Исследование № 14
Из-за конкуренции самцов страдают самки
Удивительно, насколько логичными становятся самые сложные явления, если смотреть на них сквозь призму отбора. Конфликт полов – многоликий и многообразный – становится понятным и объяснимым с позиций полового отбора. Добавим к половому отбору отбор на выживаемость – получим поддержание полиморфизма в популяции (см. Исследование № 10). Скомбинируем половой отбор и родственный – и вот уже у родственников острота конкуренции за партнеров снижается, а конфликт полов сглаживается. Еще бы, ведь эволюционная задача у родственников общая – распространить и передать следующим поколениям сходные, родственные наборы генов. Именно так, согласно теории, должно обстоять дело. Кому-то это может показаться удивительным, но даже в неоднозначных ситуациях, когда действуют разнонаправленные векторы отбора – полового и родственного, – практика подтверждает теорию. Так, были проведены эксперименты, в которых удалось смоделировать совместное действие полового и родственного отбора. Мы рассмотрим два из них: один – на плодовых мушках дрозофилах, другой – на корневых клещах. Оба исследования показывают, насколько предсказуемыми могут быть результаты отбора (если, конечно, в распоряжении исследователей есть адекватные теории, позволяющие эти результаты предсказывать). В первом эксперименте, поставленном биологами из Оксфордского университета (Великобритания), за самок конкурировали либо родственные друг другу самцы, либо неродственные. Самцы-чужаки, помещенные в пробирку с самкой, чаще дрались и агрессивнее ухаживали за дамой, чем родные братья в такой же ситуации. Из-за этого самка быстрее теряла с возрастом плодовитость и за свою жизнь успевала оставить меньше потомков. А если за самкой ухаживали братья, то она дольше оставалась плодовитой и производила в итоге больше потомства. Так что братские гены тоже оставались в выигрыше. О втором эксперименте мы расскажем в следующей главе.


Различие мужских и женских репродуктивных стратегий изначально основано на том, что самец может произвести гораздо больше сперматозоидов, чем самка – яйцеклеток. Поэтому в типичном случае женский репродуктивный ресурс – в дефиците, а мужской – в избытке. Как следствие, репродуктивный успех самца сильно зависит от исхода его конкуренции с другими самцами. Для самок в большинстве случаев конкуренция за самцов менее актуальна. Самцу выгодно спариться с максимальным количеством самок, а для этого нужно соревноваться с другими самцами, преследующими ту же цель. Репродуктивный успех самки обычно зависит скорее от “качества”, чем от количества ее половых партнеров (хотя иногда количество тоже бывает важно; см. Исследования № 11 и 12).
Несовпадение оптимальных линий поведения самцов и самок ведет, как мы уже говорили, к конфликту полов. Напомним, что это ситуация, когда признаки, повышающие репродуктивный успех одного из полов, снижают приспособленность другого. Например, самцы многих видов животных (от гуппи и гусей до овец и орангутанов) практикуют насильственную копуляцию, хотя порой это плохо сказывается на плодовитости самок и здоровье потомства (Gasparini et al., 2012).
Острый конфликт полов может угрожать выживанию популяции. В общем случае для популяции было бы лучше, если бы самцы не изводили самок, снижая тем самым их плодовитость. Но каждый отдельный самец все же оставит больше потомства, если будет вести себя агрессивно и назойливо. Поэтому “гены сексуальной агрессии” распространяются в генофонде вопреки тому, что они вредны для популяции. Это способно даже привести к вымиранию. Печальный пример – горные гориллы, вымирающий вид, который сейчас насчитывает всего 800-900 особей. У них ярко выражен конфликт полов, проявляющийся в жестоком мужском инфантициде. С позиций каждого отдельного самца убийство чужих детенышей вполне оправданно, а что это ускоряет вымирание популяции – для них значения не имеет, ведь отбор очень близорук, отдаленное будущее его не заботит, равно как и “благо вида”.
