Исследование № 13
Половой отбор может довести до вымирания
Как показывают рассмотренные выше примеры, эволюционные последствия полового отбора могут быть весьма разнообразными. С одной стороны, половой отбор способствует отбраковке вредных мутаций, что может положительно сказаться на долгосрочных перспективах существования вида. С другой стороны, иногда он приводит к развитию адаптаций, выгодных одному из полов, но потенциально вредных для вида (яркий пример такой адаптации – мужской инфантицид; см. Исследование № 12). Как выясняется, половой отбор способен даже доводить виды до вымирания. На это указывают факты, опубликованные американскими палеонтологами в 2018 году. Проанализировав распространение 93 видов остракод (ракушковых раков) в верхнемеловых отложениях юго-востока США, ученые обнаружили связь между силой полового отбора и вероятностью вымирания вида. Характеристикой силы полового отбора служила степень полового диморфизма (межполовых различий) по размеру и форме раковины. В основе этих различий у остракод лежит гипертрофия мужских половых органов, порождаемая спермовыми войнами. Оказалось, что темпы вымирания у видов с максимальным диморфизмом на порядок выше, чем у видов с минимальными межполовыми различиями. Результаты согласуются с предположением, согласно которому половой отбор может ускорять вымирание видов, стимулируя развитие громоздких, дорогостоящих адаптаций, повышающих конкурентоспособность в борьбе за половых партнеров ценой снижения приспособляемости к переменам среды.


Влияние полового отбора на риск вымирания вида теоретически может быть двояким. С одной стороны, половой отбор способствует эффективной отбраковке вредных мутаций, тем самым уменьшая генетический груз. Этот эффект был подтвержден в изящном эволюционном эксперименте на жуках, о котором рассказано выше (см. Исследование № 8). Уменьшение генетического груза, по идее, должно снижать вероятность вымирания вида.
С другой стороны, половой отбор часто поддерживает развитие дорогостоящих адаптаций, снижающих жизнеспособность либо того пола, который обладает этими адаптациями (как в случае с громоздкими украшениями и турнирным оружием самцов), либо противоположного (в этом случае говорят о “конфликте полов”). Логично предположить, что это должно повышать вероятность вымирания.
Проверять подобные предположения лучше всего на палеонтологическом материале, который позволяет оценить реальные темпы вымирания видов (конечно, если палеонтологическая летопись достаточно полна). Риск вымирания современных животных приходится оценивать косвенно – например, по охранному статусу или по величине генетического груза, который, в свою очередь, оценивается по другим косвенным признакам, таким как темп снижения приспособленности в инбредных линиях.
Проблема, однако, в том, что найти подходящих для такого анализа ископаемых животных очень непросто. Во-первых, их “захоронения” должны быть массовыми, а ископаемая летопись – подробной, чтобы моменты появления и вымирания видов поддавались точному датированию. Во-вторых, у этих животных должен быть четкий половой диморфизм, по выраженности которого можно судить о силе полового отбора. Причем диморфизм должен затрагивать не только мягкие ткани, крайне редко сохраняющиеся в ископаемом состоянии, но и скелет. Желательно также, чтобы группа не была полностью вымершей (как, например, аммониты) и имела живых представителей, которые позволили бы убедиться, что различия, принимаемые нами за межполовые, действительно таковы (а не межвидовые или, скажем, возрастные).
Чуть ли не единственной группой, удовлетворяющей этим критериям, являются остракоды, или ракушковые, – вездесущие мелкие ракообразные, населяющие моря и пресные воды с кембрийского периода по настоящее время. Тело остракод заключено в прочную двустворчатую раковину, отлично сохраняющуюся в ископаемом состоянии. При этом у многих остракод хорошо выражен половой диморфизм (рис. 13.1).
У современных представителей цитероидных остракод (надсемейство Cytheroidea) раковинки самцов сильнее вытянуты в передне-заднем направлении. Это связано с тем, что в задней части раковины у самца помещается громоздкий копулятивный аппарат. Хотя сам аппарат не сохраняется в ископаемом состоянии, форма створок недвусмысленно свидетельствует о его наличии или отсутствии, что позволяет уверенно отличать самцов от самок на ископаемом материале (рис. 13.2).
У современных цитероидных остракод размер мужских половых органов не связан с какими-либо особенностями брачного поведения (ухаживанием или сигналами, подаваемыми самке), а связан лишь с количеством производимой спермы и силой мышц, которые эту сперму выбрасывают во время спаривания. По-видимому, гипертрофия половых органов у самцов является результатом спермовых войн.
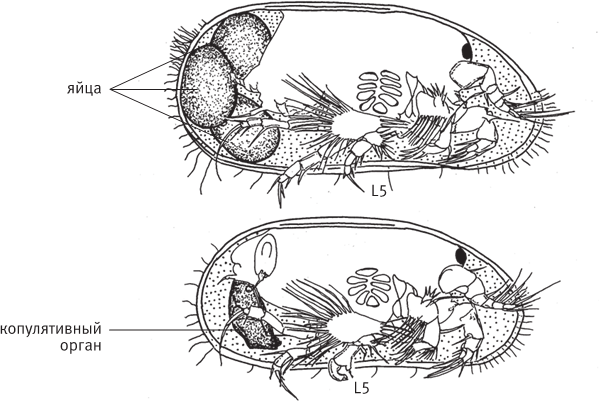
рис. 13.1. Половой диморфизм у современной остракоды Vestalenula cornelia. Вид справа, правая створка удалена. Вверху – самка, внизу – самец с крупным копулятивным органом. Половой диморфизм затрагивает также строение пятой пары конечностей (L5), которую самцы используют для удержания самки во время копуляции. По рисунку из Ozawa, 2013.
Если самцу приходится тратить ресурсы на выращивание огромных половых органов, то у него, при прочих равных условиях, должно оставаться меньше сил на все остальное. Спермовые войны предполагают острую конкуренцию между самцами, что может вредить здоровью самок. В таком случае самки в ответ могут выработать защитные адаптации, тоже затратные. По остракодам таких данных нет, но это следует из общих соображений и показано на других членистоногих (см. Исследование № 15). Меняя морфологию и физиологию самцов и самок, сильный половой отбор, очевидно, уводит их от того оптимального строения, к которому привел бы их обычный естественный отбор в отсутствие полового. Иными словами, организмы приобретают строение, неоптимальное с точки зрения выживаемости и способности переносить невзгоды, но связанное с взаимоотношениями между полами. Поэтому логично предположить, что виды остракод с сильным половым диморфизмом будут вымирать в среднем чаще, чем виды со слабым диморфизмом.

