Книга: Восхождение на гору Невероятности
Назад: Глава 2 Шелковые путы
Дальше: Глава 4 Взлет разрешен
Глава 3
Сигнал с горы
Высоко над равниной вздымается гора Невероятности, вознося свои пики в головокружительную заоблачную бездну неба. Кажется, никто никогда не доберется до макушек горных башен. Точно крохотные букашки, ползут и карабкаются упрямые альпинисты, безнадежно глядя на далекие, недостижимые вершины. Огорченно качают маленькими головками и объявляют нависающую над ними махину неприступной.
Наши скалолазы чересчур амбициозны. Умопомрачительная картина вертикальных склонов захватила их, им даже не приходит в голову обогнуть гору и посмотреть, что там, с другой стороны. Они увидели бы не отвесные скалы с гулкими расселинами, а живописные луга, отлого восходящие к удаленным плато. Местами попадаются небольшие каменистые утесы, но их всегда можно обойти, так что хорошо экипированный альпинист в прочных ботинках, которому некуда спешить, легко преодолеет не бог весть какой сложный подъем. Если не пытаться покорить вершину одним марш-броском, сама по себе ее высота уже не имеет такого значения. Наметьте доступный маршрут, и если вы располагаете достаточным запасом времени, взойти на гору вам будет не труднее, чем сделать очередной шаг. Конечно, гора Невероятности – это аллегория. В этой и последующих главах мы узнаем, что она означает.
То, что вы прочтете ниже, взято из одного письма, опубликованного в лондонской “Таймс” несколько лет назад. Автор – имени я не назову, дабы не ставить его в неловкое положение, – весьма уважаемый коллегами физик, заслуженно избранный членом Королевского общества, самого авторитетного в Англии содружества ученых.
Сэр, я один из тех физиков… кого смущает дарвиновская теория эволюции. Мои сомнения вызваны не религиозными мотивами и не желанием лить воду на мельницу любой из сторон в споре – на мой взгляд, дарвинизм просто несостоятелен с научной точки зрения.…Нам ничего не остается, кроме как принять факт эволюции – его подтверждают все находки палеонтологов. Вопрос только в том, почему она происходит. Как утверждает Дарвин, все дело в случайности: в процессе смены одного поколения другим происходят случайные малозначительные, сохраняющиеся впоследствии изменения, в результате чего некоторые преимущества вида становятся более выраженными. Таким образом, живые существа постепенно вырабатывают более выгодные качества – например, силу, чтобы добывать себе пищу и бороться с врагами. Этот процесс Дарвин назвал естественным отбором.Как физик, я не могу с этим согласиться. Не думаю, что такая потрясающая машина, как организм человека, могла образоваться благодаря лишь случайным ошибкам. Взять хотя бы глаз. Дарвин признает свою слабость в этом вопросе – он не мог понять, как примитивный орган, реагирующий на свет, мог эволюционировать до глаза… Лично я не вижу альтернативы гипотезе о преднамеренном конструировании живой материи. Ни происхождение жизни, ни то, что за тысячи миллионов лет существования нашей планеты живые создания удивительным образом видоизменялись от поколения к поколению, нельзя объяснить в терминах традиционной науки.Но кто Конструктор?Искренне ваш…
Автор дважды сделал акцент на том, что он физик, и это придает его мнению особый вес. Другой ученый, профессор химии Университета штата из калифорнийского города Сан-Хосе, вдруг обратился к биологии, опубликовав статью под заголовком “Без Бога не было бы инжира”. Описывая достаточно сложные взаимодействия фигового дерева со своими опылителями – осами (см. главу 10), он делает следующее заключение: “Молодая оса всю зиму спит в плоде фиги, но в точно назначенный час вылетает, чтобы отложить яйца в плод летнего урожая, без чего опыление не произойдет. Для этого необходимо строго соблюдать режим – стало быть, Господь за этим следит!” Восклицательный знак мой. “Глупо думать, будто случайности эволюции могли привести к столь точному таймингу. Такие виды, как смоковница, без божественного вмешательства не смогли бы существовать… Сторонники теории эволюции утверждают, будто подобные вещи происходят спонтанно, без определенной цели и вовсе не по плану”.
Похожие соображения, но только касательно макромолекул, таких как ферменты, для которых проще, чем для глаза или инжира, просчитать их собственную “невероятность” – то есть вероятность того, что они появились по воле случая, – не раз высказывал сэр Фред Хойл, один из самых именитых британских физиков и, между прочим, автор научно-фантастического романа “Черное облако”, который входит в число лучших литературных произведений своего жанра. Скорее, ферменты служат чем‐то вроде огромного множества станков для массового производства молекул. Производительность ферментов определяется их трехмерной формой, форма – спиральной структурой, а спиральная структура – последовательностью аминокислот, соединенных в цепочку. Соблюдение этой последовательности непосредственно контролируется генами, что очень важно. Неужели все это случайно?
Нет, говорит Хойл, и он прав. Число аминокислот фиксировано – а именно, их двадцать. В типичной молекуле фермента несколько сотен цепочек, составленных из этих двадцати аминокислот. Как видно из простого расчета, вероятность случайного образования одной последовательности, скажем, ста аминокислот равна одному к (20 х 20 х 20… х 20), где 100 сомножителей, то есть 1 к 20100. Такое число трудно себе даже представить, оно намного превышает количество элементарных частиц во всей вселенной. Сэр Фред, желая быть честным по отношению к своим оппонентам-дарвинистам, делает реверанс в их сторону и широким жестом сокращает эту величину до 1:1020. Выглядит более приемлемо, но все равно вероятность удручающе мала. Профессор Чандра Уикрамасингхе, астрофизик, соавтор и коллега Хойла, цитируя его, сравнивает произвольное, “по воле случая”, образование способного работать фермента с ураганом, который пронесся над кладбищем старых автомобилей и случайно собрал из обломков “Боинг-747”. Однако Хойл и Уикрамасингхе упустили из виду, что дарвинизм – это не теория случайностей. Это теория случайных мутаций в сочетании с неслучайным кумулятивным эффектом естественного отбора. Почему, недоумеваю я, даже умудренным опытом ученым тяжело усвоить такую простую мысль?
Самому Дарвину приходилось воевать с физиками из предыдущего поколения, которые заявляли, что его теорию якобы разрушает одно слабое место – “случайность”. Одним из самых ярых научных противников Дарвина был Уильям Томсон, лорд Кельвин, пожалуй, величайший физик той эпохи. Ему принадлежит масса открытий – например, он рассчитал возраст Земли по скорости ее остывания, исходя из того, что когда‐то она была частью солнца и сформировалась из “выброса” его вещества. Согласно его выводу, возраст Земли составлял какие‐то десятки миллионов лет. По современным оценкам, наша планета живет уже тысячи миллионов лет. То, что лорд Кельвин ошибся в сто раз, нисколько не умаляет его заслуг. В те времена не был известен метод расчета по радиоактивному распаду, никто ничего не знал и о термоядерном синтезе, настоящем солнечном “огне”, – метод расчета по скорости остывания с самого начала был обречен на неудачу. Куда менее простительно то, что он высокомерно – “как физик” – не принимал во внимание биологически обоснованные аргументы Дарвина; якобы Земля молода, и дарвиновская эволюция не могла привести к тем результатам, которые мы наблюдаем, так как прошло слишком мало времени; очевидно, биологические факторы нельзя принимать в расчет, они меркнут на фоне более убедительных физических факторов. Но и Дарвин мог бы в свою очередь возразить – чего он не сделал, – что биологические признаки явно указывают на эволюционный процесс, следовательно, было достаточно времени для эволюции, и стало быть, физика ошибается!
Возвращаясь к вопросу о “случайности”, лорд Кельвин воспользовался своим привилегированным положением и не преминул процитировать в президентском послании Британской научной ассоциации другого известного физика, сэра Джона Гершеля, который назвал дарвинизм “законом чепухи”:
По сути, принцип произвольной, случайной изменчивости и естественного отбора, per se, не более подходит нам для удовлетворительного описания живой природы в прошлом и настоящем, нежели лапутянский метод сочинительства книг (доведенный до крайности) – для Шекспира и “Начал”.
Гершель подразумевал “Путешествия Гулливера”, где Свифт с издевкой описывал методику хаотического комбинирования слов, по которой сочиняли книги жители острова Лапута. Гершель и Кельвин, Хойл и Уикрамасингхе, анонимный физик, чье письмо я здесь привел, и всякие свидетели Иеговы ошибочно толкуют принцип естественного отбора, будто учение Дарвина и лапутянская писанина – одно и то же. Дарвинизм по сей день принято трактовать как теорию “случайностей”, причем даже в тех кругах, где могли бы и разобраться в сути предмета.
