Вкладка
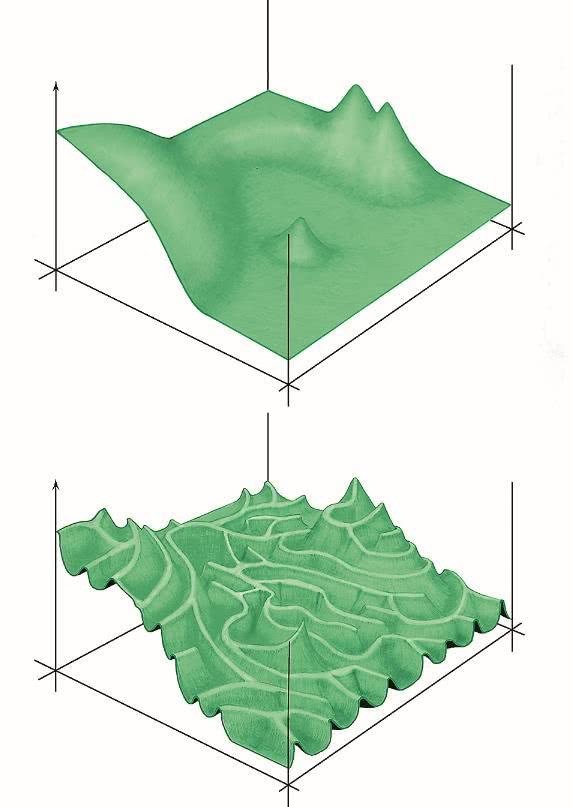
Илл. I. Примерно так можно изобразить ландшафт приспособленности.
Два горизонтальных измерения символизируют “пространство последовательностей”, то есть все возможные варианты аминокислотной последовательности белка.
В действительности пространство последовательностей многомерно: в нем столько же измерений, сколько аминокислот в белке, и по каждому измерению белок может занимать одну из 20 позиций. Но такое пространство невозможно изобразить на бумаге, поэтому для простоты и наглядности его обычно схлопывают в два измерения.
Вертикальное измерение – эффективность выполнения белком своей функции (здесь мы опять сильно упрощаем реальность, предполагая, что чем эффективнее белок выполняет свою функцию, тем выше приспособленность организма). Естественный отбор двигает эволюционирующий белок вверх по склонам и препятствует спускам. Если эффекты мутаций слабо зависят друг от друга, ландшафт оказывается легкопроходимым (график вверху), что позволяет эволюции быстро освоить все потенциально доступные области.
В случае сильного эпистаза ландшафт превращается в лабиринт тропинок, разделенных пропастями (график внизу). В такой ситуации многие области пространства последовательностей могут остаться не разведанными эволюцией – просто потому, что до них трудно добраться.
Рисунок Андрея Томиленко.
