Книга: Восхождение на гору Невероятности
Назад: Глава 4 Взлет разрешен
Дальше: Глава 6 Музей Всех Возможных Раковин
Глава 5
Сорокаполосный путь к просветлению
Животным приходится иметь дело с окружающей средой и с другими живыми и неживыми телами. Они ходят по ним, ползают под ними, стараются избежать контакта или спариться с ними, ловят их и едят, убегают от них. На заре геологической истории и эволюции животные вступали в физический контакт с другими телами раньше, чем догадывались о том, что рядом есть кто‐то или что‐то. Перед тем первым счастливчиком, у которого выработалось дистанционное сенсорное восприятие, открывались огромные перспективы – он мог заметить препятствие раньше, чем налетал на него, увидеть хищника раньше, чем тот успевал его схватить, раздобыть где‐то еду, даже если ее не оказывалось в непосредственной близости. Какие же высокие технологии могли дать ему подобный бонус?
Не только химические шестеренки живой природы пришли в движение от энергии солнца. Она запустила и процесс дистанционного поиска. Каждый квадратный миллиметр нашей планеты подвергся бомбардировке лавиной фотонов – элементарных частиц, которые с максимально возможной в нашем мире скоростью разлетаются по прямым траекториям во все стороны, проникая во все щели и дырочки, не пропуская ни одного уголка и закоулка во вселенной. Стремительные и прямолинейные потоки фотонов, различия в поглощении и отражении фотонов разными веществами, а также высокая проникающая способность света – все это делает возможным развитие мощных и высокоточных технологий дистанционного слежения. Надо только зарегистрировать фотоны и, что труднее, выяснить, откуда они прилетели. Был ли использован этот шанс? Раз вы читаете мой текст, значит, спустя три миллиарда лет можете ответить на этот вопрос.
Известно, что раздел “Органы крайней степени совершенства и сложности” Чарльз Дарвин начал со слов о глазе:
В высшей степени абсурдным, откровенно говоря, может показаться предположение, что путем естественного отбора мог образоваться глаз со всеми его неподражаемыми изобретениями для регуляции фокусного расстояния, для регулирования количества проникающего света, для поправки на сферическую и хроматическую аберрацию.
Возможно, на Дарвина повлияло мнение его жены Эммы по этому вопросу. За пятнадцать лет до “Происхождения видов” он написал длинное эссе, где в общих чертах обрисовал свою теорию эволюции путем естественного отбора. Он хотел, чтобы Эмма опубликовала эту работу, если он умрет, и дал ей ее почитать. Сохранились ее заметки на полях, и любопытно, что ее внимание привлекло предположение Дарвина о том, что глаз, “с некоторой вероятностью мог сформироваться в ходе постепенного отбора незначительных, но непременно полезных отклонений в развитии”. “Прекрасное предположение / Э. Д.”, – прокомментировала Эмма. Много позже выхода в свет “Происхождения видов” Дарвин признался в письме американскому коллеге: “Меня по сей день из‐за глаза дрожь пробирает, но когда я вспоминаю о том, сколько удивительных превращений он претерпел на пути развития, мой разум велит мне унять дрожь”. Наверное, временные колебания Дарвина имели нечто общее с сомнениями того физика, которого я цитировал в начале 3 главы. Но для Дарвина сомнения стали поводом не отступиться от своих идей, а размышлять дальше.
Надо сказать, что, говоря о некоем глазе, мы недооцениваем глубину проблемы. По достоверным оценкам ученых, глаза эволюционировали не менее сорока, а возможно и больше шестидесяти раз, независимо в разных филогенетических группах царства животных. В конструкции разных типов глаз часто используются совершенно различные принципы. Если анализировать те самые сорок или шестьдесят независимо эволюционировавших глаз, можно обнаружить девять принципиально разных схем строения. Далее я расскажу о некоторых из девяти типов строения глаза – мы будем рассматривать это как девять отдельных пиков в разных частях горного массива Невероятности.
Кстати, откуда мы вообще знаем, что у двух разных групп животных какая‐либо структура эволюционировала независимо? Скажем, откуда нам знать, что у летучих мышей и птиц крылья развивались независимо? Среди млекопитающих настоящие крылья есть только у летучих мышей. Теоретически древние млекопитающие могли иметь крылья, однако все, кроме летучих мышей, их потеряли. Но для этого должно было реализоваться невообразимое множество не связанных друг с другом сценариев исчезновения крыльев, чего, как подсказывает здравый смысл – и что подкреплено массой доказательств, – не произошло. Древним млекопитающим, как и большинству их потомков, передние конечности служили не для полетов, а для передвижения по земле. Следуя той же логике, мы можем предположить, что и глаза не раз заново возникали в животном мире, и каждый случай был уникальным. Мы владеем и другой информацией – нам известны детали развития глаза у эмбрионов различных животных. Например, и у лягушек, и у головоногих моллюсков имеются полноценные глаза камерного типа, но у их эмбрионов эти органы образуются настолько по‐разному, что мы можем быть абсолютно уверены – эти глаза эволюционировали независимо. Отсюда не следует, что у общего предка лягушки и кальмара вовсе отсутствовали глаза. Не удивлюсь, если выяснится, что еще миллиард лет назад общий предок всех выживших животных был зрячим. Возможно, у него были какие‐то точечные скопления светочувствительного пигмента и он отличал день от ночи. Но глаза как высокотехнологичные приборы, способные формировать изображение, независимо прошли множество стадий эволюции. Иногда глаза разных животных конвергировали к какой‐то одной схеме строения, а иногда приходили к абсолютно уникальному дизайну. Совсем недавно появились новые, впечатляющие факты, касающиеся независимой эволюции глаза в разных группах животных. Мы еще вернемся к ним в конце главы.
Анализируя глаза животных, я во многих случаях буду указывать, на каком склоне горы Невероятности следует искать тот или иной тип глаз. Но не забывайте о том, что речь идет о современных животных, а не об их далеких предках. Разумно предположить, что по ним можно судить о строении глаз древних животных. По крайней мере, это свидетельствует о том, что те глаза, которые мы оставляем на полпути к вершине горы Невероятности, могли выполнять свои функции. Это важно, так как, повторяю, ни одно животное ни в одной ветви эволюции не имело статуса промежуточного звена. То, что мы принимаем за краткий привал во время восхождения к вершине более развитого глаза, для самого животного могло быть жизненно важным органом, и скорее всего, при его образе жизни лучшего и не требовалось. Например, совсем крохотному существу ни к чему глаза, способные формировать изображение высокой четкости. Глаза высшего качества должны быть не меньше определенного размера, чем больше – тем лучше, и не относительно размеров самого животного, а в абсолютном выражении. Крошечному существу, наверное, слишком дорого далось бы развитие таких глаз, да и не под силу было бы носить массивный, объемистый орган зрения. Если бы улитке достались такие же зоркие глаза, как у человека, вид у нее был бы весьма несуразный (рис. 5.1). Может статься, улитки с глазами хоть капельку крупнее средних будут видеть лучше своих конкуренток. Но они и поплатятся за это – придется таскать на себе более тяжелую ношу, и, следовательно, их шансы на выживание станут меньше. Диаметр самого большого глаза, известного науке, равен аж 37 см. Тот могучий великан, которому принадлежит этот рекорд, – гигантский кальмар с десятиметровыми щупальцами.
Давайте спустимся вниз по склону Зрения, не забывая об ограничениях, принятых в концепции горы Невероятности. Мы обнаружим там глаза до того примитивные, что они даже не заслуживают права так называться. Собственно, поверхность всего тела слабо реагирует на свет. Например, некоторые одноклеточные, медузы, морские звезды, пиявки и разные виды червей не способны формировать изображение или хотя бы распознавать, откуда идет свет. Они лишь чувствуют, и то смутно, что где‐то поблизости есть источник света – яркого света. Вы не поверите, но есть неопровержимые доказательства наличия чувствительных к свету клеток на гениталиях бабочек, как самцов, так и самок. Это не глаза, которые формируют изображение, но разницу между светом и темнотой они ощущают – и могут послужить той самой отправной точкой, которую мы имеем в виду, когда говорим о начале эволюции глаза в древности. Никто не знает, как бабочки оперируют этим органом, даже Уильям Эберхард, из чьей очень занимательной книги под названием “Половой отбор и половые аппараты животных” (Sexual Selection and Animal Genitalia) я почерпнул эти сведения.
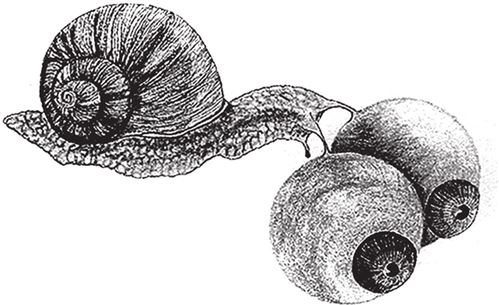
Рис. 5.1. Фантастическая улитка видела бы так же хорошо, как человек, если бы у нее были такие огромные глаза.
Если допустить, что в долине у подножия горы Невероятности обитают предшественники животных, абсолютно не восприимчивые к свету, то светочувствительные, но не способные определить источник света кожные покровы морских звезд и пиявок, равно как и половой аппарат бабочек, уже ведут нас чуть выше по склону, к началу горной тропы. Найти тропу легко. И впрямь, долина Абсолютной невосприимчивости к свету, возможно, никогда и не была такой уж необъятной. Может быть, все живые клетки в той или иной степени могут реагировать на свет – и тогда светочувствительные половые органы бабочек уже не должны вызывать недоумение. Луч света – это прямолинейный поток фотонов. В результате соударения с молекулой окрашенного вещества фотон может прекратить свой полет, а молекула пигмента может перейти в другое состояние, хотя останется той же молекулой. При этом выделяется энергия. Зеленые растения и бактерии используют эту энергию в серии реакций – фотосинтезе – для производства молекул, которыми они питаются. У животных эта энергия может спровоцировать реакцию в нервной клетке, и это можно считать первой стадией процесса, который мы и называем “зрение”, даже если у животного нет глаз в нашем понимании. На начальном уровне сгодится любой из широчайшей палитры пигментов (окрашенных веществ). Существует огромное множество таких пигментов, которые используются далеко не только для поглощения света. Вероятно, постепенное усовершенствование молекул пигментов позволило сделать первые робкие шаги вверх на гору Невероятности. Это пологий склон непрерывных усовершенствований, по нему легко подниматься маленькими шажками.
Низинный пандус ускорил путь к эволюции живого аналога фоторецептора – клетки, специализирующейся на улавливании фотонов пигментом и преобразующей выделяющуюся энергию в нервные импульсы. Клетки сетчатки (у нас это палочки и колбочки), которые специализированы для улавливания фотонов, я буду впредь называть фоторецепторами. Все они используют один мудрый прием – увеличение числа слоев пигмента, способного улавливать фотоны. Это существенно, ибо очень вероятно, что через один слой фотон пролетит, не претерпев ни малейших изменений. Чем больше пигментных слоев, тем больше шансов изловить хотя бы один фотон. Какая разница, сколько фотонов будет поймано, а сколько проскочит насквозь? Разве нет у нас достаточного их запаса? Нет, и это ключевой момент для понимания того, как устроены глаза. В своего рода “экономике” фотонов действуют те же монетарные отношения, основанные на стремлении хапнуть побольше и отдать поменьше, с такими же неизбежными потерями и компромиссами, как в нашем обществе.
Даже не вдаваясь в нюансы экономических выгод и потерь, мы точно знаем, что иногда фотонов попросту не хватает. Однажды звездной, холодной ночью 1986 года я разбудил свою двухлетнюю дочь Джульет, укутал ее в одеяло, вынес в сад и повернул сонным личиком туда, где, как сообщалось, должна была пролетать комета Галлея. Она не понимала моей речи, но я упорно шептал ей на ушко про комету и про то, что мне уже точно не доведется увидеть ее во второй раз, а вот самой Джульет еще выпадет шанс, когда ей будет семьдесят восемь лет. Я разбудил ее, объяснял я дочке, чтобы в 2062 году она могла сказать внукам, что видала эту комету раньше, а может, и вспомнила бы своего папу с его восторженной блажью, который потащил ее в ночной сад любоваться кометой Галлея. Кажется, я так и сказал – восторженная блажь, – потому что маленьким детям нравятся непонятные слова, если их четко произнести.
Наверное, тогда, в 1986 году, какие‐то фотоны достигли сетчатки Джульет, но, честно говоря, мне и себя‐то нелегко было убедить, что я видел комету. Иногда мне мерещилось какое‐то едва различимое сероватое облачко примерно там, где она должна была пролетать. Потом оно исчезало. Беда в том, что на сетчатки наших глаз попадало близкое к нулю количество фотонов.
Фотоны долетают до нас спорадически, как капли дождя. Если дождь льет вовсю, мы точно это видим и уповаем на то, что никто не прихватил с собой наш зонтик. Но когда он только-только начинается, как определить момент его начала? Мы чувствуем одну каплю и задумчиво глядим на небо – будет вторая и третья или нет. При столь незначительных осадках один человек скажет, что пошел дождь, а другой ему возразит. Капли падают редко, так что один человек ощутит касание за минуту до того, как другая капля упадет на его соседа. Чтобы не сомневаться в том, светло вокруг нас или темно, частота попадания фотонов на сетчатку должна быть достаточно высокой. Надо полагать, фотоны кометы Галлея, когда мы с Джульет устремили взоры предположительно в ее сторону, достигали отдельных фоторецепторов наших сетчаток с низкой до обидного частотой – может, по одному за сорок минут! То есть от одного какого‐нибудь фоторецептора можно было получить сигнал, что свет есть, в то время как колоссальное множество соседних фоторецепторов никаких сигналов не передавали. У меня вообще возникло чувство, что я видел некий объект, похожий на комету, лишь благодаря моему мозгу, который подытожил вердикты, вынесенные сотнями фоторецепторов. Два фоторецептора ловят больше фотонов, чем один. Три больше, чем два – и так далее вверх по склону горы Невероятности. В столь совершенных глазах, как наши, плотно, как ворс ковра, упакованы миллионы фоторецепторов, и каждый настроен ловить как можно больше фотонов.
