Книга: Восхождение на гору Невероятности
Назад: Глава 3 Сигнал с горы
Дальше: Глава 5 Сорокаполосный путь к просветлению
Глава 4
Взлет разрешен
Люди веками лелеяли несбыточную мечту подняться в небеса, а добившись все же своего, взлетают с такой натугой, что легко переоценить связанные с полетом трудности. Для большинства представителей животного мира полет – естественное состояние. Перефразирую изречение моего коллеги Роберта Мэя: в первом приближении летают все виды животных. Главным образом потому, как он сказал на самом деле, что в первом приближении все существующие виды относятся к насекомым. Но даже только про теплокровных позвоночных можно сказать, что летает больше половины всех видов: видов птиц вдвое больше, чем видов млекопитающих, а четверть всех видов млекопитающих составляют виды летучих мышей. Мы слишком крупные звери, потому‐то нам так тяжело летать. Бывают животные и покрупнее – слоны и носороги, например, – и естественно, мы их опасаемся, но в общем и целом все животные меньше нас (рис. 4.1).
Если ты очень маленький, сопротивление воздуха тебе не мешает. Для крошечного существа куда более непростая задача – устоять на земле. Такая принципиальная разница между большими и маленькими животными вытекает из некоторых непреложных физических законов.
При заданной форме тела его вес увеличивается не пропорционально приросту длины, а в третьей степени. Если страусиное яйцо втрое длиннее куриного при той же форме, оно весит не втрое больше, а в (3 х 3 х 3), то есть в 27 раз. Эта мысль давит на психику, пока ее не переваришь. Один человек съедает на завтрак одно куриное яйцо, а взвод полицейских – двадцать семь человек – поделит на всех яйцо страуса. Объем, а следовательно, и вес тела, увеличивается пропорционально кубу линейного размера. А вот площадь поверхности пропорциональна квадрату длины. Это легко показать на примере кубических коробок, но общее правило верно для любых форм.
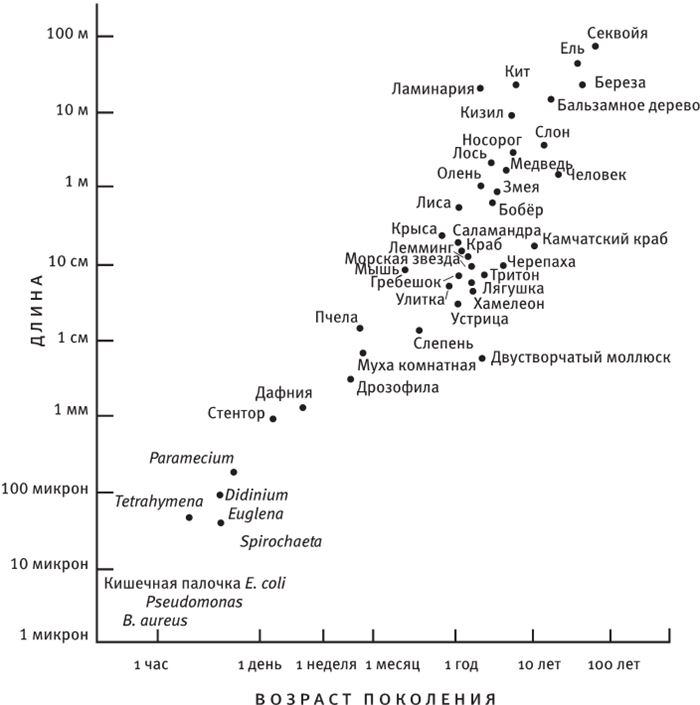
Рис. 4.1. Живые существа и растения могут отличаться по величине на восемь порядков. Для наглядности они показаны в координатах возраст поколения – размер (эти параметры взаимосвязаны, но сейчас мы не будем отвлекаться на причины корреляции). Шкала по обеим осям – логарифмическая, иначе для того, чтобы уместить на одном графике бактерию и секвойю, нам понадобился бы лист бумаги шириной более полутора километров.
Представьте себе коробку-кубик. Сколько в нее поместится таких же коробочек с ребром вдвое короче? Нарисовав кубики, вы легко получите ответ – восемь. В большую коробку можно уложить не в два, а в восемь раз больше яблок, чем в маленькую; упаковать не в два, а в восемь раз больше банок с краской. Но если вы захотите покрасить большую коробку, насколько больше вам понадобится краски по сравнению с маленькой коробкой? Опять же по рисунку вы увидите, что краски надо взять не вдвое и не в восемь раз, а в четыре раза больше.
Если сравнивать сильно отличающиеся по размеру предметы, площадь поверхности и объем разойдутся еще больше. Предположим, производитель спичек хочет изготовить для рекламы спичечный коробок величиной с человека – ящик, высота которого, если положить его на землю, составляет 2 м. Высота стандартного коробка – 2 см, поэтому по высоте ящика помещается 100 коробков. Вдоль одного края большого ящика укладывается подряд 100 спичечных коробков. По ширине тоже помещается 100 коробков. Итак, сколько коробков будет в заполненном большом ящике? 100 х 100 х 100, то есть миллион. Вроде бы ящик всего в сто раз больше обычного коробка, и по простоте душевной человек может так примерно и оценить его размеры: больше всего в сто раз. Но если смотреть иначе, ящик больше коробка в миллион раз, и в него можно уложить в миллион раз больше спичек – а на самом деле, еще больше, потому что картонные стенки съедают относительно немного объема.
Если договориться, что гигантский и обычный коробки сделаны из одного и того же материала, каковы будут относительные затраты на картон? Это зависит не от объема и не от длины сторон, а от площади поверхности. Для коробка-гиганта потребуется не в миллион раз больше материала, чем для обычного коробка, а только в 10000 раз. Относительно веса площадь поверхности обычного маленького коробка кажется намного больше, чем площадь поверхности огромного коробка относительно его веса. Если вы разрежете маленький коробок, сложенная картонка еле‐еле влезет в другой такой же коробок. Но если вы разрежете большую коробку, она едва прикроет дно другой большой коробки. Соотношение площади поверхности и объема – очень важный параметр. В то время как объем растет в третьей степени, поверхность увеличивается только во второй. Это можно выразить математически: если размеры тела увеличиваются пропорционально, то отношение площади поверхности к объему увеличивается в степени 2/3. Для мелких объектов отношение площади поверхности к их объему больше, чем для крупных. У маленького тела как бы “больше поверхности”, чем у большого такой же формы.
Далее, в жизни иногда важнее всего площадь поверхности, иногда объем, иногда габариты, а бывает, что и эти параметры вместе в различных комбинациях. Представьте себе гиппопотама, уменьшенного до размеров блохи с точным соблюдением всех пропорций. Рост (длина или толщина) настоящего гиппопотама могут быть в тысячу раз больше, чем у “блохопотама”. Следовательно, он в миллиард раз тяжелее. Площадь его поверхности больше всего лишь в миллион раз. Отсюда – для своего веса уменьшенный гиппопотам располагает поверхностью в 1000 раз большей, чем крупное животное. Простой здравый смысл подсказывает нам, что малютке было бы гораздо легче парить в воздухе, чем гиппопотаму нормального размера, но порой бывает полезно взглянуть, на что опирается здравый смысл.
