одновременно питаться и дышать. В новой носовой полости появились необычные новые структуры – носовые раковины: закрученные костные или хрящевые завитки, выступающие посредине воздушного потока. Они могли бы показаться помехой, но в действительности играют ключевую роль в поддержании постоянной температуры тела при вдыхании большого количества воздуха. Некоторые носовые раковины покрыты кровеносными сосудами, согревающими и увлажняющими воздух при вдохе, прежде чем он попадет в легкие. У нас они тоже есть – внутри черепа, между ноздрями и задней стенкой горла. Они гарантируют нам, что даже самый холодный и сухой воздух в самые морозные зимние дни за доли секунды нагревается и увлажняется до уровня, соответствующего условиям тропического леса. К тому же они работают и в обратную сторону, компенсируя потерю драгоценной влаги при выдохе.
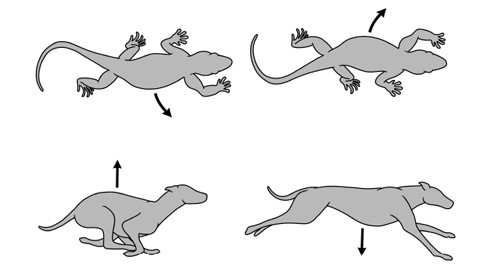
Различия в локомоции рептилий, которые перемещаются, двигая туловищем вправо-влево (вверху), и млекопитающих, которые двигаются вверх-вниз (внизу). Стрелками показано направление движения.
Рисунок Сары Шелли
Все эти, казалось бы, тонкие анатомические нюансы способствовали одному важному качеству: легкие усваивали намного больше кислорода. Одновременно с этим цинодонты повышали потребление калорий благодаря усовершенствованной технике укуса.
Первым шагом, по-видимому, стало разделение мышц, смыкающих челюсти, на отдельные пучки, способствовавшие более мощным и сложным движениям челюстей. Эти раздельные мышцы – характерная для млекопитающих система из височной, жевательной и крыловидных мышц – тоже укрупнялись, требуя больше места и более прочного крепления. Кости верхней части черепа упростились и затвердели, пока в конце концов не стали единой структурой – вот почему у нас есть «свод черепа», а не десятки раздельных, слабо связанных между собой костей, как у рептилий и амфибий. Боковое височное отверстие, появившееся еще во времена каменноугольных болот как признак линии синапсид, слилось с глазницей, образовав единую главную камеру для крепления мышц.
Однако самой глубокой перестройке подверглась нижняя челюсть. Предки млекопитающих обладали сложными челюстями, состоявшими из множества костей с разными названиями. У них были зубная кость, на которой, как и следует из названия, крепились зубы, сочленовная кость, образующая челюстной сустав посредством соединения с верхней частью черепа, и многие другие: угловая, надугловая, предсочленовная, пластинчатая, венечная. Но у нас – и у всех млекопитающих – осталась только одна, зубная. А все из-за того, что происходило с цинодонтами в триасе. По мере того как их челюстные мышцы становились больше, они сдвигались и в конце концов стали крепиться только к зубной кости. Это было совершенно логично: так как зубная кость – единственная, несущая зубы, то подобная перестройка мышц позволяла усилить укус и оптимизировать силу сжатия зубов на разных участках зубного ряда во время кусания.
Но это потребовало полного преобразования самих костей. Зубная кость стала больше и намного глубже. В ней появился массивный венечный отросток для крепления мышц, выступавший далеко за зубной ряд. Одновременно с этим другие кости нижней челюсти, к которым больше не крепились мышцы, стали редуцироваться. Они уменьшались и сдвигались назад, пока не отделились от зубной кости. Некоторые из них даже сошлись со стремечком – слуховой косточкой – и теперь, освободившись от функции кусания, стали помогать передавать звук.
Оставалась, правда, еще одна большая проблема: верхняя и нижняя челюсти все еще соединялись при помощи сочленовной кости, которая уменьшалась и сходила на нет. Редуцировалась и кость верхней части черепа, к которой она крепилась, – так называемая квадратная кость. У некоторых цинодонтов развился выступ надугловой кости нижней челюсти для соединения с верхней, но, поскольку надугловая кость тоже уменьшалась, это не приносило особой пользы. Ни один инженер не сконструировал бы систему смыкания челюстей подобным образом, и работала она только потому, что цинодонты настолько уменьшились, что им уже не требовалось выдерживать такие нагрузки при укусе, как их более крупным предкам.
