Книга: Кости, гены и культура
Назад: Нейрохимическая гипотеза происхождения человека
Дальше: Глава 10 Социальность и интеллект
Эволюционные корни агрессии
Раз уж мы заговорили об эволюции и нейрохимии поведения, нельзя обойти молчанием и такую тему, как агрессия, тем более что и в этом вопросе наука за последние годы заметно продвинулась. В частности, появились новые данные по летальной агрессии у наших ближайших родственников шимпанзе.
Систематическое убийство себе подобных – крайне редкое явление среди обезьян, которое на сегодняшний день доподлинно известно только у одного вида – у обыкновенного шимпанзе, Pan troglodytes. На первый взгляд кажется, что естественный отбор мог бы чаще поддерживать такое поведение, ведь физическое устранение конкурентов, по идее, должно повышать приспособленность агрессоров, то есть их шансы на успешное размножение (книга 2, глава 5, раздел “Окситоцин и парохиальность”, подраздел “Обезьяньи войны”).
Однако этого не наблюдается, а причины редкости смертельных схваток у обезьян тоже, в общем-то, очевидны. По мнению многих исследователей, главная причина состоит просто-напросто в том, что эскалация агрессии повышает риск получить сдачи, который сильно перевешивает ожидаемые выгоды от расправы над конкурентом. Даже нападение на одинокого детеныша у высокосоциальных приматов чревато последствиями, ведь малыш обязательно криком позовет на помощь родню. Еще одно препятствие на пути эволюции крайних форм агрессии состоит в том, что весь риск берут на себя сами агрессоры, тогда как выигрыш от ослабления соседнего сообщества и расширения территории делится на всю группу. В такой ситуации больше потомства в среднем оставят как раз менее агрессивные члены сообщества (в теории эволюции альтруизма это называют “проблемой безбилетника”; книга 2, глава 5).
И все же у шимпанзе схватки с летальным исходом происходят систематически. Это привлекает внимание исследователей в том числе и потому, что многие склонны усматривать здесь истоки человеческой воинственности. Как-никак шимпанзе и куда менее агрессивные бонобо – два наших ближайших родственника из всех ныне живущих видов.
Согласно самой распространенной среди специалистов точке зрения, агрессивное поведение шимпанзе – это не случайность и не патология, а нормальная адаптация, то есть признак, повышающий приспособленность и закрепившийся под действием естественного отбора. Предполагается, что особенности экологии и социальной организации шимпанзе (в отличие от других обезьян) способствуют увеличению выигрышей и снижению рисков, связанных с убийством себе подобных. Например, сообщества шимпанзе постоянно то разбиваются на подгруппы, сильно различающиеся по численности, то снова объединяются. Это позволяет агрессорам время от времени использовать подавляющее численное превосходство, сводя к минимуму риск получения нападающими серьезных травм. Для многих популяций шимпанзе характерна острая конкуренция между самцами за самок, а успешный военный рейд на территорию соседнего сообщества дает доступ к дополнительным половым партнершам. Такие особенности шимпанзе, как развитый социальный интеллект и огромная роль личных взаимоотношений и репутации, вероятно, позволяют этим умным животным успешно решать проблему безбилетника и обходить другие трудности, связанные с организацией рискованных коллективных предприятий.
Впрочем, не все антропологи согласны с таким эволюционным объяснением. Альтернативная точка зрения состоит в том, что убийство себе подобных – не адаптация, а случайный побочный результат человеческого вмешательства в обезьянью жизнь. Например, в национальном парке Гомбе, где Джейн Гудолл начала наблюдения за дикими шимпанзе в 1960 году, она поначалу подкармливала обезьян, чтобы они перестали ее бояться. В результате шимпанзе стали больше времени проводить около лагеря и чаще ссориться друг с другом (ожидаемое следствие конкуренции за обильный ресурс, сконцентрированный в одном месте). Подобные наблюдения навели некоторых исследователей на мысль, что замеченные позже смертельные схватки у шимпанзе вызваны антропогенным влиянием: если не подкармливанием, то охотой (ведь некоторые африканцы до сих пор охотятся на шимпанзе ради мяса!), разрушением местообитаний, хозяйственной деятельностью человека в местах проживания обезьяньих сообществ. Все это гипотетически может способствовать росту общего уровня агрессии у шимпанзе, и в таком случае рассматривать убийство себе подобных как эволюционно обусловленную адаптацию было бы неверно.
Чтобы проверить эти гипотезы, большой международный коллектив приматологов, долгие годы наблюдавших за 22 дикими сообществами шимпанзе и бонобо, проанализировал все накопленные сведения о стычках с летальным исходом. Объем информации внушает уважение: были обработаны данные, сложившиеся в сумме в 426 лет наблюдений за коллективами шимпанзе и 92 года наблюдений за коллективами бонобо (Wilson et al., 2014).