Со всей очевидностью, без тени сомнения можно утверждать, что если бы теория Дарвина и впрямь сводилась к простым случайностям, она не работала бы. Не надо быть физиком или математиком, чтобы понять: можно до второго пришествия ждать, пока нам повезет и сами собой как‐то образуются глаз или молекула гемоглобина. Астрономическая невероятность таких чудес живой природы, как глаза, ферменты, коленные и локтевые суставы, – узкое место не только дарвиновской концепции, а вообще любой теории о жизни, и именно дарвинизм предлагает ответ на этот вопрос. В теории Дарвина невероятные процессы разбиваются на небольшие, доступные восприятию отрезки, необходимое везение распределяется равномерно, предлагается обойти гору Невероятности и шаг за шагом подняться по пологому склону длиною в миллион лет. Одному Богу под силу совершить гигантский прыжок через пропасть. Соглашаясь с его ролью Всевышнего Конструктора, мы остаемся ровно там, откуда начали. Тот, кто способен создать поражающую воображение стройную, живую конструкцию, должен быть неправдоподобно мудрым и тонким дизайнером. Неправдоподобие – синоним ничтожно малой вероятности, что тоже нуждается в объяснении. Господь прост, как все гениальное, возражает богослов и аккуратно – впрочем, не вполне – увиливает от ответа, так как немудрящий бог при всех своих прочих достоинствах был бы чересчур примитивен для того, чтобы создать вселенную, не говоря уж о том, чтобы простить грехи, внять молитве, благословить брак, превратить воду в вино и вообще исполнить все, чего от него ждут. Это вещи несовместимые. Или ваш бог может сотворить мир и совершать прочие божественные чудеса – и в таком случае его самого не так просто понять, – или он на это не способен, и тогда не может дать нам разъяснения. Фред Хойл сказал бы, что Господь – это предельный случай “Боинга-747”.
Высота горы Невероятности символизирует сочетание идеала с бесконечно малой вероятностью, которое реализуется в строении глаза и в молекулах ферментов – а также и в способности Бога их сотворить. Если создать такие объекты, как глаз и молекула белка, почти невозможно, значит, для этого требуется точный расчет. Объект состоит из множества упорядоченных особым образом элементов. Число возможных комбинаций этих элементов чрезвычайно велико. Для белковой молекулы – прямо огромно. Айзек Азимов рассчитал количество вариантов для молекулы гемоглобина – гемоглобиновое число, как он его назвал. Оно оканчивается на 190 нулей. Столько раз можно поменять местами фрагменты гемоглобина так, чтобы собственно молекулы гемоглобина не получилось. Для глаза нельзя провести подобный расчет, не сделав ряд допущений, однако интуитивно мы понимаем, что получили бы еще одно колоссальное число. На самом деле та комбинация, которую мы наблюдаем, – лишь одна из триллионов допустимых, и в этом смысле она невероятна.
Далее, если вернуться назад, можно заметить еще один небезынтересный факт – вероятность любой перестановки элементов в равной степени близка к нулю. Даже сама свалка металлолома не более вероятна, чем самолет, поскольку существует бесчисленное множество вариантов комбинации ее элементов. Загвоздка в том, что все эти комбинации тоже дадут свалку. Тут надо вспомнить о качественных характеристиках. Как ни перекладывай куски металла на кладбище самолетов, летать они не смогут. Подходящих вариантов крайне мало. Среди всех возможных перестановок элементов глаза ничтожно мало таких, при которых глаз будет видеть. Глаз человека формирует изображение на сетчатке с поправкой на сферические и цветовые аберрации; сам сужает и расширяет зрачок, чтобы интенсивность светового потока внутри глаза оставалась более или менее постоянной, хотя извне может поступать свет разной интенсивности; сам меняет фокусное расстояние хрусталика в зависимости от удаленности предмета; различает цвета, сравнивая возбуждение светочувствительных клеток (колбочек) трех типов. Практически ни одна из вероятных перестановок элементов глаза не позволяет выполнять столь сложную и тонкую работу. У этой единственной существующей комбинации есть одна особенность. Вероятность реализации любого из вариантов одинаково низка. Но неработающих комбинаций неизмеримо больше, чем тех, от которых есть толк. Действующее устройство невероятно и нуждается в дополнительных пояснениях.
Великий математик и генетик, основатель современной статистики Р. Э. Фишер с присущей ему педантичностью указал в 1930 году (я лично с ним не был знаком, но так и слышишь, как он четко и размеренно диктует своей многострадальной жене):
Можно считать, что организм адаптировался к данным условиям и вообще к окружающей его среде лишь постольку, поскольку мы можем представить себе совокупность других, в чем‐то иных условий, к которым животное в целом адаптировалось хуже, и в той же мере поскольку мы можем представить себе незначительно отличающиеся от него живые природные объекты, которые хуже адаптировались к данной окружающей среде.
Глаза, уши, сердце, крылья летающих тварей, паутина – техническое совершенство этих органов и структур поражает нас независимо от того, где мы их видим, нам не надо наблюдать за ними в естественной для них среде, чтобы понять: они позволяют решать конкретные задачи, но если поменять местами какие‐либо их части, будет только хуже. На них впору повесить ярлык “невероятный идеал”. Инженер подумал бы, что если бы ему пришлось решать одну из этих задач, точно такое устройство он и сконструировал бы.
Есть и другое объяснение тому, что объекты такого рода не могли образоваться спонтанно. Как мы уже поняли, списать все на случайное стечение обстоятельств – то же самое, что одним махом запрыгнуть на вершину горы Невероятности с ее скалистой, неприступной стороны. Тогда что было бы равносильно постепенному подъему по другому склону – удобному, пологому, покрытому мягкой травкой и цветами? Медленное, кумулятивное, постепенное и неслучайное выживание случайных экземпляров – то, что Дарвин называл естественным отбором. Метафора горы Невероятности подчеркивает заблуждения скептиков, чьи высказывания приведены в начале этой главы. Их ошибка заключалась в том, что они смотрели вверх, задрав головы к устремленной ввысь вершине. Они полагали, что к ней – к глазу, белковым молекулам и прочим абсолютно невероятным сочетаниям элементов – можно добраться только по отвесному утесу. Дарвин показал плавный подъем с другой стороны горы – вот в чем суть его величайшего достижения.
Однако, может, и впрямь нет дыма без огня? Ложное толкование учения Дарвина как теории случайностей широко распространено. Ведь не просто так пошла молва? Ну что ж, ошибочные дискуссии и искаженные представления на чем‐то основаны, их поддерживает некий шаткий фундамент. Один из этапов дарвиновского процесса, а именно этап мутаций, действительно носит чисто случайный характер. Благодаря мутациям – как правило, спонтанным – появляются новые генетические варианты, доступные для отбора. Однако дарвинисты так настаивают на “спонтанности” мутаций только потому, что хотят противопоставить их другой части эволюционного процесса – отнюдь не случайному отбору. Для того чтобы происходил естественный отбор, мутации вовсе не должны быть случайными. Произвольно что‐то меняется или нет – отбор все равно происходит. Настаивая на непредсказуемости мутаций, мы делаем акцент на том, что естественный отбор, напротив, ни в коей мере не случаен. Забавно, что все дарвиновское учение воспринимается как теория случайностей как раз из‐за противопоставления мутаций и закономерности естественного отбора.
В сущности, и сами мутации во многих отношениях неслучайны, хотя эти их особенности не вносят вклада в невероятное совершенство организмов и поэтому мы не будем подробно о них говорить. Например, мутации, вызванные хорошо изученными физическими факторами, до известной степени закономерны. Операторы рентгеновских установок отходят в сторону, прежде чем нажать кнопку, или надевают просвинцованные фартуки именно потому, что рентгеновское излучение может спровоцировать мутацию. Некоторые гены подвержены изменчивости более других. В хромосомах есть “горячие точки”, где мутации в среднем происходят заметно чаще, чем в других звеньях цепи. Это пример закономерности иного рода. Бывают обратные (реверсивные) мутации. Как правило, прямая и обратная мутации гена равновероятны. Иногда ген в одном направлении мутирует быстрее, чем в противоположном. Возникает так называемое “мутационное давление” – тенденция к эволюции в определенном направлении независимо от направления естественного отбора. Впрочем, это скорее очередной пример неслучайных мутаций. Отметим, что мутационное давление далеко не всегда работает на улучшение признака. Как и рентгеновские лучи. Вовсе даже наоборот: в том, что касается качественных изменений, в большинстве своем мутации, какова бы ни была их причина, случайны, а это означает, что они неблагоприятны для организма, так как испортить что‐то можно разными способами, а путей улучшения немного.
Можно выдумать абстрактный мир, в котором мутации чаще ведут к улучшению. В такой гипотетической природе неслучайные мутации отличались бы по характеру от мутаций, спровоцированных рентгеновскими лучами – была бы заметна тенденция к опережению отбора и предвосхищению потребностей организма. Но вопреки разнообразным теоретическим мечтаниям, как раз для такого типа мутаций практически нет оснований, ибо маловероятно, что мутации систематически будут угадывать потребности организма, да и неясно, как вообще можно что‐то предвидеть. Что значит предвидеть потребности организма? Допустим, в регионе с умеренным климатом вот-вот начнется оледенение и тамошним оленям с их легкими шубками грозит вымирание. Большинство животных так или иначе погибнет, но если бы ровно в это время олени, как нельзя кстати, начали бы обрастать густым мехом, точно овцебыки, вид мог бы сохраниться. Теоретически можно придумать механизм запуска полезных мутаций в нужный момент. Как мы знаем, рентгеновское излучение повышает общий мутационный уровень, то есть повышается вероятность любых мутаций, приводящих к формированию как более теплого, так и более тонкого меха. Что если бы морозы могли как‐нибудь избирательно повлиять на направление мутаций – в сторону роста густой шерсти? И наоборот – что если бы жара могла вызвать обратную мутацию и шерстяной покров стал бы легче?
