Книга: История биологии с древнейших времен до начала XX века
Назад: Глава 26 Создание и развитие эволюционной палеонтологии
Дальше: Глава 28 Перестройка сравнительной анатомии на основе дарвинизма
Глава 27
Создание эволюционной эмбриологии животных
Среди многих отраслей биологии, испытавших благотворное влияние учения Дарвина, одно из первых мест занимает эмбриология. В свою очередь открытия в области сравнительного изучения эмбрионального развития имели немаловажное значение для эволюционного учения, так как они дали убедительные доказательства кровного родства далеких по своей организации групп животных и способствовали построению родословной животного царства.
Сравнительное изучение эмбрионального развития.
Эмбриологи середины XIX в. продолжали в основном линию Бэра, ограничиваясь в своих работах сравнением зародышевого развития позвоночных, главным образом птиц и амфибии. Такие работы ни в какой мере не затрагивали теорию типов, прочно вошедшую в сознание зоологов. Отрывочные наблюдения, свидетельствовавшие о том, что у некоторых беспозвоночных, подобно позвоночным, зародыш состоит из отдельных слоев, привлекали к себе мало внимания и не могли вызвать, сомнений в справедливости теории типов Кювье-Бэра. Правда, еще в 1829 г. Г. Ратке попытался гомологизировать зародышевые слои речного рака с зародышевыми листками позвоночных. Такую же попытку позднее сделали А. Келликер (1842) и Г. Цаддах (1854) при изучении эмбриологии насекомых. Стремясь сопоставить эмбриональное развитие насекомых и позвоночных, Келликер дал слоям, на которые расщепляется зародышевая полоска на брюшной стороне бластодермы у насекомых, название серозного и слизистого листков. Эта схема перестала удовлетворять эмбриологов после того, как Ремак реформировал учение о зародышевых листках и стал говорить о них не только как о зачатках будущих частей сформировавшегося животного, но и как о функционирующих зародышевых органах.
Отголоском представлений Келликера в области эмбриологии насекомых явилась работа Г. Цаддаха. Он описал появление зародышевой полоски, расщепляющейся на два листка, которые, однако, по его мнению, соответствуют не серозному и слизистому листкам позвоночных как думал Келликер, а их роговому и мышечному листкам. Цаддах утверждал далее, что первичные сегменты насекомых соответствуют первичным позвонкам (сомитам) и что так называемые зародышевые лики являются не чем иным, как спинными пластинками, описанными Бэром у зародышей позвоночных. Вскоре выяснилось, что наружный листок, описанный Цаддахом у зародышей ручейника, является не зародышевым листком, а провизорной внезародышевой оболочкой. А. Вейсман, установивший эту ошибку, пришел к отрицанию существования зародышевых листков у насекомых. Оценивая позднее эту работу Вейсмана, И.И. Мечников писал: «Главный ее вывод сводился к тому, что насекомые развиваются по совершенно своеобразному типу и что немыслимо проводить какую бы то ни было параллель между эмбриологией этих суставчатоногих и развитием позвоночных… Выводы Вейсмана являлись, таким образом, новой опорой мнению, которое в те времена было общепринято, что каждый тип животных — позвоночные, мягкотелые, суставчатоногие и пр. — представляет особое, строго замкнутое целое и что поэтому нет никакой возможности проводить параллель между анатомическим устройством и историей развития представителей этих разных типов».
Мысль о зародышевых листках как основе для сопоставления способов развития животных, относящихся к различным типам, была столь чужда эмбриологам середины XIX в., что даже в тех случаях, когда отдельные листки были отчетливо видны у зародышей, они не связывали эти наблюдения с учением о зародышевых листках. Так обстояло дело и в тех случаях, когда А. Крон у медуз, и иглокожих или К. Гегенбаур у сагитты видели двуслойную стадию, позднее получившую название гаструлы.
Теория типов, сформулированная Кювье на сравнительно-анатомической основе и подтвержденная эмбриологическими исследованиями Бэра оставалась незыблемой вплоть до того времени, когда Дарвин обосновал идею единства происхождения всего животного мира. Дарвин очень рано, задолго до выхода в свет «Происхождения видов», осознал значение закономерностей эмбрионального развития для обоснования эволюционной теории.
В основных своих произведениях — «Происхождение видов», «Изменение домашних животных и культурных растений» и «Происхождение человека и половой отбор» — Дарвин целеустремленно и детально обсуждал проблемы эмбриологии и с большим вниманием следил за работами современных ему эмбриологов. Наибольшее значение он придавал тем, тогда еще немногочисленным исследованиям, которые демонстрировали единство закономерностей эмбрионального развития позвоночных и беспозвоночных, и прежде всего работам А.О. Ковалевского.
Создание А.О. Ковалевским и И.И. Мечниковым эволюционной эмбриологии.