Все убийства подразделили на три группы: 1) наблюдавшиеся непосредственно, 2) установленные на основе неопровержимых улик (таких как найденный труп с характерными ранами от зубов шимпанзе), 3) предполагаемые (например, когда особь после жестокого избиения исчезла и больше никогда не попадалась на глаза исследователям). В общей сложности ученые насчитали 152 убийства у шимпанзе (58 наблюдавшихся, 41 доказанное и 53 предполагаемых) и одно-единственное предполагаемое убийство у бонобо.
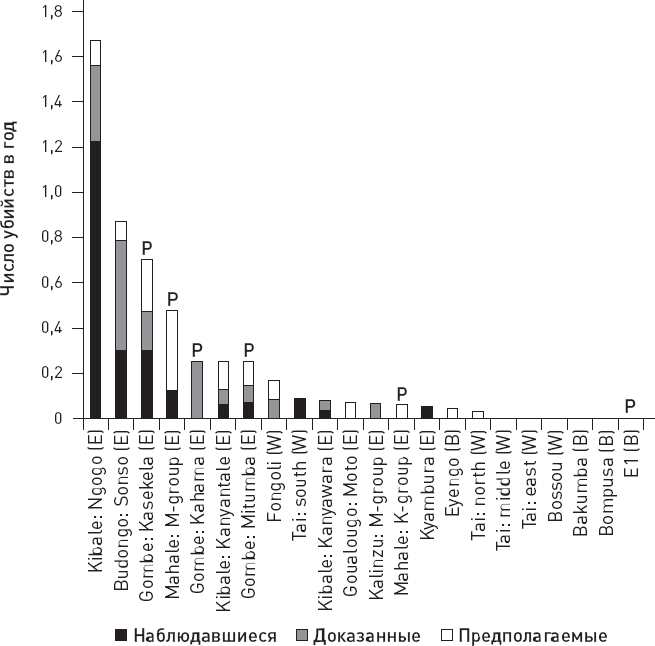
Рис. 9.10. Частота убийств, совершаемых членами 22 сообществ шимпанзе и бонобо, в расчете на год. Черным цветом обозначены наблюдавшиеся убийства, серым – доказанные, белым – предполагаемые. Для каждого сообщества указаны его название и район наблюдений (например, для первого сообщества Kibale – район, Ngogo – название). E – восточные популяции шимпанзе, W – западные, B – бонобо, P – сообщества, подкармливавшиеся людьми. По рисунку из Wilson et al., 2014.
Исследованные сообщества сильно различаются по частоте убийств (рис. 9.10). Некоторые группы западных шимпанзе, обитающие в относительно сухих и голодных районах, ни разу не замарали рук убийством сородичей, тогда как в самых жестоких сообществах (все они относятся к восточным популяциям, живущим в высокопродуктивных тропических лесах) частота убийств достигает устрашающих значений: 0,7–1,7 убийства в год на сообщество, что с учетом численности сообществ соответствует уровню 1,1–1,4 убийства на 100 особей в год. Это в 50–70 раз больше, чем в сообществах Homo sapiens, населяющих ныне Центральную и Западную Африку (по данным ООН за 2012 год).
Ученые исходили из того, что две гипотезы – эволюционная и антропогенная – по-разному предсказывают, с какими факторами должны коррелировать межгрупповые различия по частоте убийств и каким должен быть характер этих убийств, то есть кто кого и при каких обстоятельствах должен чаще убивать.
Если во всем виновато человеческое влияние, логично предположить, что больше убийств будет там, где это влияние сильнее. Исследователи принимали во внимание три показателя, связанные с антропогенным воздействием: 1) наличие или отсутствие подкормки, 2) площадь охраняемой территории (большинство изученных сообществ обезьян, хотя и не все, живут в заповедниках и национальных парках) и 3) совокупный показатель “человеческого воздействия”, учитывающий данные по интенсивности охоты на обезьян, разрушению местообитаний, хозяйственной деятельности и наличию крупных хищников (предполагается, что в районах с многочисленным человеческим населением или с его активной хозяйственной деятельностью крупных хищников должно быть мало). Проверялись гипотезы как о совместном, так и об индивидуальном влиянии каждого из трех показателей на частоту убийств у шимпанзе.