Приверженцы учения Дарвина вовсе не возражали бы против таких провидческих мутаций, если бы таковые происходили. Конечно, пришлось бы отказаться от претензий на исключительность, но устои дарвинизма не были бы подорваны: при попутном ветре вы быстрее долетите трансатлантическим рейсом до пункта назначения, но это не переубедит вас в том, что домой вас доставила все‐таки мощная машина. Однако если бы был открыт механизм подобных благоприятных мутаций, дарвинисты очень удивились бы – и были бы заинтригованы – по трем причинам.
Во-первых, несмотря на активные научные изыскания, такой механизм пока не открыт – по крайней мере для животных и растений (есть кое‐какие весьма спорные результаты для частных случаев бактерий). Во-вторых, на сегодня нет адекватного теоретического объяснения способности организма выбирать и запускать нужную мутацию. Наверное, можно представить себе, что если бы миллионы лет назад прокатилась волна десятков ледниковых периодов, то мог бы сформироваться некий “архетип”, до сих пор не известная разновидность естественного отбора на более высоком уровне, со склонностью к мутации в нужном направлении при первых признаках нового ледникового периода. Но, повторяю, нет ни доказательств, ни, более того, адекватных теоретических объяснений такого эффекта. В-третьих – вернемся к первому пункту, – по мнению некоторых дарвинистов (и я нахожусь в их числе), гипотетический механизм направленной мутации выглядит некрасиво и противоестественно. Конечно, нельзя отдавать приоритет эстетическим предпочтениям. Однако идея направленной мутации потому не вызывает у нас энтузиазма, что обычно ее предлагают те, кто уверен, что такая теория непременно должна быть – и эти люди ошибаются, так как не понимают, что даже при хаотических мутациях собственного потенциала естественного отбора более чем достаточно. Чтобы еще раз подчеркнуть состоятельность принципа естественного отбора, напомню, что теория допускает хаотичность мутаций. Но, как я уже говорил, теория не требует, чтобы мутации были случайными, и нет ни малейших оснований вешать на теорию ярлык бессистемности. Случайными могут быть мутации, но отбор – никогда.
Прежде чем расстаться с замерзающим оленем, давайте рассмотрим еще один вариант теории счастливых мутаций – вероятно, читая три последних абзаца, вы уже о нем подумали. В самом деле, неясно, откуда организму “знать”, что грядут холода и надо как‐то меняться, чтобы обрасти теплой шерстью, – или что скоро наступит жара и требуется обратная мутация. Проще представить себе, что если условия станут более суровыми, может произойти предопределенный рост частоты мутаций во всех направлениях. На интуитивном уровне это примерно понятно. Для организма каждый новый кризис, например ледниковый период или резкое потепление – сильный стресс. Лично для меня сильный стресс, вызванный какими‐либо непредвиденными обстоятельствами, морозом, жарой, засухой, указывает на то, что на текущий момент мой организм защищен не лучшим образом. Мне‐то, может, поздновато меняться, но если быстренько мутируют гены в моих половых органах – неважно, в какую сторону, – вероятно, кому‐нибудь из моих потомков это облегчит жизнь. Те из моих отпрысков-мутантов, кому достанутся гены с неблагоприятными, как выяснится, изменениями, погибнут при любых проявлениях экологического кризиса, будь то холод, жара, засуха или наводнение, и таковых, возможно, окажется большинство. Но в условиях достаточно серьезного кризиса их все равно ждала бы смерть. Может статься, что появление ветви уродцев и особей с небольшими отклонениями даст животным шанс на производство потомства, которое успешнее справится с очередным испытанием.
Гены, продукты которых контролируют мутации других генов, действительно существуют. Теоретически можно было бы утверждать, что ген-мутатор начинает работать под воздействием стресса, и для особой разновидности естественного отбора на более высоком уровне это было бы полезной тенденцией. Но увы, на поверку выходит, что эта теория не более обоснована, чем наша предыдущая идея направленной мутации. Начать с того, что нет фактов, доказывающих ее справедливость. Мало того – любые предположения о том, что повышение частоты мутаций благоприятно для естественного отбора, спорны в теоретическом отношении. Этот довод дает нам право утверждать, что гены-мутаторы всегда в конце концов исчезают в популяции, и это согласуется с нашей гипотезой о животных, которые оказались в стрессовой ситуации.
В двух словах, главный аргумент заключается в следующем. Если животному повезло дожить до возраста родителя, оно уже должно было обладать крепким здоровьем. Если бессистемно менять что‐то и так очень хорошее, можно и испортить объект своего внимания. В реальности большинство мутаций ведет к плачевному концу. На самом деле лишь немногие мутации могут хоть что‐то улучшить – и именно они делают возможным естественный отбор. Верно и то, что ген-мутатор, увеличивая частоту мутаций вообще, помогает своему носителю поймать редкую удачу – мутировать так, чтобы стать лучше. Если это происходит, надо учитывать, что тот самый вариант гена-мутатора, который находится в организме, улучшившемся благодаря вызванной им мутации, разделит успех этого организма и закрепится в популяции. Казалось бы, это дает толчок положительному отбору, выгодному для гена-мутатора, а стало быть, частота мутаций должна была бы вырасти. Увы, идем дальше.
У следующих поколений в процессе полового размножения гены индивидов будут перетасовываться, перестраиваться и рекомбинироваться. Рано или поздно наступит момент, когда ничто не будет препятствовать разделению “хорошего” варианта гена-мутатора и того “хорошего” варианта гена, который он породил, то есть одни индивиды родятся только с “хорошим” геном, другие – только с “хорошим” мутатором. “Хороший” ген с одобрения естественного отбора будет закрепляться в будущих популяциях. Но тот, кто его сотворил – невезучий ген-мутатор, – из‐за плясок полового размножения останется за бортом. Долгосрочный прогноз для гена-мутатора, как и для любого другого гена, зависит от его усредненных эффектов – среднего эффекта, который он произвел в течение длительного периода во всех организмах, где он оказался. “Хороший” вариант гена, появившийся благодаря гену-мутатору, в среднем проявляет себя хорошо, и несущие его организмы все чаще будет появляться в популяции. А ген-мутатор в среднем дает плохой эффект и, несмотря на эпизодические выигрыши, по усредненным результатам в естественном отборе неизбежно попадет в брак. Те организмы, где он сохранится, большей частью будут иметь отклонения в развитии или погибнут.
Наши доводы против возможности положительного отбора для гена-мутатора справедливы, если размножение происходит половым путем. Иначе выпадает стадия “перемешивания”. Гены-мутаторы в течение долгого времени благополучно переходят с одной ступени естественного отбора на следующую, так как в отсутствие половых контактов они не отделяются от случайно порожденных ими “хороших” вариантов генов и путешествуют из поколения в поколение вместе с последними. При размножении неполовым путем новая “полезная” мутация приведет к рождению нового клона сильной особи. Очередная “вредная” мутация быстро исчезнет и утащит за собой субклона-уродца. Если “полезная” мутация действительно принесет пользу, клон будет совершенствоваться и дальше, и все его гены сохранятся – как “хорошие”, так и “плохие”. “Плохие” гены выживут, потому что, несмотря на губительный эффект, в среднем гены клона будут качественными. А среди этих сохранившихся генов-“попутчиков” окажется и мутатор, в первую очередь ответственный за новые “полезные” мутации. Пока речь идет о “полезной” мутации, хотелось бы иметь возможность избавиться от мертвого груза испорченных генов, и для мутатора – источника их появления – исключения сделано не будет. “Полезная” мутация, будь на то ее воля, стремилась бы к здоровому, очищающему половому процессу воспроизводства. Если бы только мои организмы решили спариться, сказала бы она, я бы послала к чертовой бабушке всю эту ораву немощных захребетников. Меня ценили бы только за мои добродетели. Может, я и сама заметила бы, что какие‐то организмы хороши, а какие‐то не очень, но в среднем могла бы выиграть от собственного положительного воздействия. С другой стороны, поврежденные гены не имеют желания воспроизводиться половым путем – они и так неплохо устроились. В одиночку на открытых массовых соревнованиях, каковыми является половое размножение, они долго не продержатся.
Все это само по себе не объясняет приоритета полового способа размножения, хотя может послужить основой для следующих рассуждений. Тот аргумент, который я привел, что “хорошим” генам половое размножение идет на пользу, а “плохим” – во вред, никак не объясняет наличие полового размножения. Существует масса теорий, объясняющих появление в эволюции полов – и ни одной достаточно убедительной. К примеру, есть так называемый “храповик Мёллера” – более строгое описание той же гипотезы о “желаниях” генов, которую я упрощенно вам изложил. Гены-мутаторы из моей истории могут послужить развитию теории Мёллера. Бесполое размножение не только приводит к накоплению в популяции поврежденных генов. Оно стимулирует появление генов-мутаторов. Вероятно, бесполые клоны начнут вымирать быстрее – иными словами, чаще защелкает храповик Мёллера. Впрочем, секс, причины его появления, теория Мёллера – все это совсем другая и сильно запутанная история. Может статься, в один прекрасный день я наберусь духу и начну ее распутывать – и тогда напишу еще одну книжку: об эволюции секса.
