Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 33 «Красный» переворот в океане
Дальше: Глава 35 Соревнование двух систем
Глава 34
Мезозойская эскалация
Чтобы на свете появилась столь разнообразная рифовая экосистема, океан должен был измениться – стать олиготрофным (от греч. ολίγος – малый, незначительный и τροφή – пища), а значит, прозрачным и заселиться хищниками нового уровня. Высокая прозрачность воды не только позволяет фототрофам освоить большие глубины, вплоть до 150 м, но и распределить по водным слоям больше разных водорослей и других уподобившихся им одноклеточных, их выедателей и т. д. – по всей пищевой цепочке. Для преобразования раннетриасовой эутрофной, насыщенной биогенными веществами «гущи» в проницаемую для дневного света водную толщу понадобились усилия новых мезо-кайнозойских фильтраторов, особенно родственников тридакны и рудистов – двустворчатых моллюсков (рис. 34.1б).
Прежде – в палеозойскую эру – их место занимали внешне несколько похожие брахиоподы (рис. 34.1а). Как и двустворки, брахиоподы питаются благодаря току воды, но создают тягу не мощные жабры и мускулистые трубки – сифоны, а мельчайшие реснички, расположенные вдоль щупалец, поддерживаемых подкововидной опорой – лофофором. Причем в отсутствие внешнего течения лофофор почти не работает, поэтому жить брахиоподам приходится на виду – на поверхности осадка, почти неподвижно и лишь слегка разворачивая раковину на своей ножке. А двустворки благодаря длинным сифонам могут скрываться от хищных глаз и клешней глубоко в осадке. Причем мезозойские виды научились, в отличие от палеозойских, проворно зарываться в ил или песок, буравя его мускулистой ногой. Даже те двустворки, что остались на поверхности, закрепляются эластичными органическими нитями – биссусом, по прочности на разрыв не уступающим кевлару, из которого делают тросы для альпинистов, или наглухо цементируются к твердой поверхности. Некоторые из них без труда, хотя и не слишком быстро, перемещаются с места на место: самые подвижные – гребешки; распространившиеся как раз в мезозойскую эру, хлопая створками, они способны даже проплыть до 10 м со скоростью 0,73 м/с (рис. 34.1б). Вдобавок быстрый рост двустворок сокращает возрастной период, в течение которого они особенно рискуют быть съеденными. Эти моллюски лучше брахиопод размножаются и меньше страдают от засыпания осадком, переворачивания и прочих невзгод. Благодаря всем этим новшествам бойкие двустворки быстро восстановились после позднепермского вымирания и заполнили пространство, где прежде обитали брахиоподы.

Это замещение привело к очень серьезным последствиям для всех морских обитателей, ведь процеживают воду двустворки в среднем от двух с половиной до четырех раз интенсивнее брахиопод (113–3155 мл/ч против 39–516 мл/ч). Причем с увеличением массы (а растут они быстрее) эта разница существенно сдвигается в пользу двустворок (до восьми раз), а крупные двустворки далеко не редкость по сравнению с большими брахиоподами (рис. 34.2е). Рост биомассы донных фильтраторов заметен по разности в мощности ракушняков: в брахиоподовом палеозое этот показатель в среднем составлял менее 0,2 м (и не более 0,65 м), а в двустворчатом кайнозое, когда обычными стали устрицы и гребешки, вырос до 0,5 м, и нередко стали появляться даже 10-метровые скопления ракушек. После перехода океана во власть моллюсков он стал заметно чище: они могут пропустить через себя весь объем океанической воды за полгода, а верхнюю, наиболее обитаемую 500-метровую толщу процедить всего за 20 дней. Может быть, темпы работ мезозойских двустворок и мшанок были несколько ниже, чем современных, но все равно прочим организмам пришлось приспосабливаться к голодным – олиготрофным – условиям.