Если же верна эволюционная гипотеза, то следует ожидать, что интенсивность убийств и их характер будут определяться теми факторами, которые влияют на эволюцию экстремальных форм агрессии. Так, теория родственного отбора предсказывает, что чужаки (члены других сообществ) должны подвергаться жестоким атакам существенно чаще, чем свои, хотя встречи и конфликты со своими происходят у шимпанзе в природе во много раз чаще, чем с иноплеменниками. Кроме того, в роли агрессоров должны чаще выступать самцы, а не самки. Дело в том, что самцы получают наибольший выигрыш от устранения соперников, поскольку их репродуктивный успех зависит от числа доступных половых партнеров намного сильнее, чем у самок. Логично также предположить, что чем больше самцов в группе (и чем выше плотность популяции), тем острее конкуренция и, следовательно, тем чаще самцы будут проявлять экстремальные формы агрессии. Кроме того, следует ожидать, что жертвами убийц чаще всего будут самцы (как прямые конкуренты) и маленькие детеныши (поскольку самка, лишившаяся грудного детеныша, быстрее становится готовой к новому зачатию). Наконец, баланс рисков и выигрышей, направляющий действие отбора на поведенческие признаки, должен способствовать тому, чтобы агрессоры старались использовать численное преимущество, атакуя противника всерьез лишь при максимальных шансах на победу и минимальной вероятности ранений.
Статистический анализ данных не подтвердил ни одного из предсказаний антропогенной гипотезы. Частота убийств у шимпанзе не зависит ни от подкормки, ни от интенсивности хозяйственной деятельности людей, ни от природоохранных мер.
Предсказания же эволюционной гипотезы, напротив, получили весомые подтверждения. Оказалось, что для того, чтобы предсказать максимально точно уровень убийств в сообществе, достаточно знать только две переменные из всех рассмотренных, а именно плотность популяции и число самцов в группе. Чем эти показатели выше, тем чаще происходят убийства. Прочие показатели существенного влияния на частоту убийств, по-видимому, не оказывают.
Например, максимальный уровень убийств отмечен в сообществе Нгого (Уганда) с высокой плотностью популяции и низким уровнем антропогенного воздействия, тогда как в сообществе Боссу (Гвинея) с максимальным антропогенным вмешательством и низкой плотностью популяции убийств ни разу не наблюдали. При этом плотность популяции и у шимпанзе, и у бонобо достоверно коррелирует с обилием пищевых ресурсов (которых много в дождевом лесу и мало в сухой саванне), но не связана с уровнем антропогенного воздействия.
В последние годы приматологи стали регистрировать у шимпанзе больше убийств, чем раньше. Было высказано предположение, что это связано с ростом антропогенного воздействия. Исследователи проверили это и выяснили, что на самом деле никакого роста кровопролитности у шимпанзе не происходит. Суммарное количество регистрируемых убийств выросло из-за того, что в фокус внимания ученых попало большее число сообществ шимпанзе. Если взять не сумму, а частоту убийств в расчете на сообщество или на 100 особей, то никаких значимых изменений за период наблюдений не было.
В полном соответствии с предсказаниями эволюционной гипотезы, среди шимпанзе-убийц резко преобладают самцы. Если суммировать все 64 случая, когда атакующих обезьян удалось точно распознать, то выходит, что самцы составляли 92 % от общей численности нападающих (338 особей из 366). Самки изредка присоединялись к самцам во время нападения на взрослого противника (3 случая), но самки-убийцы, действующие без помощи самцов, умерщвляли лишь беспомощных младенцев (8 случаев).
Жертвами убийств чаще всего становятся самцы и грудные детеныши, что тоже соответствует предсказаниям эволюционной гипотезы. Более того, ученые сообщают о нескольких случаях, когда атакующие убивали грудного детеныша, но оставляли в живых его мать, хотя явно могли расправиться и с ней тоже.
Из 99 смертоносных атак, для которых известно, к какому сообществу принадлежали нападающие и их жертвы, 62 (63 %) были межгрупповыми, а не внутригрупповыми стычками. Как видим, соответствующее предсказание эволюционной гипотезы тоже подтвердилось, хотя возможностей и поводов для конфликтов со своими у шимпанзе куда больше, чем с чужаками. С членами своего сообщества обезьяны контактируют ежедневно, а с чужаками – лишь изредка (например, в сообществе Каньявара в Уганде такие встречи наблюдались в среднем лишь раз в 50 дней).
В межгрупповых стычках команда атакующих обычно включала много самцов (от 2 до 28, медианное значение – 9), а нападали они почти всегда или на самок с грудными детенышами, или, реже, на самцов. При этом численное превосходство агрессоров, как правило, было подавляющим. Только одна из всех наблюдавшихся стычек началась при равных силах атакующих и обороняющихся, а в среднем агрессоры имели восьмикратный численный перевес (восемь на одного: нечестно, зато надежно).
Таким образом, полученные результаты – весомый аргумент против идеи о зловредном (буквально убийственном) влиянии антропогенного фактора на поведение шимпанзе. По-видимому, склонность к агрессии у шимпанзе представляет собой адаптацию, повышающую репродуктивный успех агрессора, когда риск получить сдачи сведен к минимуму.