К началу 60-х годов XIX в. эмбриология позвоночных была разработана достаточно детально. Что касается эмбриологии беспозвоночных, то в этой области было накоплено много разрозненных наблюдений, которые не удавалось еще связать общей руководящей идеей. Было описано, в частности, дробление яиц некоторых кишечнополостных, червей, моллюсков и иглокожих, строение и превращение личинок многих беспозвоночных, однако о внутренних процессах их развития, о способах закладки и дифференцирования органов у них почти ничего не было известно, а главное, не удавалось с достоверностью найти общие черты; в эмбриональных процессах у животных, относящихся к разным типам. Поэтому эволюционная эмбриология как наука, основывающаяся на историческом принципе, не могла еще возникнуть. Датой ее зарождении следует считать середину 60-х годов — начало эмбриологических исследований основоположников эволюционной сравнительной эмбриологии А.О. Ковалевского и И.И. Мечникова.

Александр Онуфриевич Ковалевский. 1840–1901.
К этому времени общие принципы теории эволюции, сформулированные Дарвином, уже нашли подтверждение в систематике и сравнительной анатомии, в работах некоторых палеонтологов, в данных географического распределения организмов, в практике разведения домашних животных и культурных растений. Общность происхождения в пределах таких хорошо очерченных групп животных, как позвоночные и членистоногие, не вызывала сомнений. Перед зоологами начала второй половины XIX в. стояла задача доказать родство между позвоночными и беспозвоночными и установить истинное систематическое положение, а следовательно, филогенетические отношения с остальным животным миром некоторых групп такого сборного типа, как черви, сомнительных в систематическом отношении форм — бесчерепных, оболочников, мшанок, плеченогих, щетинкочелюстных и т. д., а также тех классов животных, эмбриональное развитие которых, судя по их внешним особенностям, протекает совершенно отлично от позвоночных, в частности губок, кишечнополостных, щетинконогих кольчецов, боконервных, лопатоногих и головоногих моллюсков, ракообразных, паукообразных и насекомых.
Именно эти формы и привлекли к себе внимание Ковалевского и Мечникова. Они потратили 20 лет упорного труда на раскрытие закономерностей их эмбрионального развития. Плодом исследования этих ученых, за которыми пошли многие зоологи и эмбриологи, как в России, так и за ее пределами, явилось не только детальное знакомство с разнообразными формами эмбрионального развития у представителей разных систематических групп животных, но и принципиальные обобщения, совокупность которых по справедливости получила название эволюционной сравнительной эмбриологии.
Первая эмбриологическая работа А.О. Ковалевского, его магистерская диссертация, опубликованная в 1865 г., была посвящена развитию ланцетника. В то время ланцетника, несмотря на отсутствие у него позвоночника и головного мозга, дифференцированных органов чувств и парных плавников, относили к позвоночным. Сам Ковалевский называл его «замечательной рыбкой». Именно в эмбриональном развитии этого «позвоночного», строение которого убедительно говорило о его древности, Ковалевский рассчитывал обнаружить общие черты развития беспозвоночных и позвоночных. Это ожидание блестяще оправдалось, так как ранние стадии развития ланцетника оказались чрезвычайно сходными с соответствующими этапами онтогенеза многих беспозвоночных, в том числе и низших, например кишечнополостных.
Дробление яйца ланцетника, по наблюдениям Ковалевского, отличается от дробления яиц миноги и лягушки равномерностью, расположение же бластомеров оказалось менее упорядоченным, чем у позвоночных с полным дроблением, и напоминает дробление яиц низших беспозвоночных. Бластомеры зародыша ланцетника образуют полый шар, одна из половин которого затем впячивается в другую. Наружная поверхность полушаровидного зародыша (гаструлы) покрывается ресничками; на этой стадии происходит вылупление. Края отверстия, ведущего в гастральную полость, сближаются, и зародыш начинает вытягиваться в длину. «С этой стадии, — писал Ковалевский, — наша мешкообразная личинка, столь сходная с личинкой любой медузы, начинает уже развиваться дальше по типу позвоночного». В этом беглом замечании содержится глубокая и смелая мысль: Ковалевский утверждает, что низшее позвоночное (точнее, низшее хордовое) животное начинает развиваться по типу беспозвоночных, именно кишечнополостных, и лишь позднее приобретает признаки позвоночного. Первым из этих признаков является способ образования центральной нервной системы. Ковалевский показал, что эта стадия соответствует стадии образования медуллярной трубки у зародышей лягушки и миноги. Под нервной трубкой появляется хорда, состоящая из пузырчатых клеток, сходных с клетками хорды зародышей всех позвоночных. Описав далее образование ротового отверстия на переднем конце, противоположном месту закрывшегося отверстия гастральной полости, образование жаберных отверстий, хвостового плавника и пульсирующего брюшного кровеносного сосуда, Ковалевский сделал заключение, что развитие ланцетника может служить ключом к пониманию развития позвоночных.