рис. 13.2. Половой диморфизм у двух видов позднемеловых цитероидных остракод. а – очертания раковин самцов и самок Krithe cushmani, б – Veenia ponderosana. На графике (в) по вертикальной оси отложена степень удлиненности раковины (логарифм отношения длины к высоте), по горизонтальной – размер раковины (логарифм площади створки). Видно, что у обоих видов самцы имеют более удлиненную раковину, чем самки (это справедливо и для всех остальных цитероидных остракод). Что касается диморфизма по размеру, то он может иметь разную направленность: у K. cushmani самцы мельче самок, у V. ponderosana – крупнее. По рисунку из Martins et al., 2018.
Для проверки этого предположения американские палеонтологи проанализировали подробнейшие данные по распространению 93 видов цитероидных остракод в верхнемеловых отложениях юго-востока США, штаты Миссисипи и Алабама (Martins et al., 2018). Возраст изученных слоев – от 66 до 84 млн лет. Исследованные виды сильно различаются по выраженности полового диморфизма.
Ученые пытались обнаружить связь наблюдаемой динамики появления и вымирания видов со всевозможными комбинациями таких параметров, как форма раковины, ее размер, степень полового диморфизма по различным признакам, частота встречаемости вида, его таксономическое положение и продолжительность существования.
Выяснилось, что вероятность вымирания видов тесно связана с половым диморфизмом по форме раковины и чуть слабее – с диморфизмом по размеру. Средние темпы вымирания у видов с максимальным диморфизмом примерно в десять раз выше (а средняя продолжительность существования, соответственно, меньше), чем у видов с минимальным диморфизмом. При этом диморфизм по размеру повышает вероятность вымирания только в том случае, если самцы крупнее самок. В обратной ситуации – когда самцы мельче самок – диморфизм по размеру не увеличивает, а скорее даже немного уменьшает вероятность вымирания. Ни форма раковины, ни ее размер сами по себе не влияют на риск вымирания – только величина различий между самцами и самками.
Таким образом, наиболее склонными к вымиранию оказались виды с признаками, указывающими на то, что самцы вкладывают в размножение очень много ресурсов.
Кроме диморфизма, на вероятность вымирания вида влияет его таксономическая принадлежность: в одних семействах виды вымирали чаще, в других реже. Это значит, что в данных по скорости вымирания есть “филогенетический сигнал”. Иными словами, виды, происходящие от короткоживущих видов, с повышенной вероятностью сами окажутся короткоживущими, и наоборот. Впрочем, это не новость, а общая закономерность в палеонтологии. Разные таксоны имеют свои характерные скорости вымирания родов и видов. К примеру, у головоногих моллюсков среднее время существования родов в фанерозое составляло всего лишь 6-7 млн лет, а у двустворчатых моллюсков – примерно на порядок больше.
Еще один параметр, влияющий на вероятность вымирания видов остракод, – частота встречаемости. Виды с высокой численностью особей и широкими ареалами вымирали реже, чем малочисленные виды с небольшими ареалами. Это опять-таки не новость, а одна из самых четких и строгих закономерностей в исторической динамике биоразнообразия.
Таким образом, полученные результаты согласуются с предположением о том, что сильный половой отбор, приводящий к резко выраженному половому диморфизму по форме и размеру раковины, повышает вероятность вымирания у цитероидных остракод.
Теоретически половой отбор может способствовать ускоренному видообразованию (см. Исследование № 11). Однако данные по верхнемеловым остракодам юго-востока США не подтвердили эту идею: достоверной связи между темпами появления новых видов и выраженностью у них полового диморфизма ученые не обнаружили.
Означают ли полученные результаты, что половой отбор никогда не помогает видам противостоять вымиранию (на что вроде бы указывают данные по современным животным; см. Исследование № 8), а всегда только мешает? Вряд ли. Скорее всего, влияние полового отбора на эволюционную судьбу видов может быть как положительным, так и отрицательным. Положительное влияние связано с более эффективной отбраковкой слабовредных мутаций и, возможно, с ускорением дивергенции и видообразования. Отрицательное – с конфликтом полов и развитием дорогостоящих адаптаций, повышающих шансы на победу в конкуренции за половых партнеров ценой снижения жизнеспособности и приспособляемости.

Будем надеяться, что дальнейшие исследования прольют свет на факторы, от которых зависит, какой из двух эффектов окажется сильнее в каждом конкретном случае. Однако палеонтологическая летопись, вероятнее всего, сможет дать нам лишь примеры негативного влияния сильного полового отбора на выживаемость видов. Ведь если половой отбор действует мягко и не становится причиной возникновения гротескных межполовых различий, мы просто не сможем заметить его признаки на ископаемом материале.