Вывод об адаптивном характере склонности к убийству у шимпанзе не следует трактовать в духе “наивного биологического детерминизма”. Иными словами, не нужно думать, что раз наши ближайшие родственники выработали в ходе эволюции такую адаптацию, то и наша человеческая воинственность имеет глубокие эволюционные корни и потому не может быть преодолена. Эволюционные корни она, скорее всего, действительно имеет (книга 2, глава 5; ниже мы еще вернемся к этой теме), но это вовсе не делает ее непреодолимой. Ведь человеческое поведение – результат не только биологической, но и культурной эволюции; эти два процесса у рода Homo с давних пор тесно переплетены (книга 2, глава 3; глава 12 здесь). Если общество выработает соответствующие моральные нормы и институты, то мрачное биологическое наследие вполне может быть если не искоренено, то в достаточной мере подавлено и обуздано. И потом, миролюбивые, сексуальные и демократичные бонобо являются нашими родственниками ровно в той же степени, что и воинственные, склонные к построению жестких “вертикалей власти” шимпанзе. Так на чье же наследие мы станем ориентироваться? Осмысленный выбор за нами.
Еще одно важное исследование, результаты которого были опубликованы в 2017 году, пролило несколько неожиданный свет на роль нейропептида окситоцина в агрессивном поведении наших ближайших родичей (Samuni et al., 2017).
О роли окситоцина в регуляции поведения млекопитающих, включая человека, рассказано во второй книге (главы 3 и 5). Напомним, что, похоже, у всех млекопитающих окситоцин стимулирует заботу о потомстве. Например, если девственной крысе ввести в мозг окситоцин, она начинает заботиться о чужих крысятах, которые в норме ей безразличны. Если же у крысы-матери блокировать окситоциновые рецепторы, она перестает заботиться о своих детенышах. Окситоцин, по-видимому, подавляет чувство страха, что способствует эффективной родительской заботе, например бесстрашной защите любимого детеныша от опасного хищника (книга 2, глава 3, раздел “Нейрохимия личных отношений”).
У видов, ведущих социальный образ жизни или формирующих устойчивые брачные пары, окситоциновая система регуляции родительской привязанности, судя по всему, неоднократно кооптировалась для обслуживания дружеских взаимодействий и супружеской привязанности.
В последние годы стали появляться данные, показывающие, что у наших предков окситоциновая система, по всей видимости, была привлечена для обслуживания еще одной функции – парохиальности, то есть хорошего отношения к своим в противовес чужакам (книга 2, глава 5, раздел “Окситоцин и парохиальность”). Парохиальность – один из механизмов выживания социальных животных в условиях острой межгрупповой конкуренции. По-видимому, у людей в ситуации межгрупповой вражды окситоцин способствует доброму отношению только к тем, кого мы считаем своими, а по отношению к чужакам он может стимулировать враждебные действия, правда, скорее оборонительные, чем наступательные, хотя грань тут бывает очень зыбкой (De Dreu et al., 2012; De Dreu, 2016).
Эти данные хорошо согласуются с гипотезой сопряженной эволюции парохиального альтруизма и войн у наших предков. Согласно этой гипотезе, в условиях чрезвычайно острой межгрупповой вражды повышенная склонность к внутригрупповой кооперации и альтруизму развивалась в неразрывной связке с ксенофобией (книга 2, глава 5).
Для наших ближайших родственников шимпанзе (но не для наших столь же близких родственников бонобо), как мы уяснили из предыдущего исследования, характерна ярко выраженная парохиальность. Она проявляется в более частых нападениях на чужаков, чем на членов своего коллектива. Если соображения о связи между окситоцином, внутригрупповой сплоченностью и враждебностью к чужакам верны, то следует ожидать, что межгрупповые конфликты у шимпанзе должны сопровождаться усиленным выделением окситоцина нейронами гипоталамуса. Ведь сплоченность группы повышает шансы на успех в борьбе с внешними врагами, и если такая борьба происходит постоянно, а окситоцин повышает сплоченность, то отбор должен способствовать усиленному производству окситоцина в контексте межгруппового конфликта.
Германские антропологи решили проверить, так ли это, на примере двух враждующих сообществ диких шимпанзе в национальном парке Тай в Кот-д’Ивуаре (Samuni et al., 2017). У тамошних шимпанзе, по сравнению с другими популяциями, в межгрупповых стычках дело редко доходит до убийства (см. рис. 9.10). Но все же они регулярно патрулируют границы своих территорий и по-честному дерутся с членами соседних групп. Может быть, относительно низкий уровень риска является причиной того, что в парке Тай в рейдах по патрулированию границ и в драках активно участвуют самки (даже обремененные грудными детенышами), тогда как в популяциях с высоким уровнем кровопролитности воюют в основном самцы. Или, наоборот, женское участие снижает кровопролитность конфликтов?