Двустворки были не единственными «новыми» фильтраторами. В юрском периоде начался расцвет переживших позднепермское вымирание мшанок-тубулипорат (Tubuliporata, от лат. tubula – трубка и греч. πόρος – пора) – колониальных организмов с мельчайшими особями-зооидами. Хотя мшанки меньше похожи на брахиопод, чем те – на двустворок, их внутреннее строение такое же, как у других щупальцевых: рот каждого зооида окружен лофофором с ресничными щупальцами. У тубулипорат в отличие от палеозойских мшанок появились выводковые камеры, и созревшая личинка сразу по выходе могла закрепиться на грунте вместо того, чтобы подрастать, болтаясь в полной опасностей водной толще. Мшанки стали почковаться (размножаться бесполым путем) не только вдоль поверхности прикрепления колонии, но и перпендикулярно ей: так оказалось проще обрастать препятствия и нерасторопных соперников. Обызвествление крышек, прикрывавших зооиды, мешало паразитам пролезать внутрь колонии, а гибкое сочленение зооидов позволяло колонии гнуться под нагрузкой, не ломаясь. Гребнеротые мшанки (Ctenostomata, от греч. κτενός – гребень и στόμα – рот), пришедшие на смену тубулипоратам в позднемеловую эпоху, в дополнение обзавелись подвижными шипами для обороны от прожорливых голожаберных моллюсков. Гребенчатые фронтальные щитки (по ним эти мшанки и названы) надежно прикрыли поверхность зооидов. Принадлежащие к одной колонии зооиды образовывали сплошную зону облова так, что щупальца соседей почти не соприкасались, но синхронно действовали в едином темпе. Водный ток устремлялся к центру колонии, лишенному зооидов, где восходил вверх в виде сильной струи. Слаженная работа ускоряла фильтрацию, и пищи поступало больше. Щупальца у этих зооидов тоже были длиннее, чем у отдельных особей других мшанок. У автозооидов, за счет которых кормились прочие зооиды, рот открывался шире и выходящий поток не мешал входящему. В целом новшества гребнеротых мшанок оказались весомее, а обмен веществ – активнее. Их колонии развивались быстрее и подавляли рост колоний других мшанок. Пусть не в той же мере, как двустворки, но гребнеротые мшанки тоже приняли посильное участие в очищении океана.
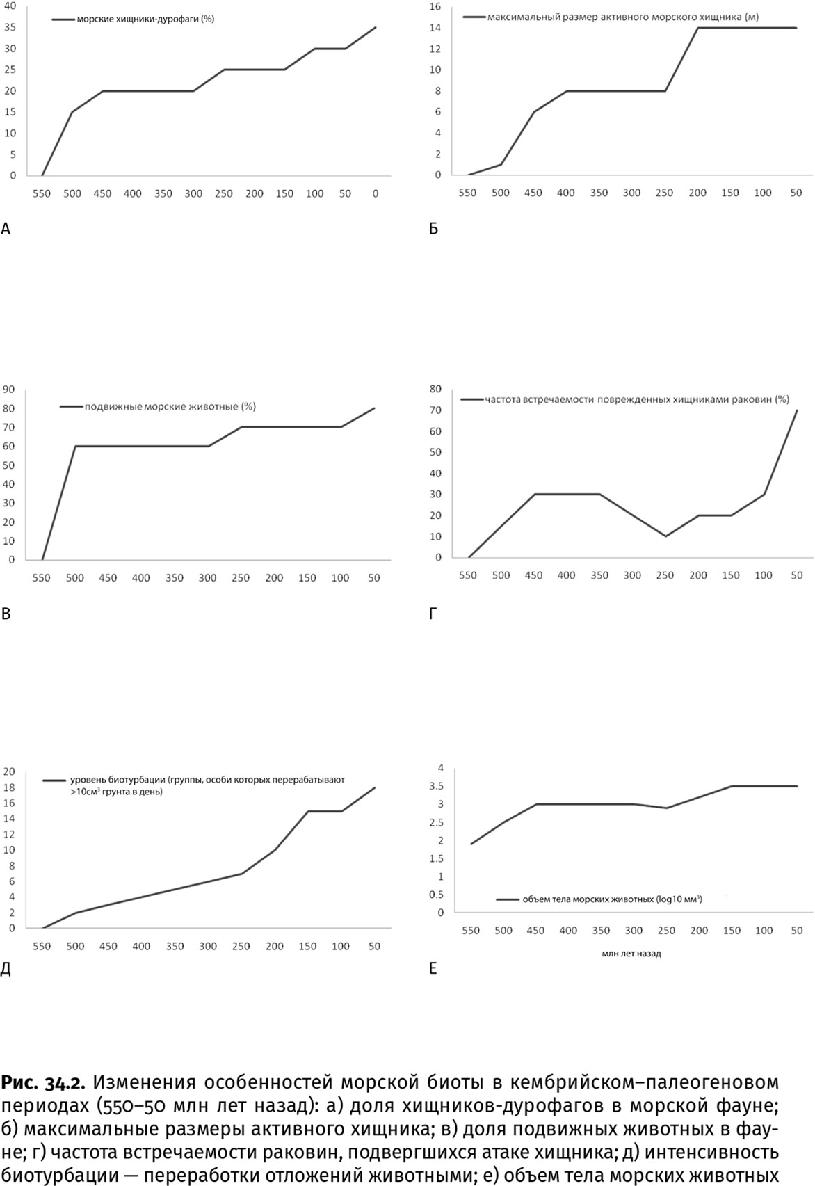
Конечно, будь то сообщество кораллового рифа или плотное поселение двустворок, подобное устричной банке, оно нуждается в помощи животных, выедающих обрастателей и не дающих тому или иному виду сообщества заполонить все пространство исключительно своими особями. Однако этого недостаточно: «увлекшись», первичные консументы могут сглодать весь риф или банку до дна. Так бы, наверное, и происходило, если бы не существовал многоуровневый «контрольный аппарат» из хищников (костные и хрящевые рыбы). Например, в упомянутом эоценовом сообществе Итальянских Альп уже существовала подобная система: крупные суповые акулы, судя по содержимому желудка, целиком заглатывали хищников поменьше – барракуд. Такая многоступенчатая трофическая пирамида обладает высокой устойчивостью. Ведь если бы крупные хищники не следили за мелкими, те съели бы всех скусывателей и выедателей, риф покрылся бы обрастателями, в итоге фотосинтез и рост замедлились бы, деструкторы-разрушители снесли бы пирамиду до основания и «все нажитое непосильным трудом» сгинуло бы в одночасье. Неслучайно мезо-кайнозойский этап развития биосферы ознаменовался ростом разнообразия морских хищников (до 35 % всех видов против 15 % в палеозойскую эру) и появлением самых крупных и прожорливых созданий (рис. 34.2а, б).
Впервые на бурный рост разнообразия морских хищников на мезо-кайнозойском интервале земной истории обратил внимание в 1977 г. американский палеонтолог Герат Вермей, назвавший эту экологическую перестройку «мезозойской морской революцией». Он же отметил, что в это время особенно стала заметна роль дурофагов (от лат. durus – твердый, крепкий и греч. φάγος – обжора) – дробителей и взломщиков крепких известковых раковин. Заметный «всплеск преступности», запечатленный в следах нападения, произошел между среднетриасовой и позднемеловой эпохами (240–66 млн лет назад). Головоногие наутилоиды, которые еще в палеозойскую эру обзавелись обызвествленной клювовидной верхней челюстью-ринхолитом (греч. ρίν – нос и λίθος – камень) для пробивания раковин, в раннеюрскую эпоху начали использовать ее как режущий и колющий инструмент. Другие головоногие – аммониты – обрели обызвествленные челюсти, устроенные, как щипцы. Многие аммониты были вооружены теркой-радулой из нескольких зубных рядов, как у осьминогов. В триасовом периоде морские ящеры – покрытые панцирем толстые короткомордые плакодонты (Placodontia, от греч. πλακόεις – плоский и οδούς – зуб) с пластинами в челюстях и некоторые ихтиозавры (Ichthyosauria, от греч. ιχθύς – рыба и σαύρα – ящерица) с широкими зубами – плющили раковины. Тогда же на смену прежним рыбам-«щелкунчикам» пришли новые: скаты и акулы с зубами, слившимися в давящие пластинки, – гибодонтные (Hybodontiformes, от греч. ύβος – горб), распространившиеся в триасовом и меловом периодах, и гетеродонтные (Heterodontiformes, от греч. ετερος – разный) – в раннеюрскую эпоху. Омары и большие креветки с клешнями для взламывания раковин расплодились в триасовом периоде, крабы и раки-богомолы – в раннеюрскую эпоху. Новые семейства моллюскоядных возникли среди рыб, морских млекопитающих и прибрежных птиц в кайнозойскую эру.
