Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 32 Возвращение в протерозой
Дальше: Глава 34 Мезозойская эскалация
Глава 33
«Красный» переворот в океане
Необычная обстановка способствовала резкому обновлению состава водорослевого одноклеточного планктона. Вместо практически исчезнувших акритарх, представлявших зеленые водоросли, в триасовом океане всплыли динофлагелляты (250 млн лет назад), кокколитофориды (227 млн лет назад) и диатомовые (не позднее 205 млн лет назад), которые использовали пигменты, восприимчивые к разным по длине световым волнам. Впрочем, заметной силой эти водоросли стали в юрском периоде и тогда, по словам американского океанографа Пола Фальковски, совершили в океане «красную» революцию, ведь их предшественники использовали для фотосинтеза в дополнение к хлорофиллу а еще и b, т. е. зеленые пигменты, а неофиты предпочли хлорофилл с и фикоэритрин, придающий клеткам золотисто-оранжевый или красноватый оттенок. Сама по себе «цветная» революция не удивительна – пигменты, обеспечившие ее, оказались более выгодны для фотосинтеза в тусклых водах океана Тетис, обогащенных биогенными веществами и нередко «цветущих». В результате преимущество и получили те, кто мог выжить в подобных условиях. Стимуляторами роста у новых групп водорослей служат те микроэлементы, которые хорошо растворимы в насыщенных кислородом водах, – марганец, кобальт, кадмий. (В отличие от них зеленые полагаются на железо, медь и цинк, характерные для анаэробных условий.)
Различаются эти водоросли по дополнительным пигментам, составу и форме раковинки, набору органелл, количеству и строению жгутиков. Так, у кокколитофорид (Coccolithophorida, от греч. κόκκος – зерно, λίθος – камень и φορέω – носить) клетка заключена в ажурную раковинку, состоящую из многочисленных известковых элементов – кокколитов, которые похожи на сдвоенные, соединенные короткой трубочкой колесики. У этих водорослей есть два двигательных жгутика на одном из полюсов клетки. Между ними располагается третий – совсем тонкий и свернутый в спираль, который в основном используется для прикрепления к субстрату и другим клеткам. [Этот жгутик, или гаптонема (от от греч. άπτω – прицеплять и νημα – нить), дал другое название этим существам – Haptophyta]. Кокколитофориды – одни из самых мелких живых существ (менее 0,03 мм): в литре морской воды (а в другой они не живут) может поместиться до 200 млн особей.
Диатомовые (Diatomeae, от греч. διατομή – рассечение), наоборот, довольно крупные по меркам микромира существа – до 1 мм в поперечнике. Эти организмы лишены жгутиков на зрелой стадии, окрашены в коричневатые тона и строят округлые или продолговатые двустворчатые пористые раковинки, своего рода коробочки с крышечками, из легко растворимой разности кремнезема – опала. Когда клетка размножается бесполым путем, она делится – рассекается надвое, и одному потомку достается большая половинка, а другому – маленькая (оба достраивают недостающую створку меньшего размера). Успеху диатомовых сопутствовали появление внутриклеточных вакуолей, в которых можно было запасать биогенные вещества, и улучшение контроля над внутриклеточными циклами углерода и главным образом азота, а также особого генного комплекса, существенно улучшившего способность поглощать ортокремневую кислоту и утилизировать кремний. Диатомовые распространились в высокоширотных океанах и озерах. Они удивительно живучи: могут размножаться в горячих источниках, прижились в почве и даже на деревьях и настолько хорошо приспособились к жизни в холодном климате, что заселили толщу снега и льда, придавая замерзшей воде красноватый оттенок. Во льду в некотором отношении жизнь даже легче – меньше тяжелых изотопов водорода и кислорода, и съесть буроватое мороженое с диатомовыми непросто. Полярными ночами они впадают в «спячку» (анабиоз) и могут пролежать так до 180 000 лет. Возможно, именно изо льда диатомовые переселились в пресные водоемы. Плавают они большими скоплениями, погруженными в липкую слизь. Скопления оседают на дно, унося с собой опал, поэтому эти одноклеточные стали главным источником осадочного кремнезема как в морях, так и в озерах и даже болотах.