Еще одна особенность шимпанзе из парка Тай состоит в том, что они давно привыкли к присутствию антропологов и уже не обращают на них внимания. Это позволило ученым следовать за обезьянами буквально по пятам, снимая все их действия на видео и оперативно собирая пробы мочи с опавшей листвы при помощи пластиковых пипеток. Эти пробы замораживались и отправлялись в Европу для определения уровня окситоцина. В научной литературе существуют разногласия по поводу того, насколько точно уровень окситоцина в моче отражает его концентрацию в мозге. Окситоцин, производимый нейронами некоторых участков (ядер) гипоталамуса, попадает, с одной стороны, в кровь и затем в мочу, с другой – работает нейромодулятором в мозге, влияя на функционирование отделов, связанных с системой внутреннего подкрепления, социальным поведением и привязанностями. На сегодняшний день большинство специалистов склоняется к тому, что уровень окситоцина в моче служит адекватной мерой концентрации окситоцина в мозге. Если происходит какое-то событие, способствующее выбросу окситоцина, то его уровень в моче повышается через 15 минут после события и остается повышенным около часа. Исследователи, конечно, учитывали это при анализе данных.
Наблюдения и сбор проб проводились в 2013–2015 годах. В общей сложности было отснято около 4500 часов наблюдений и собрано 482 пробы мочи от 20 индивидов (по 5 самцов и 5 самок из каждого сообщества).
Для начала ученые проверили гипотезу о том, что межгрупповой конфликт стимулирует внутригрупповую сплоченность. Для этого они подсчитали случаи, когда какой-нибудь индивид “дезертировал”, то есть покидал свою компанию во время межгрупповых конфликтов (к конфликтам относили как групповое патрулирование границ, так и собственно драки с чужаками), и когда особь отходила от коллектива в мирных обстоятельствах. Контрольные наблюдения (мирные) были проведены таким образом, чтобы они максимально точно соответствовали наблюдениям, проведенным во время конфликтов, как по продолжительности, так и по числу обезьян, попадающих в поле зрения камер.
Оказалось, что во время боевых действий обезьяны втрое реже покидают свою группу, чем в спокойной обстановке. Между патрулированием и собственно дракой по этому показателю различий не обнаружилось: в обеих ситуациях случаи дезертирства были немногочисленны. Дополнительные расчеты показали, что дело тут не в близости границ участка: если компания обезьян находится на краю своей территории, но занимается мирными делами, а не патрулированием, то особи свободно уходят и возвращаются. Когда же начинается боевой рейд (а начинается он с весьма специфического группового поведения), сплоченность группы резко возрастает. Исследователи отмечают со ссылками на психологическую литературу, что повышенная сплоченность и дружеские чувства по отношению к своим характерны и для человеческих солдат, отправляющихся в атаку.
Результаты анализа уровня окситоцина в моче показаны на рисунке 9.11. Оказалось, что участие в военных действиях сопровождается повышением уровня окситоцина. Это справедливо как для патрулирования, не закончившегося дракой, так и для драк, причем между этими двумя вариантами разницы не обнаружилось. Аффилиативные проявления во время подготовки к рейду (объятия, груминг, коллективные игры) не оказывают значимого влияния на уровень окситоцина – тот подскакивает у бойцов независимо от того, обнимались ли они с товарищами перед выходом в рейд или нет (две правых позиции на рис. 9.11). В мирное время уровень окситоцина у обезьян значительно ниже – и тоже вне зависимости от наблюдаемых проявлений дружбы, таких как груминг (две левых позиции). Ранее уже было показано, что груминг у шимпанзе сам по себе не ведет к повышению уровня окситоцина – здесь все зависит от социального контекста и, наверное, от “истинных чувств”, которые трудно измерить. Однако известно, что окситоцин у шимпанзе повышается, когда они делятся с сородичами пищей – и потому такие случаи в данном исследовании исключались из рассмотрения.
