Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 34 Мезозойская эскалация
Дальше: Глава 36 Зеленая эпитафия динозаврам
Глава 35
Соревнование двух систем
Вне морской среды в мезозойскую и кайнозойскую эры происходили не менее драматичные изменения. Потепление при наличии обширной суши привело к тому, что триасовый период стал самым сухим в истории Земли. В сухости, при сильных перепадах температур вольготнее всего себя почувствовали семенные папоротники (кейтониевые, пельтаспермовые) и голосеменные. Кейтониевые (Caytoniales, название которых происходит от бухты Кейтон в Англии, на берегу которой собраны их многочисленные остатки) отличались пальчатыми листьями в мелкую сеточку и разнообразными по форме коробочками семян, свисавшими с веток. У пельтаспермовых (Peltaspermales, от греч. πέλτη – щит и σπείρον – покров) листья были перистые, часто сросшиеся, как щиток. Широко распространились леса, где было много деревьев с игольчатыми листьями (не только хвойных), которые прекрасно удерживали влагу. Уже существовали деревья, похожие на сосны и араукарии.
Господство голосеменных продолжилось в юрском периоде и в раннемеловую эпоху. Более разнообразными стали гинкговые, саговники и беннеттиты; появилось много сугубо мезозойских групп, таких как чекановскиевые и пентоксилеевые. Последние (Pentoxyales, от греч. πέντε – пять и ξύλον – древесина) получили название по пятичастному срезу ствола, где каждый сегмент был обрисован своими годичными кольцами. Листочки чекановскиевых (Czekanowskiales, названы в честь ссыльного польского геолога Александра Чекановского, в чьих сибирских коллекциях они были описаны) напоминали хвоинки, многократно раздваивавшиеся в одной плоскости. Саговники и их близкие родственники – беннеттиты (Bennettitales, названные в честь английского ботаника Джона Беннетта) – образовывали подлесок. У одних беннеттитов стебли были тонкие, ветвящиеся, у других – толстые, похожие на бочонок или несколько сросшихся ананасов; листья длинные, перистые, как у саговников, но семена сидели не в шишках, а в пальчатых или чашевидных органах, напоминавших цветы. Эти «цветы» опыляли насекомые, а мясистые плоды поедали крупные животные и таким образом разносили семена. Среди саговников были и деревья, и растущие на ветвях и коре других видов растения-паразиты. После лесных пожаров саговники первыми выпускали зеленые ростки, подкармливая уцелевших животных. Открытые пространства заполняли травянистые папоротники. На влажных склонах и в низинах, вблизи озер и устьев рек последние древовидные хвощи-неокаламиты все еще тянули свои кроны на 10 м вверх.
В триасовом периоде растительность по всей Пангее не отличалась многообразием, но с середины мезозойской эры, по мере установления климатических поясов с разными температурами и количеством осадков, а также по мере раскола последнего (на сегодня) суперконтинента стали формироваться растительные царства. На севере Пангеи, в бывшей Ангариде с ее сухим и прохладным в зимнее время климатом, преобладали листопадные хвойные, особенно метасеквойи, и гинкговые. Единственные существующие ныне виды метасеквойи (Metasequoia glyptostroboides) и гинкго (Ginkgo biloba) выжили только на небольших горных территориях Китая (причем гинк-го – только в монастырских парках), а метасеквойя впервые была описана как ископаемое растение и лишь потом открыта в природе (рис. 35.1). Двулопастные, похожие на маленькие зеленые веера листочки гинкго невозможно спутать ни с какими другими, даже в ископаемом виде. На ветвях мезозойских гинкговых шелестели разные листочки – похожие на хвоинки, на сердечки, на перышки, но всегда раздвоенные. И насекомые – скорпионницы – научились скрываться среди них: их четыре раскрытых крыла повторяли очертания листовой пластинки гинкго, пронизанной жилками.