Цинодонтам все же было необходимо найти какое-то решение. Они нашли – и получился признак, определяющий млекопитающих.
Классификация – человеческое занятие. Природа не вешает ярлыков, их вешают люди. Мышь – млекопитающее, змея – рептилия, тираннозавр – динозавр.
Млекопитающие, рептилии, динозавры. Все это группы на древе жизни, но как они устанавливаются?
Несложно определить, что такое млекопитающее, если рассматривать только современный мир. Мыши, слоны, люди, кенгуру, летучие мыши и тысячи других видов обладают общим набором уникальных признаков: у всех нас есть волосы, теплокровность, большой мозг, дифференциация зубов на резцы, клыки, предкоренные и коренные, мы выкармливаем потомство молоком и т. д. Но, как мы видели, эти «маммальные» признаки развивались по отдельности, в ходе 100 млн лет эволюции, когда пеликозавры каменноугольных болот породили терапсид, терапсиды пережили пермско-триасовое вымирание в виде мелких цинодонтов, а цинодонты в триасе продолжали уменьшаться. Где на этой эволюционной линии провести границу между немлекопитающими и млекопитающими?
Столкнувшись с изобилием новых находок пермских и триасовых синапсид в середине XX в., палеонтологи пришли к решению. Млекопитающими следует считать всех потомков первого животного, у которого появилась ключевая инновация – новый челюстной сустав между зубной костью нижней челюсти и чешуйчатой костью верхней части черепа. Этот новый сустав оказался простым, но элегантным решением проблемы непрерывного уменьшения костей в основании челюсти. Открытый Вальтером Кюне олигокифус не обладал сочленением зубной и чешуйчатой костей, а значит, не был млекопитающим. Но одна из ветвей родословной, куда входили виды типа морганукодона (Morganucodon) – еще одно из открытий Кюне в пещере Уэльса, – обладала. Итак, их договорились считать млекопитающими. Как и всех животных – живших в триасе и появившихся позже, включая нас, – родословную которых можно проследить вплоть до этого предка с новым челюстным суставом.
Это может показаться несколько неубедительным, субъективным. В некотором смысле это так. Но палеонтологи не просто выбрали случайный признак и привязали к нему название млекопитающих. Они выбрали один из фирменных признаков всех современных млекопитающих, четко отделяющий нас от амфибий, рептилий и птиц. Развитие нового, более крепкого сустава между зубными и чешуйчатыми костями стало также заметным поворотным моментом эволюции и, как мы скоро увидим, запустило каскад изменений, связанных с питанием, интеллектом и размножением млекопитающих. Это были финальные штрихи формирования млекопитающих после долгого процесса эволюционной сборки, последние кусочки мозаики, довершающие картину.
Здесь мне придется сделать отступление и признать, что приведенное выше определение в наши дни не используется большинством палеонтологов. В последние лет двадцать млекопитающих стали определять иначе – через так называемую кроновую группу. Этот подход объединяет всех ныне живущих млекопитающих – более 6000 видов однопроходных, сумчатых и плацентарных – и прослеживает их происхождение до последнего общего предка на родословном древе. Считается, что именно с него началось движение от немлекопитающих к млекопитающим. У «кронового» определения есть свои преимущества, и в первую очередь простота. Но у него есть и свои недостатки: сотни ископаемых видов, которые выглядели, вели себя, росли, метаболизировали, питались, выкармливали потомство и ухаживали за своей шерстью как млекопитающие, появились до того, как общий предок современных млекопитающих откололся от родословного древа, и получается, что все эти ископаемые животные не млекопитающие. Как, например, морганукодон.