Но мы отвлеклись. Главное, необходимо понять – при размножении половым путем мутации наказуемы, хотя в отдельных, очень редких, случаях естественный отбор может предпочесть мутантов. Это справедливо даже для стрессовых ситуаций, когда, казалось бы, рост частоты мутаций наиболее обоснован и оправдан. Склонность к мутациям – это всегда плохо, несмотря на то, что бывают и полезные изменения. Как ни странно, правильнее всего было бы считать, что естественный отбор – за нулевую частоту мутаций. К счастью для нас и для эволюции, которая должна продолжаться, генетическая нирвана недостижима. Естественный отбор – вторая ступень дарвиновского процесса – это сила, закономерно направленная в сторону усовершенствования признаков. Следовательно, любым улучшением мы прежде всего обязаны везению – вот почему дарвинизм имеет репутацию теории случайностей. Что есть глубокое заблуждение.
Уверенность в том, что мутации произвольны и для естественного отбора предпочтительна их нулевая частота, вовсе не мешает нам рассмотреть весьма любопытную возможность, так сказать, “эволюции эволюционной способности” – я писал об этом в одноименной статье. В 7 главе, посвященной калейдоскопической эмбриологии, я изложу вам свежую версию этой идеи. А пока давайте вернемся к естественному отбору, этой второй половине дарвиновской пары. Несмотря на то, что мутации могут быть спорадическими и, в одном очень важном смысле, они происходят спорадически почти всегда, сам естественный отбор по сути отнюдь не спорадичен. Из всех волков, которые могли бы выжить, выживут и передадут следующим поколениям свои гены не любые, произвольно взятые, а самые быстроногие, самые хитрые и коварные, самые чуткие и клыкастые. Следовательно, у нынешних волков мы наблюдаем гены, скопированные с генов неслучайно отобранных особей, живших много лет назад. Каждое поколение являет собой генный фильтр. Отфильтрованные за миллионы поколений варианты генов обладают качествами, необходимыми для того, чтобы пройти отбор. Они участвовали в производстве эмбрионов миллионов особей и ни разу не подвели. Каждая из этих миллионов особей благополучно дожила до зрелого возраста. Ни одна не оказалась настолько непривлекательной, чтобы не найти себе пару – под непривлекательностью мы понимаем любые качества, которые могут оттолкнуть потенциального партнера по спариванию. Все эти особи смогли дать жизнь хотя бы одному детенышу. Фильтр очень тонкий. Попавшие в будущее варианты генов – не случайные образцы, а самая что ни на есть элита. Они пережили периоды оледенения и зноя, “бэби-бумы” и спады рождаемости, справились с болезнями и хищниками. Вынесли не только капризы погоды – ливни, морозы и засухи. Они вынесли все капризы генов-компаньонов, ибо такова уж доля гена в половом размножении – менять партнеров в каждом поколении; выживают те гены, которые укрепили здоровье, “общаясь” с удачливыми партнерами из совокупного фонда, то есть с другими генами, в свою очередь выигравшими от взаимодействия еще с какими‐то генами в популяции. В формировании условий, к которым должен приспособиться ген, главную роль играют другие гены популяции – его товарищи по команде в реке, текущей из Эдема и несущей свои воды вниз по поколениям удачливых особей. Каждая отдельная популяция в том или ином рукаве реки создает микроклимат, в котором предстоит выживать отдельному набору генов.
Для упрощения мы считаем мутацию первой стадией эволюционного процесса, а естественный отбор – второй. Но это неверный подход, коль скоро предполагается, что естественный отбор зависает в ожидании мутации, а потом либо выбраковывает измененные гены, либо хватается за них – и снова приостанавливается. Вероятно, естественный отбор такого рода мог бы работать – а может, где‐то во вселенной и работает. Но на нашей планете ничего подобного не происходит. В реальности есть обширный пул изменений, который накопился благодаря медленному притоку мутаций и в котором благодаря половому размножению произошло интенсивное перемешивание и приумножение изменений. Причина изменчивости – мутация, но к тому моменту, как ею “заинтересуется” естественный отбор, ей может быть уже много лет.
Приведу пример: мой оксфордский коллега, покойный Бернард Кеттлуэлл, прославился своим исследованием эволюции темных, почти черных, а некогда светлых бабочек одного вида. Темноокрашенные особи данного вида – березовой пяденицы Biston betularia – как правило, чуть устойчивее к воздействию неблагоприятных факторов, чем светлые, но в сельских районах, где нет промышленного загрязнения, встречаются реже, так как они лучше видны птицам, то есть их съедают первыми. В промышленных зонах, где стволы деревьев черны от копоти, они не так заметны, как их светлые сородичи, поэтому реже попадаются птицам на обед. Это позволяет им более эффективно использовать свое природное преимущество – устойчивость. Дальнейшее стало одним из лучших доказательств эффективности естественного отбора: начиная где‐то с середины XIX века численность темноокрашенных особей в промышленных районах стала стремительно увеличиваться, и в конце концов эта разновидность возобладала. Сейчас вы поймете, почему этот пример показателен. Многие ошибочно полагают, что после промышленной революции естественный отбор “культивировал” одну-единственную новую мутацию. Мы же, напротив, уверены, что бабочки с темной окраской появлялись всегда, но не выживали. Эта мутация, как и большинство других, периодически воспроизводилась, но птицы успевали склевать всех темных бабочек. После промышленной революции условия изменились, и естественный отбор нашел себе в генофонде немножко уже готовых “темных” генов.
Итак, прежде чем запустить эволюционный процесс, надо иметь две составляющие – мутацию и естественный отбор. Обе они начнут действовать автоматически на любой планете благодаря наличию предшествующих компонентов, один из которых добыть нелегко, но, очевидно, возможно. Этот базовый компонент – наследственность. Для того чтобы где бы то ни было во вселенной заработал естественный отбор, особи должны выстроиться в генеалогические ряды – походить на непосредственных предков больше, чем на других особей во всей популяции. Наследственность – это не то же самое, что размножение. Бывает размножение без воспроизводства наследственных признаков. Лесные пожары размножаются, но их особенности не передаются от одного пожара другому.
Вообразите себе выжженную зноем степь, куда ни кинь взгляд – кругом сухой ковыль. И вот где‐то в этой степи легкомысленный курильщик бросает спичку – в один миг трава вспыхивает и занимается пожар. Наш недотепа бежит прочь, пыхтя и отдуваясь во всю мощь своих отравленных легких, но нас больше интересует то, как распространяется пламя. Огонь быстро расползается во всех направлениях. В небо летят искры. Ветер относит искры и горящие травинки далеко от эпицентра начавшегося пожара. Там, где на пересохшую землю упала искра, возникает новый очаг. Снова летят искры, которые поджигают траву уже в других местах. Можно сказать, для пожара это своего рода процесс воспроизводства, или размножения. У каждого пожара есть свой родитель – пожар, от которого полетела искра-зародыш. Есть и дедушка с прадедушкой – и так далее вплоть до родоначальника, загоревшегося от спички. Каждый новый пожар раскидывает снопы искр в разные стороны, поэтому у него один отец, но сыновей может быть более одного. Если бы вы смогли понаблюдать за процессом сверху и записать “биографию” каждого пожара, у вас получилось бы полное генеалогическое древо степных пожаров.
Однако суть в том, что хотя пожары и размножаются, как таковой наследственности тут нет. Иначе каждый пожар должен был бы иметь больше общих черт со своим непосредственным “родителем”, чем с другими пожарами. В самой идее о сходстве пожара и его предшественника нет ничего ложного. Так вполне может случиться. Пожары меняются и обретают индивидуальные отличия – как и люди. Каждому пожару присущи свои цвет и высота пламени, оттенок дыма, уровень шума и так далее. Любой из этих признаков может быть таким же, как у предыдущего пожара. Если бы все пожары имели такое сходство со своими “отцами”, можно было бы говорить о настоящей наследственности. Но на самом деле пожар похож на своего предка не больше, чем в среднем на любой другой очаг огня в степи. Свойства отдельно взятого очага пламени – масштабы возгорания, цвет дыма, треск – зависят от внешних условий, от того, насколько высокая и сухая трава там, куда попала искра, от скорости и направления ветра. Это все характеристики того места, куда попадает искра, а не пожара, в котором она образовалась.
Чтобы претендовать на наследственное сходство, каждая искра должна переносить какие‐то качества – характерные признаки – родительского пожара. Допустим, в очаге может вспыхнуть пламя какого‐то цвета – желтое, красное или голубое. Если от желтого огня будут лететь желтые искры, от которых разгораются новые желтые пожары, а от красного – красные искры, вновь дающие красное пламя, можно говорить о подлинной наследственности. Но так не бывает. Заметив синий огонь, мы скажем: “Видимо, в этих местах есть соли меди”. Нельзя утверждать, что огонь разгорелся от искры, зародившейся неизвестно где в синем пламени.
Именно этим кролики, люди и одуванчики принципиально отличаются от огня. Но пусть вас не сбивает с толку тот факт, что у кролика два родителя, два деда и две бабки, а у пожара только один непосредственный родитель и один дед. Это существенное отличие, но в данном случае речь не о том. Возможно, с палочниками и тлёй вам будет понятнее, чем с кроликами – самки этих насекомых могут произвести на свет дочерей, внучек и правнучек без участия самца. Форма, окраска, величина и повадки палочника, безусловно, зависят от того, в каком районе он вылупился и какой там климат. Но важную роль играет также искра, которая летит только от родителя к потомку.