Самыми крупными мезозойскими пелагическими хищниками стали ихтиозавры. Уже в триасовом периоде они достигли 14-метровой длины (рис. 34.3). [Эти размеры превзошли только эоценовые древние киты (Archaeoceti), более поздние зубатые киты (Odontoceti), такие как кашалот, и, возможно, некоторые головоногие моллюски.] В меловом периоде с ихтиозаврами сравнялись мозазавры (Mosasauroidea, название от реки Маас и греч. σαύρα – ящерица), плезиозавры (Plesiosauroidea, от греч. πλησίος – близкий), плиозавры (Pliosauroidea, от греч. πλείον – большой) и морские крокодилы (рис. 34.2б). Приобщались к водному образу жизни все морские ящеры независимо друг от друга, но, возможно, по одной и той же причине: водная стихия снимала проблему сочетания движения и дыхания, с которой пресмыкающиеся столкнулись на суше. Бочковидная грудная клетка укрепилась брюшными ребрами, так что получился жесткий каркас, не мешавший дышать при движении и одновременно придававший телу плавучесть. Появилось у морских ящеров и много других общих признаков, позволявших им стать быстрыми пловцами и хорошими ныряльщиками. Вытянутые челюсти были усажены рядами крепких остроконечных зубов. Кости были облегченные, с полостями, разделенными костными перегородками – для прочности. Хрящевые кольца трахеи усилились, а костные пластинки, развивавшиеся в глазной оболочке – склере, прикрыли глазные яблоки, чтобы эти органы выдерживали высокое давление при погружении. Дополнительные пальцы и фаланги расширили и удлинили ласты, и в большинстве своем юрские и меловые ящеры «летали» в воде, попеременно взмахивая передней и задней парой широких ласт; позвоночник у них выгнулся, и мощный хвост с плавником как бы сам толкал тело вперед. (И спинной плавник, и верхняя лопасть хвостового развились заново из кожной складки.) Плотный, но упругий кожный покров (его фрагменты нередко встречаются в лагерштеттах) снизил лобовое сопротивление тела. (Только у мозазавров – потомков сухопутных ящериц, близких к варанам, сохранилась такая же чешуя.) Не исключено, что ихтиозавры и плезиозавры обладали способностью регулировать температуру тела, о чем свидетельствует изотопный состав их костей – не такой, как у крокодилов и холоднокровных рыб. У плезиозавров, кроме того, в скелете конечностей преобладала пластинчатая кость, характерная для быстрорастущих гомойотермных животных, а в шейных позвонках сохранились артериальные каналы. Эти дополнительные сосуды, исчезающие у большинства позвоночных по мере развития, усиливали кровоток, обеспечивая терморегуляцию и противодействуя гипоксии при погружении. Уже в триасовом периоде ихтиозавры и предки плезиозавров перешли на живорождение: в лагерштеттах того времени встречаются скелеты самок с эмбрионами в полости тела. Разительно отличалась лишь конструкция длинношеих плезиозавров, среди которых в меловом периоде появились существа с 7-метровой шеей, поддерживаемой 80 позвонками, на которой сидела маленькая голова с пастью, полной тонких и гладких загнутых зубов. Биомеханика скелета предполагает, что, поскольку шея у них хорошо сгибалась вниз, но не в стороны и не вверх, плезиозавры были удильщиками мелкой стайной рыбы или креветок и плавали у поверхности с опущенной ко дну головой.

Огромные морские ящеры могли заглатывать довольно большую рыбу, черепах, птиц, белемнитов и аммонитов общим весом до 350 кг. Впрочем, об особенно крупную добычу они сами ломали зубы, так и оставшиеся торчать в ископаемых раковинах. На глубине с ящерами за пищевые ресурсы соперничали гигантские кальмары и вампироморфы, похожие на осьминогов с перепонкой между щупальцами и парой кожистых плавников. В юрских отложениях встречаются крючья от щупальцев по 0,2 м длиной; какой величины был сам кальмар, и вообразить трудно, но никак не меньше 20 м, а позднемеловые челюсти шириной 0,1 м могли бы пригодиться 10–15-метровому вампироморфному моллюску.
Едва ящеры вымерли – а разные их группы сменяли друг друга на протяжении всей мезозойской эры (от ихтиозавров до плиозавров и мозазавров с крокодилами), – в морях подросли «новые акулы» (Neoselachii), сравнявшиеся по длине со своими хищными предшественниками; распространились разнообразные морские птицы (впрочем, они начали осваивать эту среду еще в меловом периоде, породив особую группу зубастых пернатых), а позднее, в эоценовую эпоху, к ним присоединились ластоногие и потомки сухопутных парнокопытных млекопитающих – гигантские киты-археоцеты. Уже большинство палеозойских акул обзавелось острыми зубами с дополнительными зубчиками, годными для захвата и удержания плавающей добычи. Изощренные способы разделки жертвы освоили «новые акулы»: сочленение челюстей и мозговой коробки изменилось, так что челюсти стали выдвигаться вперед и вверх относительно черепной коробки или опускаться вниз. Орудуя выдвижными челюстями, такие акулы вырывали куски плоти из противника, который был гораздо крупнее их самих, а пилообразные зубы глубоко взрезали его тело. Резкие движения головы из стороны в сторону помогали разорвать даже очень крупное животное, а увеличение массы мозга выразилось в развитии сложных поведенческих навыков. Ископаемый предшественник большой белой акулы – мегалодон (Otodus megalodon) – вырастал вдвое длиннее современного вида (до 14–16 м против 6,4 м), а зубы у него были до 0,18–0,2 м высотой (у белой акулы – 0,08 м). Распределение нагрузок в черепе мегалодона, что можно смоделировать на компьютере, позволяло акуле создавать усилие в задней части челюстей более 100 000 Н. Своими массивными пилообразными зубами с мощными корнями мегахищник мог дробить любые кости. (Давление челюстей у этой акулы более чем в 20 раз превышает максимум, доступный современному чемпиону по силе укуса – аллигатору.)