На рис. 5.2 изображен типичный, в данном случае человеческий, фоторецептор, но другие устроены почти так же. Клубок червячков в центре рисунка – это митохондрии, внутриклеточные органеллы. Они произошли от бактерий-паразитов, но стали незаменимыми элементами клетки, без которых не вырабатывается энергия. За левое поле рисунка уходит нервное окончание. Ровный, красивый ряд мембран в правой части, выстроившихся, словно на параде, и есть та самая часть рецептора, куда попадают фотоны. В эти слои встроены молекулы жизненно важного пигмента, поглощающего фотоны. На этом рисунке я насчитал 91 мембранный слой. Неважно, сколько именно слоев – когда речь идет об улавливании фотонов, лишним ни один не будет, хотя их количество лимитируется общими затратами организма. Идея в том, что 91 мембрана эффективнее ловит фотоны, чем 90, 90 эффективнее 89 и так далее вплоть до одиночной мембраны, чей КПД все‐таки выше нулевого. Вот что я имею в виду, когда говорю о плавном подъеме на гору Невероятности. Резкий перепад высоты встретился бы нам в том случае, если бы существовало некое пороговое значение – скажем, до сорока пяти слоев мембраны работали бы вообще не эффективно, а после сорока пяти очень эффективно. Здравый смысл выступает против таких непредвиденных скачков, да и нет фактов, свидетельствующих о том, что они могли бы быть.
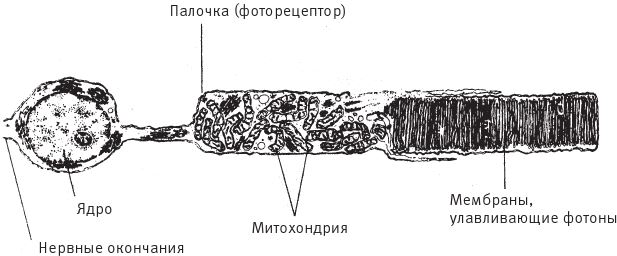
Рис. 5.2. Фоторецептор, единичная клетка сетчатки человека, – биологический “фотоэлемент” для улавливания фотонов.
Как мы уже видели, у головоногих моллюсков независимо от позвоночных развились похожие глаза. Даже фоторецепторы у них почти такие же. Разница лишь в том, что у кальмаров мембрана состоит не из дисков, а из столбика колец на полой трубке. В эволюции полно таких несущественных отличий, в которых не больше логики, чем, скажем, в устройстве выключателя: в Англии при опущенном тумблере свет зажигается, а в Америке – выключается. У всех животных хорошо развитые фоторецепторы пытаются любым способом обзавестись дополнительными мембранами с пигментом – и пусть фотон попробует через них проскочить. В рамках концепции горы Невероятности важно, что каждый новый слой – сколько бы их уже ни было – хоть немножко, да повышает шансы заловить фотоны. В конце концов захватывается уже львиная доля фотонов, и начинает действовать закон убывающей доходности для роста затрат на прибавку слоев.
Конечно, в природе редко у кого возникает нужда отследить комету Галлея с ее ничтожно слабым потоком фотонов, которая к тому же появляется раз в семьдесят шесть лет. Однако зоркие глаза, способные видеть при лунном свете и даже при свете звезд – если вы сова, – весьма полезная вещь. Обычно ночью один наш фоторецептор улавливает примерно по фотону в секунду, то есть существенно быстрее, чем при наблюдении за кометой, но все равно слишком медленно для полного поглощения всех фотонов до единого – если это вообще возможно. Но коли уж мы заговорили о суровых экономических законах в применении к фотонам, было бы большой ошибкой полагать, что они действуют только ночью. В ясный солнечный день тоже не все так просто, хотя на сетчатку может обрушиться настоящий фотонный ливень. Формирование отчетливого, детального изображения сводится к тому, что на разных участках сетчатки фоторецепторы реагируют на свет разной интенсивности, а значит, частота попаданий фотонов, из которых складываются отдельные струи фотонного ливня, тоже должна быть разной. Мелкозернистое изображение предполагает распределение фотонов, а это может приводить к таким же значительным локальным ослаблениям их потока, как и сокращение его вообще в ночное время. Вот этим распределением мы сейчас и займемся.
Сами по себе фоторецепторы сообщают животному лишь о том, светло вокруг него или темно. Животное отличает день от ночи и понимает, с какой стороны падает тень – например, может вовремя заметить приближение хищника. На следующей, более высокой стадии развития, должна была выработаться способность определять направление света и перемещения, скажем, зловещей тени. Для этого проще всего закрасить одну сторону фоторецептора темной краской. Прозрачный фоторецептор без покрытия одной из сторон пропускает свет отовсюду и не несет информации о местонахождении его источника. Животное с единственным фоторецептором в голове, затемненным с одной стороны, может идти на свет или от него. Определить направление немудрено – надо поворачивать голову, как маятник, то в одну сторону, то в другую; если интенсивность света с разных сторон различна, надо изменить направление так, чтобы уравнять яркость света. Некоторые личинки удирают от света, руководствуясь этим правилом.
Впрочем, вертеть головой, пытаясь понять, откуда льется свет, можно в самом начале восхождения на гору Невероятности, на самом примитивном уровне. Хорошо бы иметь побольше затемненных сзади фоторецепторов, направленных в разные стороны. Тогда можно будет сравнить частоты попадания фотонов на два фоторецептора и догадаться, где находится источник излучения. Если же есть целый коврик из фоторецепторов с закрашенным задником, то лучше всего согнуть его полусферой, так чтобы каждый его элемент всегда был ориентирован одинаково. Из выпуклой поверхности в конечном итоге может получиться фасеточный глаз, как у насекомых, и позже мы его рассмотрим. Поверхность, вогнутая в виде чаши, образует другой типичный орган зрения – наш, человеческий, глаз камерного типа. Поступающий с разных направлений свет активизирует разные фоторецепторы, и чем их больше, тем тоньше структура изображения.
На пути световых лучей (параллельные белые линии со стрелками на рис. 5.3) встает толстый черный экран, выстилающий заднюю поверхность камеры-чаши. Мозг определяет, где находится источник света, по тому, какие фоторецепторы включаются в работу. Для нас с нашей теорией о горе Невероятности важно то, что путь наверх пролегает по пологому склону – животных, чьи фоторецепторы расположены в плоскости и на вогнутой/выпуклой поверхности, связывает длинная череда плавных эволюционных переходов. Мало-помалу, постепенно, кривизна чаши может меняться в ту или иную сторону. Чем сильнее кривизна, тем увереннее глаз различает лучи света, которые падают под разными углами. По дороге к вершине вам не придется запрыгивать на высокие уступы.
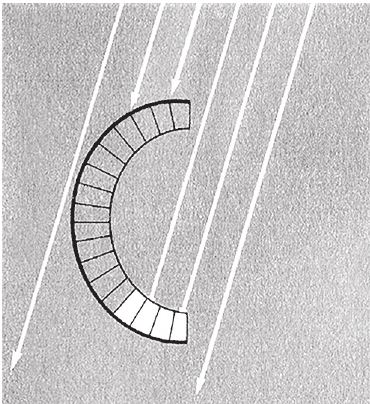
Рис. 5.3. Примитивный глаз типа чаши способен определить направление света.
Такими глазами обладают животные. На рис. 5.4 показаны глаза морского блюдечка, полихеты (многощетинкового червя), двустворчатого моллюска и плоского червя. Вероятно, форму чаши эти глаза приобрели независимо друг от друга. Так, строение глаза плоского червя, бесспорно, говорит о его независимой эволюции – фоторецепторы находятся во внутреннем слое чаши. На первый взгляд, какая‐то нелепица – прежде чем свет попадет к фоторецепторам, он должен пройти сквозь толщу соединяющих нервов, – но нам ли чваниться, ведь и наши, куда более чувствительные и высокоразвитые, органы зрения страдают тем же пороком. Я еще вернусь к этому вопросу и покажу, что здесь не все так неумно, как кажется.
В любом случае сам по себе глаз-чаша не способен формировать хорошее изображение – хорошее в нашем понимании, то есть удовлетворяющее нашему придирчивому глазу. Тут надо пояснить, как формируется изображение в глазу человека по законам оптики. Чтобы разобраться в этой проблеме, давайте выясним, почему без дополнительных органов слой фоторецепторов в вогнутой чаше (камере) не покажет, скажем, дельфина, даже если этот самый дельфин находится прямо перед ними.
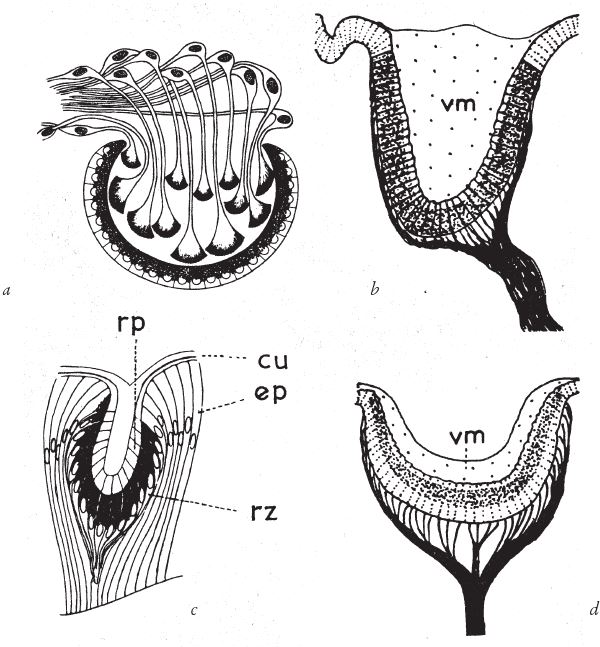
Рис. 5.4. Глаза камерного типа разных животных: (а) плоского червя; (b) двустворчатого моллюска; (с) многощетинкового червя полихеты; (d) моллюска блюдечко.
vm – стекловидная масса; cu – кутикулярная линза; еp – эпителий; rz – сетчатка с фоторецепторными клетками.
Если бы световые лучи шли так, как показано на рис. 5.5, все было бы проще простого и на сетчатке сформировалось бы прекрасное изображение дельфина. Увы, реальная картина иная. Точнее, некоторые лучи проходят именно так, как я нарисовал. Загвоздка в том, что их перекрывают потоки других лучей, которые разлетаются во всех направлениях одновременно с теми, что нас интересуют. На каждую единицу площади сетчатки попадают лучи, отраженные от каждой единицы поверхности дельфина. И не только его, но и всего остального, что находится за дельфином и вокруг него. Тогда, подумаете вы, должно получиться бесчисленное множество изображений дельфина – во всех точках чаши и во всевозможных положениях. Однако в итоге, конечно, не будет никакого изображения – только световое пятно, размазанное по всей поверхности (рис. 5.6).
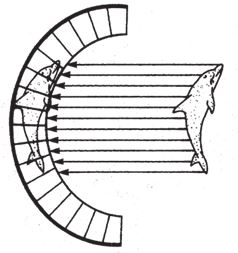
Рис. 5.5. Так не бывает. Если бы лучи света падали так ровно!
Мы обозначили проблему. Глаз видит слишком много – не одного дельфина, а многотысячную стаю. Напрашивается решение: одного дельфина оставить, всех остальных ликвидировать. Неважно, какого выбрать – главное, как избавиться от остальных? Можно попробовать перелезть на тот же склон горы Невероятности, по которому мы добрались до чаши, планомерно углубляя чашу и сужая вход, пока он не превратится в крохотную дырочку. Тогда львиная доля светового потока пройдет мимо камеры глаза. Останется лишь малая часть – та, что формирует небольшое количество одинаковых картинок перевернутого дельфина (рис. 5.7). При очень маленьком входном отверстии можно будет избежать размытости контуров и сформируется единственное, четкое изображение дельфина (на самом деле чересчур узкое отверстие становится причиной размытости другого рода, но сейчас не будем на это отвлекаться). Крохотная дырочка играет роль фильтра изображений, который позволяет прекратить дикую свистопляску в глазах и видеть только одного дельфина.
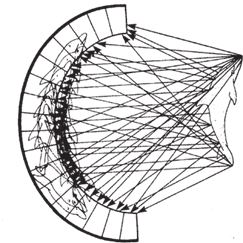
Рис. 5.6. Световые лучи из каждой точки расходятся во все стороны, изображение не формируется. Из-за наложения бесчисленного множества изображений дельфина отчетливой картинки не будет.
Эффект крохотного входного отверстия – это предельный случай уже знакомого нам эффекта чаши, благодаря которому можно понять, откуда поступает свет. Оно занимает место на том же склоне горы Невероятности, чуть повыше, и между этими двумя уровнями нет резких перепадов высоты. Эволюционный путь от глаза-чаши к узкому входу в камеру глаза ничем не затруднен, так же как ничто не мешает плоскому слою фоторецепторов эволюционировать в глаз-чашу. Путь от плоского слоя к маленькому входному отверстию не утомителен. Вы как бы отсекаете одну картинку за другой, пока не доберетесь до вершины, где останется единственное изображение.
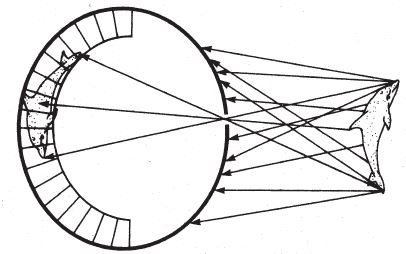
Рис. 5.7. Принцип действия глаза с маленьким отверстием. Конкурирующих изображений дельфина больше нет. Попавшие в отверстие лучи формируют одно четкое перевернутое изображение.