Так что же за таинственные искры передаются от родителя потомку, но не наследуются пожаром от предыдущего пожара? На нашей планете это ДНК. Самые удивительные молекулы в мире. ДНК удобно представить себе как информацию, необходимую для создания организмом другого, подобного себе организма. Правильнее было бы рассматривать организм как транспорт, который ДНК использует для производства таких же, как она сама, ДНК. В каждый отрезок времени, например, на нынешнем этапе, все имеющиеся в наличии ДНК ведут род от цельных цепочек успешно развивавшихся предков. Никакие два индивида, за исключением однояйцевых близнецов, не имеют одинаковых ДНК. Индивиды выживают и способны производить такие же ДНК именно благодаря различиям в своих ДНК. Повторю, потому что это важно: вниз по реке спустились те ДНК, которые в течение сотен миллионов лет сохранялись в организмах выживших предков. Многие особи, которые могли бы стать чьими‐нибудь предками, погибли в молодости или не нашли партнера для размножения. Но ни одна из их ДНК не дошла до нас, ныне живущих.
Здесь легко ошибиться – подумать, будто здоровые и благополучные предки передают “переходящей” ДНК некий эликсир удачи, ореол святости. Ничего подобного. От нас в будущее течет чистый (если не принимать во внимание мутации) поток ДНК, который уходит от нас точно так же, как приходит. Точнее, он постоянно перемешивается в процессе полового размножения. Половину ДНК вы получаете от отца, половину – от матери. Различные комбинации ДНК в каждом вашем сперматозоиде и каждой яйцеклетке составлены из генетических ручейков, берущих начало в организмах ваших родителей. Но мои слова остаются в силе. Здоровые предки, “провожая” свои гены в далекое будущее, ничего в них не вкладывают.
Успех живых организмов дарвиновская теория объясняет очень просто. Они процветают благодаря накопленной мудрости своих предков. Но это не та мудрость, которой можно научиться или обрести на жизненном пути. Такая мудрость достается чисто случайно, в результате полезных спорадических мутаций, и избирательно, неслучайно записывается в генетической базе вида. Одному поколению перепадает не слишком много, даже скептически настроенные физики, которых я цитировал, поверили бы в возможность столь малого везения. Но поколение за поколением приумножали успех, и теперь мы имеем результат настолько поразительный, что это кажется невероятным. Все действо по Дарвину зависит от – и вытекает из – факта наследственности. Говоря о наследственности как основной составляющей, я имею в виду, что на любой планете во вселенной, где наблюдается наследственность в каком‐либо виде, почти непременно должен пойти процесс эволюции, то есть должна быть жизнь.
Вот мы и вернулись к горе Невероятности, к “равномерному распределению” удачи, то есть к объяснению того, каким образом некое колоссальное везение – без него, скажем, не сформировался бы глаз там, где его не было, – делится на множество удачных этапов, на каждом из которых предыдущие успехи последовательно закрепляются текущими. Мы уже это видели на примере накопления в ДНК множества небольших “кусочков удачи” выживших предков. Выжило удачливое, с точки зрения генетики, меньшинство, а масса менее везучих индивидов погибла. В каждом поколении кто‐то проигрывает дарвиновский процесс, но любая отдельная особь является потомком только счастливчиков, составлявших меньшинство в предыдущих поколениях.
Послание, которое поступило с горы, содержит три пункта. Первый нам уже известен: нельзя вот так вдруг запрыгнуть наверх, скачкообразное упорядоченное усложнение невозможно. Второе – пути назад, то есть вниз, нет; популяция не может стать менее здоровой в расчете на то, что поправит дела позже. Третье – пик может быть не один, то есть одну и ту же проблему одинаково успешно можно решить разными способами.
Какой орган любого животного или растения ни возьми, возникает резонный вопрос – как постепенные изменения того или иного органа предков привели к его формированию. Иногда удается проследить этот процесс в череде относительно молодых ископаемых животных и растений. Самый популярный пример – эволюция трех слуховых косточек у млекопитающих, благодаря которым звук передается от барабанной перепонки к внутреннему уху с безупречным согласованием импедансов, если этот термин вам что‐нибудь говорит. Судя по ископаемым остаткам, эти косточки – молоточек, наковальня и стремечко – образовались непосредственно из трех соответствующих челюстных костей наших предков-рептилий.
Далеко не все находки палеонтологов так легко трактуются. Иногда о вероятных промежуточных вариантах можно только догадываться и приходится включать воображение, глядя на современных животных, то ли имеющих отношение к интересующей нас проблеме, то ли нет. В слоновьем хоботе нет костей, поэтому не сохранилось и окаменелостей, однако и без таковых нетрудно догадаться, что когда‐то он был обычным носом. Теперь же… впрочем, позвольте привести выдержку из книги Иэна и Ории Дуглас-Хэмилтонов “Битва за слонов” (Iain and Oria Douglas-Hamilton, Battle for the Elephants), перечитывая которую, я каждый раз тайком утираю слезы. В книге чередуются главы, написанные Иэном и Орией, и вот какую горестную картину “выбраковки” слонов, увиденную в Зимбабве, рисует Ория:
Я глядела на отсеченный хобот и думала, сколько же миллионов лет понадобилось эволюции, чтобы сотворить это чудо. Сила захвата и удара оснащенного пятьюдесятью тысячами мышц хобота – сложнейшего органа, работа которого координируется мозгом, – измеряется тоннами. Но вместе с тем он способен выполнять самые отточенные движения, например, аккуратно заправить в рот маленький бобовый стручок. Этот универсальный инструмент служит сифоном на целых четыре литра воды, чтобы слон мог напиться или принять душ, а заодно указательным пальцем и рупором.Хобот – это еще и социально важный орган, с его помощью можно проявить заботу, подбодрить и поприветствовать сородича, пообниматься и выразить готовность к спариванию, а для самца это оружие в схватке, когда соперники сцепляются бивнями и в шутку, а то и всерьез стараются доказать свое превосходство. И вот он валяется, отрубленный, – сколько я видела их по всей Африке!
Вот опять – все тот же эффект, стоило мне прочесть этот абзац…
Таким образом, в послании с горы говорится, что в ряду слоновьих предков должна быть длинная непрерывная цепочка промежуточных вариантов – животных с более или менее вытянутыми носами, таких как тапиры, длинноухие прыгунчики, носачи (кахау), морские слоны. Все они – не родня ни слону, ни друг другу. Все отрастили свои носы независимо друг от друга и, может статься, совершенно по разным причинам (рис. 3.1)
За время эволюции слона из его коротконосых предков носы должны были удлиняться постепенно, без резких изменений, и так же плавно и непрерывно должны были нарастать мышцы и развиваться более сложная и разветвленная нервная регуляция. На каждой следующей ступени, прибавляя дюйм за дюймом, хобот средней длины должен был быть лучше приспособлен для каких‐то новых функций. Если промежуточный хобот ни на что не годится – ни то ни се, ни два ни полтора, нельзя успокаивать себя, что не стоит волноваться, дескать, еще несколько миллионов лет – и все будет прекрасно. Жизнеобеспечение зверей основано не на их неуклонном движении к совершенству в процессе эволюции. Чтобы жить, звери должны есть, не попадаться на обед другим зверям и размножаться. Если бы хобот средней длины всегда отвечал этим требованиям хуже, чем короткий или длинный, последний так никогда и не развился бы.

Рис. 3.1. Африканский слон и другие млекопитающие, которые не имеют к нему никакого отношения, а свои длинные носы приобрели, по-видимому, независимо друг от друга и по разным причинам; против часовой стрелки, начиная сверху слева: носач, Nasalis larvatus; длинноухий прыгунчик, Rhynchocyon petersi; чепрачный тапир, Tapirus indicus; южный морской слон, Mirounga leonina.
Из того, что хобот должен был эффективно функционировать на каждой промежуточной стадии, вовсе не следует, что его функции должны были быть одинаковы. Вероятно, поначалу возможность поднять что‐то с земли не была его главным преимуществом. Может быть, длинный нос прежде всего гарантировал более тонкое обоняние – как у длинноухих прыгунчиков – или играл роль акустического резонатора, как у морских слонов, или, как у обезьян носачей, был призван понравиться самцу – хотя, на наш вкус, это сомнительное украшение. С другой стороны, уже на ранних стадиях эволюции слонов довольно короткий еще хобот мог использоваться в качестве своего рода руки. Предположение не такое уж неправдоподобное, если взглянуть на тапира, который носом срывает листья и отправляет их в рот. Независимая эволюция одного и того же органа у разных животных может многое нам объяснить.
В данном случае, со слоновьим хоботом, кое‐какие выводы можно сделать, если изучить окаменелости твердых частей черепа, особенно бивней и прилегающих к ним костей. До наших дней дошли лишь два вида потомков всех тех разнообразных обладателей бивней, которые некогда бродили по всем континентам. Бивни современного слона – это несоразмерно увеличенные верхние резцы, но у многих ископаемых животных, например у мастодонтов, торчали вперед еще и более выдающиеся нижние резцы. Они могли быть такими же огромными и острыми, как и те бивни, что сейчас мы видим только на нижней челюсти. У других видов бивни были плоские, так что два больших зуба, продолжая линию нижней челюсти, вместе образовывали широкий совок или лопату из кости, с помощью которой можно было выкопать корешки и клубни корнеплодов. Этот совок так далеко выступал за пределы нижней челюсти, что верхняя губа не доставала до пищи, которую удавалось выкопать. Не исключено, что изначально хобот вырос как раз для компенсации функции совка и захвата подкопанной еды. Можно предположить, что позднее зарождающийся хобот оказался весьма удобным инструментом для добывания пищи, и совок с лопатой стали лишними. Еще позже – по крайней мере в сохранившихся генеалогических ветвях – лопата сама собой уменьшилась, в то время как хобот сохранился, словно предмет, оставшийся на суше после отлива. Нижняя челюсть сократилась до примерно исходных пропорций, а новый хобот стал полностью самостоятельным органом. Более подробно об эволюции хобота можно прочесть в блестящей книге Джона Мейнарда Смита “Теория эволюции” (John Maynard Smiths, The Theory of Evolution).