Признаюсь честно: в своих научных работах я использую определение через кроновую группу. В научной статье я назову морганукодона не млекопитающим, а «базальным маммалиаформом» или «немаммальным маммалиаформом». Как видите, терминология быстро становится громоздкой. Чтобы избавить нас от выговаривания всех этих названий «почти млекопитающих», чуть-чуть не попавших в кроновую группу, для простоты я буду называть млекопитающими всех, у кого есть челюстной сустав, образованный зубной и чешуйчатой костями. Надеюсь, коллеги меня простят.
Независимо от определения, главное то, что в конце триасового периода цинодонты типа морганукодона нашли решение проблемы с челюстями. Зубная кость увеличилась и углубилась настолько, что, по-видимому, неизбежно сошлась с верхней частью черепа. Образовался шаровидный сустав, то есть новый шарнир для челюсти. Прежде слабое – и все ослабевавшее – сочленение челюсти и черепа укрепилось. Помимо соединения редуцировавшейся сочленовной кости с квадратной появилась вторая точка соединения между зубной костью и чешуйчатой. Некоторое время оба сустава сосуществовали – зубно-чешуйчатый снаружи квадратно-сочленовного. Потом квадратная и сочленовная кости редуцировались настолько, что больше не могли участвовать в открывании рта, поэтому подверглись дальнейшему уменьшению и в конце концов отделились от челюсти. Однако они не исчезли, а взяли на себя неожиданную новую функцию, о которой мы поговорим в следующей главе.
Зубно-чешуйчатый сустав полностью изменил положение. Челюсти, прежде довольно условно крепившиеся к черепу, вдруг обрели прочное крепление. Этот новый сустав – которым управляли челюстные мышцы, увеличившиеся на более раннем этапе эволюции цинодонтов, – теперь давал еще более сильный укус. Кроме того, укус становился намного более прицельным благодаря слаженным усилиям разделившихся височной, жевательной и крыловидной мышц – трех ниточек кукловода. Челюсти теперь могли точно прилагать основную силу укуса к определенным зубам в определенное время. Это открыло путь к совершенно новому способу питания, который мы воспринимаем как должное, но среди животных это редкость, – к жеванию. Пережевывая пищу в кашицу, первые млекопитающие смогли осуществлять немалую часть ее переработки во рту – фактически начинать переваривание еще до того, как пища попадет в желудок. Это был еще один способ получать больше калорий и более эффективно питаться.
Предки млекопитающих – различные пеликозавры, терапсиды и цинодонты, с которыми мы успели познакомиться, – по большей части обладали простым укусом. В этом отношении они походили на ящериц или хищных динозавров: они просто смыкали пасть, и все. Вверх-вниз, чик-чик. Они питались, отрывая и заглатывая куски пищи, особой переработки во рту не происходило. Исключением были некоторые прогрессивные цинодонты вроде олигокифуса, научившиеся при смыкании пасти двигать нижней челюстью назад и таким образом перетирать растения. Но это было нечастое явление.
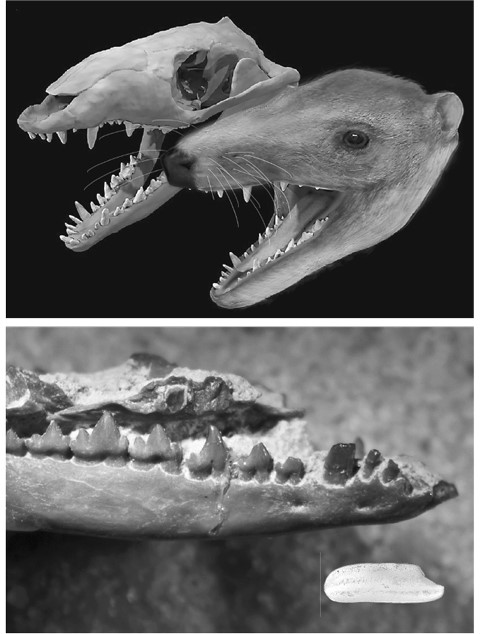
Морганукодон, одно из первых млекопитающих. Череп и реконструкция головы на основании компьютерной томограммы (вверху) и ископаемая нижняя челюсть (внизу; для масштаба рядом рисовое зерно).