Конечно же, ни одно крупное животное не может быть увеличенной копией маленького зверька, и сейчас мы поймем, почему. Механическое пропорциональное увеличение не прошло бы естественный отбор, потому что пришлось бы компенсировать возникший дисбаланс между площадью поверхности и объемом. У гиппопотама примерно в миллиард раз больше клеток, чем было бы у его крошечного двойника, а поверхностных клеток в его коже всего в миллион раз больше. Каждой клетке требуются кислород и питательные вещества, каждая клетка выделяет ненужные вещества, так что гиппопотам пропускал бы через себя в миллиард раз больше всех веществ в обе стороны. У блохопотама кожный покров занимал бы существенную долю поверхности, через которую происходил бы обмен кислорода и продуктов жизнедеятельности. У нормального гиппопотама, напротив, кожи мало относительно его веса, и для того чтобы обслужить весь свой миллиарднократный запас клеток, он вынужден сильно увеличить площадь поверхности. Задача решается за счет длинного, уложенного петлями кишечника, альвеолярной структуры легких и снабженных микроканальцами почек – причем все эти органы пронизаны густой сетью разветвляющихся кровеносных сосудов. Благодаря этому в организме крупного животного площадь внутренней поверхности значительно превышает наружную площадь его шкуры. Чем меньше животное, тем менее важны для него легкие, жабры, кишечник и кровеносные сосуды: его тело имеет достаточно большую площадь поверхности, чтобы самостоятельно справиться с обменными процессами, которые происходят в относительно небольшом количестве внутренних клеток. Можно сформулировать это короче: у маленького животного доля клеток, контактирующих с внешней средой, выше. У крупного животного, например гиппопотама, доля контактирующих с внешней средой клеток настолько невелика, что ему приходится наращивать ее за счет органов с развитой поверхностью – таких как легкие, почки и капилляры кровеносной системы.
От площади поверхности зависит скорость обмена веществ, а также другие важные свойства. К таковым относится способность подниматься в воздух и летать. Нашего блохопотама мог бы унести легчайший ветерок. Зверек взмыл бы к облакам вместе с восходящим потоком теплого воздуха, а потом, целый и невредимый, плавно опустился бы на землю. Если с такой же высоты упадет настоящий гиппопотам, он костей не соберет, а рухнув вниз с пропорционально увеличенной высоты, прокопает себе могилу. Живой гиппопотам может летать только во сне. Блохопотам при всем желании не смог бы не взлететь. Для того чтобы оторвать от земли обычного гиппопотама, надо привязать к нему такие большие крылья, что… впрочем, проект с самого начала обречен на провал, потому что даже гигантским крыльям не поднять колоссальную мышечную массу, которая должна будет приводить их в движение. Если уж вам придет в голову вывести летающее животное, не стоит начинать с гиппопотама.
Дело в том, что если крупный зверь захочет оторваться от земли, ему придется отрастить большие, широкие крылья, точно так же, как ему необходимо иметь почки и легкие с хорошо развитой поверхностью. А маленькому зверю, чтобы оторваться от земли, ничего отращивать не надо. Его тело и так имеет достаточно большую площадь поверхности. Высоко над землей по всему миру роятся тучи мелких насекомых и прочих созданий – так называемый воздушный планктон. Безусловно, у многих из них есть крылышки. Но к воздушному планктону относятся и различные бескрылые существа – они парят в воздухе, хотя и не имеют специальных органов, которые обеспечили бы им несущую поверхность. Просто они маленькие, вот и летают; как мы не тонем в воде, так и они не падают на землю. Можно пойти и дальше в этом сравнении: даже имея крылышки, крошечное насекомое энергично машет ими не столько для того, чтобы удержаться в воздухе, сколько чтобы “плыть” в нем. В данном случае слово “плыть” уместно, ведь с очень маленькими созданиями происходят и другие странные вещи. В таком масштабе огромную роль играет поверхностное натяжение, и маленькое насекомое воспринимает воздух как густую, вязкую среду. Взмахивая крылышками, насекомое, должно быть, ощущает примерно то же, что ощущали бы мы, плавая в сиропе.
Что толку болтаться в воздухе, не имея возможности менять высоту и поворачивать в нужную сторону, спросите вы. Не буду вдаваться в пространные объяснения, но с позиций генетики даже такой полет – уже большое достоинство, тем более для организмов, которые ведут прикрепленный (седентарный) образ жизни. Это особенно актуально для растений – любой участок земли время от времени, скажем, после пожара или наводнения, становится не пригодным для жизни. Растениям нужно много солнца, поэтому для них весь лес, кроме просветов, образовавшихся из‐за падения деревьев, не пригоден. Предки почти всех животных и растений обитали где‐то еще, и, скорее всего, у них есть гены, необходимые для того, чтобы попадать в новые места – неважно, куда. Поэтому семена одуванчика крепятся к невесомым пушинкам, а репейнику нужны крючки – чтобы цепляться за шерсть и мех животных. По той же причине множество насекомых дрейфует вместе с воздушным планктоном и опускается на неосвоенную территорию с каплями дождя.
Легкость, с которой мелкие твари дрейфуют по ветру, наводит нас на единственно возможную мысль: способность летать изначально эволюционировала у маленьких организмов – и пик Полетов на горе Невероятности кажется не таким уж недосягаемым. Очень маленькие существа могут летать вовсе без крыльев. Тех, что чуть покрупнее, поднимают в воздух крошечные крылышки – и вот мы уже направляемся к настоящим крыльям по ровному, отлогому склону горы Невероятности. На самом деле, как показало весьма оригинальное исследование Джоэла Кингсолвера и Мими Коэль из Калифорнийского университета Беркли, все не так примитивно. Кингсолвер и Коэль проверяли гипотезу об иной первоначальной функции крылышек у насекомых – они служили своеобразными солнечными элементами, необходимыми для обогрева. В те незапамятные времена крыльями не махали. Это были просто небольшие выросты на грудной клетке.
Авторы разработали хитроумную методику исследования. Сверяясь с палеонтологическими находками остатков первых насекомых, они изготовили простые деревянные модели. Одни без крыльев, другие с крылышками разной длины, причем у многих моделей крылышки были такие короткие, что их и крыльями‐то не назовешь, да и летать с такими трудновато. Модели насекомых разной величины испытывали в аэродинамической трубе, чтобы определить, насколько они эффективны с точки зрения аэродинамики. Кроме того, встроенные миниатюрные термометры показывали способность моделей улавливать свет яркого прожектора, имитирующий солнечные лучи.
Исследователи выяснили, что самые крохотные насекомые прекрасно летают и без крыльев – о чем мы и говорили. Моя приятная прогулка по склону горы Невероятности несколько осложняется тем выводом, что при очень маленьких размерах крылышки, похоже, не улучшают аэродинамических характеристик насекомого. Пока их длина не достигала сколько‐нибудь существенного значения, они не обеспечивали полезную подъемную силу. При двухсантиметровом туловище заметная подъемная сила возникала при длине крыльев, равной длине тельца. Крылышки, длина которых составляла всего 20 % длины тельца, не приносили модели ни малейшей пользы. На первый взгляд, у нас на пути к вершине возникает неожиданное препятствие: чтобы крылья одним махом выросли до необходимого размера, нужна мутация с очень сильным эффектом. Впрочем, его не так уж трудно обойти, если принять во внимание два дополнительных обстоятельства.
Во-первых, относительно большие крылья дают преимущества в полете только совсем крошечным насекомым. Насекомым покрупнее достаточную подъемную силу обеспечивают даже коротенькие крылья. Если постепенно наращивать длину крыльев при длине туловища 10 см, в какой‐то момент произойдет скачкообразное усиление аэродинамических свойств.