Термин “преадаптация” употребляется в тех случаях, когда орган исходно выполнял какую‐то одну функцию, а потом в процессе эволюции стал использоваться для выполнения совершенно другой функции. Этот термин вносит ясность в ход наших мыслей, поскольку нередко избавляет нас от мучительных раздумий об истоках эволюции. В наши дни дикобраз имеет на вооружении страшные иглы. Они выросли не на пустом месте – это модифицированная шерсть, “преадаптированная” для совершенно другой функции – поддержания температуры тела. У многих млекопитающих есть специфические, хорошо развитые запаховые железы. И не поймешь, откуда они взялись, пока не изучишь их под микроскопом и не увидишь, что они образовались из желез меньшего размера, необходимых для выделения пота и снижения температуры тела. У тех же животных есть и неизменившиеся потовые железы, только иначе расположенные, поэтому нетрудно сравнить железы разного назначения. Другие запаховые железы, по‐видимому, развились из сальных желез, изначально выделявших воскоподобный секрет для защиты шерсти. Прежняя преадаптация и ее современное следствие зачастую не так уж отличаются друг от друга. Пот обычно пахнет и, возможно, выделяется, когда животное нервничает (да и людей часто бросает в пот от страха – по крайней мере, я всегда взмокаю, если во время важного выступления что‐то начинает идти не по плану). Поэтому преадаптация, то есть постепенное формирование тех свойств, которые помогли сместиться в сторону выполнения той функции, что мы наблюдаем сейчас, – процесс вполне естественный.
Порой трудно понять, что было вначале – где преадаптация, а где ее следствие. Изучая эволюционные корни легкого, Дарвин искал ответ в строении и роли плавательного пузыря рыб. С помощью этого наполненного воздухом пузыря, действующего по принципу картезианского водолаза (человечка в бутылке, который то ныряет, то всплывает, если слегка надавить на крышку), костистые рыбы управляют своей плавучестью. Работая мышцами, рыба меняет объем воды в плавательном пузыре, что позволяет ей зависать на определенной глубине. Это относится только к обычной костистой рыбе. У акул, которые, несмотря на вполне рыбью внешность, еще меньше костистые рыбы, чем мы с вами, нет плавательного пузыря, поэтому им, для того чтобы оставаться на нужной глубине, приходится активнее работать плавниками. Плавательный пузырь похож на легкое, и Дарвин подумал, что он мог быть преадаптационным органом, позже развившимся в легкое. Современные зоологи склонны ставить телегу впереди лошади и полагают, что как раз плавательный пузырь относительно недавно развился из примитивного легкого: рыбы, которые дышат воздухом, и сейчас не редкость. Как бы то ни было, мы должны задаться вопросом, какой орган предшествовал более раннему из этих двух. Возможно, легкое или плавательный пузырь образовались из дивертикула кишечника и поначалу участвовали в пищеварении. На каждом этапе эволюции, с каждым шагом на пути к вершине горы Невероятности дивертикул, полость или легкое должны были приносить своему обладателю какую‐то пользу.
Не мог ли слоновий хобот вырасти одним махом, не мог ли это быть один мощный рывок? Почему детенышу родителей с носами, как у тапира, не видать хобота, как у слона? На самом деле здесь кроется три вопроса. Первый – возможны ли мутации с сильным эффектом, или макромутации? Второй – если да, то может ли естественный отбор благоприятствовать таким мутациям? Третий вопрос требует не такого однозначного ответа – что подразумевать под мутацией с сильным эффектом? Вернусь к различиям между макромутациями типа “Боинг-747”и типа “удлиненный Дуглас ДС-8”, которые я описал в моей прежней книге.
На первый из трех вопросов можно ответить положительно. Макромутации действительно происходят. Иногда потомство решительно, до боли в сердце не похоже ни на одного из родителей, ни на других представителей вида. По словам Скотта Гарднера, фотографа из газеты Hamilton Spectator, жабу, изображенную на рис. 3.2, нашли у себя в саду две девочки из города Гамильтона, который находится в канадской провинции Онтарио. У нее отсутствуют глаза на поверхности головы. Когда животное разинуло рот, рассказывал Гарднер, стало ясно, что оно все‐таки реагирует на окружающее. Жабу отправили на ветеринарный факультет Гуэлфского университета для исследования, но сколько‐нибудь подробного отчета я до сих пор не видел. Такие бедолаги интересны тем, что часто дают информацию о нормальном развитии эмбриона. Не все врожденные аномалии развития обусловлены генетикой – взять хотя бы побочные эффекты приема талидомида, – но многие таковы. Ахондороплазия, тяжелое заболевание, связанное с укорочением костей конечностей и приводящее к карликовости и искажению пропорций тела, связана с вариантом одного-единственного гена. Подобные значительные мутации – “макромутации” – иногда называют еще сальтациями. Как правило, вариант гена, являющийся причиной заболевания ахондроплазией, передает потомку один из родителей, но бывает, что болезнь развивается случайно в результате резкого изменения – должно быть, так этот вариант гена и возник когда‐то впервые. Теоретически, хотя я очень сомневаюсь, что это могло произойти на самом деле, в каком‐то одном поколении в результате такой же грандиозной мутации вытянутый нос тапира мог спонтанно вырасти до размеров слоновьего хобота.

Рис. 3.2. Макромутации возможны. В саду одного дома в Канаде нашли вот такого маленького уродца с глазками на нёбе, и он был жив. Впервые фото было опубликовано в городской газете The Hamilton Spectator.
Что касается второго вопроса – может ли глубокая, приведшая к появлению фрика мутация, коль скоро она произошла, оказаться полезной с точки зрения естественного отбора, – то, пожалуй, универсального ответа не найти. Бывает ведь по‐разному – скажем, в случае с ахондроплазией может, а с двухголовым теленком – нет. Собачий аналог мутантного варианта гена ахондроплазии принес огромную пользу селекционерам, которые выводили новую породу не только интереса ради, но и чтобы получить отличного четвероногого помощника. Таксу “укоротили” до формата барсука, и существенную роль в генетических экспериментах, в ходе которых родилась эта порода, сыграло появление мутантного варианта гена ахондроплазии. Вероятно, иногда, как в случае с ахондроплазией, в природе происходят мутации с сильным эффектом, и нежданно-негаданно открывается возможность использования новой жизненной стратегии или пищевого ресурса: вдруг выясняется, что маленький зверек, вечный аутсайдер в гонках на открытой местности, в отличие от своих сородичей способен настичь животное в норе.
Время от времени теоретики-эволюционисты высказывали предположения, что в природе основные сальтации являются неотъемлемой частью эволюционных изменений. Знаменитый американский генетик немецкого происхождения Рихард Гольдшмидт выдвинул не менее знаменитую теорию “удачных монстров”. Об одном из примеров я расскажу в 7 главе. Однако гипотеза Гольдшмидта так и не получила широкого признания, и есть веские причины задуматься, так ли уж важны для эволюционного процесса макромутации. Живой организм – это невероятно сложная и очень хорошо отлаженная машина. Крайне маловероятно, что непродуманные кардинальные изменения в устройстве одного из ее узлов, даже если он работает хуже, чем хотелось бы, дадут положительный эффект. Но за счет случайного незначительного изменения устройства можно заставить механизм работать лучше. Если телевизионная антенна ловит плохо, ваши шансы исправить положение, слегка повернув ее в любую сторону, равны пятьдесят на пятьдесят. Просто вероятность того, что ваше случайное движение приблизит антенну к идеальному положению, каково бы оно ни было, составляет 50 %. Но если вы будете вертеть ее во все стороны и сильно наклонять туда-сюда, то, скорее всего, будет только хуже. Даже угадав направление, вы с большой вероятностью проскочите мимо. В общем и целом, испортить настройку легко, а найти единственно верный угол – трудно. Если мудреный механизм хоть как‐то работает, вряд ли его настройки далеки от идеальных. Можно чуть‐чуть изменить их – неважно, как, – и он станет функционировать более правильно, а если и нет, то разладится не слишком сильно. Но резкие и сумбурные перемены производят тот же эффект, что проверка сразу всех возможных вариантов. И может статься, что почти все эти варианты окажутся неудачными.
Даже распространенное убеждение, что заглохшую машину можно привести в чувство хорошим пинком, не противоречит моим рассуждениям. Удар может быть очень сильным, однако телевизор – достаточно прочная конструкция, и ее элементы необязательно сместятся из‐за толчка. А вот не очень хорошо закрепленная деталь может сдвинуться со своего места, и вполне вероятно, что именно она и является причиной неисправности.
