Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 21 Изучение морских осадков с полным погружением
Дальше: Глава 23 Все наверх!
Глава 22
Строители трофических пирамид
Понадобилось еще некоторое время – около 20 млн лет (до середины следующего, ордовикского периода), чтобы хищники подросли, а вместе с ними выросла и трофическая, или пищевая, пирамида (греч. τροφή – пища). Основу такой пирамиды составляют продуценты – бактерии, водоросли и растения, которые создают органическое вещество из неорганических соединений и элементов. Те, кто питается непосредственно продуцентами, являются консументами первого порядка (от лат. consumo – съедать, потреблять); эти растительноядные животные занимают следующий ярус пирамиды. Их в свою очередь потребляют консументы второго порядка – хищники. Последние способны занять несколько ярусов пищевой постройки, ибо «более крупные едят более мелких». Поскольку биомасса продуцентов всегда больше таковой консументов первого порядка и т. д. – по всем ярусам, и получается устойчивая пирамидальная конструкция. Особую прочность ей добавляют как раз хищники высшего уровня: без их влияния на хищников низшего и среднего звена продуцентам грозило бы выедание и пирамиду ждала бы участь зиккурата – Вавилонской башни.
В эдиакарских морях вся пищевая цепочка начиналась с осмотрофов (реже – фильтраторов) и ими же заканчивалась, т. е. состояла из одного звена. Кембрийские хищники продолжили ее строительство, а аномалокаридиды и их донные родственники – головохоботные черви той же длины – обосновались на вершине. Однако их относительно небольшой максимальный размер – в пределах метра – не позволял создать достаточно длинную и прочную трофическую цепочку. Кроме того, если нынешние плотоядные животные специализированны (разборчивы в еде), а их отношения выстроены в виде жесткой иерархической лестницы (кому и чьей пищей стать, строго регулируется), то их кембрийские предшественники ели все подряд и не особо церемонились друг с другом. (Ископаемые с набитыми желудками, а также колалиты и копролиты являются явным тому свидетельством.) В результате пирамиду то и дело приходилось «подновлять и ремонтировать». В реальном мире этот процесс выразился в частых вымираниях кембрийских организмов (порядка семи эпизодов массовой гибели, один из которых сравним по масштабам с крупнейшими вымираниями фанерозойского эона), в формировании черных сланцев из избыточных, непотребленных остатков продуцентов и в резких зигзагах изотопной подписи углерода.
Хотя в ордовикском периоде аномалокаридиды еще были на плаву (последние представители этой группы, так и не ставшие членистоногими, вымрут в девонском периоде), с вершины этой постройки они были низвергнуты и стали весьма редкими видами. Их по размерам и в проворстве обогнали настоящие членистоногие – ракоскорпионы – и наутилоидные головоногие моллюски (Nautiloidea), достигавшие длины 2 и 8 м соответственно. (Ныне от ракоскорпионов остались только их измельчавшие сухопутные потомки, а от наутилоидных головоногих – два-три вида, обитающих в глубинах океана.) Конечно, одних размеров для «смены власти» было недостаточно – всегда необходимо опираться на массы: в ордовикском периоде доля кембрийских хищников (примитивные членистоногие, аномалокаридиды, головохоботные черви) в общем разнообразии этой трофической группы стремительно падает до 25 %, а остальные 75 % как раз начинают составлять хелицеровые (в том числе ракоскорпионы), многощетинковые черви, головоногие и брюхоногие моллюски, некоторые иглокожие, конодонтофориды и другие позвоночные.
Ракоскорпионы, или эуриптериды (Eurypterida), будучи прямыми предками наземных скорпионов, были вооружены клешневидными педипальпами. В отличие от сухопутных потомков, хватательными у них были и передняя пара конечностей – хелицеры, и первая пара ходных ног, а судя по весловидной последней паре конечностей, они хорошо плавали. Эуриптериды, вероятно, вели начало от кембрийских агласпидид через мечехвостов, единственную группу морских хелицеровых, уцелевшую до наших дней, и часматаспидид (Chasmataspidida). По строению тела часматаспидиды больше напоминали ракоскорпионов, но конечности у них были как у мечехвостов. Мечехвосты, прожившие долгую эволюционную жизнь, конечно, тоже не могли не измениться: так, раннесилурийские представители этой группы еще сохраняли двуветвистые конечности, включая хелицеры, подобно древним членистоногим.