Уже в этой ранней работе А.О. Ковалевский выступил как убежденный эволюционист-дарвинист. Он привел доказательства ошибочности антиэволюционных представлений Катрфажа, считавшего, что ланцетник — это деградированное позвоночное, а не форма, близкая к предкам позвоночных, и полемизировал с Келликером, полагавшим, что изменение видов зависит не от естественного отбора незначительных уклонений взрослых форм, а от внезапных превращений развивающихся зародышей.
Дальнейшие работы Ковалевского, посвященные развитию ланцетника (1867, 1870, 1876), уточнили и расширили его первоначальные наблюдения. Оценивая их, австрийский зоолог Б. Гатчек писал: «Мы можем считать эту работу началом новой эпохи в сравнительной эмбриологии, для которой она проложила путь».
Почти одновременно с первой работой о развитии ланцетника Ковалевский напечатал статью по эмбриологии асцидий (1866). Результаты этих исследований, как писал Ковалевский, превзошли его ожидания. Они привели его к выводам, которые первоначально казались совершенно парадоксальными. Ковалевский установил, что после дробления, образования шаровидной бластулы и инвагинационной гаструляции, сходной с описанной для ланцетника, наружный слой клеток образует два валика, которые через некоторое время смыкаются в нервную трубку, погружающуюся под слой покровных клеток. Тем самым, как это ясно сознавал Ковалевский, было установлено полное соответствие в образовании нервной системы у позвоночных и асцидий. Описав клеточный тяж, состоящий из одного ряда пузырчатых клеток и расположенный в хвосте личинки асцидии, Ковалевский смело приравнял его к хорде ланцетника и зародышей позвоночных и тем самым установил несомненное родство асцидий, которых в то время причисляли к моллюскам, с позвоночными. К заключению об их родстве на основании собственных исследований, подтвердивших наблюдения Ковалевского, вскоре присоединились немецкий зоолог К. Купфер (1870) и русский зоолог и эмбриолог М.С. Ганин (1870).
Годом позже в книге «Происхождение человека и половой отбор» Дарвин по достоинству оценил важность этих новых научных факторов и сделанных из них выводов. «Г-н Ковалевский, — писал он, — наблюдал недавно, что личинки асцидий сходны с позвоночными по способу развития, по относительному положению нервной системы и по присутствию одного органа, совершенно сходного с chorda dorsalis позвоночных животных, и в этом его данные были подтверждены проф. Купфером… Когда его результаты будут прочно подтверждены, то все это составит открытие величайшего значения. Таким образом, если верить эмбриологии, оказывавшейся всегда самой верной руководительницей в деле классификации, мы получим, наконец, ключ к источнику, из которого произошли позвоночные. Мы теперь имеем право думать, что в чрезвычайно отдаленный период времени существовала группа животных, сходных во многих отношениях с личинками теперешних асцидий, и что эта группа разделилась на две большие ветви, из которых одна регрессировала в развитии и образовала теперешний класс асцидий, другая же поднялась до венца и вершины животного царства, дав начало позвоночным».
Естественно, что открытия Ковалевского и вытекающие из них выводы не сразу встретили всеобщее признание. Более того, они сделались ареной борьбы новых, эволюционных, взглядов на мир животных и старых представлений, базирующихся на теории типов. Сторонники идеи эволюции органического мира, в частности Э. Геккель и К. Гегенбаур, увидели в факте общности эмбрионального развития позвоночных и беспозвоночных убедительное свидетельство филогенетическго родства между ними. Геккель рассказывал брату А.О. Ковалевского палеонтологу В.О. Ковалевскому, что Гегенбаур, прочтя работу о развитии асцидий, «проходил в волнении целую ночь, не ложась в постель».
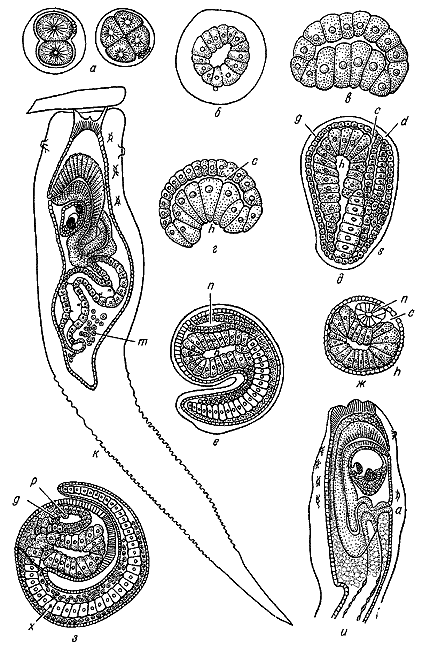
Развитие асцидий по А.О. Ковалевскому. 1866.