Что во‐вторых, мы поймем, вернувшись к самым мелким моделям насекомых. Для них доказано, что маленькие крылышки уже дают термический эффект. Крылышки чуть‐чуть большего размера лучше работают как солнечные батареи, но не создают подъемную силу. Судя по всему, для самых маленьких насекомых КПД “солнечных батарей” нарастает постепенно. Крылышки длиной 1 мм лучше, чем полное их отсутствие, 2 мм лучше, чем 1 мм – и так далее. Но так далее не до бесконечности. По достижении определенной длины эффективность “солнечных батарей” увеличивается уже гораздо медленнее. Следовательно, можно утверждать, что если преобладает аэродинамическая функция, само по себе постепенное повышение эффективности обогрева не стимулирует рост крыльев. Однако Кингсолвер и Коэль обнаружили хорошее решение этой проблемы. Как только у мелких насекомых развились крылышки ради аккумуляции солнечной энергии, некоторые виды по разным причинам стали увеличиваться в размерах. Тому есть несколько причин, развитие более крупных форм с течением времени – не редкость в животном мире. Может быть, насекомым выгоднее быть большими, потому что их с меньшей вероятностью съедят. Если в процессе эволюции они почему‐либо выросли, то вместе с ними должны были бы вырасти и их “солнечные батареи”, они же крылышки. В таком случае, увеличившись в размерах, насекомое со всеми своими крылышками и прочими органами автоматически перешло бы в ту категорию, для которой важнее аэродинамические преимущества, – и продолжило восхождение на гору Невероятности, пусть и к другому ее пику по другому склону.
Нельзя надеяться на точное воспроизведение в аэродинамической трубе тех процессов, которые происходили 400 миллионов лет назад в девонском периоде. Неизвестно, правда ли крылышки когда‐то служили насекомым лишь обогревательными элементами и до тех пор, пока все туловище по той или иной причине не увеличилось в размерах, были совершенно бесполезны в воздухе. Возможно, в естественных условиях шли иные физические процессы, отличные от смоделированных, и зачатки крыльев с самого начала развивались так, что становились более пригодными для летания. Однако эксперимент Кингсолвера и Коэль преподносит нам интереснейший урок. Он показывает нам новый тонкий маневр, учит лавировать и искать удобные подходы к вершине горы Невероятности.
У позвоночных, в большинстве своем относительно крупных животных, способность к полету, вероятно, вырабатывалась по другому сценарию. Функция управляемого полета эволюционировала независимо у птиц, летучих мышей (видимо, по меньшей мере в двух различных группах) и птерозавров. Многие животные, пусть они и не летали по‐настоящему, планировали с ветки на ветку, и это мог быть один из путей эволюции. В кронах деревьев кипит жизнь. Мы видим лес, который растет на земле. Пробираясь меж стволов, мы смотрим на лес с позиции больших, тяжеловесных и неуклюжих животных, устраивающих себе жилища на земле. Для нас дремучий лес подобен колоссальному сумрачному собору с арками и гулкими сводами и высоченным зеленым потолком. Но большинство его обитателей предпочитает верхние ярусы и видит его с другой стороны. Их лес – просторный, мягко колышущийся, залитый солнцем лиственный луг, который вообще‐то растет “на сваях”, хотя вряд ли их это интересует. На этом высотном лугу, никогда не спускаясь, проводит всю жизнь бесчисленное множество представителей животного мира. Луг – это место, где много зелени, а зелени много, потому что там много солнца, а солнечный свет – источник всей энергии на земле.
Рельеф поверхности не сплошной. Ажурный луг пестрит провалами, чуть зазеваешься – и долетишь до самой земли; тут не помешал бы мост. Многие виды животных хорошо экипированы, каждый по‐своему, и легко преодолевают довольно широкие пропасти. Попасть на противоположный край – зачастую вопрос жизни и смерти. Преимущества дают любые видоизменения в строении тела, которые помогают хоть немного, даже самую малость, увеличить дальность прыжка. Главное отличие белки от крысы – ее хвост. Хвост – не крыло, на нем далеко не улетишь. Но его “оперение” – пушистый мех – обеспечивает поверхность для создания подъемной силы. Крыса с беличьим хвостом, безусловно, могла бы перепрыгнуть через более широкую пропасть, чем крыса с обычным хвостом. И если у предков белки хвост напоминал крысиный, он должен был меняться долго и постепенно, становясь все более пушистым, пока не превратился бы в нормальный беличий хвост.
Говоря о беличьем хвосте, я употребил слово “оперение”, но это в гораздо большей мере относится к совершенно другому млекопитающему – карликовому летающему поссуму (рис. 4.2). У этого сумчатого животного больше общего с поссумом и кенгуру, чем с белками или крысами. Оно живет высоко на ветвях австралийских эвкалиптовых рощ. Его хвост, конечно, не настоящее перо с хитрой системой крошечных крючков и бородок – это определенно изобретение птиц. Но хвост карликового поссума очень похож на перо, и работает он точно так же.
Кроме того, карликовый летающий поссум имеет перепонки – складки кожи, натянутые между локтем и коленом, благодаря которым он способен “перелететь” почти на 20 м. Другое семейство австралийских поссумов (семейство Petiuridae) еще больше преуспело в развитии кожистой перепонки. Гигантский летающий поссум (Petauroides volans) преодолевает расстояния до 100 м и может поворачивать на 90 градусов, хотя и у него перепонка достает только до локтевого сгиба. Еще увереннее чувствует себя в воздухе большая сумчатая летяга (Petauroides australis). Ее перепонка расположена между лучезапястным и голеностопным суставами, как и у сахарного летающего поссума (карликовой сумчатой летяги Petaurus breviceps) и более крупного беличьего поссума (сумчатой летяги Petaurus norfolcensis).

Рис. 4.2. Карликовый летающий поссум, Acrobates pygmaeus, австралийское млекопитающее.
Гигантская летяга, обитающая в дальневосточных лесах, и североамериканская летяга с виду почти такие же, хотя принадлежат к абсолютно другой группе млекопитающих. Это самые настоящие белки, грызуны, но у них, как у более ярких представителей сумчатых летяг, кожистые перепонки натянуты между лучезапястными и голеностопными суставами. Летают они ничуть не хуже сумчатых “планеристов”. В Африке встречаются и другие грызуны, у которых тоже прекрасно развита техника планирования в воздухе. Это шипохвостая летяга (шипохвост) и карликовая летяга (летяга-пигмей) – они и не белки даже и прыгать на большие расстояния научились явно самостоятельно, американские белки-летяги тут ни при чем. На Филиппинах обитают экзотические зверьки шерстокрылы, они же кагуаны, или так называемые лемуры-летяги, с еще более мощной кожистой мембраной – от шеи до хвоста и от кончиков передних лап до кончиков задних. Кто они такие на самом деле, неясно, но точно не лемуры – настоящие лемуры живут на Мадагаскаре и, хотя некоторые их виды могут прыгать достаточно далеко, ни летать, ни просто планировать в прыжке они не способны. Как бы то ни было, шерстокрылы – не грызуны и не сумчатые, и свою кожистую перепонку, как и связанные с ее наличием повадки, они обрели независимо ни от кого.
Шерстокрылы, все виды белок-летяг и сумчатых летяг планируют, перелетая довольно далеко, примерно с равным успехом. Но если бы эволюция на этом не остановилась, у них могли бы развиться крылья нового типа, поскольку кожистая перепонка соединяет пальцы – хотя у других животных она достигает только кистей передних лап и голеностопа. То же самое, даже с еще большей очевидностью, справедливо для ящериц с красивым названием “летучие драконы” (рис. 4.3). Это древесные ящерицы, которые также обитают в кронах деревьев на Филиппинах и в Индонезии. В отличие от млекопитающих летяг, их кожистые перепонки никак не связаны с конечностями, а натянуты на длинных ребрах, и, когда надо, летучие драконы их расправляют. Из всех “летучих” животных мне больше всего нравится веслоногая лягушка – древесная лягушка родом из джунглей Юго-Восточной Азии. Вытянутые пальцы ее конечностей соединены перепонками, с помощью которых она, как и все остальные уже знакомые нам планирующие животные, “порхает” с ветки на ветку.