В виде обглоданных соперниками скелетов или после естественной смерти остатки гигантов опускались на дно, где на огромных трупах морских ящеров и рыб, а позднее китов образовался свой мир. Рыбы, морские ежи и аммониты спешили полакомиться падалью, оставляя на костях следы зубов и челюстей. А сидячие кольчатые черви запускали в костную ткань выросты, похожие на корни, и высасывали органическое вещество с помощью живущих в этих «корнях» бактерий: на костях морских ящеров и черепах после них образуются характерные следы растворения. Эти падальщики, точнее костоеды – название оседакс (Osedax) означает «пожиратель костей», – из-за венчика щупалец похожие на розово-красные хризантемы, настолько необычны, что зоологи долгое время и за кольчецов их не считали: родство было установлено лишь с помощью методов молекулярной биологии. (Видимые черви 2–3 см длиной, со щупальцами, исключительно самки; миллиметровые самцы застывают в своем развитии на стадии личинки и живут, порой гаремами из 14 штук, в полости тела супруги.) Сегодня поселение «пожирателей» за два месяца вычищает до белизны многометровый китовый каркас.
В мезозойскую эру среди хищников важное место заняли и костистые рыбы. Полностью окостеневший осевой скелет, а иногда и межмышечные косточки придали их телу жесткость, необходимую для быстрого плавания, а плавательный пузырь снизил затраты на поддержание тела на плаву. Именно среди костных рыб появились стремительные пловцы – тунцы и меч-рыбы, развивающие скорость до 90–130 км/ч. Среди иглокожих крупными хищниками стали морские звезды: выворачивая наружу желудок, они приспособились захватывать сравнимую с ними по размеру добычу. И если в палеозойскую эру хищничали не более 20 % брюхоногих моллюсков, позднее такой способ питания освоили 60 % видов. Появились хищники даже среди двустворок: аномалодесматы (Anomalodesmata, от греч. ανωμαλία – несоразмерность и δεσμός – связка) заглатывали добычу – рачков – целиком, всасывая ее мощным сифоном. К концу кайнозойской эры хищниками стали свыше 60 % подвижных морских животных (в палеозойскую эру их число не превышало 40 %). Скажем, в эоценовом лагерштетте Болька остатки хищников в количественном выражении и по разнообразию резко преобладают над окаменелостями растительно- и планктоноядных рыб. (Здесь во множестве встречаются даже специализированные ночные охотники – лупоглазые рыбы-солдаты и апогоновые.)