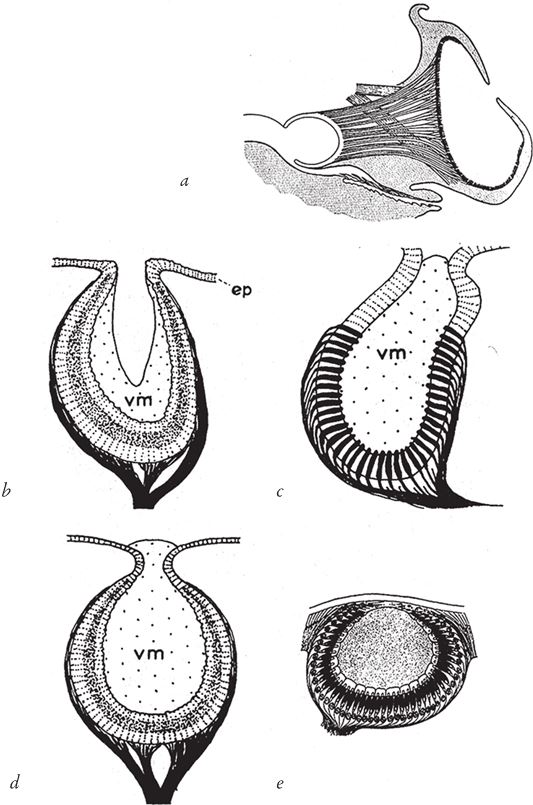
Рис. 5.8. Путь к формированию неидеального, но достаточно четкого изображения. Глаза беспозвоночных: (а) узкое отверстие моллюска Nautilus; (b) морской улитки; (с) двустворчатого моллюска; (d) морского ушка; (е) нереиды.
vm – стекловидная масса; еp – эпителий
Различные варианты глаз с точечным входным отверстием широко распространены в животном мире. Наиболее выражено это свойство у загадочного моллюска наутилуса (рис. 5.8а), родственника ископаемых аммонитов (и более далекого кузена осьминога, только с витой раковиной). Глаза других животных – например, морского моллюска (рис. 5.8b), пожалуй, ближе к глубокой чаше, чем к узкому отверстию. Все они служат наглядным подтверждением того, что данный склон горы Невероятности достаточно удобен для восхождения.
Первое, что приходит в голову – если входное отверстие достаточно мало, глаз должен функционировать хорошо. При микроскопическом отверстии почти все лишние изображения, накладывающиеся на основное, отсекаются, и вроде бы должно получиться почти безупречное изображение. Но тут возникают два новых препятствия. Одно из них – дифракция. Пока не будем ее учитывать. Проблема в том, что свет имеет волновую природу, и волны интерферируют, т. е. накладываются друг на друга, из‐за чего изображение становится нечетким. При очень малом диаметре отверстия резкость падает сильнее. Вторая трудность связана с “экономикой фотонов” – из‐за малых размеров отверстия приходится делать непростой выбор. Если дырочка достаточно мала для того, чтобы изображение получилось резким, через нее поступает такой слабый поток света, что увидеть предмет можно только при очень ярком – недостижимо ярком – освещении. При обычном уровне освещенности сквозь входное отверстие в глаз попадет слишком мало фотонов, и вы не поймете, что же вы видите. С такой крошечной дырочкой мы оказываемся в той же ситуации, что и с кометой Галлея. Чтобы решить возникшую проблему, можно опять расширить вход. Но так вы вернетесь туда, где уже были – к путанице с многочисленными изображениями дельфина. Экономика фотонов завела нас в тупик и оставила все у того же подножия горы Невероятности. Узкий вход в камеру глаза дает либо четкое, но темное изображение, либо яркое, но расплывчатое. И того и другого одновременно мы не получим. Экономисты обожают подобные игры с компромиссами, поэтому я и предложил экономическую схему для фотонов. Но неужели никак нельзя добиться яркого и вместе с тем отчетливого изображения? По счастью, можно.
Для начала попробуем все рассчитать. Допустим, мы расширим малюсенькую дырочку и увеличим силу входящего светового потока. Но мы не оставим ее открытой, а закроем “волшебным окном” – умным электронным устройством, запаянным в стекло и подключенным к компьютеру (рис. 5.9). Управляемое с помощью компьютера стеклышко имеет одно свойство. Попадающий в него луч света выходит не прямо, а под определенным углом. Компьютер подбирает угол так, чтобы все лучи, исходящие из одной точки (например от кончика дельфиньего носа), после преломления сходились в соответствующей точке на сетчатке. Я нарисовал только те лучи, что идут от носа, но пред волшебным окном все точки, естественно, равнозначны, и для каждой выполняются одинаковые расчеты. Лучи, исходящие от кончика хвоста, преломляются и встречаются на сетчатке в соответствующей точке хвоста – и так для всех точек. Эффект волшебного окна – четкое изображение дельфина на сетчатке. Но благодаря тому, что во всех точках носа, хвоста и вообще всего тела дельфина на сетчатке сошлись целые пучки лучей (то есть, целые потоки фотонов), изображение получилось не такое темное, как если бы лучи непосредственно прошли сквозь узкое отверстие. Волшебное окно обладает плюсами узкого входного отверстия, но лишено его недостатков.
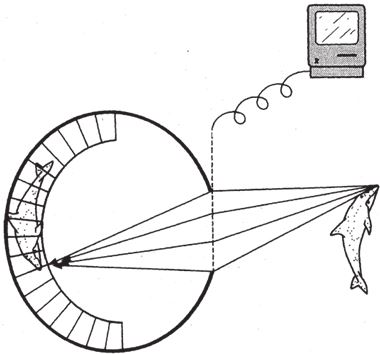
Рис. 5.9. “Компьютерный хрусталик” – сложный и крайне нерациональный гипотетический способ добиться резкого и одновременно яркого изображения.
Хорошо вот так взять и сотворить из ничего “волшебное окно” в своем воображении. Легко сказать – а сделать? Подумайте о том, какие сложные расчеты выполняет подключенная к волшебному окну машина. Компьютер обрабатывает данные о миллионах световых лучей, исходящих из миллионов всевозможных точек. Каждая частица поверхности дельфина посылает миллионы лучей под миллионом разных углов во все точки поверхности волшебного окна. Лучи пересекаются, словно прямые трассы на путаных многоуровневых дорожных развязках. Волшебное окно и соединенный с ним компьютер должны пропустить через себя все лучи по очереди и для каждого точно рассчитать свой угол преломления. Где раздобыть такой прекрасный компьютер – разве что обратиться к волшебнику? Неужели нас ждет горькое разочарование, неужели непреодолимое препятствие преградило нам путь к вершине?
Не все так печально. Компьютер на рисунке – всего лишь гипотетический пример того, сколь неразрешимой может показаться задача, если рассматривать ее только с одной стороны. Но если подойти к проблеме с другого бока, можно найти до смешного простой выход. Есть, оказывается, нехитрая вещь, которая обладает свойствами нашего волшебного окна и работает без компьютера, без высокотехнологичной электронной начинки и вообще без каких‐либо дополнительных условий. Это хрусталик. Компьютер больше не понадобится, потому что ничего не надо будет вычислять. Кусочек прозрачного материала с закругленными поверхностями автоматически, мгновенно выполняет сложнейшие расчеты углов для миллионов лучей. Прежде чем показать вам, что эволюция хрусталика должна была пройти без серьезных помех, я отвлекусь на небольшую прелюдию и объясню принцип действия линз.
По законам физики лучи, попадающие из одной прозрачной среды в другую, меняют направление (рис. 5.10). Угол выхода луча зависит от природы обеих сред, поскольку разные вещества имеют различные показатели преломления – этой величиной измеряется способность вещества менять направление световых лучей. У воды и стекла показатели преломления почти одинаковы, и в этом случае при переходе из одной среды в другую ход луча меняется незначительно. На границе стекла с воздухом луч преломляется гораздо сильнее, потому что воздух имеет относительно низкий показатель преломления. Излом луча на границе воды и воздуха настолько явственный, что нам кажется, будто весло погнулось.
На рис. 5.10 показан стеклянный кубик в воздухе. Жирная линия – это луч, который входит в кубик, преломляется, проходит его насквозь и вновь выходит под тем же углом, под каким вошел. Но, конечно, кусок прозрачного материала вовсе не обязательно имеет ровные, параллельные грани. В зависимости от кривизны поверхности луч может пойти как угодно. А если поверхность кубика состоит из множества лежащих под разными углами граней, пучок лучей может разойтись во все стороны (рис. 5.11). Выпуклый с одной стороны или двояковыпуклый кусок прозрачного материала представляет собой линзу и работает точно так же, как наше волшебное стекло. В природе достаточно прозрачных материалов. Вода и воздух, основные вещества на нашей планете, тоже прозрачные. Прозрачны и многие жидкости. Подойдут и кристаллы, отшлифованные морской волной почти до идеальной гладкости. Вспомните округлые прозрачные камешки различной формы, которые отполировал прибой. В зависимости от углов на поверхности такой камешек разбрасывает световые лучи от одного и того же источника в разные стороны. Форма камешка может быть любой. Чаще всего встречаются двояковыпуклые. Как они могут повлиять на ход световых лучей, падающих от одного источника – например, от лампочки?
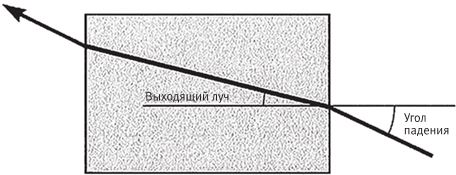
Рис. 5.10. Преломление луча в стеклянном кубике.
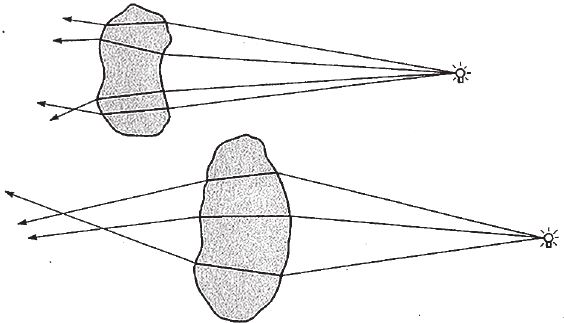
Рис. 5.11. Необработанные камешки преломляют лучи как попало.
Пройдя сквозь кристалл с сильно выгнутыми сторонами, лучи сойдутся. Правда, не настолько точно, чтобы сформировалось четкое изображение источника света, как в нашей модели с воображаемым “волшебным стеклом”. На это можно только надеяться. Но тенденция к правильному ходу лучей будет заметна. Любой гладкий, округлый с обеих сторон обломок кварца будет работать не хуже “волшебного стекла” и, как самая настоящая линза, даст пусть нерезкие, но куда более яркие изображения, чем это позволяет сделать крохотная дырочка. На самом деле природные камешки, обработанные водой, почти всегда округлые и двояковыпуклые. Будь они прозрачными, многие из них могли бы послужить если не самыми красивыми, то вполне пригодными линзами.
Галька – лишь один из природных объектов, которые приобрели свойства линзы чисто случайно, без чьего‐либо умысла. Есть и другие. Повисшая на листике капля воды имеет округлую форму. Всегда, без исключения. Она могла бы стать примитивной линзой сама собой, без нашего вмешательства. Если нет противодействия каких‐нибудь сил, в частности силы тяжести, жидкости и гели неизбежно примут форму шарика. Зачастую линзы из них получаются просто потому, что иначе и быть не может. Так же ведут себя и многие другие биологические субстанции. Молодые медузы напоминают линзу как формой, так и изумительной прозрачностью. Они могут заменить собой линзы, хотя им самим это никакой пользы не принесет, и нет никаких оснований полагать, что естественный отбор благоприятствовал развитию у медуз свойств линзы. Прозрачная медуза не так заметна для врагов, что, наверное, дает преимущество, а плюсы округлой формы связаны вовсе не с оптикой, а со строением тела.
С помощью разных подручных средств, не предназначенных для этой цели, я получил кое‐какие картинки. На рис. 5.12а вы видите большую букву А, отображенную на листе бумаги, который я поместил на заднюю стенку камеры-обскура – глухой картонной коробки с маленьким отверстием на одной из сторон. Без подсказки вы, возможно, и не опознали бы букву. Чтобы она получилась достаточно яркой и ее вообще можно было бы разглядеть, я просверлил довольно большую дырку, диаметром примерно в сантиметр. Я мог бы сделать ее поменьше, и тогда картинка получилась бы более резкой, но не хватило бы чувствительности пленки: мы снова вынуждены идти на тот же компромисс.
Теперь посмотрим, как можно добиться совершенно иного результата даже с самыми нехитрыми “линзами”, вовсе не рассчитанными на такое применение. На рис. 5.12b изображена та же буква А на задней стенке той же коробки, снятая сквозь ту же дырочку. Но на этот раз я подвесил перед отверстием полиэтиленовый пакет с водой. Производители пакета не думали, что из него сделают линзу. Но если налить в него воды и подвесить, по естественным причинам он примет округлую форму. Пожалуй, если бы медуза была не такой складчатой, а более гладкой, она дала бы изображение даже более высокого качества. Надпись на рис. 5.12с (CAN YOU READ THIS?) получена с помощью той же продырявленной коробки, но перед отверстием я поместил уже не пакет, а круглый бокал с водой. Бокал для вина – безусловно, творение рук человеческих, но в намерения дизайнера не входило делать линзу, и круглую форму ему придали с иными целями. И вновь вещь, которую создавали вовсе не для этого, оказалась более или менее сносной линзой.
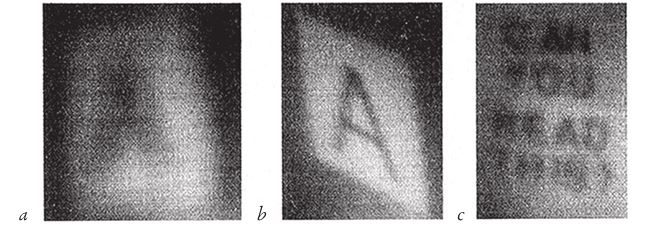
Рис. 5.12. С помощью дырочки и самодельных заменителей линз можно получить вот такие изображения: (а) дырочка без дополнительных приспособлений; (b) с полиэтиленовым пакетом, в который налили воду; (с) с бокалом, в который налили воду.
Конечно, древние животные не пользовались ни полиэтиленовыми пакетами, ни стеклянными бокалами. Я и не предполагаю, что на одном из этапов эволюции глаза использовались как полиэтиленовые мешки или картонные коробки. Смысл всех этих примеров – с пакетом, дождевыми каплями, медузами и кварцевой галькой – сводится к тому, что ни один из перечисленных предметов и природных объектов не должен был заменять собой линзу. Нашлось достаточно иных веских причин, по которым они приняли форму линзы.