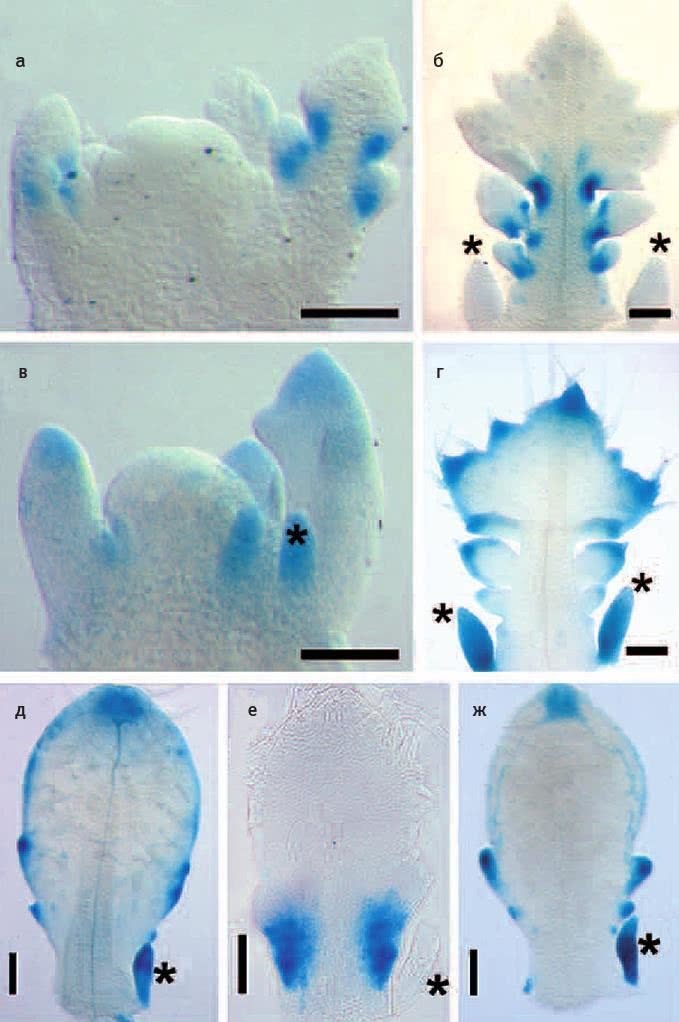
Илл. II. Области экспрессии генов RCO и LMI1. а – г: в зачатках листьев сердечника регуляторная область RCO обеспечивает экспрессию в основаниях лопастей (будущих вторичных листочков; а, б), а регуляторная область LMI1 срабатывает на концах лопастей (в, г). д – ж: в зачатках листьев резуховидки регуляторная область LMI1 (д – своя, ж – сердечника) обеспечивает экспрессию на конце листа и на вершинах мелких выступов по краю листовой пластинки, а регуляторная область RCO сердечника (е) срабатывает во внутренних областях листовой пластинки у ее основания. Звездочками отмечены прилистники. Длина масштабных линеек – 100 мкм. Из Vlad et al., 2014.

Илл. III. Полиморфизм окраски у тлей Macrosiphoniella yomogicola.

Илл. IV. Тринидадские гуппи – удобный объект для изучения быстрых эволюционных изменений. Из синопсиса к Ghalambor et al., 2015.
С сайта blog.tamagaro.net.

Илл. V. Восхитительные узоры на крыльях бабочек складываются из отдельных чешуек, различающихся по цвету, форме и строению. Окраска одних чешуек определяется пигментами, других – тонкослойной интерференцией. Каждая чешуйка одноцветна и образуется единственной клеткой.
Cornel Constantin / Используется по лицензии Shutterstock.com.

Илл. VI. Примеры параллельной эволюции цихлид в трех больших африканских озерах. Звездочками отмечены виды, геномы которых были прочтены.
По рисунку из синопсиса к Brawand et al., 2014.
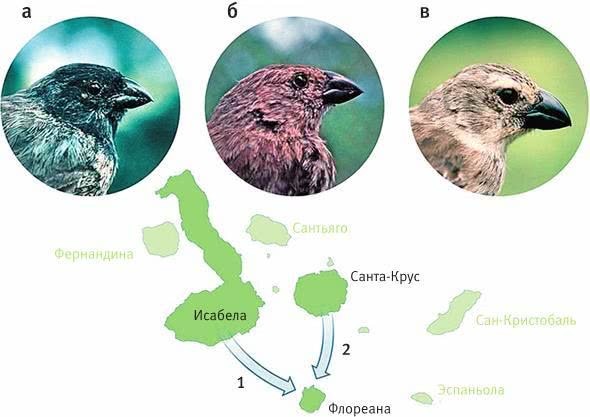
Илл. VII. Три вида древесных вьюрков с острова Флореана: малый (а), большой (б) и попугайный (в).
Большие древесные вьюрки встречаются только на Флореане. Они произошли от попугайных древесных вьюрков, прилетевших некогда с острова Исабела (стрелка 1) и впоследствии измельчавших и превратившихся в отдельный эндемичный вид. Другая группа попугайных вьюрков, попавшая на Флореану с острова Санта-Крус (стрелка 2), не претерпела существенных изменений, но к настоящему времени уже вымерла. Малые древесные вьюрки обитают на многих островах архипелага. Откуда они прилетели на Флореану, точно не известно.