Всю мезозойскую эру аммониты пытались противостоять более крупным, чем они сами, хищникам: сначала обзавелись плотно свернутыми раковинами, более устойчивыми на излом, чем слабо свернутые, искривленные или прямые (рис. 34.4). При этом им удалось не потерять в массе тела, что очень важно для существ, плавающих за счет усилий собственных мускулов. А ушки и кили на раковинах придавали аммонитам остойчивость при движении. Шипы, ребра и утолщенные отвороты, конечно, добавляли раковине прочности, но, поскольку плотность известковой стенки примерно в два с половиной раза больше плотности морской воды, такую раковину труднее поддерживать на плаву. Мешает она и ускорению при движении. Совместить, казалось бы, несовместимое аммонитам отчасти удалось, разгородив раковину внутренними подпорками на отсеки: фестончатые перегородки позволили обрести более прочную и притом не отягощенную скульптурой раковину. Ребристые раковины преобладали до середины мелового периода, затем их доля начала сокращаться. Все позднейшие мелководные аммониты жили в обтекаемых раковинах с очень сложными и частыми перегородками. Именно тогда они достигли своего расцвета: освоили самые разные глубины и породили гигантов до 1,7 м в поперечнике. Однако в позднемеловую эпоху хищничество и соперничество настолько усилилось, что возникли непреодолимые противоречия между требованиями защиты, плавучести и скорости передвижения. Выжили те, кто совсем отказался от раковины, – белемниты, кальмары (их близкие родственники), осьминоги и другие внутрираковинные головоногие. От торпедовидных белемнитов с двумя широкими горизонтальными плавниками, как правило, сохраняется только похожий на наконечник стрелы твердый и тяжелый ростр. При жизни он служил белемнитам грузилом, уравновешивавшим большую голову с длинными щупальцами. В углублении на переднем конце ростра иногда сохраняется маленький конус с перегородками – рудимент многокамерной раковины, от которой вперед отходит длинная узкая пластина. У кальмаров осталась только прозрачная тень этой пластины – органический гладиус. Разворачивая воронку, белемниты и кальмары двигались любым концом вперед, что повышало их маневренность. Кальмары развивают скорость до 55 км/ч и, разогнавшись, пролетают над водой до 45 м. А вот заключенные в раковину наутилусы проплывают в час менее 1 км, их и осталось всего пять видов. Позднемеловые аммониты тоже пытались превратить раковину во внутреннюю, и, возможно, их потомками стали осьминоги.

Давление хищников привело к тому, что появилось намного больше двустворок с волнистым краем и быстро зарывавшихся улиток с прикрытым устьем. Неплотно свернутые улитки стали редки уже в палеозойских морях и почти пропали в мезозойских. Башенковидные раковины с толстым отворотом или суженным устьем надежно защищали моллюсков от посягательств через естественный вход, а крупные размеры, толстая стенка, узкое удлиненное устье с зубцами и другими защитными ухищрениями и усиленная наружная скульптура раковины оберегали от лузгальщиков с клешнями.
Лучшая защита – плотно сомкнутые створки, которые не позволяют выйти даже запаху, ведь многие хищники полагаются на обоняние. Если в девонском периоде двустворок с волнистым краем раковины не было (за ненадобностью усиленной обороны), то в юрском доля таких особей в сообществах выросла до 25 %, а ныне составляет почти половину. Среди усоногих раков преимущество получили морские желуди, которые закрепляются на скалах (или чужой шкуре) с помощью цементирующих выделений, превосходящих по прочности любой самый надежный клей, а ножки используют только для фильтрации. Хрупкие животные, такие как стебельчатые лилии, брюхоногие и двустворки с тонкими гладкими раковинами, в юрском периоде спустились в глубины океана, где низкая температура отрицательно влияет на скорость обмена веществ и тем самым ограничивает возможности хищников (рис. 34.5). А медленнорастущие обызвествленные губки (родственники когда-то обильных строматопорат и хететид) и древние голоротые мшанки нашли убежище в подводных пещерах. Там, где хорошему хищнику повернуться негде, эти пережитки палеозойской эры и скрываются.
История двух последних крупных групп брахиопод с известковой раковиной особенно показательна: ринхонеллиды (Rhynchonellida), наращивая толстые и высокие ребра на раковине, долго сопротивлялись хищникам, но с появлением в меловом периоде новых, хорошо вооруженных взломщиков уступили и спрятались в подводных гротах или холодных водах. Наоборот, ребристые теребратулиды (Terebratulida), которые не только усилили раковину, но и начали выделять опасные для сверлильщиков вещества, все еще живут на опасном мелководье (рис. 34.1а). Одновременно число родов ринхонеллид сократилось вдвое, а теребратулид возросло вчетверо.
Сложнее оказалось укрыться от сверлильщиков, к которым добавились самые «профессиональные» в этом отношении группы – круглые черви, осьминоги, мурицидные (Muricidae), натицидные (Naticidae) и некоторые другие улитки. Взломы и сверления заметно участились к концу мезозойской и в кайнозойскую эру, затронув не менее 80 % особей, причем до 44 % попыток заканчивались для жертвы летальным исходом (в палеозойскую эру такие попытки составляли 40 и 20 % соответственно). В целом же с ордовикского по четвертичный период средний размер сквозных отверстий в раковинах увеличился почти в 10 раз (с 0,35 до 3,25 мм) (рис. 34.2 г). Этот тренд указывает и на рост средних размеров самих хищников, и на повышение их активности (сверление – очень энергоемкий процесс). Причем соотношение размеров в паре «жертва – хищник» выросло почти в 70 раз (!) и, конечно, в пользу хищников. Среди брахиопод и двустворок от сверлильщиков меньше страдали формы с толстой раковиной и более плотно расположенными на ее поверхности ребрами. Вдвое участились и случаи залечивания. В мезо-кайнозойское время сверлильщики предпочитали охотиться на двустворчатых моллюсков, а не на брахиопод, что вполне логично: толщина раковины, которую необходимо преодолеть, сопоставима, а съедобных мускулов в двустворках намного больше. В итоге брахиоподы продолжили угасать, а двустворки – в противостоянии с хищниками – достигли невиданного разнообразия.