Вернемся к живым созданиям; как я писал в “Слепом часовщике”, выжить можно самыми разными способами, но способов погибнуть, безусловно, гораздо больше (ничто человеческое мне не чуждо, и не скрою, приятно было увидеть эту фразу в “Оксфордском словаре цитат” (Oxford Dictionary of Quotations)). Почти все воображаемые животные, которые могли бы получиться путем перестановки фрагментов целого существа, обречены на смерть. Точнее – почти ни одно из них и не могло родиться. Любой вид животных или растений – это островок жизни в океане вероятных конфигураций, которые, даже появись они паче чаяния на свет, умерли бы. Среди них могли бы быть звери с глазами на пятках, хрусталиками в ушах, а не в глазах, с одним только левым крылом и правым плавником, с черепом, внутри которого находится не мозг, а желудок. Дальше изощряться в выдумках нет смысла. Из всего сказанного и так ясно, что самые крупные острова выживаемости, сколько бы их ни нашлось, ничтожно малы, как ничтожно мало и их количество по сравнению с бесконечным множеством мертвых, нежизнеспособных тварей.
Выживший родитель ребенка-мутанта должен был найти прибежище на одном из таких островков. Дитя с незначительными изменениями – вроде небольшого удлинения костей ног или легкого изменения челюстного угла – просто окажется в другой части того же островка. Возможно, малыш освоит кусочек суши в море недалеко от береговой линии и присоединит его к “большой земле”. Но серьезная и резкая, уродующая мутация, которая может привести к кардинальным переменам, равнозначна гигантскому прыжку в неведомые дали. Макромутанта может занести куда угодно, за многие сотни и тысячи километров от родного дома. Не исключено, что он попадет на другой остров. Но острова столь малы и столь редко встречаются в огромном океане, что вероятность такого события крайне невелика. Подобные редчайшие катаклизмы теоретически могут случиться один раз за несколько миллионов лет, и если это все‐таки случится, эволюция может пойти по новому направлению.
Однако не стоит чересчур увлекаться островными пейзажами. Эта схема во многом ошибочна. Все особи родственны друг другу, то есть между разными дорогами жизни в океане вероятностей должны быть связующие пути. Тут нам скорее поможет аналогия с горой Невероятности, а не с островами. Острова лучше всего помогают проиллюстрировать важный факт – чем заметнее и уродливее отклонение в развитии, тем менее вероятно, что оно приживется.
Кроме того, надо различать типы макромутаций. Я придумал зверей с глазами на пятках и хрусталиками в ушах, чтобы показать возможные перестановки частей организма. Вероятность того, что подобные мутации окажутся удачными и особи с ними выживут, близка к нулю. Но изменение в какой‐либо части может быть довольно сильным и без перестановок. В качестве примера можно было бы взять стремительное удлинение носа тапира до слоновьего хобота, если бы при этом изменился только его размер. Менее очевидно то, что крутые повороты в развитии всегда сопряжены со скачком в океан нерациональности и смерти.
Я обещал вернуться к макромутациям “Боинга-747” и “удлиненного Дугласа ДC-8”. Помните дискуссионный тезис о кладбище самолетов и авиалайнерах, выдвинутый сэром Фредом Хойлом? Говорят, он сравнил эволюцию сложной структуры – скажем, белковой молекулы и, как следствие, глаза или сердца, – которая происходит под влиянием естественного отбора, с ураганом, так продуктивно покружившим над кладбищем старых самолетов, что из поднятых им обломков сложился “Боинг-747”. Он был бы прав, если бы вместо слов “естественный отбор” употребил слово “случай”. К сожалению, я вынужден поставить его на одну доску с теми упрямцами, которые упорно считают естественный отбор делом случая. Теория отводит слишком большую роль случайности, если эволюция должна за один прием, из ничего, собрать новый, сложный механизм вроде глаза или молекулы гемоглобина. Согласно такой концепции, естественному отбору вообще ничего не надо делать. Всю “инженерную” работу берет на себя мутация – одна сильная мутация. В аналогии с “Боингом” подразумевается мутация именно такого типа, и я называю это макромутацией “Боинг-747”. Макромутации “Боинг-747” в природе не происходят и никакого отношения к учению Дарвина не имеют.
В другом моем примере с авиалайнерами удлиненная модель “Дугласа ДС-8” отличается от обычной только относительно вытянутым корпусом. В целом конструкция сохранена, но увеличена средняя часть фюзеляжа. В длинном самолете больше кресел, багажных камер и всего прочего, что может уместиться вдоль салона. Очевидно, по всей длине фюзеляжа протянуто больше проводов, трубок и уложено больше ковровых покрытий. Не так очевидно, но все же понятно, что в связи с новой задачей – поднять в воздух более длинную машину – должны быть изменены и другие детали конструкции. Но в основном различия между прежней версией авиалайнера “Дуглас ДС-8” и удлиненной сводятся к одной макромутации – фюзеляж нового самолета значительно и за один прием стал больше, чем был у его предшественника. Не было постепенного перехода через промежуточные стадии.
Предки жирафов больше напоминали современных окапи (рис. 3.3). Самое заметное, бросающееся в глаза отличие – это вытянутая шея. Могло ли это стать результатом одной мощной мутации? Спешу вас уверить – нет, не могло. Но из этого “нет” вытекает другой вопрос. Сложнейшая “новинка” – глаз со зрачком-диафрагмой и хрусталиком, способным менять фокусное расстояние, – не могла родиться в одночасье, словно богиня Афина из головы Зевса, точно так же, как никогда, жди хоть миллиард миллиардов лет, не произойдет мутация “Боинг-747”. Но шея жирафа могла (держу пари, что все было иначе) вырасти в результате одной-единственной мутации, как можно было взять и построить “Дуглас ДС-8” с удлиненным фюзеляжем. В чем разница? Дело не в том, что шея – куда более простой орган, чем глаз. (Как бы не посложнее глаза!) Вовсе нет – суть в степени сложности различий между двумя последовательными стадиями ее развития. А эти различия невелики – по крайней мере, по сравнению с различиями между нашим глазом и его отсутствием. Строение шеи жирафа и окапи одинаково – и, вероятно, у собственно короткошеего предка жирафа эта часть тела была устроена так же. У них одна и та же последовательность семи позвонков с присущими им кровеносными сосудами, нервами, связками и мышцами. Разница лишь в том, что все позвонки гораздо длиннее – и в той же пропорции увеличивается расстояние между сопряженными тканями и сосудами.
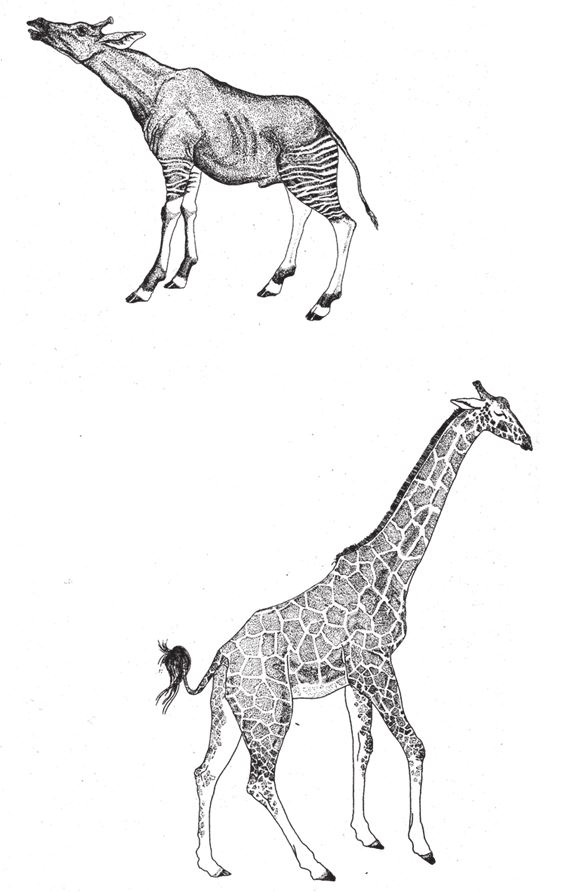
Рис. 3.3. Развитие длинной шеи. Окапи, Okapia johnstoni, возможно, предок современного жирафа, и жираф, Giraffa camelopardalis reticulate.
Вот что главное – для того чтобы шея стала длиннее в четыре раза, достаточно внести какое‐то одно изменение в ход развития эмбриона. Скажем, просто изменить скорость роста зачатков позвонков, а дальше все пойдет по плану. Но для того чтобы на нетронутой коже появился глаз, надо поменять не один параметр, а сотни (см. 5 главу). Если бы у окапи в результате мутации выросла шея, как у жирафа, он изменился бы по типу “удлиненного ДС-8”, а не “Боинга-747”. Следовательно, такой вариант в принципе допустим. Ничего нового не добавляется, меняется только путь формирования органа. Фюзеляж со всем, что к нему прилагается, становится длиннее, а новые компоненты сложной структуры не добавляются, просто растягивается уже существующая структура. Этот принцип работал бы и в том случае, если бы у жирафа было больше семи шейных позвонков. У разных видов змей число позвонков варьируется от 200 до 350. Поскольку все змеи – родственники, а позвонки не могут прирастать четвертинками или половинками, это значит, что время от времени рождаются змеи, у которых на один позвонок больше или меньше, чем у их родителей. Подобные изменения можно отнести к макромутациям, а раз все эти змеи существуют, стало быть, такие макромутации участвуют в эволюционном процессе. Это мутации типа “Дуглас ДС-8”, потому что в данном случае не происходит внедрения новой структуры, как при мутации типа “Боинг-747”, а дублируется та, что уже есть.