Иллюстрации Стефана Лаутеншлягера и Памелы Джилл соответственно
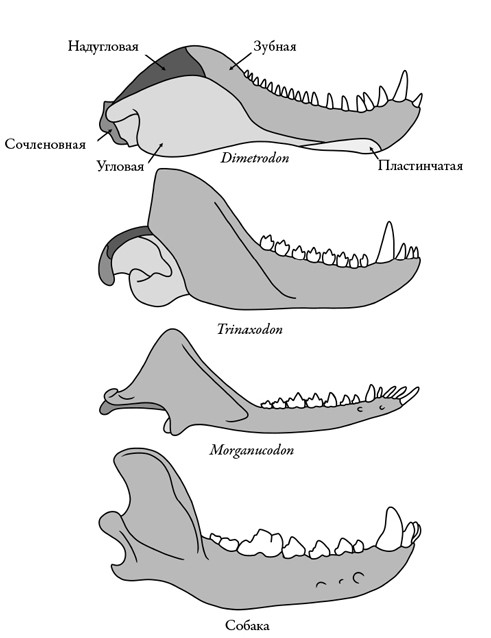
Редукция и упрощение челюстных костей синапсид со временем, пока не осталась единая кость нижней челюсти (зубная) млекопитающих.
Рисунок Сары Шелли
А вот триасовые млекопитающие стали жевательными машинами. Их челюсти теперь могли двигаться вверх-вниз, вперед-назад и из стороны в сторону. Они овладели жевательным циклом из трех последовательных действий, похожих на согласованные элементы плавания брассом. Вначале нижняя челюсть движется вверх и внутрь, смыкаясь с верхней челюстью (силовое движение), при этом рот закрывается, затем вниз, открывая рот (возвратное движение), а затем слегка наружу (подготовительное движение), и цикл повторяется вновь и вновь. Все это стало возможным благодаря уникальной способности нижней челюсти выдвигаться то наружу, то внутрь при открывании и закрывании рта. Покачайте как следует челюстями и почувствуйте, как они умеют смещаться в любых направлениях. Это и есть новые жевательные движения, впервые освоенные нашими отдаленными предками триасового периода.
Сложные скоординированные движения челюстей были только частью уравнения. Для того чтобы жевать, верхние и нижние зубы млекопитающих должны были сойтись вместе. Это называется окклюзией, и у динозавров или рептилий ее нет – у них верхние зубы обычно перекрывают нижние, подобно ножницам, или смыкаются зигзагообразно, но не совпадают поверхностями, когда пасть закрыта. Млекопитающие разительно отличаются от них, в чем каждый из нас может убедиться, закрыв рот. Хотя верхние резцы у нас слегка перекрывают нижние, наши щечные зубы плотно смыкаются – верхние стыкуются с нижними. У нас такая хорошая окклюзия зубов, что, когда мы полностью сжимаем челюсти, они фактически запираются. Именно большая площадь контакта и плотное смыкание верхних и нижних зубов обеспечили появление жевательной поверхности. Она возникла у триасовых млекопитающих типа морганукодона, сделавших последний шаг в развитии классической маммальной зубной системы – заклыковые зубы у них разделились на предкоренные и коренные.
Однако окклюзия создает одну проблему. Верхние и нижние зубы должны точно соответствовать друг другу, чтобы выступы одних входили в ямки других. Иначе они не будут смыкаться, и тогда жевание станет в лучшем случае неэффективным, в худшем – невозможным. Так произойдет с животными, меняющими зубы на протяжении всей своей жизни, – такими, как предки млекопитающих, – если они попытаются жевать. Даже если верхние и нижние зубы идеально подходили друг другу, при выпадении верхнего зуба нижнему будет больше не с чем смыкаться. Конечно, верхний отрастет, но не сразу, и пока зуб не встанет на свое место, смыкания с нижним не будет. Понятно, что для жующего животного это неудачный сценарий и млекопитающие снова пришли к оригинальному решению. Они прекратили менять зубы всю жизнь, обменяв неисчерпаемые поколения зубов, которыми обладали их предки, на всего лишь два набора: молочные зубы в период роста и затем постоянный набор взрослых зубов. Это называется дифиодонтизмом. Теперь, когда у вас сломается (или выпадет) зуб и вам понадобятся дорогостоящие услуги стоматолога, можете винить ваших триасовых предков.