Динофлагелляты (Dinoflagellata, от греч. δινέω – вертеть и лат. flagellum – плеть, кнут) используют два жгутика разной длины: один, расположенный в продольной бороздке, – в качестве руля, другой, поперечный, – винтового мотора. Поперечный жгутик волнообразно изгибается вокруг клетки, создавая волну, на «гребне» которой клетка и плывет, вращаясь. Панцирь у динофлагеллят гибкий, состоит из отдельных целлюлозных пластинок (размером от 0,005 до 2 мм), образующих нечто вроде шлема странной формы. У силикофлагеллят (Silicoflagellata, от лат. silex – кремень и flagellum – плеть, кнут), появившихся в меловом периоде (115 млн лет назад), есть только один жгутик и множество тонких ложноножек, а тельце (0,02–0,1 мм в диаметре) покрыто кремневыми пластинками, что и отразилось в их названии.
Хотя все эти водоросли имеют красноватые пигменты, они являются родственниками не в большей степени, чем посетители ресторана, заказавшие одно и то же блюдо. Просто хищные предки каждой из этих групп в мезозойскую эру «съели» красную водоросль – обзавелись вторичным эндосимбионтом. Если первичные эндосимбионты – митохондрии и пластиды – окружены в клетке двумя оболочками, то вторичный – тремя. (Встречается и третичный эндосимбиоз – у водорослей, которые поглотили целую диатомею.) Некоторые динофлагелляты еще помнят о своем хищном прошлом и, убивая рыбу ядом, потом питаются ее мясом. Гаптофиты с помощью гаптонемы захватывают пищу; диатомовые образуют мочевину, которую, правда, не выделяют наружу, а используют как исходный субстрат в цикле азота, поэтому все эти одноклеточные водоросли считаются миксотрофами – организмами, способными и к фотосинтезу, и к другим типам питания. Кокколитофориды, диатомовые и силикофлагелляты, более близкие родственники: они относятся к страменопилам, а динофлагелляты представляют совсем другую ветвь органического мира – альвеолят.
Появившись на свет, новый водорослевый планктон горы своротил, точнее, наворотил. Ведь притом что фитопланктон составляет менее 1 % всей растительной биомассы планеты, на него приходится почти половина ежегодной продуктивности. За миллионы лет из раковинок диатомовых и силикофлагеллят образовались осадочные кремнеземы – диатомит и опока, а из чешуек кокколитофорид – писчий мел (рис. 19.10 г). Чтобы эти осадочные толщи формировались, раковинки погружались в холодные глубины океана, а вместе с отмершими клетками уходила и часть атмосферного углекислого газа. Ведь двуокись углерода растворятся в океане и включается водорослевым планктоном в обмен веществ, причем 15 % органического вещества, которое образуется из СО2 при фотосинтезе, возвращается обратно лишь через сотни лет, а небольшая доля органики (менее 2 %) попадает на дно. Однако за десятки миллионов лет эта «небольшая доля» преобразуется в нефтематеринские породы – источники нефти и природного газа. Крупнейшие месторождения природного «топлива» (не менее 70 % всего существующего нефтегазового сырья) в Северной и Западной Африке, на Ближнем и Среднем Востоке, на Аравийском полуострове, в Индостане, Индокитае и Индонезии, Южной Америке и Карибском бассейне, Западной Сибири, Арктике и Северном море (практически в любой стране, добывающей более 10 000 кг нефти в день) образовались за время существования этого планктона в мезозойскую и кайнозойскую эры.