По рисунку из Grant, Grant, 2014.
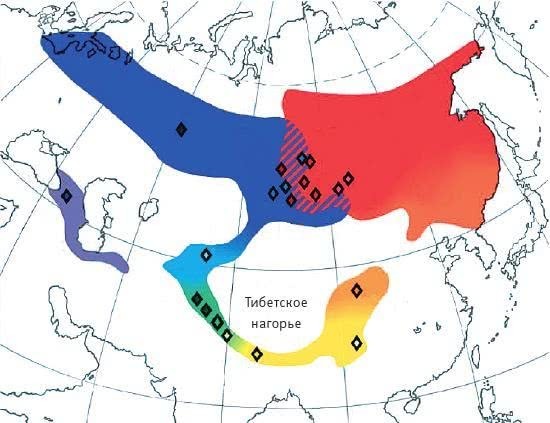
Илл. VIII. Ареал зеленой пеночки (Phylloscopus trochiloides). Разными цветами обозначены территории обитания разных подвидов: желтый – P. t. trochiloides, зеленый – ludlowi, сиреневый – nitidus, синий – viridanus, красный – plumbeitarsus, оранжевый – obscuratus. Ромбиками обозначены точки сбора материала.
По рисунку из Alcaide et al., 2014.
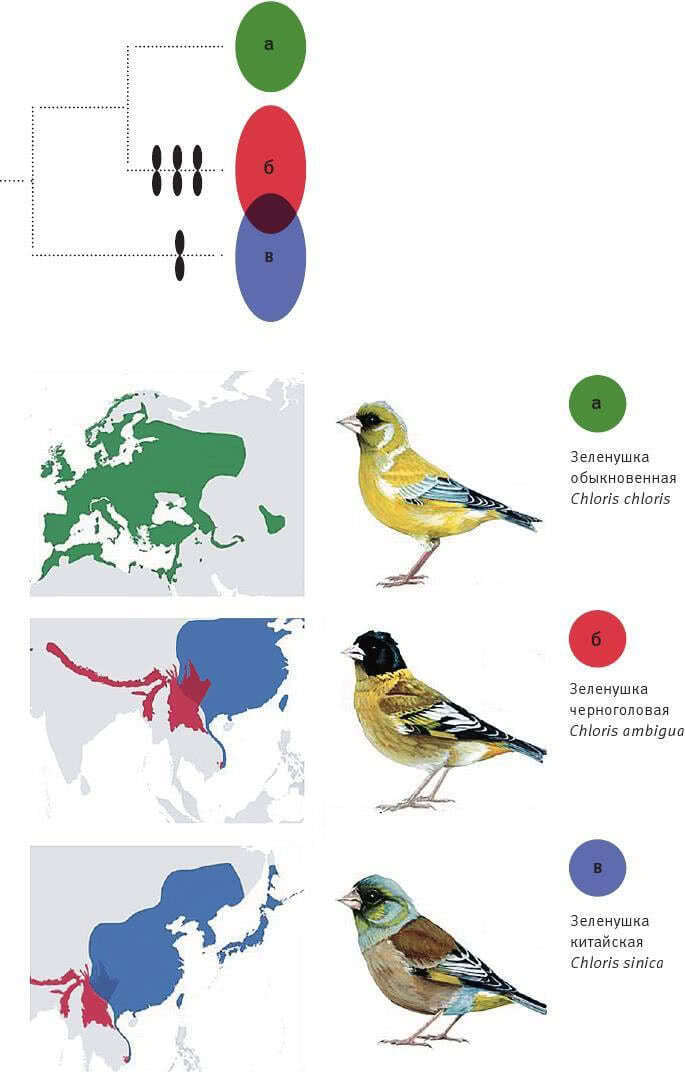
Илл. IX. Фиксация хромосомных инверсий у зеленушек (обыкновенной – Chloris chloris, черноголовой – C. ambigua и китайской – C. sinica).