Противники эволюционных представлений и просто скептически настроенные естествоиспытатели попытались подвергнуть сомнению наблюдения и выводы Ковалевского.
Против сопоставления асцидий с позвоночными выступил К.М. Бэр, опубликовавший в «Записках Петербургской Академии наук» обширную полемическую статью на немецком языке под заглавием: «Развивается ли личинка простых асцидий первое время по типу позвоночных животных?» (1873). Возражения Бэра основывались не на проверке эмбриологических открытий, а на сравнительно-анатомических соображениях. По положению сифонов, нервных узлов и другим анатомическим признакам Бэр стремился подтвердить ошибочное заключение Кювье о систематической близости асцидий к двустворчатым моллюскам. Движущей силой полемического выступления Бэра было его отрицательное отношение к дарвинизму.
С утверждением Ковалевского о родстве между оболочниками и позвоночными не сразу согласился даже Мечников, посвятивший полемике с ним несколько статей. Опираясь на господствовавшие тогда неверные мнения о принадлежности асцидий к типу моллюсков и на представление, что нервная система моллюсков в отличие от других животных происходит не из верхнего, а из среднего зародышевого листка, Мечников пришел к заключению, что «самое первое образование органов у асцидий ни в коем случае не напоминает собой типичного развития позвоночных, как это утверждал Ковалевский». Мечников считал также, что содержимое хордальной оболочки асцидий, т. е. вещество самой хорды, жидкое и вовсе не состоит из клеток; тем самым, по его мнению, осевой тяж личинок асцидий не может сравниваться с хордой позвоночных. Парируя эти возражения, Ковалевский писал: «Сейчас едва ли кто будет сомневаться в том, что хорда асцидий не только аналогична, но и гомологична хорде позвоночных».
Мечников лишь постепенно, шаг за шагом, уступал свои прежние позиции. Уже в статье 1871 г. Ковалевский ссылается на письмо Мечникова, в котором последний выражает согласие с его основными соображениями о развитии нервной системы и хорды у асцидий. Правильность наблюдений и выводов Ковалевского в дальнейшем подтвердили бельгийские эмбриологи П. ван Бенеден и Ш. Жюлен (1884) и другие авторы.
Вспоминая о своих прежних разногласиях с Ковалевским, Мечников в статье, посвященной памяти своего друга (1902), писал: «Мне сначала казалось, что наблюденные мною факты не вяжутся с выводами Ковалевского. Но лотом я сам и многие другие естествоиспытатели вполне подтвердили точность данных, добытых А.О.».
Считая сравнительную эмбриологию источником доказательств единого происхождения всех многоклеточных животных, Ковалевский в докторской диссертации, посвященной развитию загадочной в систематическом отношении группы форонид (1867), наметил ту основную задачу, решение которой позволит, по его мнению, заложить основы этой новой науки. Он полагал, что для всех животных должен быть прежде всего выяснен способ развития главных органов пищеварительного канала, стенок полости тела и нервной системы, т. е. тех образований, которые у позвоночных возникают соответственно из нижнего, среднего и верхнего зародышевых листков. Он с большой проницательностью показал, что основой сравнительной эволюционной эмбриологии является учение о гомологии зародышевых листков у всех типов животных.
Подтверждение гомологии зародышевых листков позвоночных и беспозвоночных.
Пандер и Бэр в первой трети XIX в. создали топографическую теорию зародышевых листков, показав, что взаимное расположение наружного, среднего и внутреннего листков у позвоночных в принципе одинаково и что из каждого такого листка у всех классов позвоночных развиваются одни и те же системы органов. В 60-80-х годах А.О. Ковалевский и И.И. Мечников впервые высказали мысль, что листкам, или клеточным слоям, из которых состоят зародыши позвоночных, вполне гомологичны соответствующие образования зародышей самых различных типов беспозвоночных. Тем самым они вложили в теорию зародышевых листков генеалогическое, или филогенетическое, содержание, обосновав эмбриологическими данными учение Дарвина о кровном родстве всего животного царства.
Ведущая роль А.О. Ковалевского в создании генеалогической теории зародышевых листков получила признание не сразу. Э. Геккель, выдвинувший под названием теории гастреи гипотезу происхождения многоклеточных животных от общего предка, имевшего, подобно эмбриональной стадии — гаструле, — структуру двуслойного мешка, писал, что «теория типов Кювье и Бэра стала неприемлемой благодаря успехам онтогении. На ее месте теория гастреи воздвигает на основе филогении новую систему, высшим классификационным признаком которой является гомология зародышевых листков». На самом деле теорию типов сменила не теория гастреи Геккеля, а теория эволюции Дарвина, в которой была использована идея гомологии зародышевых листков, т. е. обоснованное Ковалевским и Мечниковым эмбриологическое доказательство единства происхождения всех многоклеточных животных.