Ни в одном из приведенных примеров восхождение на гору Невероятности не сопряжено с серьезными трудностями. Более того, по пути к вершине летательные свойства не раз претерпевали изменения, поэтому удобных тропинок должно было быть много. Это подтверждает еще один представитель фауны тропических лесов Юго-Восточной Азии: украшенная древесная змея (золотистый древесный уж). Отталкиваясь от дерева, она запросто планирует в прыжке к другому дереву на расстояние около 20 м, хотя у нее нет ничего, подобного парусу, мембране или крылу с оперением. Все дело в том, что одна только геометрия тела обеспечивает относительно большую для ее веса площадь поверхности, а когда змея втягивает живот, так что внизу образуется вогнутая поверхность, эффект усиливается. По-видимому, первый решительный шаг к следующей ступени эволюции, к кому‐нибудь вроде летучего дракона уже с настоящей кожистой перепонкой, эта змея совершила. Второго шага так и не последовало – вероятно, потому, что при других условиях удлиненные ребра стали бы ей помехой.

Рис. 4.3. Эти позвоночные не умеют летать по-настоящему, они только планируют сверху вниз, с ветки на ветку; по часовой стрелке, начиная сверху справа: новозеландская летучая мышь колуго, Cynocephalus volans; летучий дракон, Draco volans; веслоногая лягушка, Rhacophorus nigropalmatus; карликовая сумчатая летяга, Petaurus breviceps; украшенная древесная змея, Chrysopelea paradisi.
Постепенную эволюцию таких существ, как белка-летяга, можно представить себе следующим образом. Некий предок, похожий на обычную белку, жил себе на дереве и прыгал с одной ветки на другую, ближайшую, без всяких специальных перепонок. Он отлично без них обходился, но если бы они были – хотя бы небольшие – или если бы его хвост был хоть немного пушистее, ему удалось бы перескочить на чуть более удаленную ветку, а следовательно, в критической ситуации спасти свою шкурку. Поэтому естественный отбор на стороне тех особей, у кого на лапках или суставах имеется лишняя кожа – и это становится нормой. В результате дальность прыжка среднего представителя популяции увеличивается на 10–15 см. Особи с более широким лоскутом лишней кожи прыгают еще дальше. Эта особенность закрепляется в следующем поколении – и процесс пошел. Для перепонки определенного размера существует предельное расстояние, и малейшее увеличение кожистой перепонки имеет жизненно важное значение. Средняя площадь складки кожи в популяции неуклонно растет, как растет и расстояние, на которое прыгает средняя особь. За много поколений развились животные вроде сумчатых летяг и белок-летяг, способные планировать на десятки метров и рулить в воздухе, приземляясь там, где хотят.
Однако планирование – это все еще не полет. Ни одно из этих животных не машет крыльями и не может оставаться в воздухе сколь угодно долго. Иногда они слегка меняют направление движения и немного поднимаются, прежде чем сесть на нижнюю ветку, но все равно планируют сверху вниз. Собственно управляемый полет, свойственный летучим мышам, птицам и птерозаврам, возможно, развился из планирующего спуска их предшественников. Большинство таких животных способно контролировать направление и скорость планирующего полета, а также выбирать конечную точку. Очень вероятно, что однообразная работа мышц, с помощью которых животное управляло своим движением в воздухе, постепенно перешла в полет с машущим крылом, и в течение эволюционного процесса средняя протяженность временного интервала между взлетом и приземлением все больше увеличивалась.
Впрочем, некоторые биологи склоняются к идее, что планирующие полеты на большие расстояния ведут эволюцию прыжков по деревьям в тупик. По их мнению, подлинный полет начинается не на дереве, а на земле. Безмоторный самолет, чтобы взлететь, должен стартовать с обрыва либо разогнаться по земле на буксире. Летучая рыба (рис. 4.4) использует второй способ – правда, разгоняется не по суше, а по морю, – и по дальности полета не уступает сумчатым воздухоплавателям, планирующим с ветвей деревьев. Летучая рыба набирает высокую скорость в воде, а затем взмывает вверх над волнами – надо думать, к вящему ужасу преследующего ее хищника, которому кажется, что она буквально исчезает в прозрачном воздухе. Она пролетает почти 100 м без контакта с водой. Иногда, нырнув, летучие рыбы вспенивают хвостом воду и несколько раз энергично работают плавниками, чтобы вновь набрать скорость и взлететь. Большие грудные – а у атлантических летучих рыб еще и брюшные – плавники служат им крыльями.
Не путайте истинных летучих рыб (Exocoetidae) с крылоперовыми, или долгоперовыми (Dactylopteridae), хотя по меньшей мере в двух книгах, что лежат на моем журнальном столе, их перепутали. Долгоперовые вовсе не летают – они ползают по морскому дну. Как полагают авторы многих научных работ, эти рыбы используют свои “крылья” в качестве стабилизаторов, отпугивают хищников их блеском и перепахивают ими песок в поисках жертв, а если кто‐то или что‐то нарушает их покой, то они всплывают на метр-два, расправляют “крылья” и снова плавно опускаются на дно. Единственное, чего долгоперовые не делают с помощью своих “крыльев” – так это не летают. Трудно сказать, откуда пошло заблуждение, будто они летают – может быть, из‐за явного сходства их грудных плавников с плавниками летучих рыб. Что касается самих летучих рыб, их предшественниками, очевидно, были не придонные рыбы, а быстроходные обитатели верхних слоев океана. Многие виды выпрыгивают из воды без помощи широких плавников. Очевидно, такие резвые прыгуны могли выиграть, отрастив плавники побольше и в следующих поколениях увеличив их площадь до размеров “крыльев”. Даже жаль, что до стадии развития летающих рыб не дошли одни из самых эффектных морских прыгунов – дельфины. Возможно, для того чтобы летать, они должны были бы быть меньше нынешних дельфинов, а кроме того, мелким дельфинам пришлось бы туго по другим причинам, связанным с термоизоляцией и свойствами подкожного жира. Существуют так называемые летучие кальмары – у них общие враги с летучими рыбами, например, тунец, и они спасаются тем же способом. Кальмары рода Onychoteuthis набирают в воде скорость более 70 км/ч и, пулей вылетая из воды почти на два метра, проносятся по воздуху метров на пятнадцать, а то и дальше. Столь высоких скоростей они достигают благодаря реактивной силе, а летят задом наперед, потому что все головоногие моллюски выбрасывают струи воды позади головы. Как только выталкивается очередная порция воды, запас движущей силы иссякает и остается на нуле, пока кальмар вновь не попадет в море. Тут летучие рыбы имеют преимущество: как мы уже знаем, они способны восстановить скорость, интенсивно работая хвостом в воде, пока их корпус большей частью еще в воздухе.

Рис. 4.4. Эти животные взлетают после разгона по поверхности. Атлантическая летучая рыба, Cypselurus heterurus (вверху) и летучий кальмар, Onychoteyhis.
В южноамериканских реках водятся рыбы еще одного интересного семейства – клинобрюхие (карнагиеллы, рыбы-топорики); судя по описаниям, они очень быстро, с шумом машут вверх-вниз грудными плавниками, так что им удается по‐настоящему, хоть и недолго, лететь в воздухе. Клинобрюхие не связаны близким родством ни с летучими рыбами, ни с долгоперовыми. Хотел бы я посмотреть, как мимо меня, бешено трепыхаясь, пролетает такой топорик. Не то чтобы я сомневаюсь в том, что повторяется во всех книгах, но любому рыбаку известно, да и мы уже это знаем на примере долгоперовых, что иногда рыбацким байкам надо доверять, проверяя.