Время отделения C. sinica от общих предков двух других видов – 2,2 млн лет назад. Инверсии обозначены черными “восьмерками”. Всего зафиксировалось четыре инверсии, причем между симпатрическими видами C. ambigua и C. sinica число различий по инверсиям максимально (четыре), а у C. chloris, чей ареал не пересекается с ареалами близких видов, не зафиксировалось ни одной инверсии.

Илл. X. Табак обыкновенный (Nicotiana tabacum), табачное дерево (N. glauca) и их аллополиплоидный потомок N. tabauca.
Верхний ряд демонстрирует быстрый рост нового растения (показаны экземпляры одинакового, 44-дневного, возраста).
Второй и третий ряды – различия в форме и окраске листьев: у N. glauca пигменты антоцианы распределены по всему листу, у N. tabacum они отсутствуют, у гибридного N. tabauca – имеются только возле черешка.
Нижние ряды показывают различия в строении цветков. Длина масштабной линейки – 20 см. Из Fuentes et al., 2014.
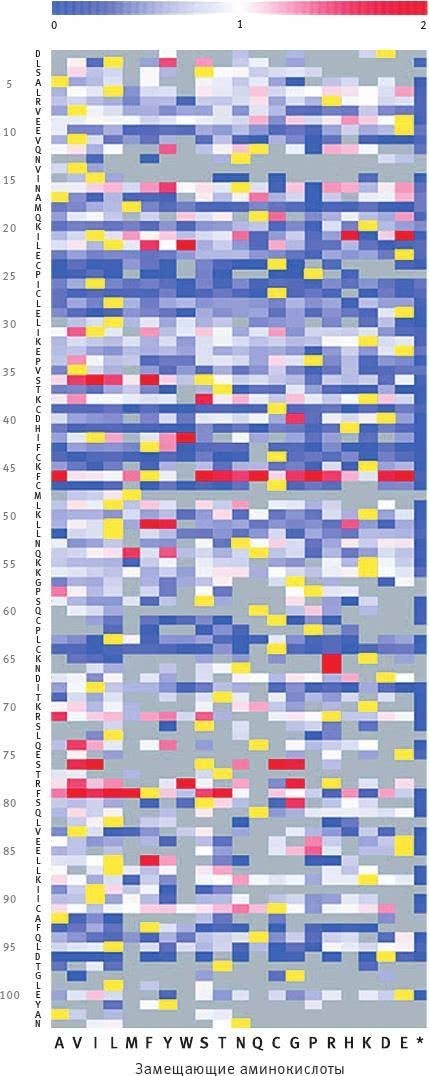
Илл. XI. Результаты проверки того, как влияют на убиквитин-лигазную функцию BRCA1 аминокислотные замены в каждой из 102 аминокислотных позиций исследованного фрагмента белка.
Аминокислотные позиции расположены вдоль вертикальной оси и подписаны в левой части диаграммы.
Для каждой позиции разными цветами показаны эффекты от замен. По горизонтальной оси отложены 20 аминокислот, которыми исследователи поочередно заменяли исходную в каждой позиции (звездочка – выпадение аминокислоты).
Желтыми прямоугольничками отмечены аминокислоты “дикого типа”, то есть те, что стоят в данной позиции в нормальном, немутантном белке. Синий цвет обозначает ослабление функции, красный – ее усиление, белый – сохранение исходного уровня активности белка (см. цветовую шкалу вверху; единица соответствует исходному состоянию, то есть норме). Наконец, серый цвет обозначает, что для данной замены результаты получены не были.
По количеству белых и почти белых прямоугольничков можно судить о толерантности данного белка к мутациям, то есть о спектре несинонимичных (ведущих к замене аминокислоты) мутаций, которые не приводят к драматическим изменениям функциональности белка.
Из Starita et al., 2015.