Некоторые южные гондванские древовидные папоротники-диксонии и причудливые голосеменные той поры сохранились в Тасмании, Новой Зеландии и Новой Каледонии. Хвоя у этих голосеменных длинная и плоская, на ветках торчат яркие семена на ножках, давшие им название – ногоплодники. Есть среди них деревья по 80 м высотой, похожие на елку, есть и стелющиеся ниже травы кустарники. Уцелели в Южном полушарии и араукарии. В мезозойскую эру компанию им составляли кейтониевые и пентоксилиевые, а в экваториальном поясе, охватывавшем большую часть будущих Америк, Европы, Африки и Восточной Азии, произрастали теплолюбивые, с малым количеством вторичной древесины, беннеттиты, саговники и папоротники.
Упавшие стволы голосеменных не особенно разрушались, поскольку эти растения прекрасно овладели средствами химзащиты. Неслучайно из ливанского кедра строили свои суда искусные мореплаватели древности – финикийцы, из него же выдалбливали «вечные» саркофаги для египетских фараонов, про обеззараживающие свойства гинкго ходят легенды, а у тисса в хвое накапливается яд, в больших дозах смертельный даже для млекопитающих. (Не исключено, что поэтому по всей планете сохранились окаменевшие мезозойские «леса», в основном, конечно, пни и фрагменты стволов.)


Вероятно, насекомые поначалу сосредоточились на растительных соках, пыльце и семенах: встречаются обильные копролиты с пыльцой саговников и других голосеменных, а в стробилах и шишках заметны повреждения, нанесенные личинками древних перепончатокрылых (пилильщиков) и жуков-долгоносиков. Клопы, цикады, тли и некоторые другие группы с сосущими ротовыми аппаратами поглощали соки растений. Пыльцой питались сетчатокрылые, скорпионницы, ручейники, кузнечики, пилильщики и многие другие. Позднее личинки двукрылых вслед за тараканами, жуками и скорпионницами приспособились поедать тронутую грибным разложением древесину, а личинки цикад – высасывать корни. В позднемеловую эпоху насекомые освоили листовую мякоть покрытосеменных. Среди их следов распознаются ходы гусениц моли-малютки (самых мелких и притом весьма прожорливых бабочек) – мины, проложенные в листовых пластинках. Возможно, насекомым больше «пришлись по вкусу» листья цветковых, потому что они богаче азотом, необходимым для роста и обновления хитинового панциря, и переход на этот корм вызвал бурный рост разнообразия листогрызущих групп. Тогда же личинки перепончатокрылых и одетые в прочный панцирь жуки-златки вгрызлись в стволы ослабленных деревьев. Съесть все им не дали паразитические насекомые, такие как наездники-ихневмониды, веерокрылки, мухи-жужжалы, чьи личинки высасывают чужое потомство, и хищники: на своих собратьев нападали сетчатокрылые (мантиспиды с хватательной передней парой конечностей, как у богомолов); сверчки; очень похожие на богомолов (но совсем не родственные) вымершие мезотитаниды; тараканы с хватательными конечностями; жуки и позднее – настоящие богомолы.