Стало быть, природные объекты запросто могут без чьего‐либо вмешательства приобрести свойства примитивной линзы. Если полупрозрачная студенистая масса примет округлую форму (почему – неважно), можно надеяться, что чаша и узкое отверстие будут работать с ней уже лучше. Чтобы хоть немного приблизиться к вершине горы Невероятности, всего‐то и требуется, что слегка повысить эффективность органа. А как могли выглядеть промежуточные варианты? Обратимся еще раз к рис. 5.8 – и еще раз я должен подчеркнуть, что здесь речь идет о современных животных, по ним нельзя выстраивать подлинные генеалогические линии. Обратите внимание на прозрачный слой “стекловидной массы” (vm – vitreous mass, рис. 5.8b), выстилающей полость чаши, – по‐видимому, он защищает чувствительные фоторецепторы от соленой морской воды, которая беспрепятственно попадает в глаз через входное отверстие. Этот слой несет исключительно протекторную функцию и обладает одним из обязательных для линзы свойством – прозрачностью, однако ему не хватает необходимой выпуклости и толщины. Теперь взгляните на рис. 5.8c, d и е – на глаза двустворчатого моллюска, брюхоногого моллюска морское ушко и полихеты нереидиды. Это все тоже примеры чашевидных глаз и промежуточных состояний между чашей и узким отверстием, но кроме того мы видим и значительно более толстый слой стекловидной массы. Очень многие животные имеют в той или иной степени оформленный (или бесформенный) слой стекловидной массы. Известные производители оптики не сочли бы ни один из этих шматков студенистого вещества сколько‐нибудь приличной линзой. Но тем не менее любой хотя бы немного выпуклый желеобразный прозрачный сгусток существенно увеличивает эффективность маленькой дырочки.
Между любой субстанцией вроде стекловидной массы морского ушка и качественной линзой есть одно главное различие – для того чтобы линза работала лучше, ее надо отделить и немного отодвинуть от сетчатки. Это пространство необязательно должно остаться пустым. Оно может быть заполнено студенистым стекловидным веществом. Важно, чтобы показатель преломления линзы был выше, чем у вещества, заполняющего пространство между ней и сетчаткой. Этого можно добиться разными, достаточно простыми способами. Давайте обсудим один из них – когда линза образуется из сгустка в пределах переднего отдела стекловидной массы (рис. 5.8е).
Для начала вспомним, что у любого прозрачного вещества есть характерный показатель преломления. Это мера способности вещества изменять направление светового луча. В производстве оптики показатель преломления стекла обычно принимается за постоянную величину, и от толщины слоя она не зависит. Попав в линзу и один раз изменив направление соответственно ожиданиям, до противоположной стороны линзы луч идет по прямой. Секрет производства хорошей оптики заключается в шлифовке и полировке стекла точно заданной формы и последовательном соединении разных линз.
Склеив стекла с различными характеристиками, можно изготовить составную линзу, каждый слой которой будет иметь свой показатель преломления. Так, в линзе, изображенной на рис. 5.13а, центральная часть сделана из стекла с относительно высоким показателем преломления. Здесь свойства линзы меняются дискретно. Однако нет принципиальных ограничений, которые не позволяли бы получить линзу с непрерывно меняющимся по всему ее объему показателем преломления. Такая линза показана на рис. 5.13b. Трудности при изготовлении подобной “градиентной” линзы связаны со способом обработки оптических стекол. А вот природные линзы могут быть построены таким образом, поскольку не “производятся” за одну стадию, а постепенно формируются из маленьких зачатков по мере развития животного. В сущности, линзы-хрусталики многих рыб, осьминогов и других животных имеют непрерывно изменяющийся показатель преломления. Если вы внимательно приглядитесь к рисунку 5.8е, то заметите область за входом в глаз, где, возможно, показатель преломления меняется непрерывно.
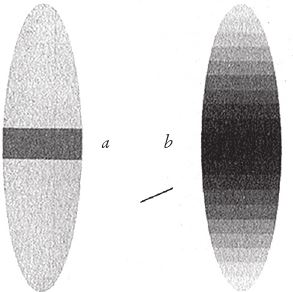
Рис. 5.13. Два типа составных линз.
Но я говорил о развитии хрусталика с нуля, из стекловидной массы, заполнившей всю полость глаза. Шведские биологи Дэн Нильссон и Сюзанна Пелгер придумали прекрасную компьютерную модель, которая наглядно показывает, как это в принципе могло бы происходить и как быстро. Я прокомментирую их элегантную идею, но зайду немного с другой стороны. Прежде чем рассказать об их эксперименте, я вернусь к нашему переходу от биоморфов к программе NetSpinner и задам такой вопрос: как вообще можно было бы подступиться к решению этой задачи – создать компьютерную модель эволюции глаза? Затем я покажу вам, что Нильссон и Пелгер проделали в сущности то же самое, хотя и не точно так же.
Вспомним, что биоморфы эволюционировали в ходе искусственного отбора по воле человека. О том, чтобы воспроизвести подлинный естественный отбор, мы и думать не могли, поэтому перешли к моделированию паутин. Это удобно, так как паутина плоская и компьютер может автоматически рассчитать ее эффективность для ловли мух. Кроме того, автоматическому расчету поддаются затраты на паутинный шелк, поэтому компьютер может автоматически “выбирать” паутину, примерно как при естественном отборе. Мы увидели, что в этом смысле паутина – уникальный объект исследования: едва ли нам удалось бы воспроизвести тот же процесс для позвоночника преследующего добычу гепарда или для хвостового плавника плывущего кита, уж очень сложно учесть все физиологические детали при оценке КПД работы трехмерного органа. Зато глаз в этом отношении подобен паутине. Компьютер сможет дать оценку двухмерной модели глаза, изображенной в плоскости. Я не утверждаю, что глаз – двухмерный объект, ибо это не так. Просто во фронтальной проекции глаз круглый, и его эффективность как объемного тела можно оценить по компьютерному изображению среднего сечения в вертикальной плоскости. Компьютер может проанализировать ход лучей и подогнать резкость изображения, которое мог бы сформировать глаз. Такая качественная оценка эквивалентна расчету эффективности смоделированной на компьютере паутины при ловле цифровых мух в программе NetSpinner.
Точно так же, как NetSpinner плодит паутины-мутанты, мы могли бы заставить модель глаза “рожать деток” – глаза с мутациями. Все глаза-детки копировали бы своего родителя внешне, но в каких‐то незначительных деталях формы могли бы спонтанно возникать отклонения. Конечно, некоторые из компьютерных “глазок” были бы совсем не похожи на нормальные глаза – и даже само слово “глаз” тут вряд ли подошло бы, – но это неважно. Они все равно воспроизведутся, а их оптические свойства получат численную оценку – скорее всего, очень низкую. Таким образом, мы могли бы усовершенствовать глаза в ходе виртуального естественного отбора, как в программе NetSpinner. За исходный образец можно было бы взять очень хороший глаз – или начать с совсем никудышного, а то и вовсе с пустого места.
Такие программы как NetSpinner, достоверно моделирующие ход эволюции, весьма информативны: вы начинаете с самого примитивного состояния и ждете финальной стадии развития. Можно прервать эксперимент на любом ключевом моменте различных направлений эволюции – это были бы альтернативные пики горы Невероятности. Мы могли бы провести эксперимент в режиме эволюции и с моделью глаза – и получили бы убедительную иллюстрацию истории. Однако в реальности эволюция модели внесет ненамного больше ясности, чем более систематический анализ пути (или путей) к вершине (или к вершинам) горы Невероятности. Естественный отбор выбирает дороги, которые выходят из конкретных точек, всегда идут вверх и никогда вниз. Если изучать модель в режиме эволюции, естественный отбор пойдет по этому пути. Поэтому, если мы будем планомерно исследовать пути к вершине и пики, которые можно покорить, выйдя из гипотетических стартовых точек, мы сэкономим машинное время. По правилам игры поворачивать вниз запрещено – это важно. Нильссон и Пелгер провели как раз такое систематическое исследование способов восхождения на гору, но вы, наверное, понимаете, почему я предпочитаю говорить об их эксперименте так, будто мы вместе с ними разрабатывали нормативные акты эволюции в духе программы NetSpinner?
Несмотря на наш выбор – работать с нашей моделью, будь то “режим естественного отбора” или “систематический анализ в режиме горы”, нам надо договориться о правилах эмбриологии – о том, как гены управляют развитием организмов. На какие внешние черты оказывают влияние мутации? Каковы будут сами мутации – с большим или малым эффектом? В случае NetSpinner мутации влияют на известные особенности поведения пауков. В случае биоморфов мутации влияют на длину и угол роста ветвей растущих деревьев. Для глаз Нильссон и Пелгер первым делом постулировали наличие трех основных типов ткани в обычном глазе камерного типа. Камера имеет внешнюю, чаще всего светонепроницаемую, оболочку. Есть слой светочувствительных “фотоэлементов” (фоторецепторов). И есть нечто вроде прозрачного вещества, которое служит защитным окном или заполняет полость камеры-чаши – если, конечно, имеется чаша, поскольку мы в нашей модели ничего просто так на веру не принимаем. Стартовая позиция Нильссона и Пелгер – подножие горы – это плоский слой фоторецепторов (на рис. 5.14 показан серым цветом), который лежит на плоском экранирующем слое (черном), а сверху закрыт плоским слоем прозрачной ткани (светло-серый на рис. 5.14). По условиям их эксперимента, эффект от мутации выражается в малом относительном изменении одного из параметров – например, в небольшом уменьшении толщины или увеличении показателя преломления ограниченной зоны прозрачного слоя. Их интересовало, насколько высоко можно подняться по склону, если выйти из базового лагеря и упорно идти вверх? Подъем подразумевает планомерные, шаг за шагом, изменения, причем учитываются только те мутации, которые приводят к улучшению оптических свойств.
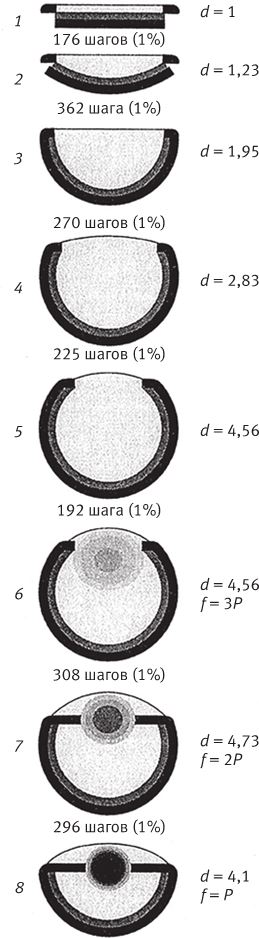
Рис. 5.14. Теоретическая последовательность стадий эволюции “рыбьего” глаза, предложенная Нильссоном и Пелгер. Число шагов от одной стадии до следующей рассчитано исходя из условия, что за один шаг какой‐нибудь один параметр меняется на 1 %. Как эти условные единицы соответствуют количеству поколений в эволюционном процессе, объясняется в тексте.
Итак, куда мы попадаем? Оттуда, где вовсе не было нормального глаза, мы потихоньку поднимаемся по тропинке с небольшим уклоном и приходим к знакомому нам рыбьему глазу с хрусталиком. В отличие от обычной искусственной линзы этот хрусталик неоднороден. У него переменный показатель преломления, примерно как на рис. 5.13. На диаграмме непрерывному изменению показателя преломления отвечают оттенки серого цвета. Показатель преломления стекловидной массы менялся от одной узкой зоны к другой, и так из нее постепенно выкристаллизовался хрусталик. Никаких фокусов. Нильссон и Пелгер не запрограммировали в модели стекловидной массы зачаточный хрусталик, чтобы дождаться рывка вперед. Они просто предоставили показателю преломления каждой узкой зоны прозрачного вещества меняться под влиянием генов. Показатель преломления каждой крупицы прозрачного вещества мог меняться произвольно в любую сторону. В стекловидной массе могло образоваться сколько угодно комбинаций зон с различными показателями преломления. Хрусталик обрел свою форму и свойства благодаря непрерывному движению вверх, которое послужило аналогом селекционной работы в каждом поколении с теми образчиками глаз, которые лучше всего видят.
Показать, что полноценный рыбий глаз мог постепенно развиться из плоского объекта, глазом не являющегося, было не единственной целью Нильссона и Пелгер. Их модель годилась и для оценки времени, которое понадобилось бы на образование глаза из ничего. При условии, что на каждой стадии какой‐либо параметр менялся на один процент, общее число стадий для их модели составило 1829. Изменение на один процент – это вовсе не чудо. Параметры могли бы меняться и на 0,005 процента на каждой стадии, и тогда таких стадий было бы 363992. Исследователям надо было выразить общее число изменений в объективных, реальных единицах – единицах генетических изменений. Для этого они сделали ряд допущений. Так, пришлось договориться об интенсивности селекции. Нильссон и Пелгер решили, что на каждого 101 выжившего животного с усовершенствованными глазами выживает 100 животных, чьи глаза не претерпели изменений к лучшему. Как видите, с позиции здравого смысла интенсивность отбора мала – можно отлично прожить и без всяких улучшений. Нильссон и Пелгер всячески старались добиться того, чтобы скорость эволюции по их оценке оказалась бы, мягко выражаясь, очень малой, поэтому они намеренно выбирали заниженные, “пессимистичные” цифры. Также они должны были обговорить еще два условия – касательно “наследуемости” и “коэффициента изменчивости”. Коэффициент изменчивости показывает, сколько изменений происходит в популяции. Естественному отбору нужны изменения, чтобы было что отбирать, и Нильссон и Пелгер здесь тоже намеренно задали неблагоприятные, низкие значения. Значение наследуемости говорит о том, сколько изменений из всех, произошедших в данной популяции, передается следующему поколению. Низкая наследуемость говорит о том, что изменения в популяции были в основном вызваны внешними условиями и что естественный отбор не оказывает заметного влияния на эволюцию, хотя и определяет, выжить особи или погибнуть. При высокой наследуемости выживаемость особей проявляется именно в выживаемости генов, поэтому влияние отбора на будущие поколения велико. Так как значение наследуемости нередко превышает 50 %, этот порог, который установили Нильссон и Пелгер, можно считать пессимистичной оценкой. И последнее неблагоприятное условие, которое они приняли, – что в одном поколении не могут одновременно меняться разные зоны глаза.