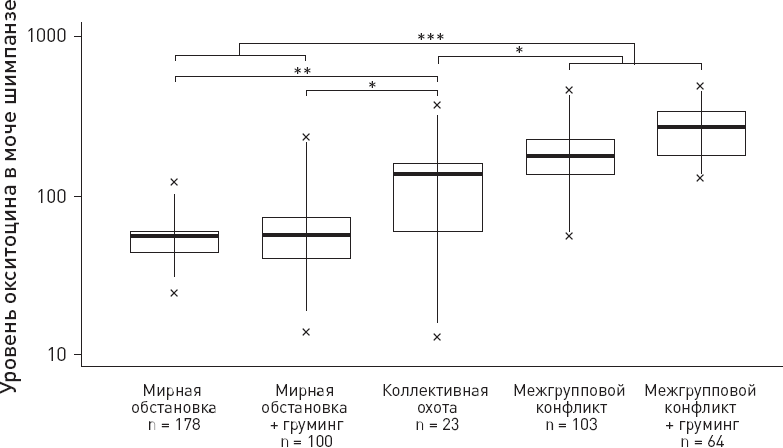
Рис. 9.11. Уровень окситоцина (в условных единицах) в моче шимпанзе в пяти ситуациях, слева направо: 1) мирная обстановка, обезьяна не участвует в груминге или иных проявлениях дружбы; 2) мирная обстановка, обезьяна участвует в продолжительном (не менее 10 минут за 90-минутный период наблюдения) коллективном груминге; 3) коллективная охота на мелких обезьян; 4) межгрупповой конфликт без предшествующего участия в аффилиативных действиях, таких как груминг или игра; 5) межгрупповой конфликт с предшествующим участием в аффилиативных действиях. Показаны медианные значения (черные горизонтальные линии), квартили (прямоугольники), перцентили (2,5 и 97,5 %, вертикальные линии), минимумы и максимумы (крестики). По рисунку из Samuni et al., 2017.
В качестве дополнительного контроля рассматривалась коллективная охота на мелких обезьян – поведение, требующее координации групповых действий, но не связанное с риском и враждебностью к чужакам. Оказалось, что у охотников концентрация окситоцина повышается, но не так сильно, как у участников боевых рейдов (третья позиция на рис. 9.11). При этом такие факторы, как пол обезьяны, ее социальный статус, число особей в группе и близость границ участка, не влияют на уровень окситоцина.
Полученные результаты согласуются с предположением о том, что окситоцин усиленно вырабатывается во время коллективных действий, боевых или охотничьих. Он способствует координации групповых действий, которые необходимы как для охоты, так и для успешной защиты территории от воинственных соседей. При этом угроза со стороны чужаков, даже только предполагаемая, как в случае рейдов, не приведших к столкновению, сильнее активирует окситоциновую систему по сравнению с охотой на безобидных колобусов.
Кроме того, выяснилось, что у шимпанзе, участвовавших в патрулировании, а перед этим – в коллективном груминге, уровень окситоцина был повышен еще до того, как начались вопли, беготня и объятия, связанные непосредственно с инициацией рейда. Это стало ясно из сравнения их с обезьянами, которые участвовали в груминге, но в рейд не пошли. Иными словами, если у шимпанзе, участвующего в коллективном груминге в мирной обстановке, измерить уровень окситоцина, то по результатам анализа можно с известной точностью предсказать, пойдет ли данная обезьяна в ближайшем будущем в боевой поход. Похоже, на войну идут те, кого переполняет любовь к сородичам. Ранее у шимпанзе было обнаружено такое же “предваряющее” повышение уровня тестостерона перед межгрупповыми конфликтами (Sobolewski et al., 2012).
Таким образом, окситоцин у шимпанзе, по-видимому, является не столько “гормоном любви и дружбы”, сколько “гормоном любви, дружбы, координации коллективных действий и войны”. Исследование показало, что нейрофизиологические основы человеческого парохиального альтруизма, возможно, сформировались уже у общего предка человека и шимпанзе, то есть раньше, чем предполагалось до сих пор. Впрочем, пока нельзя утверждать, что полученные результаты в полной мере приложимы к человеку. Брать пробы мочи у воюющих людей, следуя за ними по пятам с видеокамерами и пипетками, куда труднее, чем у воюющих шимпанзе. Но антропологи наверняка что-нибудь придумают.
В продолжение разговора о войнах и убийствах упомянем еще об одном любопытном исследовании, которое было посвящено поиску истоков человеческой жестокости и агрессивности. Вопрос этот, надо сказать, обсуждается уже не первое тысячелетие. Одни мыслители, подобно философу Томасу Гоббсу, полагали, что люди агрессивны и жестоки по самой своей природе. Другие думали, что люди от природы добродетельны, но цивилизация их портит (образ “благородного дикаря”), третьи – что все определяется культурой и воспитанием, а от природы люди не хороши и не плохи (концепция “чистого листа”).
Данные, накопленные современной наукой, в том числе психогенетикой (книга 2, глава 3), позволяют уверенно утверждать, что истина где-то посередине. Агрессивность, безусловно, зависит от культуры и среды, но она имеет также и генетическую составляющую, что проявляется в высокой наследуемости индивидуальных различий по уровню агрессии у многих млекопитающих, включая человека. А если признак имеет ненулевую наследуемость и при этом ненулевое значение для выживания и успешного размножения, то он просто по определению не может не эволюционировать под действием отбора.