Проблема гомологии зародышевых листков, возникающая при сопоставлении развития позвоночных и беспозвоночных, с наибольшей отчетливостью поставлена в классическом труде Ковалевского «Эмбриологические исследования червей и членистоногих» (1871). Ковалевский писал, что он ставил своей главной задачей исследовать спорный в то время вопрос о зародышевых листках беспозвоночных. В этом замечании нет никакого преувеличения. Как уже отмечалось, идея существования зародышевых листков у беспозвоночных до начала 70-х годов считалась малоправдоподобной. Многие эмбриологи (А. Вейсман, М.С. Ганин, Н.М. Мельников, П.Т. Степанов и др.) были уверены, что, например, при описании эмбрионального развития членистоногих и моллюсков нет оснований приравнивать клеточные слои их зародышей зародышевым листкам позвоночных. Даже И.И. Мечников, убедившись после своих исследований эмбрионального развития ракообразных, паукообразных (скорпионы) и головоногих моллюсков (каракатицы) в существовании у них, по крайней мере, двух зародышевых листков, смог привести в пользу этого заключения лишь свидетельства, основанные на прижизненных наблюдениях непрозрачных зародышей. В сущности, дело шло не о точных доказательствах, а скорее о догадках.
Учение о зародышевых листках насекомых (и кольчатых червей) встало на твердую почву только после упомянутого исследования Ковалевского, впервые применившего в эмбриологии беспозвоночных метод окрашенных микротомных срезов.
При изучении развития водных и наземных малощетинковых кольчецов Ковалевский не только установил наличие у них трех зародышевых листков, из которых, как и у позвоночных, наружный дает начало покровам и нервной системе, средний — мышцам и органам выделения, а нижний — пищеварительной трубке, но и показал возникновение каждого из этих листков из определенных бластомеров.
Э. Ланкестер (1873) также считал несомненным, что соответствие зародышевых листков у животных, относящихся к различным систематическим группам, является выражением общности их происхождения. Он попытался даже положить в основу деления всего животного царства на грунты (Animalia homoblastica, diploblastica и triploblastica) отсутствие или наличие и число (два или три) зародышевых листков.
Большая роль в обобщении данных сравнительной эмбриологии и их использовании для обоснования теории эволюции принадлежит Э. Геккелю. Его теория гастреи, т. е. гаструлоподобного гипотетического предка всех многоклеточных животных (1874), была создана главным образом на базе фактического материала, собранного Ковалевским. Одной из составных частей этой теории является учение о гомологии зародышевых листков. Для первичных, по терминологии Геккеля, «зародышевых листков» эктодермы и энтодермы (эти названия были предложены еще в 1853 г. Олменом для обозначения слоев тела кишечнополостных, а затем стали применяться к зародышевым листкам) Геккель допускал полную «в самом строгом смысле» гомологию. Это положение неоднократно оспаривалось с разных точек зрения. Против каждого из трех критериев сходства стадии гаструлы у разных животных — одинакового способа образования гаструлы, одинакового ее строения и одинаковой судьбы в развитии ее слоев, т. е. зародышевых листков, — также выдвигались в разной степени доказательные возражения. Работы Ковалевского, Мечникова и других эмбриологов показали, что двуслойный зародыш — гаструла — может возникать разными путями — посредством впячивания (инвагинации), обрастания (эпиболии), уни- и мультиполярного вселения (иммиграции) клеток и расслаивания (деламинации).
В упомянутой работе по эмбриологии червей и членистоногих Ковалевский впервые высказал мысль, что инвагинационная и эпиболическая гаструляции представляют видоизменения одного итого же процесса. С присущей ему осторожностью Ковалевский не считал возможным заранее решать, какой способ гаструляции является первичным, т. е. филогенетически более древним. Геккель, склонный к схематизации в обобщениях, с уверенностью говорил о первичности инватинационного способа обособлений зародышевых листков; из этого допущения и возникло представление о гастрее, как древней прародительской форме.
Строение готовой гаструлы разных животных тоже может быть различным, хотя, исходя из этих различий, особенно существенных возражений против гомологии зародышевых листков сделано не было. Многие примеры, приводившиеся для опровержения генеалогической теории зародышевых листков и якобы говорившие о том, что одни и те же органы у разных животных образуются не из одного и того же зародышевого листка, оказались неверными. Сюда относятся, например, ошибочные утверждения, что нервная система, возникающая у большинства животных из эктодермы, у некоторых групп (моллюсков и трехветвистокишечных турбеллярий и др.) будто бы образуется из мезодермы.
Различия в характере развития при образовании дочерней особи из яйца и из почки, а также при первичном возникновении органа в онтогенезе и вторичном развитии в результате регенерации нашли рациональное объяснение, не колеблющее основ гомологии зародышевых листков. Л. Шульце в статье «Регенерация ганглия у Ciona intestinalis и об отношении регенерации и почкования к учению о зародышевых листках» резюмировал изложенные в ней соображения следующим образом: «Учение о гомологии зародышевых листков, основанием которого является положение, что оба клеточных слоя, характеризуемые своим положением на двулистной стадии зародыша, обладают характером морфологически одинаковых органов во всех группах Metazoa, никоим образом не затрагивается явлениями почкования и регенерации».