В любом случае, разговор о планирующих летучих рыбах я завел ради прелюдии к теории о том, что за развитие подлинного полета – с машущим крылом – ответственны не обитатели верхнего лесного яруса, плавно спускавшиеся с ветки на ветку, а те животные, которые быстро бегали по земле и чьи передние конечности оставались при этом свободными. Летучие рыбы и кальмары живут в воде, но подтверждают тот факт, что животные, способные парить в воздухе, могли бы взлететь не только с края обрыва или с верхушки дерева, если бы хорошенько разогнались. И птицы могли бы, потому что они произошли от двуногих динозавров (по своей конституции птицы, можно сказать, и есть динозавры), среди которых, вероятно, были и отменные бегуны не хуже современных страусов. Можно подметить аналогию с летучей рыбой: ноги, подобно рыбьему хвосту, посылают животное вперед и сообщают ему высокую скорость, а передние конечности, видимо, первоначально помогавшие удержать равновесие и контролировать направление движения – как и плавники, – позже развились в крылья с аэродинамическим профилем. Некоторые млекопитающие – например кенгуру – набирают очень высокую скорость, отталкиваясь двумя ногами, а эволюция их передних конечностей могла повернуть в любую сторону. Кажется, из всех млекопитающих только наш вид ходит, поочередно переставляя ноги, на птичий манер, но мы довольно медлительны, и руки нам нужны не для того, чтобы летать, а чтобы носить в них и мастерить ими разные вещи. Все двуногие млекопитающие, способные бегать быстро, передвигаются, как кенгуру, отталкиваясь обеими ногами сразу. У четвероногих – собак, например, – такая манера вырабатывается естественным образом при горизонтальных прогибах позвоночника во время бега. Если проводить аналогии, киты и дельфины плавают, изгибая хребет вверх-вниз, как млекопитающие, а туловище рыбы и крокодила виляет из стороны в сторону, как у древних рыб. Кстати, нам следовало бы отдать должное невоспетым героям из числа звероподобных рептилий, первыми отработавшим тот самый стиль бега прыжками, которым теперь так восхищают нас признанные скороходы – гепарды и борзые. Наверное, и сейчас в вилянии собачьего хвоста, особенно когда пес подобострастно извивается всем телом, можно углядеть рудимент змеиных движений древней рыбы.
Среди бегающих по земле млекопитающих не только кенгуру и им подобные сумчатые передвигаются скачками. Мой коллега, доктор Стивен Кобб, в одной из своих лекций сказал студентам-зоологам из Университета Найроби, что валлаби водятся исключительно в Австралии и Новой Гвинее. “Сэр, я видел его в Кении”, – возразил кто‐то из студентов. Нет сомнений – ему повстречалось одно из существ с рисунка 4.5.
Этот зверек – долгоног; он не заяц и не кенгуру, а грызун. Как и кенгуру, удирая от хищников, долгоног скачет, чтобы набрать скорость. Точно так же ведут себя некоторые другие грызуны, например, тушканчики. Однако от следующего шага в эволюции по пути к способности летать двуногие млекопитающие явно воздержались. Среди млекопитающих по‐настоящему умеют летать только летучие мыши, причем у них кожистые перепонки охватывают как передние, так и задние конечности. Непонятно, как можно было перейти в развитии от быстрого бега к такому крылу, которое связывает летучую мышь “по рукам и ногам”. К птерозаврам это тоже относится. Мне кажется, что и у летучих мышей, и у птерозавров способность летать выработалась как следствие планирующего спуска с ветвей деревьев или с края обрыва. На каком‐то этапе кто‐нибудь из их предков должен был иметь что‐то общее с шерстокрылом.
Совсем другое дело – птицы. У них свое прошлое, главная роль в котором отводится удивительному приспособлению, а именно перу. Перья – это модифицированные чешуйки рептилий. Не исключено, что поначалу они несли иную функцию, актуальную и в наши дни, – сохраняли тепло тела. Так или иначе, они состоят из рогового вещества, пригодного для формирования легкой, ровной, гибкой и вместе с тем жесткой поверхности с аэродинамическим профилем. Птичье крыло разительно отличается от провисшей кожистой перепонки летучей мыши или птерозавра. Следовательно, у предков птицы должна была быть возможность вырастить подходящее для полета крыло, так чтобы не натягивать его между костей. Достаточно иметь передние конечности с костями. Все остальное обеспечивают перья, жесткие сами по себе. Ноги можно было оставить для бега. На земле птицы вовсе не такие неуклюжие и неповоротливые, как летучие мыши и птерозавры, – ноги позволяют им бегать, прыгать, сидеть на ветке, лазить по стволам и стенкам, охотиться и драться. Попугаи вообще работают ногами, как люди руками. Однако передние конечности предназначены для полета.

Рис. 4.5. Долгоног, Pedetes capensis.
Вот каким могло бы быть начало летной биографии у птиц. Их гипотетический предок – скажем, маленький шустрый динозавр – гонялся за насекомыми и ловил их в прыжке, отталкиваясь сильными задними лапами. Сами насекомые освоили воздушное пространство задолго до него. Они уворачивались от преследователя с удивительным проворством, и если бы динозавр мог на ходу корректировать направление прыжка, это увеличило бы его шансы на успех. В наши дни нечто в этом роде вы можете наблюдать в поведении кошек. Это нелегко, ибо в воздухе нет твердой поверхности, чтобы от нее оттолкнуться. Хорошо было бы изменить положение центра тяжести. Для этого можно переместить какие‐то одни части своего тела относительно других. Например, подвинуть голову или хвост, но самое очевидное – это подвинуть передние конечности. Теперь, когда они заняли удобное положение, их можно было бы использовать более эффективно, если бы их поверхность обеспечивала создание подъемной силы. Есть и другая гипотеза – перья, развивающиеся на первых крыльях, служили еще и ловчей сетью. Идея не такая уж далекая от реальности, как кажется – у некоторых видов летучих мышей крылья так и работают. Но согласно нашей теории, передние конечности должны служить прежде всего для управления полетом. Расчеты показывают, что наиболее эффективные для изменения крена и боковых наклонов в прыжке движения передних конечностей должны напоминать зачаточные движения машущих крыльев.
По сравнению с теорией планирующего спуска теория разбега, прыжка и промежуточной коррекции траектории переворачивает все с ног на голову. В первом случае прототип крыла поначалу должен был обеспечить плавный спуск с верхушки дерева. Лишь потом животные стали рулить с помощью крыльев и в конце концов научились ими махать. По теории прыжков за насекомыми, сначала встал вопрос управления полетом, и лишь потом сформировались передние конечности с пригодной для создания подъемной силы поверхностью. Это очень изящная теория, так как аэродинамическая поверхность должна была бы регулироваться на более поздних стадиях эволюции той же нервной цепочкой, которая отвечала за контроль над центром тяжести прыгающего древнего животного. Может быть, птицы начинали летать, подскакивая и отталкиваясь от земли, а летучие мыши – планируя вниз с ветки на ветку. А может, птицы тоже сначала только планировали, слетая с верхушек деревьев. Дискуссия пока не закрыта.