На поверхности дна спокойнее чувствовали себя те животные, которые либо намертво прирастали к субстрату, либо могли переместиться в сторону, поскольку в юрском периоде средняя глубина вскапывания осадка возросла до 1,0–1,5 м против 0,1–0,2 м, на что были способны палеозойские биотурбаторы, а приповерхностные 0,1 м превратились в подвижный, постоянно перекапываемый субстрат, неоднократно пропущенный через чьи-либо кишечные тракты. Бурная деятельность новых биотурбаторов особенно досаждала сидячим и свободно лежащим фильтраторам. В мезозойскую эру двустворкам пришлось или периодически «вспархивать», как морские гребешки, или вцепляться в скалистый грунт, как устрицы. Лишь до конца мелового периода некоторые устрицы пытались просто отлежаться – внушительные размеры останавливали слишком активных биотурбаторов. Но и они не выжили в кайнозойских морях, когда даже без волн и течений половина раковин оказывается перевернутой, а четверть – погребенной неуемными копателями всего за 40 дней. Если в раннекаменноугольную эпоху на дне неподвижно и свободно лежало более половины обитателей, то ныне таких почти не осталось. Прочие либо вымерли, либо сместились на скалы, в холодные воды, на глубины или в соленые, отрезанные от моря лагуны (рис. 34.6).

Многие биотурбаторы зарывались в осадок, чтобы просто скрыться от хищников. Закапываясь глубже, двустворки обрели длинные сифоны и подвижное сочленение створок и приспособились к пониженному содержанию кислорода; форма их раковины стала уплощенной. Ходы таких моллюсков отличаются ступенчатой формой с расширением в форме ракушки на конце, поскольку моллюск, пробиваясь в глубь осадка, то раздвигает, то смыкает створки. С юрского периода в большом числе развелись креветки, живущие большими колониями: они покрывают обширные участки дна вертикальными норками с боковыми ответвлениями, стенки которых укрепляют собственной слизью и пеллетами. Другие животные осваивали грунт в поисках пищи, и в позднетриасовую и раннеюрскую эпохи появились биотурбаторы, способные переработать огромные объемы осадка. Многощетинковые кольчецы-пескожилы, известные с триасового периода, взрыхляют 44 см3 осадка в день до глубины в 0,3 м (каждая особь). Голотурии – иглокожие, в мезозойскую эру избавившиеся от внешнего скелета, перерабатывают за то же время до 2250 см3 осадка, прокладывая очень простые вертикальные шахты до 1,8 м глубиной. Всех превзошли неправильные морские ежи, возникшие в раннеюрскую эпоху и ставшие обычными в позднемеловую: они вскапывают до 8520 см3 в день на глубине до 0,15 м, образуя широкие горизонтальные туннели с менисковидным заполнением. При этом им пришлось распрощаться с правильной шаровидной формой тела и длинными иголками – их иголки больше напоминают недельную щетину, но помогают вбуравливаться в грунт. Половина этих иглокожих перешла на такой образ жизни. Позднее к ним присоединились роющие крабы и скаты – копатели-чемпионы (12 000 см3 в день). На современные темпы переработки грунта – 20 700 км3 (или 8 с половиной Эверестов) в год – биотурбаторы вышли в неогеновом периоде (рис. 34.2д).