К числу наиболее известных примеров, вызвавших оживленное обсуждение проблемы гомологии зародышевых листков, относится развитие средней кишки у насекомых. А.О. Ковалевский (1871) сначала с удивлением отмечал, что из нижнего зародышевого листка исследованных им насекомых образуется только мезодерма, дающая, как он думал, начало и эпителию средней кишки. В более поздней работе, посвященной специально эмбриологии мух (1886), он описал образование энтодермы из двух клеточных скоплений на концах ротового и заднепроходного эктодермальных впячиваний; иначе говоря, средняя кишка насекомых оказалась вполне гомологичной энтодермальной средней кишке остальных животных. Позднее (1895) Р. Геймонс пришел к заключению, что тело сформированного насекомого состоит только из эктодермы и мезодермы, так как, по его мнению, средняя кишка развивается из переднего и заднего эктодермальных впячиваний, а энтодерма в образовании сформированного тела насекомого участия не принимает.
Может сложиться впечатление, что материал, из которого образуется средняя кишка у насекомых и энтодерма позвоночных, совершенно различен и, стало быть, этот факт полностью противоречит идее гомологии зародышевых листков позвоночных и членистоногих. На самом деле, как это убедительно показал П.Г. Светлов, соотношение в развитии кишечника позвоночных и насекомых является примером неполной гомологии. Несмотря на различие источника образования, нижний зародышевый листок, источник развития выстилки кишечника, должен быть признан гомологичным у насекомых и у тех животных, у которых эта выстилка происходит из энтодермы.
Проблема происхождения многоклеточных.
Сравнительно-эмбриологические исследования 60-80-х годов XIX в. сделали возможным обсуждение вопроса о происхождении многоклеточных животных, который имеет непосредственное отношение к проблеме эволюции животного царства. Наиболее детальные представления о происхождении многоклеточности развили Э. Геккель и И.И. Мечников. Оба они опирались на данные сравнительной эмбриологии. Геккелю принадлежит упомянутая выше теория гастреи, согласно которой у размножавшихся посредством деления простейших предков многоклеточных животных на определенном этапе эволюции дочерние особи перестали расходиться и образовали скопление, превратившееся затем в полый шар, напоминающий колонию жгутиконосцев типа Volvox. Одна из стенок этого клеточного шара впятилась, и появилось двуслойное образование, сходное с инвагинационной гаструлой, характерной для многих беспозвоночных и хордовых. Имеющему такое строение гипотетическому предку ныне живущих многоклеточных Геккель дал название гастреи.
Теория гастреи вскоре после ее опубликования получила широкую известность и возбудила оживленные дискуссии. Одно из наиболее обоснованных возражений против теории гастреи выдвинул Мечников. В монографии «Эмбриологические исследования над медузами» (1886) он в противовес геккелевской теории гастреи предложил теорию паренхимеллы, или фагоцителлы. Мечников решал вопрос о первичности одних форм гаструляции по сравнению с другими, исходя из способов питания низших многоклеточных. Открыв явления внутриклеточного пищеварения у губок и кишечнополостных, Мечников пришел к заключению, что такая форма восприятия и усвоения пищи должна была характеризовать древнейших предков современных многоклеточных. Эту мысль подтверждал тот факт, что у низших многоклеточных гаструляция совершается чаще всего путем вселения (однополюсного или многополюсного) части клеток бластулы внутрь шарообразного зародыша или путем его расслоения (деламинации).
Это обстоятельство поставило под сомнение предположение Геккеля, что гипотетический предок всех современных дву- и трехслойных животных имел близкое сходство с инвагинационной гаструлой, поскольку оно требовало допущения, что первичной, наиболее древней формой обособления зародышевых листков является гаструляция посредством впячивания и что все другие способы этого обособления представляют позднейшие видоизменения инвагинационной гаструляции.

Илья Ильич Мечников. 1845–1916.
Наблюдения Мечникова привели его к выводу, что предком многоклеточных было колониальное простейшее, часть клеток которого оставалась на поверхности, образуя эктодерму, или кинобласт, а остальные, оказавшиеся внутри, образовывали энтодерму, или фагоцитобласт. Инвагинационная гаструляция, которую Геккель считал первичной, возникла, по мнению Мечникова, в результате достаточно длительного процесса эволюции.
В целом идея гомологии зародышевых листков явилась важной составной частью учения об эволюции животного мира, но она вовсе не требовала признания первичности инвагинационного способа обособления зародышевых листков.
Проблема соотношения онтогенеза и филогенеза.