Одновременно водорослевый планктон изъял существенную часть двуокиси углерода из атмосферы, тем самым нейтрализовав последствия пермско-триасового вулканического катаклизма и способствовав понижению температуры. Кроме того, органическое вещество погребалось на дне океана без доступа кислорода, и по мере формирования нефтегазовых месторождений атмосфера насыщалась живительным газом. А новая кислородная атмосфера породила гигантов суши: сначала крупнейших из всех когда-либо ступавших по Земле существ – завроподных динозавров со своеобразным, почти птичьим дыханием, а также самых больших летающих животных – птерозавров, а затем птиц с их энергозатратным механизмом полета и плацентарных млекопитающих, развитие двух самых важных органов которых (плаценты и мозга) требует больших объемов кислорода.
На этом участие мельчайших существ в большой истории не закончилось. С помощью прямых наблюдений за изменениями температур и компьютерного симулирования установлено: одно из самых грандиозных на Земле явлений – тропические циклоны – тоже зависит от микроскопических планктонных водорослей. Из-за пигментов, наполняющих клетки, скопления планктонных водорослей придают поверхности океана более темную окраску. Если планктон отсутствует, солнечный свет коротковолнового диапазона рассеивается в глубинах, не влияя на температуру воды, а в области таких скоплений поверхность океана нагревается. Это и есть одно из условий зарождения циклона. Поскольку планктон отнюдь не однороден, а состоит из весьма «пестрых» групп водорослей, приспособленных к разным глубинам обитания и другим особенностям среды, то изменение состава планктонных сообществ может влиять на место зарождения циклона и на его силу. Изучение темпеститов – морских отложений, сформированных ураганами, показывает, что именно в меловом и палеогеновом периодах частота их встречаемости и мощность возросли как никогда. Скажем, средняя мощность ураганных наслоений достигла 0,9–1,3 м против прежних – менее 0,5 м (рис. 33.1, 33.2и, к). Дело не просто в высокой температуре на меловой Земле: в конце пермского или в силурийском периоде она была не ниже, но подобные толщи не накапливались. Значит, с распространением нового фитопланктона ураганы действительно стали более частым и разрушительным явлением – гиперганами.

Чтобы быть планктоном (плавать), одноклеточным водорослям необходимы вещества, поддерживающие плавучесть. К подобным веществам, синтезируемым водорослями, принадлежит диметилсульфониопропионат. После отмирания клетки это серосодержащее соединение распадается под действием бактерий до диметилсульфида (ДМС). Название подсказывает, что основу молекулы ДМС составляет ион серы, к которому присоединены две метильные группы (СН3). Летучие частицы ДМС поднимаются вместе с водяным паром в атмосферу, где окисляются до сульфатов и метансульфокислоты, продуктом распада которой является серная кислота. Все эти аэрозоли служат затравкой для облаков. В современном мире клетки таких водорослей, как кокколитофориды, содержащие больше атомов серы на атом углерода, чем органеллы зеленых, ежегодно поставляют в атмосферу до 37 000 кг серы – в 1,7 раза больше, чем наземные вулканы, поэтому их скопления в океане одновременно являются центрами образования облаков. Затем вместе с потоками воздуха облака перетекают в сторону суши и проливаются дождями. Сейчас более трети облаков, проплывающих над континентами, приходит со стороны океана, где появляется благодаря морскому фитопланктону. Так необходимую влагу получают наземные растения. И хотя ныне одноклеточные организмы производят серы меньше, чем люди (в 8,3 раза), благодаря своему океаническому положению именно они в первую очередь отвечают за формирование облачного покрова. В доиндустриальную эпоху фитопланктон являлся основным фактором формирования облачного покрова, особенно в юрский и меловой периоды, когда кокколитофорид было гораздо больше, чем ныне, о чем свидетельствуют меловые утесы по всей Европе – от Дувра до Волги и от острова Рюген в Балтийском море до Крита в Средиземном, почти нацело сложенные кокколитами – по 10 млрд штук в каждом 1 см3. По мере увлажнения на месте триасовой пустыни разрастались оазисы. А поскольку облака белого цвета, альбедо планеты – способность ее поверхности отражать падающий поток излучения – возросло. Ведь если бы не водоросли, облачный покров был бы тоньше и солнечное излучение сильнее нагревало бы Землю, а не рассеивалось в космосе. Этот фактор опять же повлиял на снижение температуры в конце триасового и в юрском периоде.