На фоне развития наземной растительности и благодаря появлению все более продуктивных кормов (в листьях гинкго содержится больше белка и меньше плохо усваиваемых растительных волокон, чем у травянистых и древовидных папоротников, а у покрытосеменных – еще больше) совершенствовались наземные растительноядные позвоночные. Продолжилось соревнование, по большей части заочное, двух основных линий наземных позвоночных – синапсидной и диапсидной. В мезозойскую эру вперед вырвались диапсиды – архозавры, которые, встав на задние ноги и распрямившись, более оперативно решили основную проблему пресмыкающихся – быстро перемещаться и дышать одновременно. (При четвероногой ходьбе из-за «растопыренного» положения плеч и бедер пресмыкающиеся переваливаются с боку на бок и поэтому то правое, то левое легкое пережимается.) Синапсиды смогли окончательно решить эту задачу только на стадии плацентарных млекопитающих в меловом периоде. К тому же диапсиды оказались лучше приспособлены и к пониженному содержанию кислорода в атмосфере, и к триасовой сухости (конечным продуктом обмена веществ у них является мочевая кислота, для выведения которой из организма требуется в 10 раз меньше воды, чем для выведения мочевины). Поскольку архозавры заняли области с теплым климатом, синапсиды были вытеснены в высокие широты. Возможно, именно это обстоятельство сказалось на развитии у синапсид гомойотермии (постоянной температуры всего тела, превышающей таковую окружающей среды за счет тепла, вырабатываемого органами даже в состоянии покоя; от греч. ομοιόω – равнять и θερμός – теплый). Признаками «теплокровности» у ископаемых синапсид, выраженными в строении скелета, являются окостеневшие носовые раковины, окологлазничные костные выступы, пронизанная густой сетью кровеносных сосудов пластинчатая костная ткань и редукция теменного отверстия. Вместительные носовые раковины синапсид были при жизни покрыты изнутри слизью, препятствовавшей потере влаги и тепла. Костные «бордюры» вокруг глаз не давали органам зрения «утонуть» в шерсти (подобные скелетные образования появятся в меловом периоде у некрупных хищных динозавров, чтобы обзору не мешали перья). Пластинчатая кость свидетельствует о высоких темпах обмена веществ (правда, этот признак имеет значение только в сочетании с остальными: у некоторых быстрорастущих рептилий такая кость развивается, а у воробьиных птиц и мелких млекопитающих – нет). А редукция теменного отверстия, где располагался пинеальный орган, или «третий глаз», улавливавший солнечный свет, указывает на уменьшение зависимости терморегуляции от внешнего источника тепла. Еще одним веским доказательством в пользу гомойотермии у звероподобных синапсид служит размер красных кровяных клеток. Чем они меньше, тем лучше захватывают и переносят кислород, необходимый для эффективной терморегуляции, а у этих зверей красные кровяные клетки были очень мелкие. (Конечно, сами они не сохраняются, но их размер можно рассчитать, сравнивая диаметр и плотность гаверсовых каналов и остеоцитовых полостей в компактной костной ткани: у современных млекопитающих диаметр этих образований заметно меньше, а плотность – выше, чем у земноводных и пресмыкающихся.) Исходя из расчетной размерности красных кровяных телец получается, что темпы обмена веществ в состоянии покоя у предков и древних родственников млекопитающих были не ниже, чем у морских свинок (2,58–4,58 мл О2 в час на 1–0,63 г массы; коэффициент вводится для поправки на массу, поскольку у животных разного размера темпы метаболизма отличаются). Вывод о гомойотермности этих зверей напрашивается и при анализе изотопной подписи кислорода, зависимой от температуры, при которой фосфатные минералы формировали зубы и другие части скелета наиболее продвинутых зверообразных ящеров: у них совершенствовалась гомойотермия, причем независимо в нескольких триасовых линиях предков и ближайших родственников млекопитающих. В кайнозойскую эру в крупноразмерном классе вперед вновь вырвались синапсиды – потомки звероящеров – млекопитающие, а среди мелких наземных позвоночных еще в позднемеловую эпоху установился паритет. (Конечно, если сложить вместе всех небольших черепах, ящериц, змей и птиц, то по числу видов диапсиды по-прежнему окажутся впереди, но по биомассе грызуны им не уступают.)
Диапсиды своей вершины развития достигли в юрском и меловом периоде с появлением гигантских растительноядных динозавров – рогатых (Ceratopsia, от греч. κέρας – рог и οψις – вид, наружность), птиценогих (Ornithopoda, от греч. ορνις – птица и πούς, ποδός – нога) и панцирных ящеров (Ankylosauria, от лат. ancile – щит и греч. σαύρα – ящерица) и, конечно, завропод (Sauropoda, от греч. σαύρα – ящерица и πούς, ποδός – нога), а также огромных хищников – тираннозаврид (Tyrannosauridae) и разнообразной мелкой, но очень активной плотоядной мелочи – манирапторов (Maniraptora, от лат. manus – рука, передняя лапа, и raptor – хищник). Динозавров можно назвать «позвоночным хай-теком» – в строении их тела сочетаются, казалось бы, несочетаемые признаки: небывалая для наземных животных масса при отсутствии существенно укрепленных конечностей и довольно сложное, судя по следовым дорожкам и гнездам, поведение при небольшом объеме мозга.