В любом случае с той давней поры современные птицы далеко улетели. Я бы сказал, разлетелись далеко в разные стороны, ибо они покорили великое множество пиков горы Невероятности. Сапсан, завидев добычу, несется вниз со скоростью выше 160 км/ч. Ястребы и колибри зависают на одном месте, сохраняя позицию с такой точностью, о какой вертолетчики могут только мечтать. Полярная крачка на ежегодные миграции из Арктики в Антарктику и обратно тратит по полгода, преодолевая расстояние почти в 20000 км. Там, где холодные волны будоражат ревущие сороковые, странствующий альбатрос, несомый огромными крыльями, размах которых превышает три метра, огибает полюс против часовой стрелки, неотрывно следуя за буйными ветрами и используя энергию этого естественного двигателя, а не машущего крыла. Фазан и павлин отрываются от земли – внезапно и резко – лишь иногда, если почуют близкую опасность. Другие – например, страусы, нанду и покинувшие этот мир новозеландские большие моа – слишком сильно выросли, чтобы летать, и по сравнению с мощными ногами, способными и быстро бегать, и лягнуть как следует, их крылышки оказались не развиты совсем. Полная противоположность – стрижи, которые почти всегда находятся в воздухе, с их хилыми, неуклюжими лапками и элегантными, вытянутыми кзади крыльями. Они даже спариваются и спят на лету, а садятся только на гнезда. Чтобы сесть на землю, стрижи должны выбрать высокое место, так как с ровной поверхности они не смогут взлететь. Стрижи вьют гнезда из того, что попадается им в воздухе и что они успевают сорвать с дерева, с пронзительным криком пролетая мимо. Cтрижу так же трудно и непривычно ходить по земле, как человеку – прыгать с парашютом или плавать под водой. Наш мир – это стабильный, недвижный фон для нашей деятельности. Но если смотреть черными глазами стрижа, то нормальная, естественная картина мира – это вечное мелькание бескрайних далей на крутых виражах. Для стрижа наша земная твердь сошла бы за головокружительный аттракцион.
Не все птицы машут крыльями, но, вероятно, птичьему парению и планированию предшествовал полет с машущим крылом. Последний довольно сложен и до конца не изучен. Напрашивается мысль, что подъемная сила создается непосредственно при сильных и ритмичных, направленных вниз ударах крыльев о воздух. Отчасти это верно, особенно в момент взлета, но главным образом подъемная сила, как и у самолета, обеспечивается формой крыльев – при достаточной скорости относительно воздуха. Подъемная сила возникает, когда ветер обдувает крылья, которые имеют специфический изгиб или наклонены под нужным углом, – либо, что то же самое, когда птица, неважно, по какой причине, движется вперед относительно воздуха. Взмахи крыльев в основном связаны с созданием необходимой положительной тяги (тяги для движения вперед). Суть в том, что крылья не просто так машут вверх-вниз – это залог их эффективности. Птица искусно производит вращательные движения от плеча при одновременной слаженной работе всех суставов и вдобавок получает дополнительный выигрыш просто за счет изгиба перьев. В результате всех этих вращений, сгибаний и согласованных действий энергия машущих крыльев трансформируется в положительную тягу примерно так же, как это происходит, когда кит бьет хвостом. Крылья создают подъемную силу благодаря поступательному движению в воздухе подобно крыльям самолета, хотя у самолета они неподвижны и потому устроены проще. Чем выше скорость, тем больше подъемная сила – вот почему “Боинг-747” держит высоту, несмотря на свой исполинский вес.
Крупным птицам махать крыльями намного тяжелее: против них работают законы физики. Если птица пропорционально увеличится в размерах, сохранив форму, ее вес вырастет в кубе относительно прироста длины тела, а площадь крыльев по отношению к увеличению их длины – лишь в квадрате. Чтобы не рухнуть вниз, крупная птица должна сама увеличиться непропорционально увеличению крыла и/или непропорционально увеличить скорость полета. Если допустить, что птица будет все расти и расти, рано или поздно наступит момент, когда, не имея в своем распоряжении реактивного или поршневого двигателя, она не сможет удержаться в воздухе, потому что ей попросту не хватит сил. В диапазоне птичьих размеров эта критическая точка находится где‐то чуть ниже крупных грифов и альбатросов. Как мы уже видели, некоторые крупные птицы опускают крылья, прекращая борьбу, и прекрасно живут себе на земле – и даже достигают еще больших размеров, как, например, страусы и эму. Но грифы, кондоры, орлы и альбатросы не отстранены от полетов. Почему?
Они умеют использовать внешние источники энергии. Если бы не жар солнечных лучей и не притяжение Луны, вызывающее приливы и отливы, в воздухе и на море царило бы вечное спокойствие. Внешняя энергия порождает океанские течения, гонит ветра, вздымает пылевые вихри, сотрясает атмосферу мощными толчками, способными сровнять здания с землей и изменить наезженные торговые пути; а кроме того, формирует восходящие тепловые потоки, и если воспользоваться ими с умом, можно подняться вместе с ними к облакам. Грифы, орлы и альбатросы владеют этой техникой в совершенстве. Наверное, только эти представители животного мира могут потягаться с нами в умении управлять энергией атмосферных явлений. Знания о парящих птицах я большей частью черпаю из публикаций доктора Колина Пенникука из Бристольского университета. Сам пилот-планерист, он хорошо понимает, как это делают птицы, а иногда и летает рядом с ними, чтобы изучить их повадки в естественных условиях.
Орлы и грифы извлекают пользу из восходящих воздушных потоков точно так же, как это делают пилоты на планерах. Восходящий воздушный поток представляет собой столб теплого воздуха, поднимающегося, по‐видимому, там, где почва получила влаги больше, чем солнечного тепла. Пилоты безмоторных самолетов, для которых это насущная необходимость, так навострились распознавать восходящие тепловые потоки, что замечают их издалека. На них указывают вроде бы незначительные приметы – характерная форма кучевых облаков над теплым столбом воздуха, особенности ландшафта у его основания. Чтобы совершить длительный авиаперелет на планере, можно прибегнуть к проверенному методу – подняться винтом к верхушке восходящего потока (километра на полтора), а затем парить по прямой вниз в нужном направлении. Спуск будет достаточно плавным и постепенным – гриф обычно теряет около метра высоты на каждые десять метров продвижения вперед. Прежде чем ему понадобится вновь набрать высоту на следующем тепловом потоке, он успеет пролететь больше 15 км.
Теплые восходящие потоки часто образуют своего рода “улицы”, и пилот, внимательно глядя на облака, отыскивает их загодя. Грифы тоже отлично умеют выбирать такие воздушные пути. Иногда, попав в “колею”, которая ведет в нужную сторону, гриф парит вдоль нее и поднимается вместе с очередным потоком, не утруждаясь даже покружить, чтобы встроиться в столб теплого воздуха. Это позволяет птице перемещаться на большие дистанции, не задерживаясь на кругах. Обычно грифы так ведут себя, только когда перебираются с места охоты на гнездовье. Большую часть времени грифы летят не по прямой куда‐то далеко, а нарезают круги в воздухе, высматривая падаль. При этом они не забывают поглядывать на своих сородичей. Если один увидит добычу и устремится вниз, остальные незамедлительно последуют за ним. По всему небу передается волна тревожного ожидания и готовности к действию, подобно тому, как по всей Англии вспыхивали костры на вершинах гор, распространяя сигнал о приближении Непобедимой армады.
Белые аисты во время своих долгих ежегодных миграций с севера Европы на юг Африки тоже не выпускают из поля зрения товарищей по стае, правда, с иной целью. Стая аистов насчитывает несколько сотен птиц. Как и грифы, они кружат, поднимаясь к верхним слоям восходящих воздушных потоков, а затем летят прямо к пункту назначения, пока не встретят еще один столб теплого воздуха. Поднимаются аисты толпой, но когда они выходят из потока, тесная стая перераспределяется в шеренгу. Передвигаться широким фронтом выгодно – пока птицы будут просто парить, направляясь вперед, с большей вероятностью кто‐нибудь найдет очередной восходящий поток. Заметив, что кто‐то рядом начинает подъем, все летят туда же. Таким образом, удача отдельных членов обширного коллектива, обнаруживших воздушный лифт, становится общим достоянием.
