Книга: Кости, гены и культура
Назад: Предки алтайских неандертальцев скрещивались с древними сапиенсами, а предки денисовцев – с эректусами
Дальше: В геномах поздних европейских неандертальцев не нашли следов гибридизации с сапиенсами
Предки азиатов скрещивались с несколькими разными популяциями денисовцев
Сегодня любому школьнику известно, что поздние представители рода Homo – сапиенсы, неандертальцы и денисовцы, – скитаясь по просторам Старого Света, то и дело скрещивались друг с другом. Данные об этих скрещиваниях продолжают уточняться и детализироваться. Постоянно обнаруживаются новые эпизоды гибридизации. Следить за этим интересно, вот только не запутаться все труднее.
Перечислим основные эпизоды, о которых мы уже рассказывали:
1) “неизвестные архаичные люди” (скорее всего, азиатские эректусы) с общими предками неандертальцев и денисовцев – “неандерсовцами” (см. раздел “Предки неандертальцев и денисовцев скрещивались с потомками древних евразийских эректусов” в главе 3),
2) те же или слегка другие “архаичные люди” (тоже, видимо, эректусы) с денисовцами (см. раздел “Предки алтайских неандертальцев скрещивались с древними сапиенсами, а предки денисовцев – с эректусами” в главе 6),
3) древние сапиенсы с предками алтайских неандертальцев (см. раздел “Предки алтайских неандертальцев скрещивались с древними сапиенсами, а предки денисовцев – с эректусами” в главе 6),
4) неандертальцы с денисовцами, причем не менее двух раз (см. раздел “Денни, дочь неандерталки и денисовца” в главе 3),
5) неандертальцы с вышедшими из Африки сапиенсами около 60–50 тыс. лет назад, благодаря этому эпизоду в геномах внеафриканских сапиенсов присутствует около 2 % неандертальской ДНК (книга 1, глава 5, раздел “Геном неандертальца”),
6) денисовцы с предками современных жителей Юго-Восточной Азии, Австралии и Океании (книга 1, глава 5, раздел “Люди из Денисовой пещеры”).
А вот эпизоды, о которых мы либо упоминали вскользь, либо вообще не говорили, потому что к слову не пришлось:
7) Происходили неоднократные повторные скрещивания европейских и азиатских сапиенсов с неандертальцами (Fu et al., 2015; Vernot et al., 2016).
8) В геномах некоторых африканских народов, таких как пигмеи, хадза и сандаве, обнаружены следы гибридизации с неизвестной популяцией, отделившейся от предков сапиенсов примерно тогда же, когда разделились линии сапиенсов и “неандерсовцев”, – около 750–500 тыс. лет назад (Lachance et al., 2012; Hsieh et al., 2016).
9) В геномах жителей Западной Африки обнаружена значительная генетическая примесь от неизвестной африканской популяции, обособившейся, скорее всего, еще до разделения предков неандертальцев и сапиенсов (Durvasula, Sankararaman, 2020).
Поиск следов древних межвидовых скрещиваний интересен не только сам по себе – он дает бесценную информацию и о вымерших человечествах, и об истории наших предков. Например, пункты 8 и 9 из приведенного списка сообщают нам о том, что в Африке еще относительно недавно (уже после разделения некоторых современных африканских народов) водились какие-то неведомые люди, родственные сапиенсам не более, чем неандертальцы. Археологи их пока не нашли. Кем они были? Потомками человека из Брокен-Хилл (см. раздел “Гейдельбергские люди жили в Африке одновременно с ранними сапиенсами” в главе 5)? Или какой-то вовсе неизвестной нам веточкой человеческого рода?
Кроме того, анализируя неандертальские и денисовские примеси в геномах современных людей, мы можем узнать много нового о неандертальцах и денисовцах. Например, именно таким способом удалось выяснить, что денисовцы были разными: по-видимому, существовало не менее трех ветвей денисовцев, сильно отличавшихся друг от друга. Денисовская девочка Denisova 3 – представительница лишь одной из них. Откуда же мы узнали о двух других? Их “вычислили”, анализируя денисовские примеси в геномах современных жителей Восточной Азии и Новой Гвинеи.