Особенно выделялись завроподы (рис. 35.2). Расчеты их массы, сделанные по замерам следов, а также по обмерам наиболее полных скелетов и костей (в последние годы – с помощью 3D-фотограмметрии на основе лазерного сканирования), показывают, что это были самые крупные и тяжелые создания из всех, когда-либо ступавших по суше: от 15 000 кг до 75 000 кг при длине до 26–27 м. Возможно, они были и больше, но размеры более крупных особей рассчитываются по отдельным костям методом масштабирования, который менее точен. Сравниться с завроподами может лишь синий кит – 190 000 кг (33 м), но он живет в водной среде, где легче бороться с силой притяжения и главное – с перегревом. Одно время думали, что завроподы, будучи также самыми длинношеими (до 15 м) и длиннохвостыми (до 17 м) существами, вели водный или полуводный образ жизни: ходили, волоча хвост, по дну лагун и озер и, опустив шею, выуживали со дна пучки водорослей. (Правда, следы завроподы по большей части оставили на суше, отпечатки хвоста полностью отсутствуют, а в изотопном составе их костей нет и намека на водный образ жизни.)
Все дело в конструкции скелета: короткий, по сравнению с шеей и хвостом, и выгнутый дугой спинной отдел позвоночника, словно арка в архитектуре, служил для снятия нагрузки на конечности, буквально подвешенные под ним и попарно сближенные (что видно по отпечаткам лап), и облегчал работу спинной мускулатуры, стягивая связки. Мощные связки имели дополнительные крепления на далеко отходящих вверх невральных отростках позвонков и развитых боковых, шевроновых, отростках. Последние особенно выступают в передней части хвостового отдела позвоночника, где при жизни динозавра к ним крепились объемные хвостово-бедренные мышцы. В свою очередь, суставные поверхности конечностей несли усиленный слой хряща, не дававший суставам быстро истираться, и, следовательно, вся конструкция была пригодна для передвижения по суше. Хвост уравновешивал шею, поэтому тоже был длинен и держался на весу. От перегрева – главной проблемы гигантских сухопутных животных – их спасали именно шея и хвост, создавая обширную дополнительную площадь (до 30–40 м2) для испарения влаги, т. е. для охлаждения. Кроме того, пока кровь протекала по длиннющей сонной артерии, она тоже охлаждалась. Потому у самых крупных завроподов шеи и хвосты не просто длиннее, чем у «мелких» родственников, но и сравнительно длиннее: увеличение размеров в роду этих динозавров происходило в первую очередь за счет удлинения шеи или хвоста. Метод редких изотопных мостиков позволяет измерить температуру тела завропода почти напрямую – по доле 13С–18О мостиков в фосфатных зубах (об этом методе говорилось выше): получается 32–38 °C в зависимости от размера. Это ниже, чем могло бы быть, если бы тело не охлаждалось, и, значит, механизм рассеивания тепла через шею и хвост работал.
Однако при такой длине шеи и, следовательно, трахеи, как у завроподов, их грудные мышцы просто не могли бы помочь легким втянуть воздух. Эта проблема была преодолена благодаря птичьему строению шейных и передних спинных позвонков и отчасти ребер, где размещались полости, связанные с выростами легких – воздушными мешками – и обеспечивавшие подачу свежего воздуха и на вдохе, и при выдохе. Опять же – как у птиц. Пневматизированные кости и вес ящера облегчали (на 10 %), и избыточное тепло не накапливали. Вероятно, помогало завроподам дышать и повысившееся в середине юрского и в меловом периодах содержание кислорода в атмосфере. Именно тогда они процветали на всех континентах, наплодив по меньшей мере 175 родов (четверть общего разнообразия динозавров). Кроме того, расчеты массы органов завропода среднего размера (30 000 кг) предполагают, что тело такого ящера могло вмещать сердце массой 200 кг и легкие объемом до 3000 л, что при частоте дыхания 3,5 вдоха в минуту позволяло получать более 200 л кислорода в час.