Успехи эволюционной эмбриологии заложили основу для нового подхода к проблеме соотношения онтогенеза и филогенеза. Эмбриологические работы Ковалевского позволили перебросить мост между беспозвоночными и позвоночными, принеся фактическое подтверждение закона рекапитуляции Дарвина. Исходя из этого закона, Ф. Мюллер предпринял попытку воссоздать филогению класса ракообразных, а Э. Геккель создал первое филогенетическое древо всего животного мира.
Параллельно с решением филогенетических проблем Мюллер и Геккель затронули вопрос о соотношении онтогенеза и филогенеза. Однако подошли они к этому вопросу по-разному. Мюллер исходил из положения Дарвина об изменяемости в процессе эволюции любой стадии индивидуального развития. В труде «За Дарвина» Мюллер отметил, что новые признаки формируются или путем видоизменения какого-либо отрезка онтогенеза, или путем прибавления (надставки) новой стадии к прежнему, неизменному онтогенезу. В последнем случае «историческое развитие вида будет отражаться в истории его индивидуального развития». Мюллер связал процесс преобразования личиночных стадий (так же как и процесс эволюционного преобразования онтогенеза в целом) с действием естественного отбора и борьбой за существование. Он сделал попытку выяснить факторы, обусловливающие полноту рекапитуляции. Представления Мюллера о механизме рекапитуляции не сводились к надставкам стадий; Мюллер подчеркнул значение сходства образа жизни индивида на разных возрастных ступенях; он отметил отсутствие сдвигания поздних стадий, влекущего за собой преобразование предшествующих этапов онтогенеза и приспособительного видоизменения ранних стадий. Он показал также, что темп эволюции и связанная с ним продолжительность существования данной систематической формы являются факторами, определяющими характер повторяемости в онтогенезе признаков предков.
Важным новым моментом, внесенным Мюллером в понимание соотношения онтогенеза и филогенеза, явился его вывод о многообразии типов эмбриональной дивергенции. Помимо случая, когда первоначально сходные зародыши постепенно, в ходе индивидуального развития приобретают все больше черт различия, Мюллер обнаружил, что подчас зародыши родственных форм более всего отличаются на ранних этапах онтогенеза, в других же случаях расхождение признаков падает на середину развития; наконец, сходство может наблюдаться на средних этапах индивидуального развития, в то время как на ранних стадиях и во взрослом состоянии организмы могут быть различны.
Возрастающий интерес к филогенетической проблематике повысил в глазах исследователей значение закона рекапитуляции как метода филогенетического анализа. Этому особенно содействовал Геккель, который пришел к заключению, что ведущее значение для выяснения родственных отношений между организмами имеют эмбриологические данные, ибо развитие особи есть воспроизведение истории вида. Абсолютизировав закон рекапитуляции, открытый Дарвином, Геккель дал свою формулировку биогенетического закона, который гласит: «Онтогенез является кратким и быстрым повторением филогенеза…».
Биогенетический закон в качестве важнейшей составной части метода тройного параллелизма широко использовался им в работе по созданию всеобъемлющего родословного древа, при решении проблем антропогенеза и разработке теории гастреи. Он стал ядром геккелевской теории рекапитуляции. Согласно этой теории, онтогенез включает два типа признаков: палингенезы — признаки филогенетически далеких предков и ценогенезы — вторичные признаки, возникшие в результате приспособления к определенным условиям эмбриональной или личиночной жизни. Унаследованным от далеких предков палингенетическим признакам Геккель приписывал первостепенную роль при выявлении филогенетических отношений. Ценогенез — собирательное понятие, охватывающее совокупность процессов, затемняющих проявление палингенетических признаков. Ценогенезы возникают благодаря гетерохронии — нарушению последовательности филогенетических стадий во времени и гетеротопии нарушению их последовательности в пространственном отношении.

Фриц Мюллер. 1821–1897.
Биогенетический закон верно схватил и отразил одну из сторон явления соотношения индивидуального и исторического развития — именно: момент повторяемости. Геккель в большой мере способствовал распространению биогенетического закона в качестве средства, помогающего познавать филогению; это стимулировало интенсивную исследовательскую работу в эмбриологии, сравнительной анатомии, палеонтологии и в особенности в области филогенетики. Однако Геккель упростил трактовку явления взаимосвязи онтогенеза и филогенеза. По существу в его формулировке биогенетический закон отражал одностороннюю связь между развитием особи и эволюцией вида, ибо Геккель ошибочно полагал, что филогенез является механической причиной онтогенеза и что существует полный параллелизм этих двух процессов. Наряду с другими причинами, недостаточность геккелевской трактовки биогенетического закона, вытекавшая из механистического ограниченного представления его автора о характере взаимоотношений онтогенеза и филогенеза, сделала этот закон предметом острой и продолжительной дискуссии. В ней нашло отражение углублявшееся противоречие между объективной диалектикой природы и метафизическим мышлением естествоиспытателей, которое обострило борьбу между идеализмом и материализмом в биологии.