Искать следы межвидовой гибридизации можно разными способами, и ученые постоянно изобретают новые. Можно просто искать в современных геномах фрагменты ДНК, совпадающие с известными неандертальскими и денисовскими последовательностями. Но таким способом нельзя обнаружить примеси, доставшиеся нам от вымерших популяций, по которым нет палеогенетических данных. Чтобы найти такие примеси, используют другие статистические методы, позволяющие даже и вовсе не принимать во внимание данные по древним геномам. Эти методы основаны на том, что привнесенные (интрогрессированные) фрагменты ДНК должны иметь определенное распределение длин: чем больше времени прошло с момента гибридизации, тем меньше их средняя длина. Кроме того, они должны содержать наборы тесно сцепленных, то есть расположенных по соседству, редких полиморфизмов. Если речь идет о гибридизации, происходившей уже после того, как предки разных рас и народов начали разбредаться по миру, то привнесенные участки должны встречаться только у некоторых, но не у всех современных популяций. Например, неандертальские примеси есть у всех внеафриканских сапиенсов, но отсутствуют у африканцев. Этот статистический подход, известный под названием “S*-статистика”, разрабатывается и совершенствуется уже довольно давно (Plagnol, Wall, 2006; Vernot, Akey, 2014).
В 2018 году американские генетики разработали новую, улучшенную версию S*-статистики, которую они назвали Sprime. Метод Sprime позволяет анализировать вместо последовательного обсчета коротких фрагментов сразу большие участки генома, а вместо попарных сравнений – данные сразу по множеству геномов. Кроме того, Sprime, в отличие от прежних версий, успешно справляется с таким осложняющим обстоятельством, как ограниченная миграция носителей интрогрессированных фрагментов ДНК в районы, населенные людьми без таких фрагментов. Например, этот метод позволил бы найти неандертальские примеси у неафриканцев, даже если бы некоторое (не слишком большое) число носителей этих примесей в далеком прошлом вернулось в Африку и скрестилось с местным населением. Конечно, без привлечения палеогенетических данных Sprime не определил бы, что это именно неандертальские примеси, но он бы показал, что это привнесенные фрагменты ДНК.
Проверив новый метод на виртуальных (сгенерированных компьютером) геномах с известной историей, исследователи убедились, что он обладает повышенной надежностью и чувствительностью по сравнению с прежними версиями. Так, если две популяции разошлись сотни тысяч лет назад, период гибридизации был десятки тысяч лет назад и при этом в генофонд изучаемой популяции попало 3 % чужих генов, метод Sprime позволит обнаружить половину интрогрессированных фрагментов (остальные окажутся слишком короткими или содержащими слишком мало полиморфизмов, уникальных для второй популяции), а доля ложноположительных результатов будет не более 7 %.
Убедившись, что метод работает, ученые применили его к 5639 полным геномам современных людей со всех концов Евразии, включая и папуасов (Browning et al., 2018). Количество и суммарная длина идентифицированных чужеродных фрагментов в геномах людей из разных популяций совпали с полученными ранее оценками доли неандертальской и денисовской ДНК в этих популяциях. В частности, подтвердилось, что у восточноазиатов примесей больше, чем у европейцев и южноазиатов.
Напомним, что все эти чужеродные фрагменты были выявлены при помощи метода Sprime без использования палеогенетических данных. Теперь, конечно, нужно было сравнить их с известными древними геномами. Ученые так и поступили, взяв для сравнения два наиболее качественно прочтенных генома – алтайского неандертальца (Denisova 5) и денисовской девочки (Denisova 3). Анализировались только те интрогрессированные участки, в пределах которых и у неандертальца, и у денисовца есть не менее десяти однонуклеотидных отличий от современных африканцев (у них, как считается, нет неандертальских и денисовских примесей). Для каждого такого участка определили степень его сходства, с одной стороны, с геномом алтайского неандертальца, а с другой – с геномом денисовца.