В каждом из этих случаев пессимистичность оценки подразумевает, что в результате продолжительность эволюции глаза окажется достаточно велика. Глубокий пессимизм гораздо более обоснован, нежели оптимизм. Недоверчивый скептик – например, Эмма Дарвин, – который сомневается в силе эволюции, скорее согласится с тем, что такой чрезвычайно сложный, многокомпонентный орган, как глаз, будет развиваться очень и очень долго, если вообще когда‐нибудь разовьется. По итоговой оценке Нильссона и Пелгер, этот период оказался поразительно коротким. В конце концов расчеты показали, что полноценный рыбий глаз с хрусталиком сформировался всего за каких‐то 364000 поколений. Если бы они выбрали более оптимистичные – то есть, наверное, менее реалистичные – параметры, все случилось бы еще быстрее.
Сколько лет можно отвести 364000 поколениям? Безусловно, это зависит от времени жизни поколения. Мы говорим о мелких морских животных – червях, моллюсках, небольших рыбах. Их поколение сменяется, как правило, за год или меньше. Таким образом, вывод Нильссона и Пелгер эквивалентен тому, что эволюция хрусталика могла бы завершиться менее чем за полмиллиона лет. По меркам геологии это очень-очень мало. Так мало, что в масштабе древних эр, о которых идет речь, это почти мгновение. Жаловаться, что для эволюции глаза было мало времени, – просто грех, это чудовищная, постыдная ложь.
Конечно, Нильссон и Пелгер учли не все особенности правильно сформированного глаза, которые, возможно, развивались гораздо дольше (хотя авторы исследования думают иначе). Скажем, предварительная эволюция светочувствительных клеток – ранее я называл их фоторецепторами, – по мнению Нильссона и Пелгер, закончилась еще до старта их модельной эволюционной системы. У современных глаз есть и другие полезные свойства, такие как способность менять фокусное расстояние и размер зрачка (“диафрагму объектива”) или подвижность в глазнице. Способность вращать глазами – это большой плюс не только по вполне очевидным причинам, но и потому, что жизненно важно уметь фиксировать взгляд при движении тела. Птицы удерживают голову в неподвижности – и тем самым фиксируют взгляд – с помощью мышц шеи даже при резких движениях всего туловища. В более сложных системах, обеспечивающих эту способность, задействованы тонкие механизмы мозга. Но ясно, что лучше иметь возможность хоть как‐то, на примитивном уровне, управлять движением глаз, чем не иметь ее вовсе, поэтому для нас не составит труда на пути следования к вершине горы Невероятности по пологому склону сложить фрагменты наследственных линий.
Если предмет расположен очень далеко от вас, для фокусировки исходящих от него лучей требуется линза с меньшей оптической силой, чем если бы он находился вблизи. Возможность в равной мере хорошо фокусировать лучи на дальних и ближних дистанциях – роскошь, без которой можно обойтись, но в природе ценен любой малейший шажок к увеличению шансов на выживание, и, действительно, самые разные животные могут разными способами менять фокусное расстояние хрусталика. Мы, млекопитающие, вытягиваем хрусталик, меняя его форму, с помощью мышц. Так же поступают птицы и большинство рептилий. Хамелеоны, змеи, рыбы и лягушки используют тот же принцип, по которому работает объектив камеры – слегка выдвигают хрусталик вперед или назад. Животные с маленькими глазками даже не пытаются этого делать. Их глаза подобны примитивному фотоаппарату “Кодак Брауни” – худо-бедно фокусируются на любое расстояние, пусть и неидеально. К сожалению, и наши глаза с возрастом начинают работать, как “Кодак Брауни”, а чтобы мы хорошо видели и вблизи, и вдали, нам выписывают бифокальные очки.
Нетрудно предположить, как постепенно развивался механизм регулировки фокусного расстояния. Во время опытов с полиэтиленовым пакетом и водой я сразу заметил, что могу увеличить или уменьшить резкость изображения, если сожму рукой пакет. Меня не интересовала форма пакета, я вообще не обращал на него внимания, а следил только за качеством проектируемой картинки, поэтому просто мял пакет по‐всякому, пока не добивался более четкого изображения. Любая прилегающая к какой‐либо зоне стекловидной массы мышца, предназначенная совсем для других целей, заодно могла бы скорректировать фокусное расстояние хрусталика. Это открывает широкие возможности для планомерных усовершенствований на всем пути к вершинам горы Невероятности, а высшим достижением мог бы стать способ фокусировки, которым пользуются млекопитающие или хамелеоны.
Может быть, несколько труднее, хотя и ненамного, изменить апертуру – диаметр отверстия, через которое поступают световые лучи. Нужда в этом возникает по той же причине, что и при фотосъемке. При фиксированной светочувствительности фотопленки/фоторецепторов света может поступать как слишком много (иногда он даже слепит), так и слишком мало. Более того, чем уже отверстие, тем больше глубина фокусировки – диапазон расстояний, в пределах которого изображение получается резким. Хорошая камера, как и хороший глаз, снабжена встроенным экспонометром, который автоматически затемняет линзу диафрагмой, когда солнце выходит, и открывает ее в противном случае. Зрачок человеческого глаза – пример самой умной автоматики, японские инженеры могли бы гордиться такой технологией.
Но и здесь нетрудно догадаться, как можно было бы запустить столь изощренный механизм уже на нижних уровнях горы Невероятности. Круглый зрачок, хотя мы к нему и привыкли, отнюдь не догма. Форма зрачка может быть любой. У овец и крупного рогатого скота зрачки удлиненные, ромбовидные, расположенные горизонтально. Такие же у осьминогов и некоторых змей, а у других видов змей зрачок представляет собой вертикальную щель. Кошачий зрачок меняет форму от круглой до щелевидной, ориентированной по вертикали (рис. 5.15):
Знает ли Миналуш,
Какое множество фаз,
И вспышек, и перемен
В ночных зрачках его глаз?
Миналуш крадется в траве,
Одинокой думой объят,
Возводя к неверной луне
Свой неверный взгляд.У. Б. Йейтс
Даже у многих дорогих камер зрачок объектива представляет собой не ровный круг, а многоугольник. Главное – регулировать световой поток, входящий в глаз. Если вы это поймете, то и первые этапы эволюции меняющегося зрачка покажутся вам очевидными. По низлежащим склонам горы Невероятности проходит множество удобных троп. Иридо-хрусталиковая диафрагма создает не больше препятствий на пути эволюции, чем анальный сфинктер. Важнее всего, наверное, ускорить реакцию зрачка. Раз нервная система уже есть, то и подстегнуть ее на подъеме в гору – тоже не проблема. Встаньте перед зеркалом и посветите фонариком себе в глаз – вы сможете на практике убедиться, что человеческий зрачок реагирует мгновенно. Можно провести и более эффектный эксперимент: направьте фонарик в один глаз, наблюдая за другим, и вы увидите, что оба зрачка реагируют одновременно.
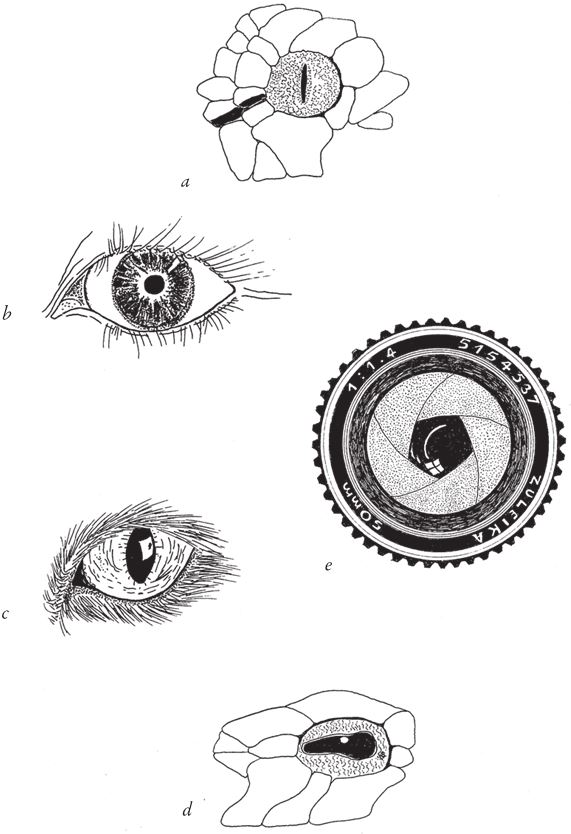
Рис. 5.15. Типы зрачков, в том числе зрачок фотообъектива. Форма зрачка не имеет критического значения, поэтому он может быть любым: (а) сетчатый питон; (b) человек; (с) кошка; (d) древесная змея длиннорылая плетевидка; (е) фотообъектив.
Как мы уже знаем, модель Нильссона и Пелгер воспроизводит хрусталик с переменным показателем преломления – у людей другой хрусталик, но у рыб, головоногих и прочих водных животных с глазами камерного типа именно такой. Линза формируется путем сгущения зоны с более высоким показателем преломления в прозрачном геле, который ранее имел однородную структуру.
Хрусталик не всегда формируется в области концентрации желеобразной массы. На рис. 5.16 показаны глаза двух насекомых, у которых хрусталики образовались иначе. В обоих случаях это так называемые простые глазки́ (оцеллии) – не путайте их с фасеточными глазами, к которым мы еще в свое время вернемся. Хрусталик первого – глаза личинки пилильщика – представляет собой утолщение роговицы, внешней прозрачной оболочки. Во втором случае (это глаз мухи подёнки) роговица не утолщается, а хрусталик образовался из скопления прозрачных бесцветных клеток. Оба пути развития хрусталика ведут к одному и тому же подъему на гору Невероятности – мы уже побывали там, когда изучали стекловидное тело глаза червя. По-видимому, хрусталики, как и сами глаза, эволюционировали независимо множество раз. У горы Невероятности далеко не один пик и много холмиков.
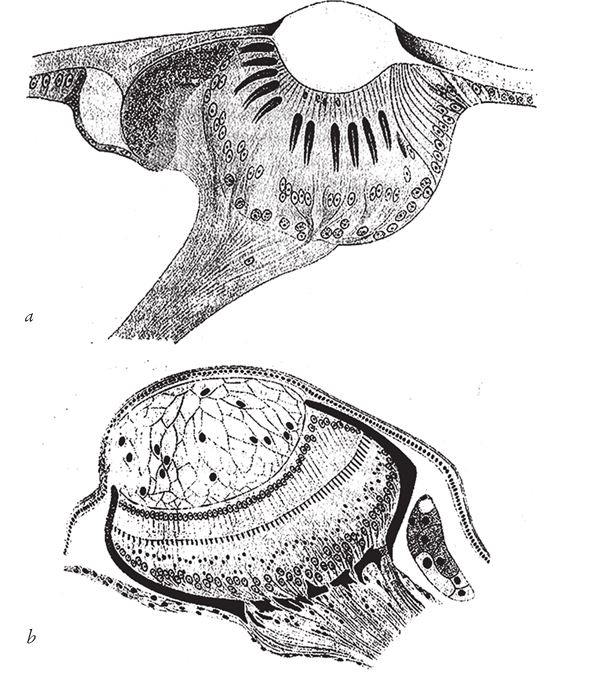
Рис. 5.16. Два варианта развития хрусталика у насекомых: (а) личинка пилильщика; (b) подёнка.
Многообразие форм сетчатки также говорит о многократной независимой эволюции этой структуры у животных. Во всех случаях, кроме одного, фоторецепторы, которые я до сих пор описывал, расположены перед нервами, соединяющими глаз с мозгом. Такая конструкция самая логичная, но не универсальная. У плоского червя (рис. 5.4а) фоторецепторы размещаются явно на “неправильной стороне” относительно нервов. Точно так же устроены и наши глаза – глаза позвоночных. Фоторецепторы ориентированы передом назад, против света. Это не так уж глупо, как кажется. Поскольку они очень маленькие и прозрачные, то неважно, куда они направлены – фотоны большей частью пролетают насквозь и попадают прямиком в ловушку мембранных экранов – складок, нагруженных пигментом. У хордовых “провода” (нервы), связывающие фоторецепторы с мозгом, уходят не к мозгу, как следовало бы ожидать, а к свету, и только в этом смысле можно говорить, что фоторецепторы направлены назад. Далее нервы проходят перед сетчаткой к ее определенной области – так называемому слепому пятну. Здесь они проникают через сетчатку к зрительному нерву – вот почему в этом месте сетчатка не воспринимает свет. Фактически мы слепы на эту зону глаза, что, впрочем, нам не мешает, ибо наш мозг ловко восполняет этот пробел. Слепое пятно можно заметить только тогда, когда в эту зону перемещается некий мелкий предмет, о существовании которого мы знаем по другим признакам – он как бы исчезает куда‐то, как свет, и явно замещается основным фоновым цветом.
Как я сказал, ориентация сетчатки не имеет никакого значения. Могло бы случиться и так, что при прочих равных нам полезнее было бы иметь сетчатку, расположенную наоборот. Пожалуй, это лишь подтверждает тот факт, что на горе Невероятности далеко не один пик и все они разделены глубокими ущельями. Коль скоро было положено начало эволюции полноценного глаза с его вывернутой наизнанку сетчаткой, возможен только один путь наверх – через усовершенствование строения того глаза, который уже есть. Радикальная смена концепции неизбежно сбросила бы нас вниз – не на пару шагов, а до самого дна каньона, что запрещено естественным отбором. Сетчатка позвоночных развивается по заданному пути уже у эмбриона, и, безусловно, этот путь развития и этот дизайн был свойственен еще древним предкам. Глаза многих беспозвоночных эволюционировали совсем по‐другому, и стало быть, их сетчатки ориентированы “правильно”.