Морфологи-идеалисты, виталисты, биологи, разделявшие автогенетические представления, приложили немало усилий, чтобы дискредитировать биогенетический закон, в котором они справедливо усматривали надежную опору эволюционной теории. Они отрицали связь между онтогенезом и филогенезом вовсе или истолковывали ее в духе механицизма, идеалистической морфологии и витализма. Предпринимались попытки отыскать противоречие между биогенетическим законом и теорией естественного отбора, противопоставить этот закон материалистическому решению проблемы целесообразности. Так, В. Гис (1870) утверждал, что изучение генеалогических отношений на основании эмбриологии должно быть навсегда прекращено, поскольку различные направления развития могут быть исчерпывающе объяснены через математически точный круг существующих способов роста. По представлениям А. Келликера (1864), изменение типичного хода индивидуального развития и появление вследствие этого новых форм обусловливается неким общим имманентным законом развития. В соответствии с ним он допускал, что без участия естественного отбора путем спонтанных скачков из зародыша губки может возникнуть гидроидный полип, из зародыша медузы — иглокожее, из зародыша сумчатого — грызун и т. п.
Г. Вольф, предпринявший попытку экспериментально доказать несостоятельность дарвиновской концепции развития, усматривал в материалистической трактовке соотношения онтогенеза и филогенеза неразрушимое противоречие. Последнее сводится им к следующему. В развитии особи наблюдается известная «целестремительность»; процессы онтогенеза ведут к вполне определенному результату. Между тем в отношении филогенеза дарвинисты подобную «целестремительность» отрицают. Следовательно, происхождение целесообразности онтогенетических процессов остается дарвинистами необъясненным. С виталистических позиций вопрос этот получает у Вольфа следующее решение. Новое в эволюционном развитии впервые проявляется в онтогенезе; однако эти новые явления не следует относить к индивидуальному развитию, между ними и процессами онтогенеза, определяемыми наследственностью, существует коренное отличие.
Таким образом, период онтогенеза, в котором обнаруживается это первично целесообразное, «чудом» появляющееся новое, не имеет, по Вольфу, никакого отношения к филогенезу.
Критическому обсуждению подвергли биогенетический закон и дарвинисты. Придавая принципу рекапитуляции важное значение, они стремились освободись закон от ошибочных наслоений и привести в соответствие с новыми фактами из области эмбриологии и сравнительной анатомии.
Так, А. Оппель (1891), проведя детальное сравнение различных органов у ряда позвоночных, находящихся на сходных стадиях эмбрионального развития, заключил, что онтогенез есть повторение филогенеза с поправкой на величину «онтогенетической разницы», т. е. отличия зародышевого развития потомков сравнительно с таковым у предков. Изменение эмбрионального развития обусловливается, по Оппелю, гетерохронией и субституцией. Обстоятельное изучение эмбрионального развития свиньи заставило Ф. Кейбеля (1897, 1898) усомниться в том, что в онтогенезе возможно воспроизведение всех филогенетических стадий. Согласно Кейбелю, гетерохронии, ведущие к сдвиганию стадий во времени и к изменению эмбриональных органов, стирают следы филогенеза.
Принимая во внимание явление гетерохронии, Э. Менерт (1898) внес поправку в формулировку биогенетического закона: онтогенез есть измененное повторение филогенеза. Ранее к сходному выводу пришел В.В. Заленский (1884), подчеркнувший, что организмы на всех этапах зародышевого развития, начиная с яйца, находятся под влиянием окружающих условий и вынуждены вести борьбу за существование. Заленский обратил внимание на неразработанность критериев разделения палингенезов и ценогенезов. Наличие ценогенезов дало К. Гегенбауру основание возражать против абсолютизации эмбриологического метода в познании филогении. Сам Гегенбаур отдавал здесь предпочтение сравнительно-анатомическому методу.
Дискуссия, вызванная биогенетическим законом, выявила необходимость критического отношения к нему, а накапливающиеся факты убедительно свидетельствовали о том, что онтогенез никогда не воспроизводит филогенез полностью, что течение онтогенеза в процессе эволюции видоизменяется под воздействием гетерохронии, гетеротопии, эмбриональных приспособлений, редукции и т. п. В результате представление Геккеля о том, что онтогенез есть краткое и быстрое повторение филогенеза, было оставлено. Однако ядро биогенетического закона — принцип рекапитуляции — выдержало все испытания. В конце XIX в. названный принцип нашел также применение в филогенетических исследованиях в области морфологии растений, гистологии, физиологии и биохимии.
Назад: Глава 26 Создание и развитие эволюционной палеонтологии
Дальше: Глава 28 Перестройка сравнительной анатомии на основе дарвинизма