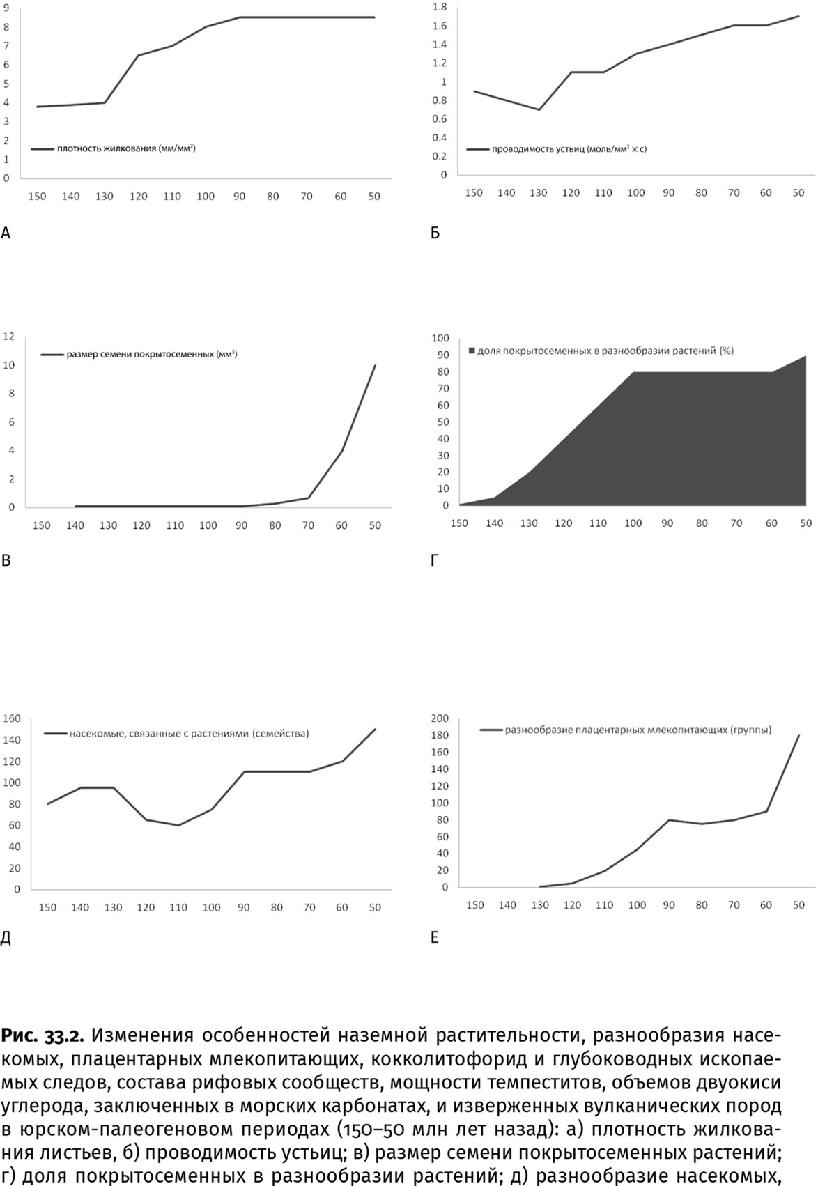
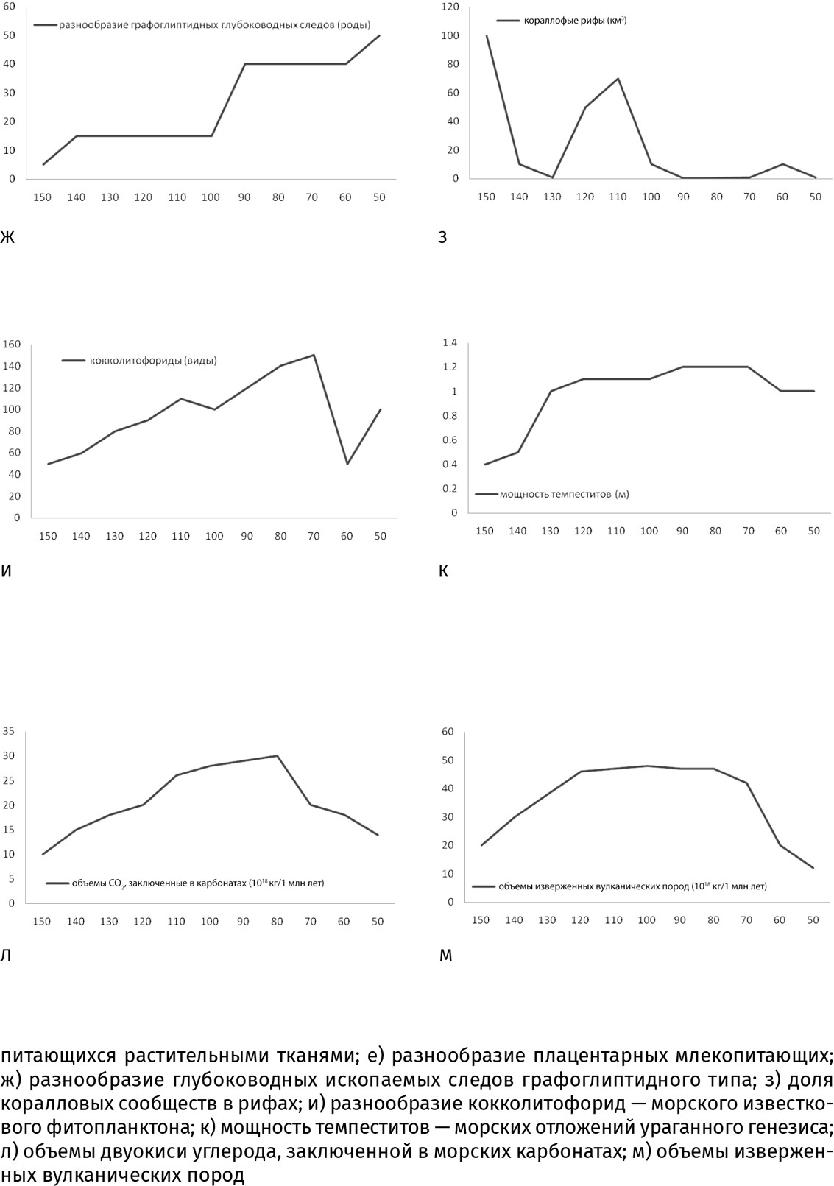
Массы кокколитов продолжали уходить на дно не только в юрском, но и в меловом периоде, когда кокколитофориды обзавелись более устойчивыми к растворению скелетиками, что привело к уже совсем иным последствиям. Ведь прежде все известняки формировались только на континентальном шельфе, и распространение известкового планктона в открытом океане сдвинуло центр карбонатонакопления в глубины – в область материкового склона и океанического ложа. Там начали накапливаться пелагические известковые илы, образовавшие километровые по мощности залежи писчего мела, когда-то покрывавшие треть всей площади дна (рис. 19.10 г). Известняки (а мел – это тоже известняк) оказались ближе к зонам субдукции, где тяжелая океаническая кора затягивается под легкую континентальную. Появление кокколитовых илов в зонах субдукции сказалось неожиданным образом: возросло число землетрясений и цунами, поскольку по мере роста температуры и давления известковые илы быстрее теряют устойчивость, чем глинистые пласты, и вдоль границ карбонатных слоев на малых сейсмогенных глубинах (от 10 до 40 км от поверхности литосферы) формируется зона разрывов, что и приводит к землетрясениям. (К тому же для субдукционной переработки карбонатов при давлении 3×109 Па их нужно нагреть до температуры всего 100 °C – меньше, чем базальты и тем более перидотиты.)