Результаты представлены на рисунке 6.2. Диаграммы, построенные для 20 внеафриканских популяций, отражают частоту встречаемости интрогрессированных фрагментов ДНК, характеризующихся разным уровнем сходства с геномами алтайского неандертальца и денисовца. Изолинии на диаграммах соединяют точки, обозначающие одинаковое количество фрагментов, для которых характерно то или иное сходство с двумя древними геномами. Эти диаграммы заслуживают внимательного разглядывания.
На всех диаграммах мы видим маленькую компактную область в левом нижнем углу. Это фрагменты, которые были определены программой Sprime как интрогрессированные, но они при этом имеют мало общего как с неандертальской, так и с денисовской ДНК. Скорее всего, это ложноположительный сигнал, то есть фрагменты, которые на самом деле интрогрессированными не являются и были определены программой ошибочно. Судя по результатам предварительного моделирования, программа должна давать 5–7 % ложноположительных результатов, что и подтверждается.
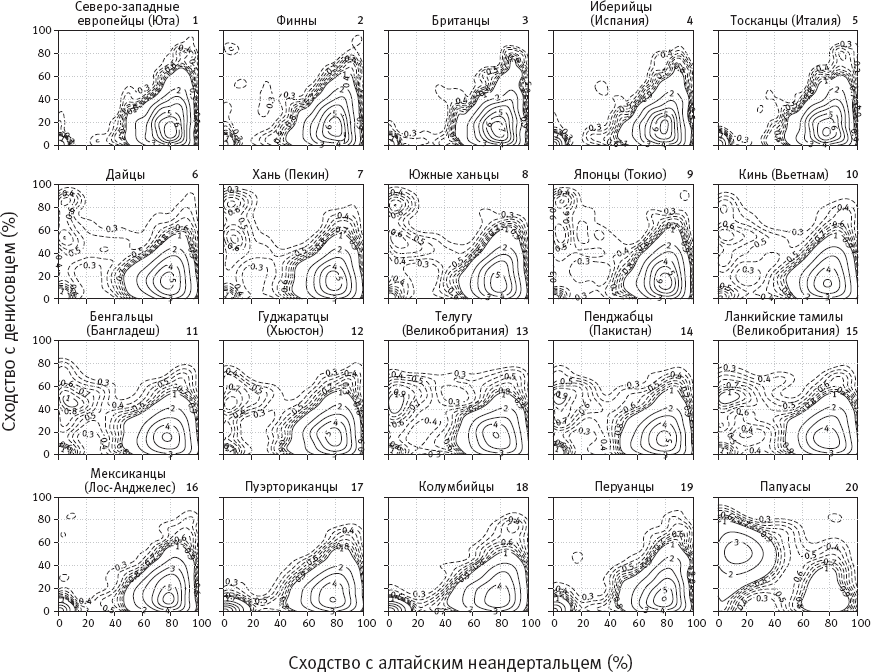
Рис. 6.2. Изолинии, отражающие частоту встречаемости интрогрессированных фрагментов ДНК с определенным уровнем сходства с геномами алтайского неандертальца и денисовца для геномов 20 внеафриканских популяций. Популяции, представленные на диаграммах 16–19 (от мексиканцев до перуанцев), – это не индейцы, а люди смешанного, преимущественно европейского происхождения. Прочие пояснения в тексте. По рисунку из Browning et al., 2018.
На всех диаграммах мы также видим внушительный подъем в правом нижнем углу. Это наше неандертальское наследие. Фрагменты, унаследованные от неандертальцев, сходны с геномом алтайского неандертальца в среднем на 80 %. При этом сходство с геномом денисовца у них низкое – порядка 10–20 %. Здесь нужно вспомнить, что по ядерному геному денисовцы ближе к неандертальцам, чем к сапиенсам, а это значит, что общие предки денисовцев и неандертальцев успели накопить какое-то количество общих отличий от сапиенсов еще до того, как разделились. Именно поэтому сходство неандертальских фрагментов с денисовским геномом достоверно отличается от нуля.