Несмотря на простоту зубной аппарат этих ящеров был достаточно разнообразен: от гладких «колышков» до зазубренных «клинышков», которые замещались новыми каждые 35–60 дней у разных видов. Одни завроподы могли счищать, как грабилкой, иголки с веток араукарий, другие – срезать метелки хвощей, третьи – обкусывать листья гинкго и папоротников. И все это не сходя с места, поскольку шея позволяла дотянуться куда угодно. А при сокращении «длины пробега» и энергии тратилось меньше: до 80 %, как показали расчеты на длинно- и короткошеих моделях. Правда, шея у них не выгибалась, как у лебедя (сочленение позвонков было жестким), а перемещалась как единая конструкция, наподобие стрелы башенного крана. Неразжеванная пища из относительно маленькой пасти отправлялась прямо в большой кишечный тракт, расчетная масса которого в среднем составляла 2500 кг. При огромной массе тела температура в кишечнике поддерживалась достаточно высокая, чтобы обеспечить бактериальное брожение.
Рогатые и птиценогие ящеры (игуанодоны и гребенчатые гадрозавры) не могли похвастаться столь внушительными размерами, хотя их поздние представители все равно превосходили большинство крупных наземных млекопитающих: гадрозавры были до 17 м длиной и до 9000 кг массой, цератопсы – до 9 м длиной и до 7000 кг массой. Но они научились тому, чего не умели завроподы, – жевать. У тех и других ящеров морда заканчивалась роговым клювом – утиным по форме у гадрозавров, попугаичьим – у рогатых ящеров. Клювом скусывались ветки или листья, а затем все это тщательно перетиралось зубной батареей (рис. 35.3).
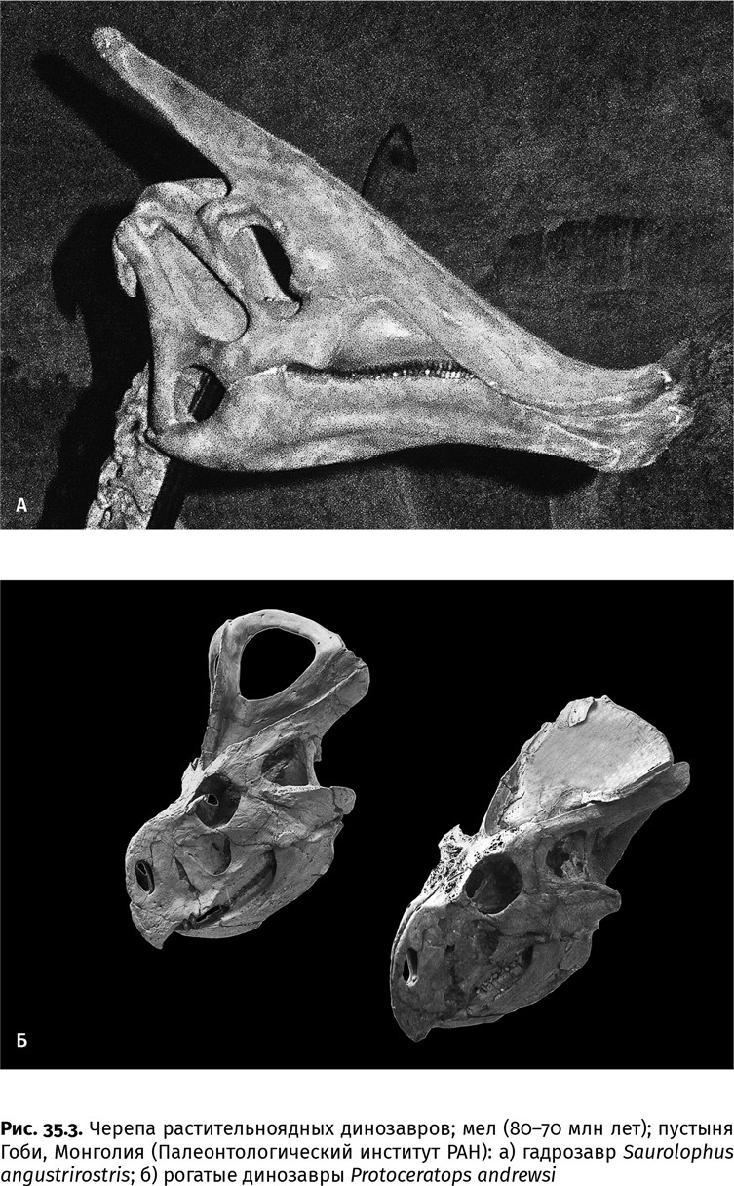