В мезозойскую эру к биотурбаторам добавились почти незаметные прежде камнеточцы. В отличие от сверлильщиков, которые трепанируют чужие раковины, чтобы поживиться их обитателями, камнеточцы скоблят и бурят рифовые и другие скальные грунты, а также ракушняки (особенно толстораковинные устричники), чтобы найти там надежное убежище или выесть бактерии и водоросли, обитающие в микрополостях. Появившиеся в палеозойских морях представители этой гильдии (водоросли, грибы, фораминиферы, губки, мшанки) проникали на глубину до 0,02–0,03 м, редко 0,05 м и оставляли очень тонкие извилистые трубчатые полости. Из этих организмов наибольшие объемы скального грунта перерабатывают губки клиониды (Clionidae) – до 7 кг/м2 в год. Их мезокайнозойские собратья по цеху (двустворчатые и брюхоногие моллюски, хитоны и морские желуди) забуривались на 0,15 м, а двустворки терединиды (Teredinidae), или корабельные черви, стали прокладывать ходы в плавающих стволах деревьев (позднее – в кораблях и деревянных сваях). Улитки морские блюдечки с колпачковой раковиной (Fissurellidae, Patellidae и некоторые другие) и многостворчатые хитоны, вооруженные железными зубами (из окислов железа – магнетита и маггемита), разъедают породу, включая кварцевый песчаник, выделяя краем ноги и мантии кислые соединения, а затем проскребывают разрыхленный материал своей теркой, оставившей царапины уже на верхнеюрских скалах. А многие морские ежи стали вгрызаться на 0,1 м в известняк и даже в базальт, перерабатывая до 10 кг/м2 грунта в год (рис. 34.7). Внедрились в твердый грунт они в юрском периоде, когда вместо желобовидных зубов обзавелись долотовидными, а отвороты панциря, поддерживавшие зубы, срослись и опора для жевательных мышц упрочилась.

По сравнению с палеозойской недвижностью донных животных в мезозойскую и кайнозойскую эры все буквально расползлось и расплылось во все стороны: рыбы, кальмары, улитки, крабы и морские ежи (рис. 34.2в). Даже морские лилии сорвались с места, помогая себя более гибкими, чем у их предков, руками, а шестилучевые кораллы и мшанки, в отличие от своих палеозойских предшественников, научились двигаться. Засыпанные осадком кораллы протискиваются сквозь ил и песок наверх; вбирая в себя воду и раздуваясь, цепляясь щупальцами, они перелезают через преграды. Мшанки ползают, используя как тягловую силу членов колонии, оказавшихся снизу. Но лучший способ перемещения заключается в том, чтобы оседлать более подвижных улитку или краба. А рыбы вообще полетели – в среднетриасовую эпоху над водой на четырех плавниках воспарили кривобрюшки: бегство в иное пространство тоже было способом озадачить хищника и спастись.
В мезозойских и особенно кайнозойских морях животные с активным обменом веществ заняли место тех, кто был слишком пассивен. Только у брюхоногих моллюсков темпы обмена веществ в течение мезозойской эры возросли на 150 %. В целом животные с активным метаболизмом в кайнозойскую эру составили 60 % против 30 % в палеозойскую, а подвижные – более 80 % против 40 %. Во много раз выросло разнообразие морских организмов – как в целом, так и в отдельных сообществах. В палеозойскую эру просто организованные сообщества (с небольшим числом видов и гильдий) составляли более половины всех донных сообществ, на мезо-кайнозойском этапе до 70 % возросла доля сложных сообществ, насыщенных видами с очень разными функциями.
Если растения поставляют биогенные вещества с суши в океан, то в конце триасового периода появились животные, возвращавшие, по крайней мере, фосфат обратно. То были летающие диапсиды – птерозавры (Pterosauria, от греч. πτερόν – крыло и σαύρα – ящерица) (рис. 34.8а, б). Судя по обильным следам этих ящеров на приморских отмелях, изотопной подписи кислорода в их костях, такой же, как у рыб, а также по особенностям биомеханики полета, предполагающей использование прибрежных восходящих потоков воздуха (термиков), это были в основном рыбоядные хищники.
Встречаются ископаемые остатки птерозавров с рыбьими костями в желудке и водных ящеров – с костями птерозавров; найден даже скелет крылоящера, намертво схваченного костной рыбой и утянутого ею на дно (поскольку проглотить жертву рыбе не удалось, погибли оба) (рис. 34.8в). Еще большее сходство с птичьими базарами проявляется в гнездовьях птерозавров, о чем свидетельствуют обильные находки яиц в прибрежных озерных отложениях. Яйца в таких захоронениях сохраняются, несмотря на мягкую скорлупу. Слабое окостенение скелета конечностей и черепа (толщина компактной кости не превышает 0,2 мм) и отсутствие зубов у зародышей подсказывают, что, вылупившись, птерозаврики не могли ни летать, ни питаться самостоятельно. Выкармливать детенышей приходилось зубастым родителям. Так что птерозавровые «базары», вероятно, стали местами накопления гуано – концентрированных фосфатных отложений.