Хосе Мария Гомес с факультета экологии Гранадского университета и его коллеги попытались реконструировать эволюционную историю летальной (то есть приводящей к гибели сородичей) внутривидовой агрессии у млекопитающих (Gómez et al., 2016). Для этого они собрали максимально подробные данные по летальной агрессии у 1024 видов, относящихся к 137 семействам (80 % всех семейств млекопитающих). Уровень летальной агрессии оценивался как число убийств, совершенных сородичами (представителями того же вида), выраженное в процентах от общего числа задокументированных смертей. Сюда входит инфантицид, каннибализм, межгрупповые стычки с летальным исходом и все прочие виды убийства сородичей. В итоге получилась внушительная сводка, содержащая данные более чем о четырех миллионах смертей у млекопитающих.
Убийства себе подобных отмечены почти у 40 % исследованных видов. Однако средний уровень летальной агрессии, характерный для млекопитающих, невелик: 0,3 ± 0,19 % (три убийства на тысячу смертей). Таким образом, летальная агрессия у млекопитающих – явление, с одной стороны, нечастое, с другой – встречающееся у значительного числа видов. Но это если данные по всем видам усреднить. Интересны в данном случае отклонения от среднего. У каких-то видов летальная агрессия повышена, а у каких-то вообще отсутствует – с чем это связано?
Первым делом исследователи проверили, имеется ли в данных по летальной агрессии “филогенетический сигнал”. Иными словами, они оценили, имеют ли родственные виды сходный уровень агрессии, или же агрессивность распределена по эволюционному дереву хаотично. Оказалось, что филогенетический сигнал есть, хотя и не очень сильный. Это значит, что агрессивность, с одной стороны, наследуется в череде происходящих друг от друга видов, с другой – может быстро меняться в отдельных ветвях. Например, в одних больших группах (таких как киты, рукокрылые и зайцеобразные) летальная агрессия встречается очень редко, а в других (приматы, некоторые семейства хищных) она вполне обычна. Но при этом встречаются случаи, когда два близкородственных вида имеют контрастно различающиеся уровни агрессии. Один из самых ярких примеров такого контраста – жестокие шимпанзе и миролюбивые бонобо.
Наличие в данных по агрессии значимого филогенетического сигнала позволило рассчитать для каждого вида (а также для глубоких ветвей и узлов дерева) “филогенетически ожидаемый” уровень агрессии. То есть оценить, каким должен быть этот уровень, исходя из положения данного вида на эволюционном дереве. Для подобных реконструкций существуют специально разработанные статистические методы и компьютерные программы.
Главный вопрос, конечно, в том, насколько надежны и информативны полученные оценки “эволюционно ожидаемого” уровня агрессивности. Проверить это в данном случае довольно просто: нужно для каждого вида взять ожидаемую величину агрессивности (которая вычисляется без учета реальных цифр по данному виду) и сравнить ее с эмпирически установленной. Проверка показала, что для 63 % видов эмпирический уровень летальной агрессии находится внутри 95-процентных доверительных интервалов для вычисленной ожидаемой величины. Иными словами, зная только позицию вида на эволюционном дереве, но ничего не зная о его реальных обстоятельствах, можно для 2/3 видов с приемлемой точностью предсказать уровень внутривидовой агрессивности. Не так уж плохо для признака, который многими по сей день считается исключительно средовым, а не врожденным.
Ученые также обнаружили, что на уровень летальной агрессии влияют, помимо эволюционного наследия, еще два параметра: социальность и территориальность. И то и другое способствует повышенной агрессии. Это вполне естественно: жизнь в группе сама по себе создает предпосылки для конфликтов, а если группа еще и охраняет свою территорию от других подобных групп, вероятность кровопролития дополнительно повышается.
Самыми агрессивными млекопитающими оказались сурикаты: 19,4 убийства на каждые 100 смертей.
Люди, как выяснилось, принадлежат к эволюционной ветви с чуть ли не самыми древними традициями убийства себе подобных. Для Homo sapiens ожидаемый уровень летальной агрессии, вычисленный на основе нашего положения на эволюционном дереве, составляет 2,0 ± 0,02 % (два убийства на 100 смертей). В ведущей к нам эволюционной линии уровень агрессии постепенно повышался. У последнего общего предка всех млекопитающих он составлял около 0,3 %, такой же уровень характерен и для общего предка всех плацентарных. Поднимаемся выше по эволюционному дереву: у общего предка приматов с их ближайшей сестринской группой, включающей грызунов с зайцами, уровень агрессии увеличился до 1,1 %. Еще выше располагается общий предок приматов, тупай и шерстокрылов (группа, объединяющая этих зверей, называется Euarchonta) – здесь уровень агрессии поднимается до 2,3 %. Этот показатель немного снизился у последнего общего предка человекообразных обезьян (1,8 %) и, наконец, слегка увеличился у предков человека (2,0 %). У шимпанзе агрессия выросла до 4,49 %, а у бонобо – снизилась до 0,68 %. По последним двум видам здесь приведены не ожидаемые, а эмпирические значения, что накладывает некоторые ограничения на прямое сравнение чисел.