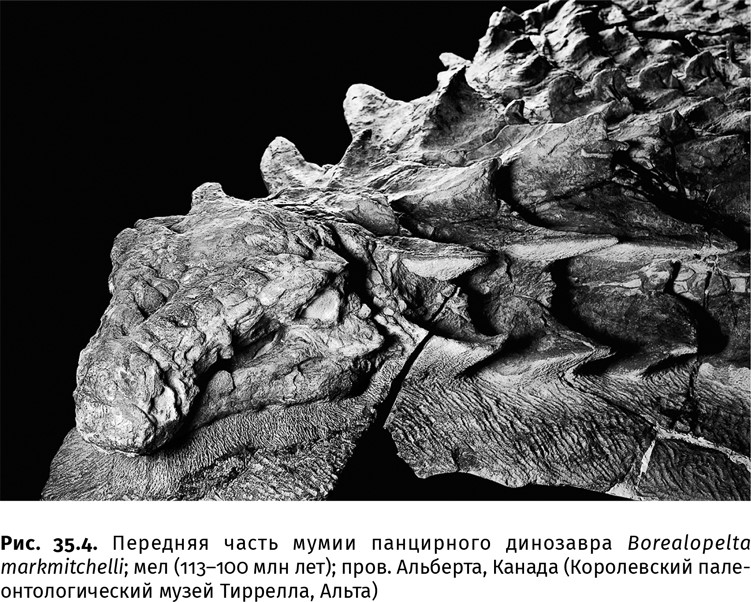
В челюстях гадрозавров наряду с рабочим зубным рядом располагалось несколько замещающих: такая батарея с острым режущим краем насчитывала до 1200 зубов, но одновременно использовалось не более трети из них. Прочие вступали в строй по мере истирания. Примерно так же была устроена зубная система цератопсов. Дополнительную прочность зубам придавали очень стойкие к стачиванию эмаль, околопульпарный и плащевой дентин, а также заполнявший углубления на жевательной поверхности цемент: вместе эти зубные ткани создавали армированную структуру. Опыты с зубами этих ящеров показывают, что твердость зубных тканей варьировала в пределах 1,7–5,6·109 Па, а темпы износа – 3,1×10–5–8,7×10–6 мм3/(Н мм) [сточенный объем зуба (мм3) к окклюзивной нагрузке (Н), приложенной вдоль жевательной поверхности (мм)] – были примерно как у современных копытных. В строении зубов эти динозавры предвосхитили крупных травоядных млекопитающих. Челюсть, судя по плотной окклюзии – характеру смыкания верхнего и нижнего зубных рядов, двигалась вперед-назад, и пища тщательно перетиралась. По подвижности челюстей, как показывает биомеханический анализ, эти две группы также превзошли всех более ранних динозавров. Панцирные динозавры, например, жевать не научились, зубы у них сменялись медленно, и эти недостатки восполнялись развитием очень сильного и подвижного языка, который был укреплен целой системой окостеневших хрящей – подъязычных костей (как у птиц, но гораздо толще). Однако такой примитивный способ питания не позволил им догнать прочих меловых растительноядных ящеров в размерах или в разнообразии (рис. 35.4).