Что бы ни послужило началом эволюции птичьего полета – планирующий спуск с ветвей деревьев или разбег и прыжок, – почти не вызывает сомнений, что пальма первенства в парящем полете не принадлежит ни грифам, ни орлам, ни аистам или альбатросам. Все они унаследовали эту способность от своих менее крупных предков, которые махали крыльями. Специально для приверженцев теории о том, что полет стал следствием планирующего спуска с дерева, современные грифы, пусть даже они поднимаются на теплом восходящем потоке воздуха, а не слетают с верхушек деревьев, подтверждают идею возвращения к парению через промежуточную стадию – полет с машущим крылом. Согласно этой теории, на промежуточном этапе в нервной системе сформировалась новая цепь и выработались новые инстинкты управления полетом и маневрирования. Когда грифы вновь стали парить, сохранившиеся инстинкты работали уже лучше. Такое нередко бывает в природе – животные возвращаются к давно забытому поведению, успев пройти другую школу, и есть все основания утверждать, что школа жизни научила их лучше справляться с прежними задачами. Возможно, парящие птицы – не самый подходящий пример, поскольку неясно, как все‐таки птицы научились летать. Более показательно возвращение к прежнему стилю жизни тех животных, которые миллионы лет ходили по суше, а потом снова оказались в воде. Вот этим я и хочу завершить главу (рис. 4.6).
Пятьдесят миллионов лет назад предки китов и морских коров (дюгоней и ламантинов) были сухопутными млекопитающими; наверное, у китов предки были хищниками, а у морских коров – травоядными. В еще более давние времена их собственные предки, как и предки других сухопутных млекопитающих, обитали в море – были рыбами. Вернувшись в океан, киты и морские коровы попали в родную стихию. Мы точно знаем, что переход происходил постепенно. Вероятно, сначала они, как знакомые нам выдры, просто искали в воде пищу. Должно быть, они проводили на суше все меньше и меньше времени, и не исключено, что на определенном этапе киты и морские коровы напоминали знакомых нам морских котиков. Теперь они уже не вылезали из воды и на берегу были абсолютно беспомощны. Тем не менее они сохранили многие признаки своих сухопутных предшественников, а кроме того, как и все млекопитающие, – рудименты прежнего, совсем уж далекого морского прошлого. Киты вдыхают воздух, так как их вышедшие на сушу предки перестали использовать для дыхания жабры. Однако эмбрионы всех млекопитающих, включая китов и морских коров, имеют зачатки жабр – бесспорно, память о далеких водных предках. Пресноводные улитки, пожив на берегу, тоже вернулись в воду и теперь дышат воздухом. Еще раньше их предки жили в море, как большинство современных брюхоногих. Судя по всему, улитки перебрались из соленых вод в пресные по “земляному мостику” – наверное, что‐то, что они приобрели во время их сухопутной жизни, облегчило этот переход. Можно вспомнить и других обитателей суши, удалившихся обратно в воду, среди них черепахи, жуки-плавунцы, паук-серебрянка, а также вымершие ихтиозавры и плезиозавры. Черепахи могут извлекать из воды кислород, но не жабрами, а через слизистый эпителий в ротовой полости (у некоторых видов – в задней кишке); мягкотелые черепахи дышат через кожу, покрывающую панцирь. Жуки-плавунцы и пауки захватывают под воду воздушный пузырь. Все эти животные возвращаются в привычную для их древних предков водную среду, но под влиянием эволюционной истории, накопленной во время наземной жизни, приспосабливаются к этой среде совсем иначе.

Рис. 4.6. Киты и морские коровы вернулись в море после сотен миллионов лет сухопутной жизни; сверху вниз: дюгонь, Dugong dugon; ламантин, Trichechus senegalensis; горбатый кит, Megaptera novaeangliae; косатка, Orcinus orca.
Почему сухопутные животные, вернувшись в водную среду, не открывают заново прежний набор приспособлений к водной жизни? Почему киты и морские коровы не сменили легкие на жабры? Сейчас гора Невероятности преподнесет нам еще один ценный урок. В частности, важно понимать, откуда мы стартуем, как в анекдоте о том, как один человек на вопрос, как пройти в Дублин, ответил: “Э-э, я пошел бы не отсюда”. На горе Невероятности много пиков. Свою жизнь в воде можно устраивать по‐разному. Можно извлекать кислород из воды с помощью жабр, а можно высунуть голову наружу и глотнуть воздуха. Казалось бы, утомительно постоянно выныривать. Может, и так, но вспомните: предки китов и морских коров начинали недалеко от пика Вдыхания воздуха. Все их внутренние органы были адаптированы к вдыханию атмосферного воздуха. Наверное, они могли как‐то преобразовать их, оживить зародышевые рудименты древних жабр и стать ближе к рыбам. Но это должно было повлечь за собой тотальную перестройку всех систем организма. Представьте себе, что вы спускаетесь в глубокое ущелье, чтобы подняться оттуда на более высокий пик. Нелишне будет еще раз вспомнить, что дарвинизм не допускает временного ухудшения ради отсроченного выигрыша.
Даже если бы киты и морские коровы спустились в долину, пик Жабр, куда они в конце концов взобрались бы, вовсе не обязательно был бы выше. В морских глубинах жабры не всегда лучше легких. Безусловно, всегда удобнее иметь возможность дышать непрерывно, а не быть обреченным то и дело всплывать на поверхность. Однако мы смотрим со своей колокольни – мы сами вынуждены делать вдох каждые несколько секунд, и даже кратковременная нехватка воздуха ввергает нас в панику. Кашалоты, которые за миллионы поколений, проживших в море, прошли естественный отбор, могут оставаться под водой пятьдесят минут, прежде чем им понадобится новый глоток воздуха. Наверное, для кита всплыть, чтобы вдохнуть, – все равно что человеку выйти на минутку в туалет. Или поесть. Если рассматривать дыхание не как жизненно необходимый непрерывный процесс, а как что‐то вроде приема пищи, то станет ясно, что обитателю подводного мира гораздо лучше без жабр. Скажем, колибри едят более или менее постоянно. Для колибри, которые должны всасывать порцию нектара каждые несколько секунд, если только они не спят, перелетать с цветка на цветок так же важно, как дышать. Асцидии, похожие на мешочек морские хордовые, весьма дальние родственники позвоночных, неустанно прокачивают сквозь себя воду, отфильтровывая микроскопические частицы пищи. Эти фильтраторы едят явно без всякого удовольствия. Пожалуй, одна мысль о том, что им придется где‐то искать новую порцию, привела бы асцидий в ужас. Им было бы невдомек, зачем самые разные животные добывают себе пропитание с риском для жизни и далеко не всегда успешно, вместо того чтобы сидеть спокойно и буквально дышать пищей весь день напролет.
Даже если это было бы так, по одному только внешнему виду китов и морских коров можно судить об их сухопутной истории. Будь они изначально созданы для жизни в море, это были бы другие существа, и у них было бы гораздо больше общего с рыбами. Животные, чья история читается по их виду и повадкам, служат для нас самыми наглядными доказательствами того, что живые существа не были созданы для их нынешнего образа жизни, а эволюционировали от весьма не похожих на них предков.