Кроме того, когда мел стал «перевариваться» в вулканических очагах и время пребывания углерода в карбонатных толщах резко сократилось, в вулканических выбросах возросла доля углекислого газа, выделявшегося при преобразовании известковых пород в силикатные (кремнеземсодержащие) (рис. 33.2и, л, м). Предполагается, что до 70 % первичного углерода, оказавшегося в зонах субдукции, возвращалось в атмосферу в виде его двуокиси. В это время континентальные и островные вулканы дополнительно извергали от 5,5×1011 до 5,5×1012 кг СО2 в год. Переплавленные известковые илы по-прежнему служат источником углекислых выделений для таких, можно сказать, знаменитых вулканов, как Попокатепетль в Мексике, Санторин в Греции, Этна и Везувий в Италии, изучение которых и позволило получить вышеприведенные значения. (Кальций из илов тоже не исчезает бесследно, а входит в состав новообразованных минералов скаполитовой группы, подобных мейониту, – Ca3Al6Si6O24 × СаСO3.)
В итоге вторая половина мелового периода и последующий палеогеновый стали одним из самых теплых интервалов в истории планеты. Температурный пик пришелся на середину палеогенового периода – эоценовую эпоху, когда змеи достигали длины 13 м (а размер этих рептилий прямо зависит от среднегодовых температур), в Арктической Канаде (75° северной широты) размножались морские крокодилы и черепахи, а на Чукотке росли теплолюбивые пальмы нипа (Nypa). Тогда же Центральная Европа больше напоминала современную Юго-Восточную Азию: росли панданы, среди которых водились панголины, лемуроподобные приматы, трогонообразные птицы и попугаи, питоны, а в озерах плавали мягкотелые черепахи и разнообразные крокодилы. Конечно, для оценки палеотемператур используются не только эти качественные показатели, но и количественные: устьичный индекс понизился, изотопные подписи бора и кислорода в ископаемых планктонных раковинах достигли заметных пиков и т. д. Все это указывает на то, что уровень углекислого газа в атмосфере 56–52 млн лет назад в три с половиной раза превышал доиндустриальный (=0,028 %), даже океанские глубины прогрелись до 12 °C (сегодня – 4 °C), а подкисление вод препятствовало осаждению карбонатных илов.
Таковы оказались некоторые (далеко еще не все) итоги «красной» революции: несмотря на мелкие размеры, кокколитофориды и другие планктонные водоросли проявили себя настоящими «большевиками».
На этом влияние «мелочи» на происходящие события не закончилось. Разнообразие плавающих водорослей вызвало в юрском периоде бурный рост планктонных фораминифер (палеозойские представители этой группы одноклеточных обитали только на дне), которые сначала питались водорослями, а затем вступили с ними в более тесный контакт: стали использовать динофлагеллят в качестве фотосимбионтов и практически перестали питаться сами. (В живом виде такие фораминиферы отличаются буроватым цветом, таким же, как у динофлагеллят, а в ископаемом – изотопной подписью углерода, характерной для остатков растений.) Они обзаводились легким пористым скелетиком: раковинка состояла из нескольких камер, но, поскольку последние из них охватывали предыдущие, превращалась в шар. Подобная форма и многочисленные шипы, усеявшие поверхность, обеспечивали оптимальное соотношение поверхности и объема и тем самым – плавучесть. Так же поступили другие плавающие одноклеточные – радиолярии с ажурной кремневой раковиной, в лучистых ложноножках которых поселились все те же динофлагелляты, известные как зооксантеллы (в основном они принадлежат к роду Symbiodinium). Планктонные фораминиферы, а также брюхоногие моллюски-птероподы с еле заметной колпачковой раковиной усилили известковый поток в глубины океана в меловом периоде.