То, что у хордовых сетчатка ориентирована задом наперед, конечно, интересно, но она и без этого взбирается на заоблачные пики горы Невероятности. Сетчатка человека насчитывает 166 миллионов фоторецепторов разных типов. Выделяют два основных вида: палочки (специализированы на восприятии света при низком уровне освещения, не различают цвета и не обеспечивают высокой остроты зрения) и колбочки (обеспечивают четкость изображения и восприятие цветов при ярком освещении). Эту книжку вы читаете с помощью одних только колбочек. Комету Галлея Джульет могла бы увидеть благодаря палочкам. Колбочки сосредоточены в небольшой зоне сетчатки – в центральной ямке (именно она позволяет вам читать), где палочек нет вовсе. По этой причине, чтобы разглядеть столь неясный объект, как комета Галлея, надо смотреть не прямо, а чуть в сторону, чтобы ее тусклый свет не попадал в центральную ямку. В теории восхождения на гору Невероятности количество фоточувствительных клеток и распределение их ролей не создают особых затруднений. Очевидно, развитие обоих свойств прокладывает легкие пути на вершину.
Для зрения лучше иметь большую сетчатку, чем маленькую. Она вмещает больше фоторецепторов, и можно разглядеть больше мелких деталей. Но за все надо платить. Вспомните сюрреалистическую улитку с рис. 5.1. И все‐таки некоторые мелкие создания могут позволить себе более широкую сетчатку. Майкл Лэнд, профессор Университета графства Суссекс, в послужном списке которого множество ярких и незаурядных открытий, касающихся глаз, и у которого я почерпнул массу сведений из этой области науки, нашел великолепный образчик – паука-скакуна. У пауков глаза не фасеточные, вот и пауки-скакуны используют глаз камерного типа в высшей степени рационально (рис. 5.17). Лэнд обнаружил весьма оригинальную сетчатку. Не широкий экран для проекции полномасштабной картинки, а длинную вертикальную полосу, узкую и не пригодную для восприятия качественного изображения. Однако паук с удивительной изобретательностью компенсирует минусы узкой сетчатки. Непрерывно перемещая сетчатку, он “сканирует” зону возможной проекции изображения. Таким образом, его “полезная” сетчатка намного больше реальной – похожий принцип используют пауки-боладоры, когда раскручивают свою единственную нить и увеличивают область захвата до площади обычной круговой паутины. Если сетчатка паука-скакуна зафиксирует достойный внимания объект – например, летящую муху или другого паука-скакуна, – “сканер” локализуется на прилегающем к цели пространстве. Это создает динамический эквивалент центральной ямки сетчатки. Находчивые пауки-скакуны оккупировали для своего глаза с хрусталиком невысокий, но вполне респектабельный пик в своем районе горы Невероятности.
Я показал вам великолепный инструмент для исправления пороков глаза-дырочки – хрусталик. Однако это не единственное средство. Эффект вогнутого и выпуклого зеркал основан на другом принципе, но они позволяют иначе решить ту же задачу – собрать потоки лучей, исходящих от каждой точки объекта, и свести их в одну точку изображения. Иногда такие зеркала работают эффективнее линз, недаром все самые большие телескопы – зеркальные (рис. 5.18а). Зеркальный телескоп имеет один небольшой недостаток: изображение формируется перед зеркалом, а именно – на пути входящих лучей. Как правило, в зеркальном телескопе есть маленькое зеркало, которое отражает сфокусированное изображение вбок – в объектив или фотокамеру. Это маленькое зеркало если и создает помехи, то несущественные, и не портит изображение. Оно лишь незначительно сокращает общий поток света, попадающий на расположенное сзади большое зеркало, а его собственного изображения не видно.

Рис. 5.17. Паук-скакун.

Рис. 5.18. Применение сферических зеркал для формирования изображения: (а) зеркальный телескоп; (b) Gigantocypris, крупное планктонное ракообразное, рисунок сэра Алистера Харди; (с) глазки гребешка в просвете створок раковины; (d) поперечный срез глаза гребешка; (е) овал Декарта.
Следовательно, теоретически вогнутое или выпуклое зеркало могло бы стать практичным физическим решением серьезной проблемы. Найдутся ли примеры “зеркальных” глаз в животном мире? Одним из первых об этом задумался сэр Алистер Харди, мой бывший профессор в Оксфорде; он прокомментировал свой рисунок интересного экземпляра глубоководных ракообразных – Gigantocypris (рис. 5.18b). В обсерваториях – например в Маунт-Вилсоновской и Паломарской – астрономы с помощью гигантских параболических зеркал улавливают даже самые слабые потоки фотонов, которые долетают от далеких звезд. Напрашивается мысль, что и Gigantocypris точно так же ловит фотоны, проникающие сквозь толщу воды в океане, но недавние исследования Майкла Лэнда решительно опровергают гипотезу о вероятном сходстве этого существа с телескопом. На сегодняшний день механизм его зрения неизвестен.
Зато другое животное для формирования изображения действительно пользуется самым что ни на есть вогнутым зеркалом, хотя и в комплекте с хрусталиком. Это открытие тоже принадлежит Майклу Лэнду, можно сказать, царю Мидасу в области исследования глаз животных. Это животное – двустворчатый моллюск морской гребешок.
На рис. 5.18с приведено увеличенное фото небольшого фрагмента щели между двумя створками раковины морского гребешка (шириной всего в 2–3 миллиметра), через которую видно тело моллюска. Между краем раковины и щупальцами расположены в ряд десятки крохотных глазок. В каждом глазу за сетчаткой есть вогнутое зеркало, которое участвует в формировании изображения. Именно эти зеркала придают глазкам морского гребешка голубоватое и зеленоватое жемчужное сияние. На рис. 5.18d показан глаз моллюска в разрезе. Как я уже говорил, помимо зеркала в нем есть и хрусталик – мы еще к нему вернемся. Сплошная серая зона между хрусталиком и вогнутым зеркалом – это сетчатка. К задней стороне хрусталика плотно прилегает часть сетчатки, видящая четкое изображение, которое проецируется зеркалом. Изображение получается перевернутое, а формируют его отраженные зеркалом лучи.
Так зачем же вообще нужен хрусталик? Сферическим зеркалам, в частности зеркалу в глазу морского гребешка, свойственна так называемая сферическая аберрация, из‐за чего изображение искажается особым образом. В конструкции одного из зеркальных телескопов – знаменитого телескопа Шмидта – этот недостаток устраняется благодаря точно подобранной комбинации линзы и зеркала. Морской гребешок, по‐видимому, справляется с проблемой по‐своему. По идее, сферическую аберрацию можно убрать с помощью специальной линзы, имеющей форму овала Декарта. На рис. 5.18e показан правильный овал Декарта. Давайте еще раз посмотрим на профиль реального хрусталика морского гребешка (рис. 5.18d). Профессор Лэнд подметил поразительное сходство форм и выдвинул идею, что в данном случае хрусталик нужен для коррекции сферической аберрации зеркала, главного элемента зрительного органа.
Исходя из нашего эмпирического опыта, мы можем предположить, откуда взялся глаз с вогнутым зеркалом на нижних уровнях отведенного ему на горе Невероятности склона. У многих животных есть отражающие слои, расположенные за сетчаткой, но они нужны не для формирования изображения, как у морского гребешка, а для других целей. Зайдите в лесную чащу, прихватив с собой мощный фонарь, и вы увидите множество устремленных на вас двойных лучей. У многих млекопитающих, особенно у тех, кто ведет ночной образ жизни (например, у западноафриканских золотистых потто, рис. 5.19b), позади сетчатки имеется отражающий слой – тапетум. Он предназначен для захвата со второй попытки фотонов, проскочивших ловушку из фоторецепторов; каждый фотон отражается обратно к тому самому фоторецептору, к которому он уже прилетал с другой стороны и который его не поймал, и поэтому изображение не искажается. Тапетум встречается и у беспозвоночных. Некоторые виды лесных пауков тоже обнаруживают себя в ярком свете фонарика. Глаза паука-волка (рис. 5.19а) горят, точно дорожные светоотражатели. Тапетумы, улавливающие все фотоны до единого, вполне могли развиться в глазах камерного типа у древних животных еще раньше, чем хрусталики. Не исключено, что тапетум – это та самая преадаптация, которая спровоцировала в отдельных случаях эволюцию глаза типа зеркального телескопа. А может быть, зеркало образовалось из другого источника. Наверняка тут ничего знать нельзя.
Получить хорошо сфокусированное изображение можно двумя способами – с линзами и с зеркалами. В обоих случаях картинка будет перевернута – верх и низ, а также правая и левая стороны поменяются местами. Правильное изображение формируется глазом совершенно другого типа – составным, или фасеточным; ему отдали предпочтение насекомые, ракообразные, отдельные виды червей и моллюсков, королевские (=камчатские) крабы (кое‐кто полагает, что эти удивительные морские животные ближе к паукам, чем к настоящим крабам) и обширная группа ныне вымерших трилобитов. На самом деле фасеточные глаза делятся на несколько разных типов. Я начну с самого простого – с аппозиционных фасеточных глаз. Чтобы понять, как функционирует аппозиционный глаз, мы должны вернуться почти к самому подножию горы Невероятности. Как нам уже известно, для того чтобы глаз видел картинку, а не просто передавал нам сигнал о наличии света, фоторецепторов должно быть много и они должны реагировать на световые лучи, приходящие по разным направлениям. Чтобы глаза видели не только то, что находится прямо перед ними, можно разместить фоторецепторы на поверхности чаши, экранированной сзади непрозрачным слоем. Все глаза, о которых шла речь до сих пор, развивались именно по этому принципу, в виде вогнутой камеры. Но, наверное, более очевидное решение – выстроить фоторецепторы на внешней стороне чаши, то есть на выпуклой поверхности, и таким образом направить их по разным осям. Это и есть простейший фасеточный глаз.

Рис. 5.19. Отраженные фотоны не пропадут зря. Сияющая отражательная оболочка, тапетум, паука-волка, Geolycosa sp. (а) и золотистого потто (b).
Вспомним о нашем первом опыте с изображением дельфина и о том, что из этого вышло. Тогда я сказал, что прежде всего мы должны справиться с множественностью изображений. Полчища “дельфинов” буквально заполонили всю сетчатку, так что в результате нельзя было разглядеть ни одного из них (рис. 5.20а). Глаз-отверстие был эффективен потому, что отсекались почти все лучи, а оставшийся узкий поток, который проникал во входное отверстие, формировал одно перевернутое изображение. Мы добавили линзу и усовершенствовали свою модель. В фасеточном глазу проблема решается еще более простым способом.
Фасеточный глаз представляет собой плотное скопление длинных прямых трубок, направленных во все стороны по радиусам от выпуклой поверхности. Каждая трубка, словно прицел винтовки, смотрит на один маленький участок пространства, который находится непосредственно на ее “линии огня”. Можно продолжить идею фильтрации – стенки трубки и поверхность купола не дают лучам других направлений попасть в нее и достигнуть фоторецепторов в ее задней части.
Таков базовый принцип работы аппозиционного фасеточного глаза. На самом деле эти маленькие трубчатые глазки – омматидии – устроены немного сложнее, чем простая трубка. Они снабжены собственными хрусталиками и крохотной сетчаткой, в которой содержится обычно около десяти фоторецепторов. Поскольку каждый омматидий формирует изображение на дне узкой трубки, оно получается перевернутое, то есть омматидий работает как несовершенный длинный глаз камерного типа. Но отдельно взятые изображения, сформированные одним омматидием, не воспринимаются мозгом. Омматидий передает только информацию об интенсивности светового потока, попадающего в трубку. А роль хрусталика сводится к тому, чтобы собрать как можно больше лучей на линии действия омматидия и сфокусировать их на сетчатке. Если объединить изображения, полученные всеми омматидиями, сформируется цельное правильное “изображение”, как показано на рис. 5.20b.
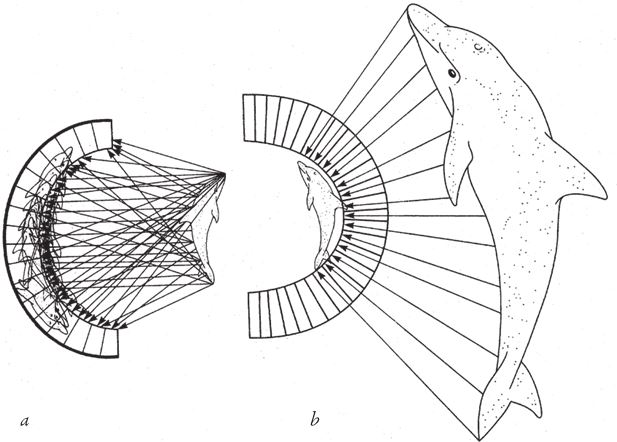
Рис. 5.20 (а). Повтор рис. 5.6; (b) вывернутая камера-чаша. Принцип устройства аппозиционного фасеточного глаза.
Как водится, вовсе не обязательно “изображение” будет правильным в нашем понимании – многоцветной картинкой, адекватно отражающей реальность. Мы только рассматриваем все варианты, которые позволяют визуально различать происходящие вокруг события. Некоторым насекомым фасеточные глаза нужны лишь для того, чтобы следить за движущимся объектом. Неподвижную картинку они могут и не увидеть. Видят ли животные так, как видят люди, – это вопрос философский, и, пожалуй, так сразу на него и не ответишь.