Откуда нам известно, что все это происходило с первыми млекопитающими? Ну, во-первых, смыкание верхней и нижней челюстей – окклюзию предкоренных и коренных зубов – можно наглядно увидеть на ископаемых черепах морганукодона и других ранних млекопитающих. Во-вторых, на трехбугорчатых коренных зубах морганукодона и прочих триасовых млекопитающих есть следы износа: уплощенные поверхности, покрытые бороздами от истирания, которые образовались в результате постоянного контакта с противоположными зубами. В части износа зубов эти млекопитающие соблюдали хрупкое равновесие. Слишком интенсивный износ приводил бы к стачиванию зубов до пеньков – катастрофа для животного, неспособного больше отращивать новые зубы сколько угодно. Но при оптимальном уровне износа формировались плоские поверхности с острыми краями, работавшие как ножницы при смыкании челюстей, что добавляло к жеванию разрезание.
Ископаемые остатки этих первых млекопитающих – с прочными челюстями, зубной окклюзией и умением жевать – примечательны еще в одном отношении. Их череп обладает вместительной полостью, содержавшей мозг намного более крупный, чем у их пеликозавровых, терапсидных и цинодонтовых предков. Заметнее всего вырос передний отдел, и трубковидный предковый мозг обратился в более шарообразный, присущий современным млекопитающим, с округлыми полушариями.
На основании компьютерной томограммы черепов морганукодона была создана цифровая реконструкция его мозга, которая, помимо увеличенных размеров и округлившейся формы, показала еще две ключевые особенности, характерные для млекопитающих. Обонятельные луковицы – отвечающие за восприятие запахов – у него массивны. А у переднего мозга появилась новая надстройка – шестислойная масса нервной ткани, которая называется новой корой. Это одно из высших достижений млекопитающих. Неврологи считают новую кору ключом к интеграции чувств, научению, памяти и интеллекту. Таким образом, первые млекопитающие триаса одновременно учились лучше жевать и умнели. То ли усиленное питание способствовало росту и усложнению мозга, то ли увеличение мозга вызвало изменения в положении челюстных мышц, сделав укус более мощным, а движения челюстями более сложными. Так или иначе, у первых триасовых млекопитающих эти существенные изменения питания и интеллекта происходили взаимосвязанно.
Морганукодон – самый знаменитый представитель этой первой волны млекопитающих и яркий образец базальной группы млекопитающих, пережившей расцвет в триасе и продолжавшей существовать в юрский период. В 1949 г. Кюне, чудесным образом ставший после своего освобождения преподавателем Лондонского университета, описал животное по единственному миллиметровому обломку зуба, который удалось отыскать в девятикилограммовом мешке гравия, собранного в одной из пещер Уэльса. В 1952 г. Кюне вернулся в Германию и стал профессором в Берлине, а исследования в Южном Уэльсе продолжила супружеская пара Кеннет и Дорис Кермак. Они нашли сотни новых костей и зубов.
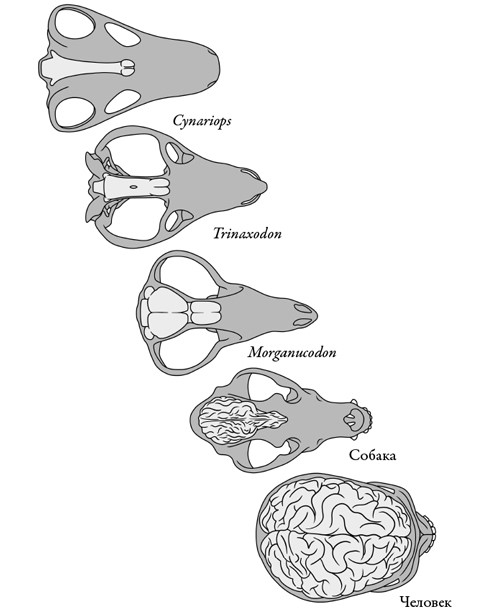
Рост мозга синапсид во времени, итогом которого стал крупный мозг млекопитающих со складчатой структурой и увеличенной новой корой переднего мозга. Масштаб 3 см.