Денисовских примесей среди исследованных популяций больше всего у папуасов (диаграмма 20). Средний уровень сходства денисовских включений в папуасских геномах с известным геномом денисовца – около 50 %, а с геномом алтайского неандертальца – те же 10–20 %. То, что сходство этих включений с геномом Denisova 3 составляет лишь 50 %, а не больше, означает, что те денисовцы, которые оставили свой след в геномах папуасов, были довольно дальней родней девочки из Денисовой пещеры. Они представляли другую популяцию денисовцев, которая обитала, возможно, далеко от Алтая.
В геномах современных жителей Южной Азии (бенгальцев, индусов и других, диаграммы 11–15) тоже есть денисовская примесь, хоть и не такая большая. Причем уровень сходства привнесенных участков с геномом денисовской девочки такой же, как у папуасов, – около 50 %. Это значит, что источником примеси, вероятно, была та же самая популяция денисовцев, не слишком близкая к алтайской.
В геномах европейцев (диаграммы 1–5) денисовских примесей практически нет, за исключением финнов (диаграмма 2), у которых есть немножко денисовской ДНК. Скорее всего, эта примесь попала к финнам вместе с порцией азиатских генов (Sikora et al., 2014).
Самый интересный и неожиданный результат связан с восточноазиатами (три китайских выборки и одна японская, диаграммы 6–9). Их денисовская примесь состоит из двух разнородных частей: одни денисовские фрагменты сходны с геномом алтайской девочки на 50 % (как у папуасов и южноазиатов), а другие – примерно на 80 %. Двугорбость распределения статистически значима. Следовательно, у общих предков китайцев и японцев, скорее всего, было два эпизода гибридизации с денисовцами. Один раз они скрещивались с той же популяцией, что оставила след в геномах папуасов и южноазиатов, а второй – с другими денисовцами, более близкими к алтайской популяции.
Эпизод с участием “50-процентных” денисовцев мог быть тем же самым, что и у предков папуасов и южноазиатов. Иными словами, в этом эпизоде могли участвовать еще общие предки папуасов и жителей Южной и Восточной Азии. Или, может быть, собственно с денисовцами скрещивались только предки папуасов, представлявшие одну из древнейших волн миграции сапиенсов на юго-восток Азии, а уже потом от них получили денисовскую примесь предки индусов, китайцев и японцев. Что же касается эпизода с участием “80-процентных” денисовцев, то в нем были задействованы только предки восточноазиатов (китайцев и японцев).
Определить временную последовательность эпизодов гибридизации в принципе можно по длине интрогрессированных фрагментов. В геномах восточноазиатов неандертальские фрагменты в среднем чуть короче, чем “50-процентные” денисовские, а те, в свою очередь, короче, чем “80-процентные” денисовские. Это указывает на возможную последовательность событий: сначала скрещивание с неандертальцами, потом с “50-процентными” денисовцами и, наконец, с “80-процентными” денисовцами. Однако исследователи честно отметили, что различия по длине фрагментов не являются статистически значимыми – и поэтому делать какие-то выводы на их основе преждевременно.
Новые данные поставили под сомнение идею о множественных эпизодах гибридизации предков современных европейцев и азиатов с неандертальцами. Неандертальские фрагменты ДНК не распадаются ни на какие кластеры, а это значит, что либо эпизод был только один, либо повторные скрещивания происходили с неандертальскими популяциями, очень близкими к той, с которой предки внеафриканских сапиенсов гибридизовались изначально около 55 тыс. лет назад. Второй вариант более вероятен, потому что у азиатов достоверно больше неандертальской ДНК, чем у европейцев, и это на сегодняшний день не удается полностью объяснить действием отбора (то есть тем, что у предков европейцев отбор эффективнее вычищал неандертальскую примесь). Скорее всего, предки азиатов повторно скрещивались с неандертальцами, являвшимися близкой родней тех, с которыми ранее скрещивались общие предки всех внеафриканских сапиенсов.