Разумеется, первое, что хочется сделать, глядя на эти цифры, – это сравнить вычисленную ожидаемую величину летальной агрессии у H. sapiens (2 %) с реальными данными по палеолиту или по современным племенам охотников-собирателей. Ученые попытались решить эту задачу, собрав из литературных источников внушительный массив данных по летальной агрессии у человека, охватывающий почти 600 человеческих популяций от палеолита (древнейшие из учтенных археологических находок имеют возраст 50 тыс. лет) до современности.
Информации в итоге набралось много, однако она крайне разнородна по своему характеру, полноте и достоверности. К тому же при сборе подобных данных и их систематизации невозможно полностью избежать субъективности. Например, в данных по неандертальцам, использованных авторами в качестве вспомогательных при оценке эволюционно ожидаемого уровня агрессивности нашего вида, материал из пещеры Эль-Сидрон (книга 1, глава 5, раздел “Людоеды”) учтен как 12 ненасильственных смертей, потому что, дескать, данные по каннибализму у неандертальцев спорны и могут быть интерпретированы по-разному. Так же авторы поступили и с другими находками неандертальцев, съеденных соплеменниками, – и в итоге неандертальцы получились у них исключительно миролюбивым видом приматов. Поэтому делать слишком детальные выводы на основе приводимых авторами данных рискованно. Общая же картина получилась (со всеми этими оговорками) следующая.
Для палеолитических (начиная с 50 тыс. лет назад) и более поздних человеческих популяций вплоть до конца бронзового века (3200 лет назад по хронологии, используемой авторами) уровень летальной агрессии у людей не демонстрирует статистически значимых отличий от эволюционно ожидаемого уровня в 2 %. То есть наши предки были кровожадны ровно настолько, насколько эту кровожадность предопределяло их эволюционное наследие. В железном веке и в Средние века летальная агрессия резко превысила ожидаемый уровень – вплоть до 15–30 %, с очень большим межпопуляционным разбросом. В Новое время (500–100 лет назад) она снизилась до значений, достоверно меньших эволюционно обусловленного уровня, – до 0,14 % по данным письменных источников. Правда, по археологическим данным за тот же период значение получилось другое – 2,4 %. Для последних 100 лет, опираясь только на письменные источники, авторы приводят значение 1,3 %.
У авторов также получилось, что современные группы охотников-собирателей, судя по имеющимся этнографическим данным, характеризуются очень высоким уровнем летальной агрессии: 10,3 % для мелких групп, 3,9 % для более крупных племен. Однако для доисторических сообществ того же типа авторы приводят, на основании археологических данных, совсем другие значения: 3,3 и 3,6 % соответственно. То ли дожившие до наших дней охотники-собиратели действительно стали кровожаднее своих палеолитических коллег (что может быть, например, следствием контактов с более продвинутыми социумами, знакомства с их оружием и так далее), то ли в исходных данных что-то не так.
Но все же исследование довольно убедительно показало, что склонность людей к убийству себе подобных отчасти унаследована от далеких предков. Мы принадлежим к одной из самых агрессивных ветвей класса млекопитающих, а это что-нибудь да значит. Вероятно, дело тут не только или даже не столько в эволюционно обусловленной психологической склонности к убийству, сколько в эволюционно обусловленных особенностях образа жизни, способствующих агрессивному поведению, таких как чрезвычайно сильно развитая социальность.
Кроме того, исследование показало, что эволюционное наследие – это вовсе не приговор. Во-первых, последний общий предок шимпанзе и бонобо, живший 2 млн лет назад, имел такую же “эволюционно обусловленную” агрессивность, что и первые H. sapiens, однако один из потомков этого предка стал агрессивным шимпанзе, а другой – мирным бонобо. Во-вторых, степень летальной агрессии у людей резко менялась в разные эпохи и в разных типах общества, то превышая в несколько раз ожидаемый уровень, то опускаясь, как в современную эпоху, до обнадеживающе низких значений. Культурные и социальные факторы, несомненно, могут самым радикальным образом модифицировать и направлять наше поведение, либо стимулируя, либо подавляя эволюционно обусловленные предрасположенности.