Рисунок Сары Шелли
Тем временем в Китае команда иезуитов-палеонтологов из Католического университета Фужэнь в Пекине занималась собственными поисками первых млекопитающих. Один из них, отец Эдгар Эйлер, отправился в провинцию Юньнань, где откопал череп длиной в 2,6 см, с нижней челюстью. Он отослал его в Пекин своему собрату по ордену отцу Гарольду Ригни. Когда в дверь Ригни постучались спецслужбы – недавно созданные новым правительством Мао, – он как раз изучал ископаемые. Его арестовали и бросили в тюрьму на четыре года. Ригни повезло меньше, чем Кюне: он не имел возможности заниматься палеонтологией в заключении, так как коммунисты всерьез вознамерились уничтожить религиозные организации в Китае. Усилиями тайной дипломатии Ригни в конце концов был освобожден из тюрьмы и вернулся в США, где его ждал череп, который вывезли втайне от коммунистов. Он отложил его в сторону и засел за автобиографию – изящно озаглавленную «Четыре года в красной преисподней» (Four Years in a Red Hell) – и только потом описал находку в знаменитой статье 1963 г., объявив, что это новый вид морганукодона. Эта великолепная окаменелость, вкупе с пополняющимся фондом костей и зубов из Уэльса, сделала морганукодона эталонным представителем ранних млекопитающих.
Были и другие ранние млекопитающие – целый набор видов, живших в триасе и юре. Многих из них нашли в Великобритании, и, как и морганукодона, их находят в пещерах Англии и Уэльса. Возможно, эти халявщики жили под землей, но более вероятно, что их смывало туда во время наводнений или они проваливались в коварные трещины и их кости со временем накапливались в этих доисторических гробницах. Один экземпляр нашел Кюне, но не распознал поначалу. Находка была представлена двумя зубами, обнаруженными им вскоре после эмиграции в Великобританию. Он продал эти зубы Рексу Паррингтону в Кембридже, и тот определил животное как новый вид, дав ему название «эозостродон» (Eozostrodon). Еще один образец нашла в уэльских пещерах Диана Кермак, назвавшая его «кюнеотерий» (Kuehneotherium) – в честь Кюне.
Были и такие, к которым Кюне не имел никакого отношения. В Южной Африке любительница-археолог Айони Раднер – самоучка, которая сначала была вольнослушательницей в Кейптаунском университете, а затем научным сотрудником Южноафриканского музея – обнаружила интересный череп и скелет животного, получившего название «мегазостродон» (Megazostrodon). По другую сторону земного шара, в Гренландии, гарвардский профессор и бывший военный моряк Фэриш Дженкинс – носивший с собой на раскопки ружья, чтобы отгонять белых медведей, и поддерживавший в экспедиционном лагере казарменные порядки – нашел челюсти и зубы млекопитающих типа морганукодона. Есть и более свежие открытия, например замечательный гадрокодиум (Hadrocodium) из Китая. Головка у него была с ваш ноготь, а туловище – с канцелярскую скрепку; таким образом, размером он не превышал самого маленького среди ныне живущих млекопитающих – свиноносой летучей мыши из Таиланда, которая весит полтора-два грамма.
Первые млекопитающие были успешными. Они диверсифицировались с головокружительной скоростью, используя преимущества своего большого мозга и новообретенного умения жевать. Они распространились по всей Пангее и вскоре стали всемирным явлением. Млекопитающие были одной из первых основных групп животных, которые широко расселились по суперконтиненту, и им не помешали мегамуссоны и пустыни. Большинство из них, по-видимому, были насекомоядными – мелкие размеры и сильный точный укус идеально подходили для того, чтобы занять нишу прытких ловцов насекомых. Но питались они не одними и теми же насекомыми. Пэм Джинн и ее коллеги сравнили износ зубов морганукодона и кюнеотерия и обнаружили, что эти два вида, сосуществовавшие в Уэльсе раннего триасового периода, специализировались на различной пище. Морганукодон ел насекомых с твердым панцирем, как у жуков, тогда как кюнеотерий предпочитал более мягкую добычу вроде бабочек. Это было первое проявление тенденции, которая будет воспроизводиться на протяжении всей эволюции млекопитающих: когда появлялась новая волна представителей этого класса, она обычно начиналась с мелких насекомоядных, которые незаметно диверсифицировались.