Например, основные свойства фасеточного глаза позволяют стрекозе выследить и догнать летящую муху, но чтобы разглядеть объект во всех подробностях, доступных нашему зрению, фасеточный глаз должен быть гораздо больше человеческого камерного. Этому можно дать примерно следующее объяснение. Очевидно, чем больше омматидиев, направленных в разные стороны, тем более детальным будет изображение. Стрекоза обладает примерно 30000 омматидиев, и этого вполне достаточно для охоты на лету (рис. 5.21). Но чтобы видеть так же хорошо, как видим мы, требуются миллионы омматидиев, а чтобы в глазу уместились миллионы омматидиев, они должны быть совсем крошечными. К сожалению, омматидий не может быть меньше определенного размера. Такие же ограничения, как мы видели, накладываются на размер глазного отверстия – это называется дифракционный предел. Из-за этого ограничения фасеточный глаз, если мы хотим добиться такого же качества изображения, какое дает наш камерный глаз, должен быть несообразно огромным – 24 м в диаметре! Немецкий ученый Куно Киршфельд нарисовал эффектную иллюстрацию – человека с фасеточными глазами и нормальной остротой зрения (рис. 5.22). Сотовая структура – это авторская интерпретация. В реальности каждая фасетка составного глаза скрывает в себе 10000 омматидиев. Киршфельд исходил из того, что люди хорошо видят только центральной зоной сетчатки, поэтому “человеческий” фасеточный глаз в его представлении вырос не до двадцати четырех метров, а лишь до одного. Двадцать четыре метра в диаметре или один – все равно с такими глазами ходить не будешь. Вывод: хотите четко видеть мир и ничего не упустить – выбирайте не составной фасеточный глаз, а простой камерный с одним, но качественным хрусталиком. Дэн Нильссон даже заметил про фасеточный глаз: “Мы не сильно погрешим против истины, если скажем, что эволюции пришлось выдержать отчаянную битву за исправление чудовищных, по сути, ошибок природы”.
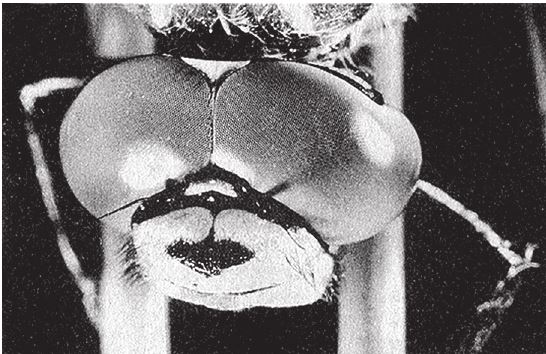
Рис. 5.21. Летающий хищник, стрекоза Aeshna cyanea, высматривает добычу огромными фасеточными глазами.
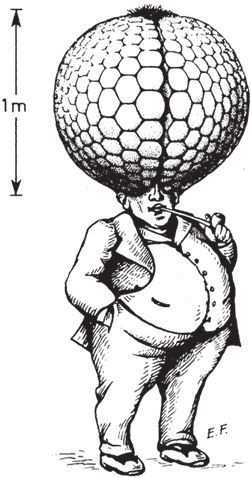
Рис. 5.22. Если бы человек имел фасеточные глаза при нормальной остроте зрения, он был бы таким, каким нарисовал его Куно Киршфельд.
Тогда почему же насекомые и ракообразные не отказались от фасеточного глаза и не развили вместо него камерный? Может статься, это был один из тех случаев, когда, заблудившись в долине у подножия горы Невероятности, вид не смог перебраться к другому склону. Нельзя спуститься в долину с тем, чтобы заново начать оттуда восхождение на более высокий пик, поэтому превращение фасеточного глаза в камерный было бы возможно только через несколько последовательных жизнеспособных промежуточных состояний. Но что мешает переходу от фасеточного глаза к камерному?
На ум приходит как минимум одно практически непреодолимое препятствие. В камерном глазу формируется перевернутое изображение. Фасеточный глаз видит правильную картинку. Найти промежуточное положение между ними, прямо скажем, непросто. Одно из вероятных состояний – полное отсутствие всякого изображения. Некоторым обитателям морских глубин, где царит почти кромешная тьма, достается так мало фотонов, что они оставили бесплодные попытки сформировать изображение. Все, что им остается – постараться хотя бы понять, есть свет или нет. Такие животные могли утратить нервный аппарат, необходимый для формирования изображения, а следовательно, оказаться в начале совсем другой тропы, ведущей на гору. Таким образом, они могли бы сыграть роль промежуточных форм на пути от фасеточного глаза к камерному.
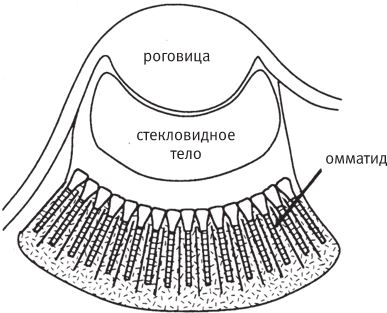
Рис. 5.23. Камерный глаз, эволюционировавший на основе фасеточного глаза. Уникальный глаз ракообразного Ampelisca.
У других глубоководных животных есть большие фасеточные глаза, но отсутствует хрусталик и оптический аппарат. Их омматидии лишились трубок, и выстроенные прямо на внешней поверхности фоторецепторы ловят все имеющиеся в наличии фотоны по всем направлениям. Отсюда всего один маленький шаг до оригинального глаза, показанного на рис. 5.23. Этот глаз принадлежит ракообразному Ampelisca, которое живет не то чтобы очень глубоко и, возможно, находится как раз на обратном пути от придонных предков. Его глаз устроен, как камерный – с одним хрусталиком, который формирует на сетчатке перевернутое изображение. Однако сетчатка с остаточными омматидиями явно заимствована у фасеточного глаза. Может, и небольшой шажок, но возможен он был только в том случае, если в переходном периоде полной слепоты мозг имел достаточно времени на эволюцию и успел “забыть”, как делается правильно ориентированное изображение.
Это пример эволюции фасеточного глаза до камерного – кстати, очередной аргумент в пользу независимого развития глаз у разных животных. Но откуда изначально мог бы взяться фасеточный глаз? Какие находки ждут нас на нижних уровнях этого обособленного пика горы Невероятности?
Было бы полезно еще раз оглядеть все современное царство животных. Помимо артропод (насекомых, ракообразных и иже с ними), фасеточными глазами обладают только некоторые полихеты (многощетинковые черви) и двустворчатые моллюски (тоже, по‐видимому, прошедшие свой собственный путь независимой эволюции). Среди многочисленных червей и моллюсков есть виды с примитивными глазами, которые могли бы составить ряд правдоподобных промежуточных вариантов у подножия тех склонов горы Невероятности, что ведут к пику фасеточного глаза, поэтому эти животные сослужат нам хорошую службу при изучении истории эволюции. Глаза на рис. 5.24 принадлежат двум разным видам многощетинковых червей. Отмечу еще раз – это не древние, а ныне существующие виды, и, возможно, их предки вовсе не имели глаз, которые можно было бы счесть промежуточными вариантами. Однако при их участии мы могли бы понять, как в ходе эволюции прогрессировали глаза от кучки разрозненных фоторецепторов слева до настоящего фасеточного глаза справа. Уверяю вас, что эта тропа не более крутая, чем дорога к обычному глазу камерного типа.
Эффективность омматидиев – в том виде, в каком мы до сих пор их рассматривали, – зависит от того, насколько они изолированы друг от друга. Лучи, исходящие от разных точек на теле дельфина, не должны попадать в омматидий, направленный, скажем, на кончик хвоста, иначе мы опять получим полчища дельфинов. Как правило, изоляцию омматидиям обеспечивает покрывающий трубку темный пигмент. Но тут возникают новые осложнения. Некоторые морские жители маскируются благодаря своей прозрачности. Они прямо‐таки сливаются с морской водой. То есть, их маскировка основана на том, что они пропускают фотоны сквозь себя. Однако смысл экранирования омматидия заключается именно в том, чтобы не пропускать фотоны. Как же избавиться от такого антагонистического противоречия?
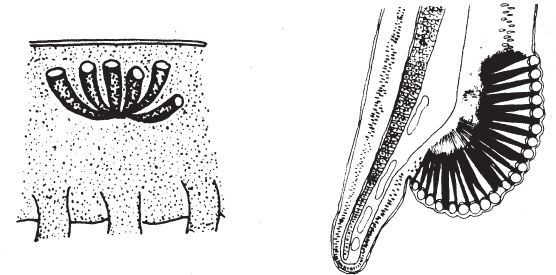
Рис. 5.24. Примитивные фасеточные глаза двух видов червей.
Среди глубоководных морских животных встречаются ракообразные, которые нашли весьма интересный компромисс (рис. 5.25). У них нет экранирующего пигмента, а их омматидии – не трубки как таковые. Скорее это прозрачные световоды, вроде наших оптико-волоконных систем. На переднем конце каждого световода есть утолщение – крошечный хрусталик с переменным показателем преломления, как у рыб. Световод вкупе с хрусталиком передает фоторецептору, расположенному в его основании, сильный поток света. Но только по прямой. Боковые лучи отражаются и в световод не попадают – и не нужен никакой пигментный экран.
Не во всех типах фасеточных глаз входящие световые потоки строго разделены. Это свойственно только глазам аппозиционного типа. Существуют по меньшей мере три разновидности так называемых суперпозиционных фасеточных глаз, в которых принимаются не такие решительные меры. Ни тебе улавливания лучей в трубки, ни оптико-волоконных технологий – свет проходит через хрусталик одного из омматидиев и попадает на фоторецептор соседнего омматидия. Хрусталики всех омматидиев дружно формируют единое изображение на общей сетчатке, образованной светочувствительными клетками всех омматидиев вместе взятых. Майкл Лэнд показал, как выглядит портрет Чарльза Дарвина, если рассматривать его сквозь составной хрусталик суперпозиционного фасеточного глаза светляка (рис. 5.26).
Рис. 5.25. Глаз глубоководного ракообразного со световодами.
В суперпозиционном фасеточном глазу, как и в аппозиционном (но не в камерном и не в изображенном на рис. 5.23 глазу ракообразного Ampelisca), получается правильно ориентированное изображение. Если предположить, что суперпозиционные глаза эволюционировали из аппозиционных предшественников, то все закономерно. Такое преобразование обосновано исторически, и по части мозга затруднений не должно было возникнуть. Но есть еще один немаловажный факт. Он связан с физическими особенностями формирования правильного изображения таким способом. В переднем отделе каждого омматидия аппозиционного фасеточного глаза есть хрусталик, который способен сформировать только перевернутое изображение. Следовательно, чтобы аппозиционный глаз превратился в суперпозиционный, необходимо каким‐то образом повернуть вышедшие из хрусталика лучи в нужную сторону. Мало того, надо аккуратно наложить друг на друга отдельные картинки, сформированные разными хрусталиками, так, чтобы получилась одна общая. Плюс этого метода в том, что итоговое изображение будет гораздо ярче. Однако повернуть лучи крайне сложно с точки зрения физики. Поразительно, что эволюция не только справилась с этой задачей, но и предложила сразу три решения – с помощью усовершенствованного хрусталика, усовершенствованных зеркал и замысловатой сети нейронов. Подробное описание всех сложнейших деталей нарушило бы логику и без того трудной главы, поэтому я буду краток.
Рис. 5.26. Майкл Лэнд сфотографировал портрет Чарльза Дарвина сквозь составной хрусталик суперпозиционного фасеточного глаза светляка.
Одна линза дает опрокинутое изображение. Вторая, установленная за первой на определенном расстоянии, по тому же закону опрокинет его обратно. Такая комбинация используется в телескопе-рефракторе Кеплера. Тот же результат можно получить и с одной линзой, показатель преломления в которой распределяется по заданной схеме. Как мы уже знаем, в отличие от искусственных, в “живых” линзах постепенная смена рефракции реализуется легко. Мухи-подёнки, сетчатокрылые насекомые, жуки, мотыльки, ручейники и представители пяти различных групп ракообразных достигают примерно такого же эффекта, который дает кеплеровская труба. Филогенетическая удаленность этих групп животных друг от друга указывает на то, что, по крайней мере, у значительной их части одно и то же свойство – механизм зрения, аналогичный принципу работы телескопа-рефрактора, – развилось независимо. Три группы ракообразных взяли на вооружение похожий метод – с зеркалами. В двух из них встречаются также виды, которые используют линзы. В самом деле, если посмотреть, какие типы фасеточных глаз развились у тех или иных видов, выяснятся удивительные вещи. Тут и там, прямо‐таки повсеместно, вы обнаружите совершенно разные способы решения одинаковых проблем, что свидетельствует о быстром, чуть ли не молниеносном эволюционном процессе.
У обширной и одной из главных групп двукрылых насекомых – мух – сформировались нейросуперпозиционные глаза. Таков же механизм зрения водных клопов гребляков (семейство Corixidae), и, скорее всего, мы вновь видим пример независимой эволюции. Нейросуперпозиционное зрение – чрезвычайно умная система. Термин “суперпозиционная” здесь даже не применим, поскольку омматидии изолированы друг от друга, как в аппозиционных глазах. Но благодаря оригинальной схеме соединения нервных клеток позади омматидиев возникает эффект наложения. Вот как это происходит. Давайте вспомним, что “сетчатка” одиночного омматидия состоит из полудюжины фоторецепторов. В обычных аппозиционных глазах зрительные нервы всех шести фоторецепторов просто складываются – учитываются все фотоны, которые достигают дна трубки, независимо от того, какой фоторецептор их зарегистрировал (поэтому я заключил в кавычки слово “сетчатка”). Несколько фоторецепторов нужны лишь затем, чтобы повысилась суммарная восприимчивость к свету. Поэтому не важно, что вообще‐то крошечное изображение на дне аппозиционного омматидия перевернуто.
Однако в глазу мухи продукты работы шести зрительных клеток не объединяются по принципу все со всеми. Каждая клетка работает совместно лишь с определенными клетками соседнего омматидия (рис. 5.27). На этом рисунке масштаб изменен для наглядности. Из тех же соображений стрелки показывают не преломленные хрусталиком лучи, а соответствие точек дельфина точкам на дне трубок. А теперь оцените гениальность этой схемы. Суть идеи в том, что клетки, которые смотрят на голову дельфина и находятся в соседних омматидиях, функционируют сообща; клетки, которые смотрят на хвост дельфина и находятся в соседних омматидиях, тоже функционируют сообща. И так далее. В итоге сигнал от каждого фрагмента тела дельфина формируется более интенсивным потоком фотонов по сравнению с обычным аппозиционным глазом и его простой системой омматидиев. Можно сказать, старая задача приумножения числа фотонов, которые излучают отдельные точки на теле дельфина, решается по‐новому – не оптическими методами, а тонким расчетом.