Услугами фотосимбионтов воспользовались не только планктонные организмы: в среднетриасовую эпоху сложился симбиоз шестилучевых кораллов и зооксантелл (от греч. ζωον – животное и ξαντός – золотисто-желтый), что сразу проявилось в изотопном составе скелета – он стал таким же, как у обызвествленных водорослей. Зооксантеллы не только обеспечивают своих хозяев сносным питанием, но и ускоряют у них обызвествление скелета в дневное время, поскольку удаляют главное препятствие для осаждения карбоната – ионы фосфата, а также поглощают углекислый газ и тем самым повышают в клетках кораллового полипа содержание ионов бикарбоната. В результате такие кораллы растут в три и более раз быстрее, чем их собратья, не разжившиеся фотосимбионтами, а коралловые рифы производят 6000 кг карбоната кальция на 1 км2 морского дна ежедневно. (Слишком медленный рост палеозойских ругоз исключает присутствие в их тканях водорослей-сожителей.) А поскольку зооксантеллы не утратили вольной привычки вырабатывать диметилсульфониопропионат, мезо-кайнозойские коралловые рифы стали еще одним фактором сгущения облаков. Для рифов, как и для планктонных водорослей, облачный покров – необходимое условие существования: благодаря альбедо часть солнечных лучей отражается, и поверхность моря не перегревается, а именно повышение температуры воды грозит распадом симбиозу зооксантелл и кораллов, болезнями и гибелью последних. Наблюдения за Большим барьерным рифом у северо-восточных берегов Австралии показали, что коралловая экосистема способна понизить температуру на 1 °C. (Интересно, что в безбрежном Тихом океане облака надолго задерживаются именно над рифовыми атоллами и островами и даже форму приобретают соответствующую, что могло помочь древним полинезийцам находить ориентиры при освоении раскиданных на морских просторах островов.)

Не отстали от кораллов и двустворки: самые большие из них, такие как 110-килограммовая тридакна, обязаны своими впечатляющими размерами как раз фотосимбионтам. Тридакна и подобные ей моллюски просто раскрывают пошире свои волнистые створки, чтобы вывалить наружу мантию, полную зооксантелл. Хитрее поступили другие: поскольку кальцит прозрачен, они начали строить раковины так, что в створках появились незамутненные окошки. Окошки служат и линзами (с индексом преломления, близким к индексу преломления морской воды), и световодами. Фотосимбионты чувствуют себя привольно, а до мягкого тела врагам не добраться. Подобные двустворки возникли в позднетриасовую эпоху: валловаконха (Wallowaconcha) по виду более всего напоминала сомбреро метровой величины; уплощенные «поля» этой «шляпы» служили своего рода солнечными панелями: в них под прозрачным арагонитом, в многочисленных отсеках, видимо, и размещались фотосимбионты. Особенно впечатляют юрские и меловые рудисты (от лат. rudis – палка), которые в заботе о своих сожителях совершенно уподобились коническим кораллам, только с крышечкой (рис. 33.3а). Толстую, нередко палковидную (до 2 м высотой) раковину пронизывали ветвящиеся каналы, куда проникала мантия и где гнездились водоросли. Возможно, именно симбиоз помог рудистам воздвигнуть рифовые постройки и во второй половине мелового периода даже потеснить кораллы. Гигантских размеров и высокой продуктивности благодаря фотосимбиозу достигли и одноклеточные: диаметр раковин нуммулитов (Nummulites от лат. nummulus – монетка) превышал 15 см. Эти бентосные фораминиферы имели уплощенную форму, изнутри раковина подразделялась на многочисленные камерки, куда вели прозрачные известковые столбики – световоды. Нуммулитового эоценового известняка с лихвой хватило для постройки всех циклопических египетских сооружений и еще больше осталось в карьерах (рис. 33.4).

Светолюбивые рифы сосредоточены в тропиках, где освещенность не меняется от сезона к сезону. Стремясь к солнцу, т. е. к поверхности океана, они становятся волноломами, и каждый ураган, обламывая и раскидывая веточки, способствует расширению кораллового поселения. Так даровая солнечная энергия позволила основным рифостроителям современности оторваться от прибрежной кромки и освоить обширные, но бедные питательными веществами просторы океана. К примеру, тихоокеанские рифы (такие, как Большой барьерный) возвышаются в прозрачных, легко проницаемых для солнечных лучей водах. В палеогеновом периоде шестилучевые кораллы окончательно вытеснили прочих рифостроителей, и именно симбиоз позволил им создать рифы, не уступающие по размерам строматолитовым и строматопоровым (рис. 33.3б, 33.5). При этом рифы превратились в центры биоразнообразия. Скажем, в Коралловом треугольнике, охватывающем моря Тихого океана вокруг Филиппин, Малайзии, Брунея, восточных островов Индонезии, Восточного Тимора, Палау, Папуа – Новой Гвинеи и Соломоновых островов, сосредоточено больше всего видов морских обитателей: 8295 – рыб, 1212 – моллюсков и ракообразных, 820 – рифостроящих кораллов, 50 – морских трав и 69 – мангровых деревьев.