Решать задачи эволюции фрику-макромутанту помогает еще и тот факт, что влияние отдельного гена зависит от других генов в организме. Глядя на ген, так просто не поймешь, какова роль его продукта в организме – каков его так называемый фенотипический эффект. Ничто в ДНК-коде гена ахондроплазии не может быть расшифровано молекулярным биологом как “укорочение” или “карликовость”. Эффект этого гена связан с укорочением конечностей только при наличии специфических вариантов множества других генов, не говоря уже об особенностях окружающей среды. Роль гена зависит от контекста, то есть от конкретных условий. Эмбрион развивается в среде, сформированной всеми генами в совокупности. Эффект, который оказывает на развитие эмбриона один ген, определяется состоянием среды. Еще Р. Э. Фишер (я уже его цитировал) много лет назад сформулировал это правило: некоторые гены работают как “модификаторы” влияния других генов. Отсюда, заметьте, вовсе не следует, что гены меняют ДНК-коды других генов. Ни в коем случае. Гены-модификаторы влияют на среду и таким образом – на эффекты, вызванные другими генами, но не на последовательность ДНК других генов.
Как видите, можно (более-менее) допустить, что в результате изменения одного гена – макромутации в одном поколении – родитель с пятнадцатисантиметровым носом тапира способен произвести на свет потомка, нос которого будет похож на почти двухметровый слоновий хобот. Крайне маловероятно, что новый нос сразу будет так же удобен и полезен, как настоящий хобот. Ситуацию теоретически может спасти работа генов-“модификаторов” и “климат”, обеспеченный работой других генов. Если макромутация хоть в какой‐то степени “полезна” и особи из‐за нее не погибают, в ходе дальнейшего отбора вариантов генов-модификаторов произойдет коррекция деталей, и недостатки будут исправлены. Главная мутация – это примерно такое же испытание для популяции, как резкая смена условий существования, например, глобальное похолодание климата. Приход ледникового периода провоцирует отбор целого комплекса генов, и резкая мутация, при которой у организма внезапно вырастает длинный нос, оказывает точно такое же воздействие на ход эволюции.
“Примкнувшие” к основной мутации гены “подчищают” не только самые очевидные последствия макромутации. Их эффекты могут проявиться в любых отделах организма, вдали от центра событий – компенсируется эффект главной мутации, сгладятся ее возможные негативные или усилятся положительные результаты. Голова с хоботом весит больше, поэтому увеличившийся в размерах нос требует более прочных костей шеи. Может сместиться центр тяжести всего тела – а далее, возможно, эстафету подхватят позвоночник и тазовые кости. В итоге отбору подвергаются десятки генов, что отражается на различных органах.
Я обрисовал картину “последующей доработки” для основных макромутаций, но отбор такого рода играет важную роль в эволюции независимо от того, случилась ли на первом шаге макромутация. Даже микромутации могут иметь такие неприятные последствия, что очень желательно было бы “доработать” перестройку организма. Любой ген может стать модификатором для другого гена. Многие гены модифицируют эффекты друг друга. Развивая этот тезис, некоторые эксперты полагают, что большинство генов, в принципе оказывающих хоть какое‐то влияние на организм (а это делают не все гены), модифицируют эффекты большинства других генов. Это еще один аспект идеи о том, что гену приходится выживать в тех условиях, которые в основном формируются под влиянием остальных генов вида.
Надо уточнить еще один момент, по которому может возникнуть недопонимание, пусть даже мы потратим на макромутации чуть больше времени, чем они того заслуживают. В научном мире существует довольно любопытная теория “прерывистого равновесия”. Подробное ее изложение вывело бы меня далеко за рамки основной темы этой книги. Но поскольку эта теория очень популярна и часто трактуется ошибочно, я хочу только подчеркнуть, что концепция прерывистого равновесия не имеет ни малейшей законной связи с макромутацией – и нельзя приписывать ей такое родство. Согласно этой теории, в течение длительного периода эволюционных изменений в филогенетических группах не происходит, они находятся в состоянии стазиса (равновесия), который периодически прерывается всплеском эволюционных изменений, что совпадает с “рождением” нового вида. Однако даже при очень быстрых переменах в эволюционный процесс оказываются вовлечены многие поколения – это все равно процесс постепенных изменений. Просто промежуточные состояния исчезают так быстро, что в ископаемых остатках они не могут быть обнаружены. “Прерывистость в виде последовательности частых перемен” и макромутация, то есть мгновенное изменение в одном поколении, – это совершенно разные понятия. В путанице отчасти виноват Стивен Гулд, один из двух апологетов этой теории (второй – Найлз Элдредж), который вообще‐то неплохо относится к отдельным типам макромутаций, но иногда отрицает значительную разницу между быстрой сменяемостью и истинной макромутацией – не подумайте только, что это относится к мутации типа “Боинг-747”. Элдредж и Гулд имели все основания сердиться на креационистов, искажавших их идею и полагавших, будто прерывистое равновесие, если пользоваться моей терминологией, аналогично гигантской мутации “Боинг-747”, где, как справедливо считают ученые, без волшебной палочки не обойтись. Гулд говорит:
Поскольку мы предлагаем модель прерывистого равновесия именно для того, чтобы объяснить существующие эволюционные тенденции, нас бесит, когда нам снова и снова – уж не знаю, сознательно или по недоумию, – приписывают утверждение, что в палеонтологической летописи нет промежуточных форм. Как правило, на уровне вида переходные формы не наблюдаются, но между большими группами они присутствуют в изобилии.
Если бы доктор Гулд более четко показал различия между быстрой сменой промежуточных состояний и сальтацией (то есть макромутацией), риск того, что его поймут неправильно, был бы меньше. Теорию прерывистого равновесия можно считать умеренной (и, возможно, верной) или революционной (и, вероятно, ложной) – это зависит от определений. Если грань между быстрым градуализмом и сальтацией размыта, теория прерывистости выглядит более радикальной. Но в то же время это постоянно действующее приглашение к ошибочной трактовке, и креационисты с удовольствием его принимают.
Причина, по которой на уровне вида переходные формы обычно отсутствуют, проста до банальности. Дети растут долго и превращаются во взрослых постепенно, но с юридической точки зрения один день рождения – как правило, восемнадцатый – особенный. То есть можно утверждать, что каждый из 55 миллионов человек, составляющих население Великобритании, либо имеет право голосовать, либо нет – промежуточного статуса не бывает. Согласно закону, как только часы пробьют полночь и наступит восемнадцатый день рождения, молодой человек превращается в избирателя – вот и зоолог должен обязательно зачислить особь в тот или иной вид. Если фактически особь занимает промежуточное положение – а таких особей немало, – зоологи, присваивая ей имя и действуя в рамках научного кодекса, все равно вынуждены что‐то выбрать. Следовательно, в отношении видов креационисты по определению должны быть правы, заявляя, что не может быть переходного состояния, но к живой природе это неприменимо – разве что к правилам классификации в зоологии.
Взять хотя бы наших собственных предков: австралопитеки, человек умелый, человек прямоходящий, древний человек разумный и современный человек разумный сменяли друг друга так плавно и постепенно, что между учеными не стихают ожесточенные споры по поводу того, к какому виду принадлежит данная окаменелость. А вот пример антиэволюционной пропаганды, взятый из одной книги: “Находку отнесли то ли к австралопитекам – к приматам, – то ли к роду Homo, а стало быть, к людскому роду. Более ста лет ведутся активные полевые изыскания и горячие дискуссии, однако витрина, зарезервированная для гипотетического предка человека, пока пустует. Недостающего звена по‐прежнему недостает”. Остается только гадать, какими должны быть окаменелые остатки, чтобы их можно было считать промежуточным видом. На самом деле в приведенной выше цитате ничего не сказано о реальном мире. Что‐то такое, довольно мутное, говорится о том, какие вещи какими именами называть. Надо всеми недостающими звеньями – казалось бы, теми самыми промежуточными видами – довлеет авторитет терминологии, которая отбрасывает его за черту или оставляет перед ней. Чтобы найти промежуточные варианты, лучше всего отвлечься от вопросов номенклатуры и посмотреть на их форму и размеры. Тогда вы поймете стройную логику палеонтологической летописи, хотя в ней есть пробелы, и порой крупные – как полагают, они остались просто потому, что когда‐то нарушился процесс окаменения. В какой‐то степени наша система присвоения имен разработана для той эпохи, когда об эволюции речи еще не шло, все со всем было четко разделено, и мы даже не надеялись найти промежуточные виды.
Мы бегло осмотрели гору Невероятности и обнаружили два различных пейзажа: неприступные скалы с одной стороны и пологие подъемы – с другой. Две следующие главы посвящены утесам чуть ли не с самыми обрывистыми склонами, мимо которых креационисты не могут пройти спокойно – а именно, крыльям (“что толку в половине крыла?”) и глазам (“пока все многочисленные элементы глаза не займут нужное положение, он не будет функционировать, поэтому глаз не мог эволюционировать постепенно”).
Назад: Глава 2 Шелковые путы
Дальше: Глава 4 Взлет разрешен