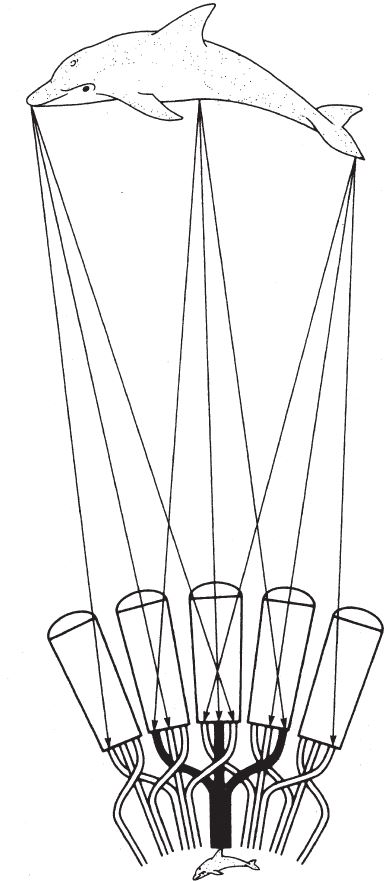
Рис. 5.27. Умная схема “подключения” омматидиев в суперпозиционном фасеточном глазу.
Понятно, почему такой глаз называется суперпозиционным, хотя в строгом смысле им не является. При истинном наложении с использованием сложных линз и зеркал проходящий через соседние фасетки свет суммируется, и все фотоны, которые излучают голова или хвост дельфина, оказываются в одних и тех же соответствующих этим участкам местах. При нейросуперпозиционном механизме фотоны по‐прежнему попадают в разные места, как и в аппозиционном глазу. Но благодаря точно рассчитанному переплетению нервных волокон, ведущих к мозгу, сигналы от них сходятся.
Вы не забыли, как Нильссон оценил темпы эволюции глаза камерного типа? По меркам геологии, это произошло едва ли не в один краткий миг! Найти окаменелости, по которым можно было бы проследить переходные стадии, – редкое везение. Для фасеточных или каких‐нибудь других типов глаз точных оценок сделано не было, но не думаю, что они развивались намного медленнее. Глаза слишком мягкие для того, чтобы окаменеть, поэтому у нас мало надежды встретить сохранившиеся с доисторических времен фрагменты глаз. Исключение составляют фасеточные глаза, поскольку значительная часть деталей их строения просматривается в аккуратном рисунке фасеток. На рис. 5.28 показан глаз трилобита девонского периода, жившего почти 400 миллионов лет назад. На вид он развит в той же степени, что и современный фасеточный глаз. Раз время, которое потребовалось для его формирования, в геологических масштабах ничтожно мало, то чего же еще ожидать?
Вся эта глава строится вокруг тезиса о легкой и быстрой эволюции глаз, образовавшихся буквально в мгновение ока. В самом начале я сослался на мнение специалистов, которые полагают, что глаза эволюционировали независимо в разных группах животных по меньшей мере сорок раз. На первый взгляд, подборка любопытных экспериментальных данных, опубликованная группой швейцарских коллег при участии профессора Вальтера Геринга, оспаривает этот тезис. Поясню в двух словах, что именно они обнаружили и почему их работа не противоречит выводам, которые мы делаем в этой главе. Но прежде я должен принести извинения за досадное и крайне неумное соглашение генетиков о названиях генов. Формирование глаз у плодовых мушек дрозофил регулирует так называемый ген eyless (ген безглазия)! Чудненько, не правда ли? Но в данном случае вопиющий терминологический ляпсус, сбивающий людей с толку, вызван довольно простой и даже интересной причиной. О действии гена мы узнаем по эффекту его мутации. Есть ген, в результате мутации которого рождаются мухи без глаз. Отсюда название места в хромосоме, занимаемого этим геном, – локус eyeless (безглазия) (locus на латыни означает “место”, в генетике это участок хромосомы, где расположены альтернативные формы данного гена). Но говоря о локусе eyeless, мы вообще‐то имеем в виду нормальный, неповрежденный ген, который занимает это место. Это и породило парадоксальное название гена, регулирующего развитие глаз. Можно было бы и динамик считать устройством беззвучия, ведь если отсоединить его от приемника, радио замолчит. Не могу с этим согласиться. Я бы переименовал ген eyeless в “ген eyemaker” (глазоделательный ген), но так мы тоже запутаемся. Слово “безглазие” мне решительно не по душе, я предлагаю использовать узнаваемую аббревиатуру названия этого гена: ey.
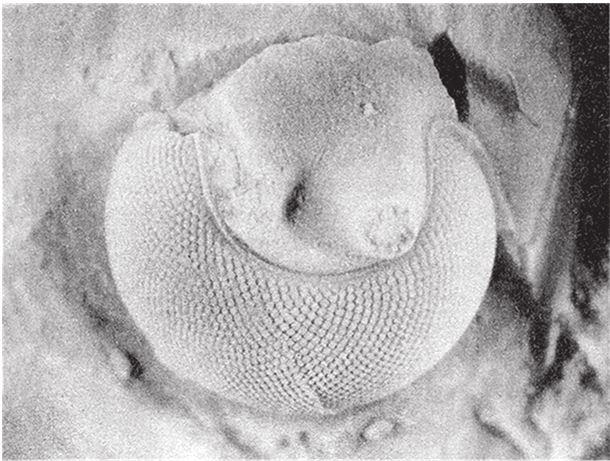
Рис. 5.28. Уже 400 миллионов лет назад фасеточные глаза были хорошо развиты: глаз трилобита девонского периода.
На сегодняшний день общеизвестно, что все клетки организма животного содержат полный набор генов, но в каждом отдельном органе и в каждой части тела находятся во “включенном” состоянии, или, другими словами, экспрессируются, лишь немногие из них. Именно поэтому печень и почки при одинаковом наборе генов ведут себя по‐разному. У взрослой дрозофилы ген ey, как правило, экспрессируется только в клетках головы, поэтому глаза там и развиваются. Георг Хайдер, Патрик Каллер и Вальтер Геринг придумали лабораторную методику активирования гена ey в совершенно не связанных с головой частях тела. Они сыграли с дрозофилой злую шутку – нашли способ запустить работу этого гена в ее антеннах, крыльях и конечностях. Результат был ошеломляющим – у подопытной особи полностью сформировались фасеточные глаза на антеннах, крыльях, ногах и в других местах (рис. 5.29). Эктопические глазки оказались меньше нормальных, но, как и нормальные фасеточные, имели множество правильно сформированных омматидиев. Они даже функционировали. Трудно сказать, чтó с их помощью видели мухи, но, судя по электрическим сигналам, поступавшим от нижней зоны омматидия, на свет они реагировали.
Это первый феноменальный результат. Второй поражает еще больше. У мышей есть ген, который называется small eye “маленький глаз”, а у человека этот ген называется aniridia – “ген аниридии”. Они тоже обязаны своими названиями генетикам – и негативному воздействию мутантных вариантов этих генов на строение организма. Их мутации приводят к недоразвитию и к полному отсутствию глаз или их структурных элементов. Сотрудники той же швейцарской лаборатории Ребекка Квиринг и Уве Вальдорф обнаружили, что последовательность ДНК в этих генах млекопитающих почти такая же, как в гене ey дрозофилы. Это означает, что их предки, столь же далекие от современных животных, как млекопитающие и насекомые, передали потомству этот ген. Мало того, в обеих филогенетических группах животных продукт этого гена, по‐видимому, принимает большое участие в развитии глаз. Третий феноменальный результат выглядит не менее эффектно. Геринг и его коллеги умудрились подсадить в клетки эмбриона дрозофилы мышиный ген. Невероятно, но факт – мышиный ген вызвал образование эктопических глаз у плодовой мушки. На рис. 5.29 (внизу) показан маленький фасеточный глаз, который вырос на ножке дрозофилы под влиянием мышиного эквивалента гена ey. Примечательно, что это не мышиный глаз, а присущий насекомому фасеточный. Продукт гена мыши просто запустил механизм формирования глаз в организме дрозофилы. Очень похожая последовательность ДНК была обнаружена и в генах моллюсков, морских червей немертин и асцидий. Ген ey, судя по всему, универсален для животных, и можно считать общей закономерностью то, что донорский ген из одного уголка царства животных способен стимулировать развитие глаза у реципиента, находящегося на совсем другом его краю.
Как эти фантастические эксперименты пересекаются с выводами, которые мы делаем в этой главе? Может, напрасно мы думаем, что могло быть сорок разных вариантов независимой эволюции глаза? Я считаю, что не напрасно. По крайней мере, не дезавуирована идея легкого и быстрого развития глаз. Наверное, эти опыты действительно показывают, что общие предки дрозофилы, мышей, человека, асцидий и прочих живых существ могли видеть. Их общий древний предок обладал каким‐то зрением, и его глаза, какими бы они ни были, развивались под влиянием последовательности ДНК, сходной с последовательностью гена ey. Но особенности глаза, характерные для конкретной группы животных – форма глаз различных типов, строение сетчатки, хрусталика и зеркал – эволюционировали быстро и независимо, так же как быстро и независимо происходил в разных группах животных выбор между простым и сложным глазом, а при наличии сложного сложного – между аппозиционным и двумя вариантами суперпозиционного. Об этом говорит разнообразие механизмов и систем, которые возникали в царстве животных тут и там без всяких закономерностей – можно сказать, по прихоти судьбы. В сущности, у животных чаще можно заметить нечто общее в глазах даже не близких, а более дальних родственников. То, что общие предки этих животных явно имели глаза того или иного типа, и то, что пути эмбрионального развитие этих глаз имели достаточно общего, чтобы регулироваться одинаковыми генами, никак не противоречит нашему выводу.
Рис. 5.29. Эктопические глаза дрозофилы, появившиеся в результате научного эксперимента; глаз на фото внизу образовался под влиянием мышиного гена.
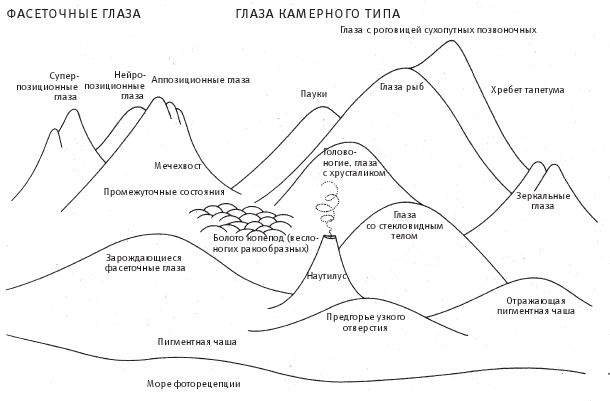
Рис. 5.30. Глазной район горы Невероятности: ландшафт эволюции глаз в интерпретации Майкла Лэнда.
Когда Майкл Лэнд согласился прочесть черновой вариант этой главы и высказал свои замечания, я попросил его нарисовать иллюстрацию к теме Глазного района горы Невероятности; на рис. 5.30 представлена его “карта”. Метафоры обычно применимы только к конкретным ситуациям, поэтому иногда приходится вносить кое‐какие поправки и даже вовсе вырезать метафору из своего рассказа. Читатель уже не раз имел случай заметить, что гора Невероятности представляет собой целую горную страну с множеством пиков, хотя мы говорим о ней как об одиночной вершине – как, например, о горе Юнгфрау.
Дэн Нильссон, еще один великий знаток зрения животных, тоже прочел черновик этой главы и обратил мое внимание на, пожалуй, самый поразительный пример специфической эволюции глаза, тем самым подытожив главную идею. Трижды, у трех различных групп рыб, абсолютно независимо развилось, скажем так, четырехглазие. Самый выдающийся представитель рыб с четырьмя глазами – батилихнопс (Bathylychnops exilis, рис. 5.31). Его обычные глаза смотрят вверх и в стороны, как у всех рыб. Но кроме них батилихнопс имеет вторую пару глаз, которые умещаются в границах основных глаз и смотрят исключительно вниз. Хотелось бы знать, что они там видят? Возможно, батилихнопс опасается, что хищник подберется снизу. Для нас самым интересным является следующее: в эмбриогенезе вторая пара глаз развивается совсем не так, как основные глаза, хотя можно предположить, что их развитие регулируется каким‐то вариантом гена ey. В частности, как написал мне доктор Нильссон, “этот вид заново изобрел для себя второй хрусталик, хотя один у него уже был. Хороший довод в пользу того, что хрусталику не так уж трудно было сформироваться”.
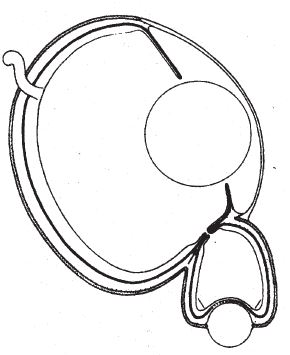
Рис. 5.31. Удивительный двойной глаз рыбы Bathylychnops exilis.
Все не так трудно, как нам порой кажется. Говоря о бессилии разума перед проблемой эволюции глаз, Дарвин преувеличивал сложности. И его жена преувеличивала, когда оставила на полях его работы свое скептическую ремарку. Дарвин знал, что делает. Креационисты обожают цитату, с которой я начал эту главу, но всегда немного недоговаривают. Дарвин пошел на уступку не для того, чтобы вызвать их на спор; далее он сказал:
Но когда в первый раз была высказана мысль, что солнце стоит, а земля вертится вокруг него, здравый человеческий смысл тоже объявил ее ложной; однако каждый философ знает, что старое изречение Vox populi – vox Dei (глас народа – глас Божий) не может пользоваться доверием в науке. Разум мне говорит: если можно показать существование многочисленных градаций от простого и несовершенного глаза к глазу сложному и совершенному, причем каждая ступень полезна для ее обладателя, а это не подлежит сомнению; если, далее, глаз когда‐либо варьировал и вариации наследовались, а это также несомненно; если, наконец, подобные вариации могли оказаться полезными животному при переменах в условиях его жизни – в таком случае затруднение, возникающее при мысли об образовании сложного и совершенного глаза путем естественного отбора, хотя и непреодолимое для нашего воображения, не может быть признано опровергающим всю теорию.
Назад: Глава 4 Взлет разрешен
Дальше: Глава 6 Музей Всех Возможных Раковин