Даже не надейтесь, что на этом изучение вклада денисовцев в генофонд современных людей завершилось или что нам уже все ясно по данному вопросу! В 2019 году были опубликованы результаты нового исследования, показавшие, что предки папуасов на самом деле тоже скрещивались не с одной, а как минимум с двумя разными популяциями денисовцев. Одна из них (получившая условное название D1) оставила свой генетический след только в геномах папуасов, а другая (D2) – папуасов и восточноазиатов. Обе эти популяции довольно далеки от алтайских денисовцев. И была, как мы уже знаем, третья популяция (D0), самая близкая к алтайским денисовцам, которая оставила свой след в геномах восточноазиатов, но не папуасов (рис. 6.3).
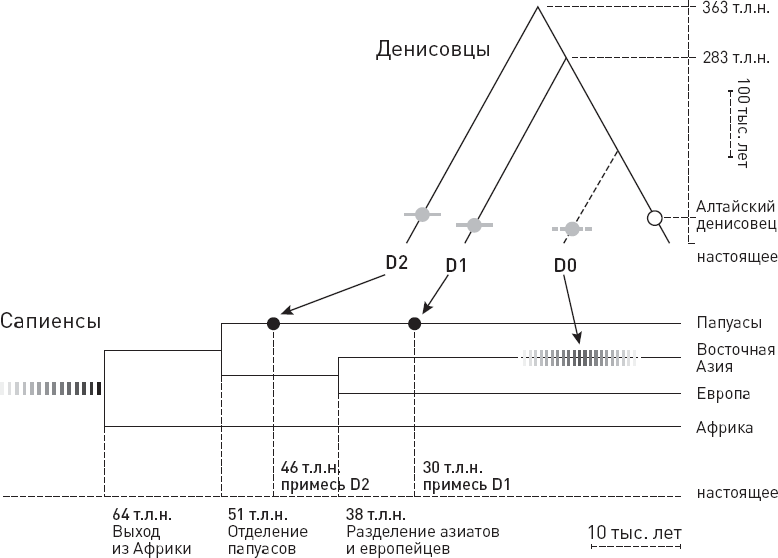
Рис. 6.3. Схема, отражающая последовательность скрещиваний разных групп сапиенсов с тремя группами денисовцев. По рисунку из Jacobs G. S. et al., 2019.
Пожалуй, самое интересное в этой истории то, что популяция D1, отделившаяся от предков алтайских денисовцев без малого 300 тыс. лет назад, скрещивалась с предками папуасов, по-видимому, прямо на Новой Гвинее, да еще и совсем недавно – всего 30–15 тыс. лет назад! На это указывает, в частности, распределение денисовских примесей D1 и D2 по территории Новой Гвинеи и близлежащих островов. Полученная раньше (46 тыс. лет назад) примесь D2 распределена равномерно, а более поздняя примесь D1 присутствует у жителей главного острова Новой Гвинеи в большем количестве, чем у аборигенов расположенной неподалеку Новой Британии, которые отделились от новогвинейцев, судя по генам, 15 тыс. лет назад. Все выглядит так, будто денисовцы жили на Новой Гвинее бок о бок с сапиенсами и продолжали скрещиваться с ними уже после того, как часть местных сапиенсов перебралась на Новую Британию (Jacobs G. S. et al., 2019).
Если все это правда, то получается, что денисовцы, во-первых, были неплохими мореплавателями. Во-вторых, они продержались на этом свете дольше, чем все остальные виды людей, за исключением сапиенсов (неандертальцы вымерли, по последним данным, 40 тыс. лет назад, флоресские хоббиты – около 50 тыс. лет назад). Еще они могли как минимум видеть древние пещерные рисунки на Сулавеси и других островах, о которых мы, правда, еще не рассказали, но скоро расскажем (так и видишь туристическую группу денисовцев, прибывших поглазеть на современную живопись народов Юго-Восточной Азии). И еще: получается, что вытеснить или истребить флоресских хоббитов в принципе могли не сапиенсы, а денисовцы… В общем, простор для фантазий и дальнейших исследований открывается широкий.
Назад: Предки алтайских неандертальцев скрещивались с древними сапиенсами, а предки денисовцев – с эректусами
Дальше: В геномах поздних европейских неандертальцев не нашли следов гибридизации с сапиенсами