Некоторые палеоэкологи не исключают, что растительноядные гиганты влияли на климат юрского и мелового периодов. В своих расчетах они исходят из данных по встречаемости костей завроподов в верхнеюрской формации Моррисон, распространенной на западе США на площади 1 млн км2. И если скорость обмена веществ у завроподов приближалась к таковой у современных пресмыкающихся, а не млекопитающих, то биомасса ящеров могла достигать 200 000 кг/1 км2 (примерно 100 апатозавров по 20 000 кг). Так как современные нежвачные млекопитающие производят метана примерно 0,18 л/1 кг собственной массы, получается, что каждый апатозавр мог бы испускать 2675 л этого газа в день, а общий объем динозавровых «выхлопов» приближался бы к 5,2×1011 л в год. Это больше, чем образуется метана в результате деятельности человека, и почти столько же, сколько его выделяют все современные источники, вместе взятые. Скорее всего, эти показатели сильно завышены, но полностью исключать пассивное участие динозавров в поддержании теплого мезозойского климата тоже не стоит.
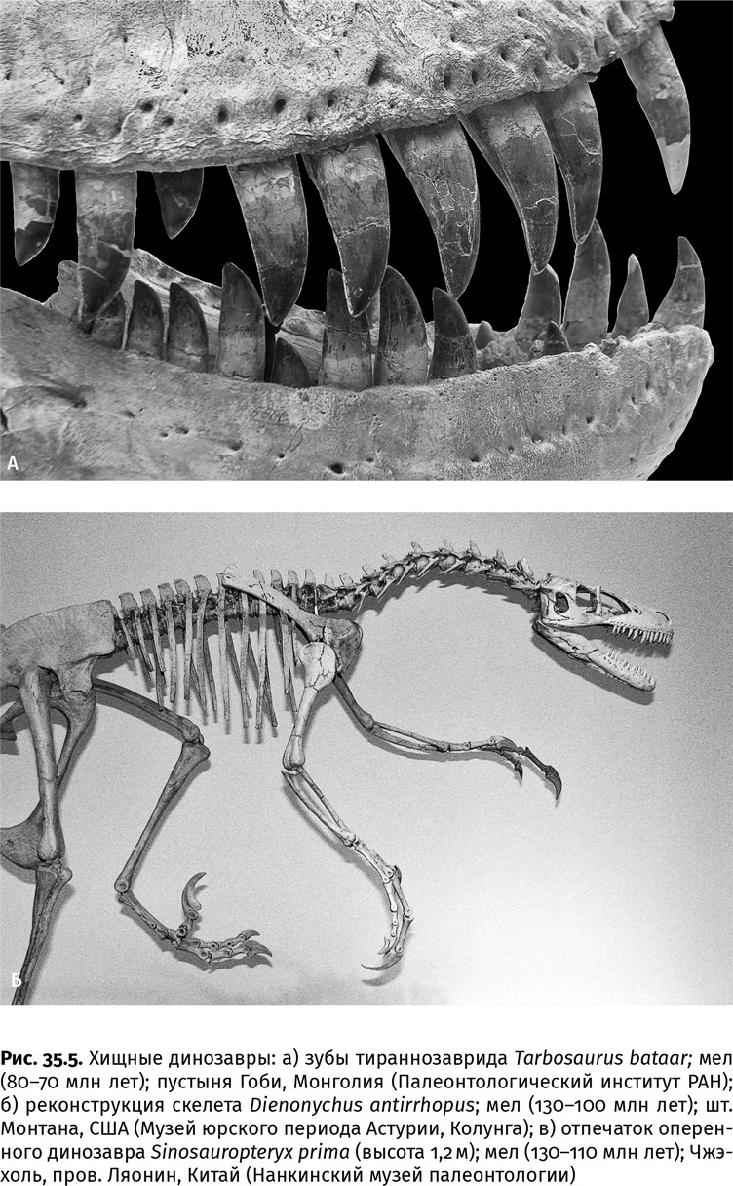
Обилие крупной добычи и огромная биомасса динозавровой молоди породили и хищников-гигантов – двуногих тираннозавридов до 14 м длиной и 5000–6000 кг массой (один только череп достигал длины 1,6 м). При внушительных габаритах тираннозавриды почти лишились передних конечностей, что компенсировалось развитием мощных челюстей и огромными крепкими зубами – округлыми спереди и уплощенными пилообразными в глубине пасти (рис. 35.5а). Сколы на зубах свидетельствуют о питании крупной добычей, а многочастичное функциональное компьютерное моделирование показывает, что сочленение и строение челюстей было достаточно прочным, чтобы пасть ящера схлопывалась с силой 35 000–57 000 Н. Только мегалодон кусался больнее. О давлении хищников-гигантов на растительноядных динозавров можно судить по залеченным ранам на костях или по патологическим изменениям костной ткани после укуса (это означает, что те не довольствовались одной падалью). Ярким свидетельством того, что вероятные жертвы пребывали в вечном страхе, является защитная окраска. Если млекопитающим массой свыше 2000 кг камуфляж не требуется (даже крупные хищники нацелены на не самую большую добычу), то, например, у панцирных динозавров размером более носорога шкура, судя по сохранившимся пигментам, была пятнистой, а в области шеи еще и покрытой острыми прочными шипами.