Однако помимо полового отбора существует еще и родственный. Мы подробно рассказали о нем во втором томе нашей книги “Эволюция человека”. Родственный отбор по праву считается одним из важнейших механизмов, способных противостоять распространению “генов эгоистичного поведения”. В основе теории родственного отбора лежит простая идея, согласно которой производство собственных потомков – не единственный способ передать свои гены следующим поколениям. Того же результата можно добиться, помогая выживать и размножаться родственникам. Поэтому, например, пожертвовать собой ради спасения трех родных братьев или сестер – выгодно с точки зрения распространения генов, влияющих на принятие такого решения. Теория предсказывает, что в определенных ситуациях отбор должен поддерживать генетические варианты, способствующие альтруизму по отношению к родственникам.
В условиях сильного конфликта полов родственный отбор теоретически может снизить остроту конкуренции за самок, если конкурирующие самцы – близкие родственники. Это предположение удалось проверить на практике (Carazo et al., 2014). Для подобных экспериментов прекрасно подходят плодовые мушки Drosophila melanogaster, потому что у дрозофил ярко выражен конфликт полов. В частности, известно, что чрезмерные домогательства самцов снижают плодовитость самок. Кроме того, дрозофилы хорошо умеют отличать по запаху своих от чужих (Tan et al., 2013).
В пробирки сажали по четыре мухи: одну самку и трех неродственных ей самцов. В одних пробирках самцы были также неродственны и друг другу (назовем эту ситуацию ABC), в других они были родными братьями (ААА). В полном соответствии с теорией родственного отбора в первом случае репродуктивный успех самок оказался ниже (рис. 14.1 и 14.2).
Самки, делившие пробирку с тремя неродственными самцами, произвели за свою жизнь в среднем по 165 потомков, тогда как у самок, посаженных с тремя братьями, потомков оказалось заметно больше, в среднем по 210 (рис. 14.2). Низкий репродуктивный успех самок в ситуации ABC объясняется тем, что у них наблюдалось ускоренное репродуктивное старение. Иными словами, их плодовитость снижалась с возрастом быстрее, чем у самок в ситуации ААА.
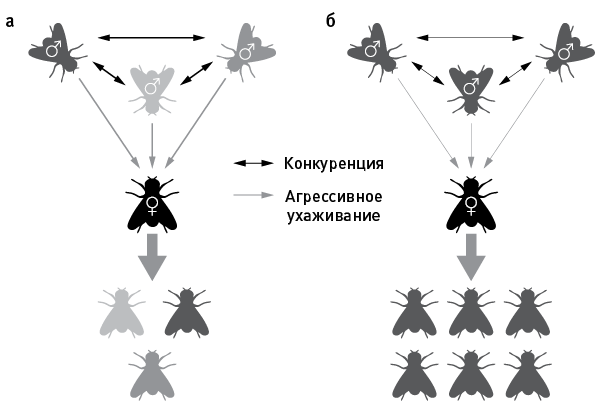
рис. 14.1. Схема эксперимента по влиянию родства между самцами на репродуктивный успех самок дрозофил. Три неродственных самца, посаженные в пробирку с самкой (а), агрессивно конкурируют друг с другом и назойливо ухаживают за дамой. В результате та быстро стареет и оставляет меньше потомков. Три родных брата в такой же ситуации (б) конфликтуют реже и не так активно пристают к самке. В результате она стареет медленнее и оставляет многочисленное потомство. По рисунку из Carazo et al., 2014.
Наблюдения за самцами показали, что в ситуации ABC те энергичнее конкурировали за самку. Они чаще дрались, отталкивая друг друга от самки, и активнее ухаживали за своей сожительницей, что проявлялось в повышенной частоте случаев, когда этим занимались два или три самца одновременно. Острая конкуренция сокращала жизнь не только самок, но и самцов: в ситуации ABC они прожили в среднем по 40 дней, а в ААА – по 47. При этом частота и продолжительность спариваний были, как ни странно, примерно одинаковы в обоих случаях. Таким образом, снижение репродуктивного успеха самок было связано не с частыми копуляциями, а с назойливыми и агрессивными ухаживаниями.