История камбаловых рыб, прямо‐таки абсурдная, тоже написана у них на лбу. Только безумный дизайнер, решив создать плоскотелую рыбу и приступив к эскизу, изобразил бы на листе ватмана нелепую фигуру с такой головой, что оба глаза неизбежно должны были разместиться на одной ее стороне. Конечно, если бы он задумал “сконструировать” ската – рыбу, которая лежит на животе и глаза которой расположены симметрично на верхней части головы (рис. 4.7), – это была бы хорошая идея. Камбаловых перекосило из‐за собственного прошлого: их предки лежали на одном боку. Скатам досталась совсем другая история – когда их предки перешли к жизни на морском дне, они лежали не на боку, а на животе, поэтому скаты получились такие изящные и симметричные. Я вовсе не имею в виду, что у них не было никаких реальных причин для того, чтобы стать другими. Скаты ведут свой род от акул, а по сравнению с высокотелыми рыбами, по форме напоминающими лопасть весла, туловище акулы уже несколько сплющенное. Высокотелая рыба-“весло” на брюхе лежать не сможет – обязательно завалится набок. Угнездившись на дне, предки камбаловых взобрались на ближайшую горку в массиве Невероятности, хотя если бы им посчастливилось найти путь в маленькую долину, к подножию более высокой горы, до пика Симметричных скатов оставалось бы всего пару раз хвостом вильнуть. Повторяю: естественный отбор запрещает спускаться с горы Невероятности, и у этих рыб просто не было выбора – им пришлось за неимением лучшего перестроить органы зрения и сместить один глаз на противоположную сторону тела. Предки скатов тоже не замедлили занять ближайшую к ним высоту Плоских рыб – и обрели красивую симметричность. Когда я говорю “не было выбора” или “не замедлили занять высоту”, вы, конечно, понимаете, что речь, как обычно, не идет об отдельно взятых особях. Имеются в виду эволюционные линии, “выбор” означает возможность развиваться по альтернативным эволюционным траекториям.

Рис. 4.7. Две плоскости для рыб: скат, Raia batis (вверху), лежит на животе, а камбала, Bothus lunatus, на боку.
Я говорил, что нельзя спускаться с горы – но кто наложил этот запрет? Возможно ли такое, что его ни разу не нарушили? А кто “запрещает” реке течь не в ту сторону, куда направляет ее уже имеющееся русло? На все эти вопросы ответ примерно один и тот же. В реальности никто не давал реке команды оставаться в своих берегах, но, по вполне понятным причинам, река течет по своему руслу. Однако она может выйти из берегов и даже размыть их, и тогда все увидят, что курс реки изменился.
Что могло бы ненадолго повернуть вспять эволюционную линию и привести ее к точке, откуда открывается путь на ранее неприступную вершину горы Невероятности? Великий генетик Сьюалл Райт задался этим вопросом и, кстати, стал первым, кто применил к эволюции язык географии – можно сказать, заложил фундамент моей горы Невероятности. Райт представлял Америку в воинственно настроенной тройке запальчивых ученых, которые в 1920‐х и 1930‐х годах создали направление в науке, известное нам сейчас как неодарвинизм. Еще двое ее членов – англичане, не имеющие себе равных, но крайне несговорчивые гении – Р. Э. Фишер и Дж. Б. С. Холдейн, и справедливости ради надо добавить, что вся агрессия исходила в основном от них, а вовсе не от Райта. Райт понял, что естественный отбор, как ни странно, иногда противодействует стремлению к совершенству. Ровно по той самой причине, о которой мы только что говорили. Естественный отбор не пускает вниз. Если он с небольшого холма заарканил какой‐то вид у подножия более высокого пика, этому виду не вырваться и на высокий пик уже не попасть. Разве что естественный отбор ненадолго ослабит хватку и вид сумеет как‐нибудь незаметно слезть пониже, чтобы пересечь долину и подобраться к пологому склону более высокой горы. И если естественный отбор снова попытается его цапнуть, он будет уже в такой точке, откуда сможет быстренько эволюционировать по уходящим вверх тропам. Таким образом, с глобальной точки зрения, чередование периодов жесткого отбора с краткими передышками могло бы пойти на пользу эволюции. Возможно, релаксация такого рода действительно играет важную роль в реальном эволюционном процессе. Когда можно было бы рассчитывать на послабление? Например, если надо “заполнить вакуум”. В частности, когда популяция не использует все возможности своего ареала и поэтому начинает расти. Когда животные заселяют девственно чистый после природной катастрофы континент, перед ними открываются безграничные возможности, и естественный отбор берет паузу. Может быть, после того как вымерли динозавры, выжившие млекопитающие получили такую свободу действий, что некоторые эволюционные линии “потеряли контроль над собой” и на короткий промежуток времени спустились вниз, обнаружив затем более высокие пики горы Невероятности, куда в обычной жизни они не попали бы.
Еще один вариант – приток свежих вариантов генов из других источников. Во второй главе, в разговоре о пауках, я обещал к этому вернуться. В компьютерной модели паутин (NetSpinner) параллельно эволюционировали сразу три “дема”, а не одна размножающаяся половым путем популяция пауков-“ткачей”. Они как бы развивались независимо в трех различных географических зонах. Но – и это важно – не совсем независимо. Из одной локальной популяции в другую идет слабая миграция генов – случайное перемещение отдельных особей. Эта миграция генов сродни вливанию свежей “крови” в другую популяцию – как я говорил, “успешная подгруппа отдает свои гены той, что послабее, тем самым ‘подсказывая’ ей более выгодный способ создания ловчей сети”. Словно открывается незаконный путь на более высокий пик метафорической горы.
Мы приближаемся к излюбленному объекту внимания креационистов и главному камню преткновения для потенциальных сторонников эволюции, который непрочно держится на самой макушке наиболее труднодоступного утеса горы Невероятности, – к глазу.
Примечание: Когда эта книга уже ушла в печать, Дж. Марден и М. Дж. Крамер опубликовали интереснейшую работу о веснянках, где указывается еще одна дорожка на вершину Полетов с машущим крылом (Marden, J. H., & Kramer, M. G. (1995) Locomotor performance of insects with rudimentary wings [“Локомоторная активность насекомых со слаборазвитыми крыльями”]. Nature, 377, 332–4). Веснянок можно отнести к первичным насекомым в том смысле, что этот вид, хотя и ныне существующий, как полагают, имеет больше сходства с предками, чем другие современные насекомые. Вид, который изучали Марден и Крамер, Allocapnia vivipara, расправляет крылышки по ветру, точно паруса, и скользит по водной глади ручья. Скорость живого “парусника” примерно пропорциональна длине крыла. Даже с самыми крохотными крылышками насекомое движется быстрее, чем совсем без них. По величине они примерно такие же, как жаберные пластины древних ископаемых насекомых. Возможно, бескрылые предки обитали на водоемах и наращивали жаберные пластины, чтобы использовать их в качестве парусов. В таком случае развившиеся жаберные пластины стали работать более эффективно и, вероятно, открыли достаточно удобный путь на гору Невероятности. Что же до следующего шага к полетам с машущим крылом, с ним связано еще одно наблюдение, которое сделали Марден и Крамер. Насекомые другого вида, Taeniopteryx burksi, тоже скользят по воде, но машут при этом крыльями. Возможно, на пути к пику Полетов насекомые, как и Allocapnia, сначала прошли стадию движения под парусом, а потом – с машущими крыльями, как Taeniopteryx. Вполне вероятно, что в один прекрасный день порыв ветра подхватил невесомых мушек, машущих крылышками и роящихся над водой. Трепещущие крылышки позволяли им все дольше и дольше держаться в воздухе, и это могло бы стать следующим пролетом пандуса, который ведет к вершине.
Назад: Глава 3 Сигнал с горы
Дальше: Глава 5 Сорокаполосный путь к просветлению