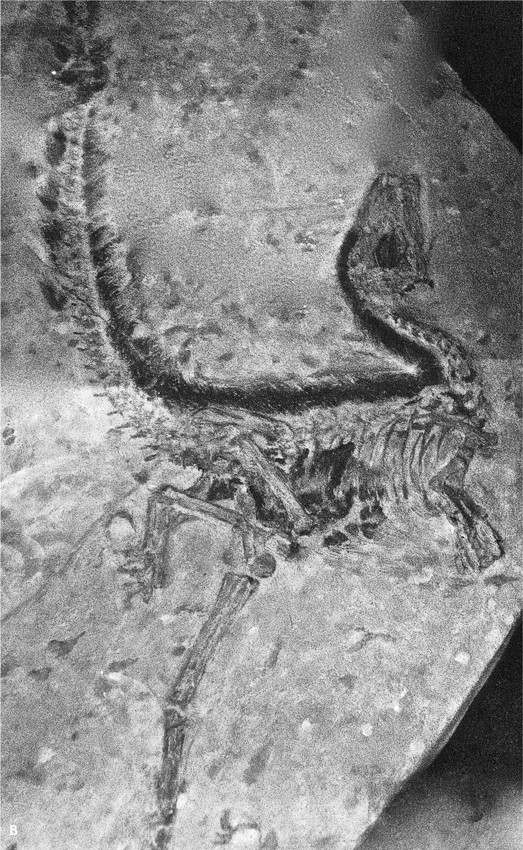
Более разнообразную группу хищников составляли манирапторы, также двуногие: подвижная нижняя челюсть и удлиненные передние конечности с тремя цепкими пальцами, приспособленными для захвата или собирания добычи, сделали их непревзойденными охотниками (рис. 35.5б, в). Конец хвоста намертво сцеплялся перекрывавшимися позвоночными отростками и плотно облекался окостеневшими сухожилиями: при беге хвост держался на весу и служил балансиром. В эволюционном развитии манирапторов размер головы уменьшался по сравнению с таковым туловища, а передние конечности удлинялись по отношению к задним: ящеры все больше становились похожими на птиц. У продвинутых манирапторов длина обеих пар лап практически сравнялась, причем передняя пара могла совершать активные маховые движения и при наличии контурных перьев создавать подъемную силу (рис. 35.6). В строении скелета, головного мозга, перьевого покрова, а также в заботе о потомстве, о чем можно судить по устройству гнезд и особенностям ямчатой скорлупы бирюзового цвета (яйца с такой скорлупой требовалось высиживать), они достигли большого сходства с птицами, в том числе современными, веерохвостыми, и, возможно, являлись предками всех пернатых. Среди наиболее рослых манирапторов возникли растительноядные формы. Они утратили или почти утратили зубы, но обрели прочный клюв, усиленный солями фосфора и кальция, которым могли ощипывать растения и раскалывать семена.

Несмотря на всю свою непохожесть, динозавры в разнообразии жизненных форм сильно уступали млекопитающим, давшим плавающих, роющих, летающих, планирующих, скачущих и бегающих зверей, причем уже в середине мелового периода, хотя то были не только сумчатые и плацентарные формы. С этого времени судьбы диапсид и синапсид вновь резко поменялись.