Когда Фэриш Дженкинс не собирал челюсти млекопитающих с ружьем за спиной, он представал в роли галантного профессора Лиги плюща, читающего лекции в безупречных костюмах. Он был артистичным лектором и забавлял несколько поколений студентов своим завуалированным юмором и умелым применением реквизита, самым скандальным образцом коего была деревянная нога, как у капитана Ахава, – ее он надевал, чтобы продемонстрировать разные типы человеческой походки. Однако больше всего его бывшим студентам запомнились его тщательные зарисовки черепов и скелетов – его основное наглядное пособие в эпоху до PowerPoint. Он приходил в аудиторию за несколько часов до занятий, иногда затемно – когда ночные млекопитающие гарвардского двора охотились на насекомых, – и рисовал мелом на доске свои шедевры.
Самый знаменитый из его рисунков – мегазостродон. Впервые этот рисунок, отредактированный профессиональным художником, был опубликован в работе Дженкинса 1976 г., где описывался скелет, найденный за десятилетие до того Айони Раднер. Рисунок стал одной из классических научных иллюстраций, которая часто фигурирует в учебниках и, да, в презентациях PowerPoint, столь популярных в наши дни. Подобно культовому портрету Че Гевары, он означает нечто большее, чем просто отдельного индивида. Это образ революции – первых млекопитающих, распространившихся по Пангее и основавших династию.

Революционное изображение древнего млекопитающего Megazostrodon, созданное Фэришем Дженкинсом.
На основе иллюстрации из Jenkins & Parrington, 1976
Малютка мегазостродон, величиной с мышь или землеройку, встревожен. Он стоит, насторожившись, у корней дерева, гордо держа на лапах гибкое тельце. Момент нерешительности. Ступни задних лап расположены так, словно он собирается взобраться на дерево и исчезнуть в ветвях. Но кисти передних направлены наружу, давая возможность удрать по земле. Он что-то чует – то ли опасность, то ли вкусную букашку, так как челюсти приоткрыты и в них полный набор резцов, клыков, предкоренных и коренных. Зубки крохотные, но острые и выглядят так, словно готовы сомкнуться за доли секунды. Глазницы маленькие, а значит, для ориентации в окружающей среде зверек полагался в основном не на зрение. Что бы ни насторожило его, он услышал это или учуял.
Если бы мы увидели это животное во плоти, а не его скелет, мы бы его узнали. Несомненно, это млекопитающее. Оно было покрыто мехом, ходило, покачиваясь туда-сюда, на своих лапках и обладало полным комплектом маммальных зубов, использовавшихся для жевания и менявшихся лишь единожды за всю жизнь. Оно было ловким, умело лазить по деревьям и бегать по земле, и оно было теплокровным или, по крайней мере, приближалось к этому, то есть обладало способностью поддерживать комфортную температуру тела, охотясь за насекомыми на ночном холоде. Внутри черепа у него находился большой мозг, обеспечивавший ему интеллект и острое обоняние, слух у него тоже работал отменно.
Если бы этот зверек пробежал перед вами в лесу или на платформе метро, уверен, вы бы подумали, что это обычная мышь.
На сцену вышли млекопитающие, наследники столь многих удивительных адаптаций, накопленных за 100 с лишним миллионов лет эволюции их предками – пеликозаврами, терапсидами и цинодонтами. Новые животные расселялись по планете и готовились завладеть Пангеей – отвоевать то, что их терапсидные прародители утратили из-за разбушевавшегося пермско-триасового вулканизма.
Или нет?
Суперконтинент начинал распадаться, а динозавры становились все больше и злее. У новоявленных млекопитающих – несмотря на все их эволюционные инновации – возможности были ограниченны.
Им пришлось на отлично освоить искусство жить инкогнито.
Назад: 1 Предки млекопитающих
Дальше: 3 Млекопитающие и динозавры