В середине палеогенового периода (эоценовая эпоха) примерно такой же треугольник располагался в Средиземноморье, тогда еще не отрезанном от Индийского океана, и сейчас в эоценовых отложениях (возрастом около 50 млн лет) Итальянских Альп находят многочисленные остатки коралловых рыбок (более 230 видов, из них 190 – лучеперых), на которые впервые обратил внимание великий Леонардо да Винчи: «…И над долинами Италии, где теперь летают стаи птиц, обычно проносились косяки рыб». Благодаря лагерштетту Болька известны древние рыбы-хирурги (названные так за острый хвостовой шип, похожий на скальпель), рыбы-ласточки, губаны, платаксовые, апогоновые, помацентровые и многие другие обитатели рифов.
Дело в том, что коралловые скелеты привлекают всевозможных обрастателей: цианобактерии и мягкие водоросли затеняют фотосимбионтов, и кораллы могли бы совсем зачахнуть. Это не происходит только потому, что на рифах существуют гильдии выедателей и скусывателей. Первые старательно выедают обрастателей (рыбы-хирурги и ласточки) или буквально очищают кораллы от органического «мусора» – детрита (губаны и платаксовые), причем рыбы-ласточки действуют избирательно – пасутся на самых быстрорастущих водорослях, чем способствуют увеличению разнообразия кораллов. Вторые скусывают отдельные веточки, покрытые обрастателями, и выгрызают целые дорожки (рыбы-попугаи). Сравнение участков современных рифов, где эти рыбы есть, с теми, где их нет, показывает, что на первых преобладают собственно коралловые заросли (их в три раза больше, чем «на безрыбье»), а водоросли успевают занять не более четверти пространства. Для рыб риф далеко не только стол, но и дом: многочисленные полости, гроты, пространство между коралловыми ветвями, даже сами жгучие щупальца полипов служат надежным убежищем, куда не каждый хищник сунется. Вполне закономерно, что рост разнообразия апогоновых, или кардиналковых, щетинозубовых (рыбы-бабочки) и помацентровых (к ним относятся рыбы-клоуны и ласточки) приходится на время видообразования у самых ветвистых и разнообразных по форме кораллов – рода акропора (Acropora). Осваиваясь в рифовых условиях, рыбы не только обрели всевозможные специализированные челюсти и зубные системы; некоторые из них также приспособились к сильным приливно-отливным течениям в условиях крайнего мелководья над рифовыми платформами, которые составляют большую, верхнюю, часть рифовой постройки. Скажем, рыбы-хирурги и губаны приобрели мощные плавники и обтекаемое тело.
Именно тесное взаимодействие шестилучевых кораллов и костных рыб сделало рифы современного типа самой богатой на виды морской экосистемой и способствовало стремительному росту биоразнообразия в кайнозойскую эру: в течение миоценовой-голоценовой эпох (последние 34 млн лет) число семейств костных рыб удвоилось, достигнув 270. Этот стремительный рост зеркально отражал увеличение видового разнообразия рифостроящих кораллов и распространение рифовых сообществ. Вдвое возросло и число родов основных рифовых рыб – хирургов, губанов, помацентровых. Там же, где сегодня рыбы перестают водиться или мельчают из-за перелова, коралловая экосистема быстро приходит в упадок.
Назад: Глава 32 Возвращение в протерозой
Дальше: Глава 34 Мезозойская эскалация