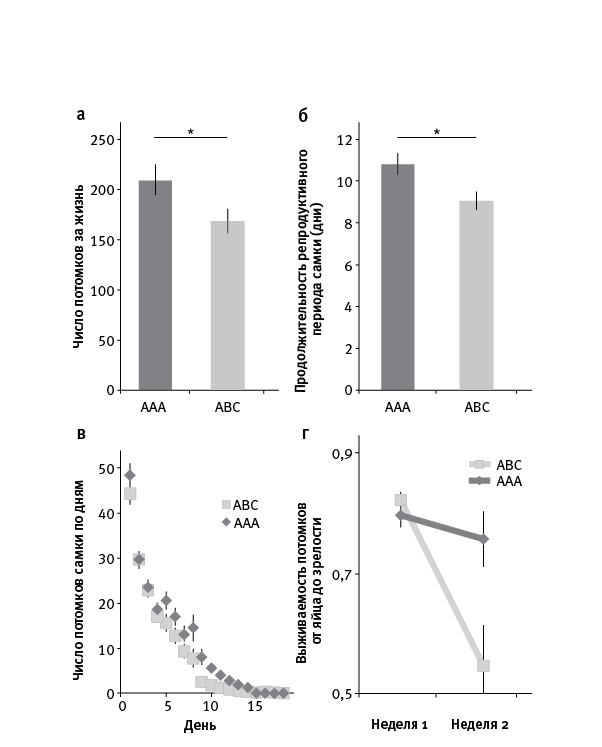
рис. 14.2. Родство между самцами повышает репродуктивный успех самок. a – общее число потомков, произведенных самкой, в двух экспериментальных ситуациях: когда конкурирующие за нее самцы родственны друг другу (AAA) и неродственны (ABC). б – число дней, в течение которых самка продолжала размножаться (от вылупления из куколки до откладки последнего яйца). в – количество яиц, отложенных самкой в каждый день жизни. Видно, что репродуктивная функция самок в ситуации ABC снижается быстрее, чем в AAA. г – выживаемость потомства (доля яиц, из которых вывелись потомки, дожившие до зрелости) в зависимости от возраста самки. Видно, что в ситуации ABC этот компонент приспособленности самок тоже снижается быстрее. Везде представлены усредненные данные. По рисунку из Carazo et al., 2014.
Пониженную конкуренцию между братьями можно рассматривать как своеобразную “родственную кооперацию”. Как известно, системы, основанные на кооперации, создают плодородную почву для развития социального паразитизма, то есть для распространения особей, использующих чужой альтруизм (под которым в данном случае понимается отказ от агрессивной конкуренции) в своих корыстных интересах. Чтобы проверить, не может ли что-то подобное происходить у дрозофил, ученые провели еще одну серию экспериментов, в которой, кроме ситуаций AAA и ABC, была также ситуация AAB, когда два брата конкурировали с одним чужаком.
По количеству драк, продолжительности жизни самцов и репродуктивному успеху самок ситуация AAB оказалась промежуточной между ААА и ABC. Ученым, однако, не удалось заметить достоверных различий между поведением братьев и третьего самца. Братья дрались с чужаком не чаще, чем друг с другом. Похоже, самцы не различали своих сожителей индивидуально, а воспринимали лишь некий “общий уровень родства” в группе. Чем этот уровень был ниже, тем агрессивнее и назойливее они себя вели.
Все три самца в ситуации AAB спаривались с самкой с одинаковой частотой. Интересно, что при этом их репродуктивный успех очень сильно различался. Самцы B оказались (в среднем) отцами половины произведенного самкой потомства, тогда как на долю каждого из самцов А пришлось лишь по четверти. Причины такого успеха самцов B пока неизвестны. Так или иначе, опыт показал, что быть единственным чужаком в компании родственников может быть весьма выгодно.