Головоногие моллюски в самом конце кембрийского периода произошли от моноплакофор или брюхоногих моллюсков (улиток) с высокой колпачковой раковиной, разделенной поперечными перегородками на камеры-отсеки. Предковым формам эти перегородки понадобились, чтобы отделить вышедшую из употребления часть раковины, куда выросшее тело уже было не втиснуть, от жилой (рис. 22.1). Головоногие слегка переоснастили пустующие отсеки: «протянули» сквозь них сифон – орган, позволявший наполнять и опустошать камеры гипоосмотической жидкостью (по сути, это морская вода, но сильно опресненная, что позволяло создавать перепад давлений между сифоном и камерами) (рис. 22.1б). Из балласта раковина превратилась в поплавок, который позволял первым головоногим воспарить над дном. Плавающий «дом» был изнутри выстлан перламутром – прочным арагонитовым слоем, что позволяло облегчить скелет, не теряя при этом в надежности. С появлением еще одного новшества – воронки, в которую превратилась часть прежней ноги, они обрели способность двигаться на реактивной тяге и со временем стали активными хищниками. Поначалу довольно медлительными, поскольку прямая, слегка согнутая или свернутая в неплотную спираль раковина не позволяла развивать скорости, доступные их современным потомкам – кальмарам и каракатицам. При движении такую раковину сильно болтало и заносило, поэтому понадобились дополнительные известковые отложения, чтобы придать ей остойчивость. Но оптимизировать параметры раковины, чтобы перемещаться быстро и по траектории, близкой к прямой, на первых порах не удавалось. Да и достаточно мощной мускулатуры для управления воронкой (и реактивной струей) у них, судя по мускульным отпечаткам на внутренней поверхности раковины, еще не было. Такие мускулы появились только у девонских бактритов (Bactritoidea) и их потомков – аммонитов (Ammonoidea). Впрочем, не было и необходимости плавать быстро: очень медленно перемещался весь мир. Судя по следам укусов, которые древние наутилоиды оставили клювовидными челюстями на глазных покровах трилобитов, охотились они вполне успешно.

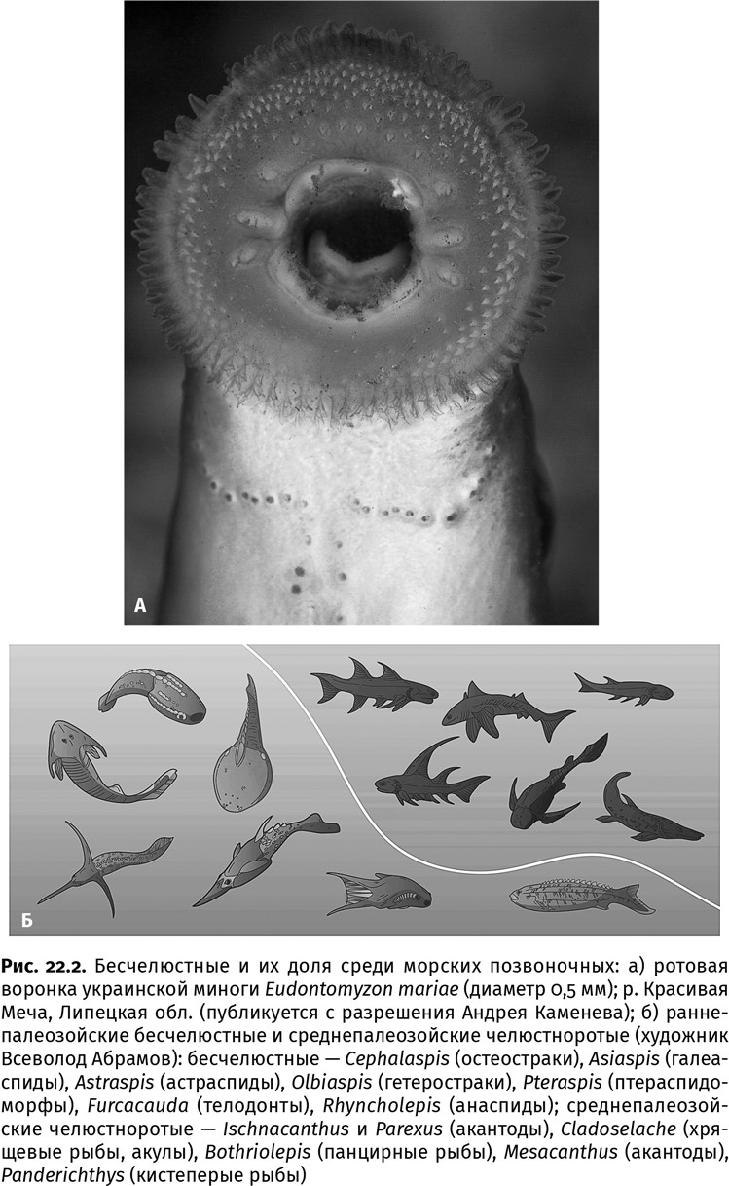
Самыми свирепыми хищниками ордовикских морей все еще оставались конодонтофориды. Недостаток роста они компенсировали высокой скоростью (мощные блоки из поперечно-полосатых мышц), способностью увидеть добычу издалека (крупные камерные глаза) и стайным образом жизни (что видно по большим скоплениям зубов). К хищникам высшего уровня подтягивались и другие позвоночные, в частности птераспидоморфы (Pteraspidomorpha) и телодонты (Thelodonti) (рис. 22.2). Голова этих рыбообразных бесчелюстных несла трехслойный костный панцирь, туловище – крупные кожные пластины с дентиновыми бугорками, а в пасти находились острые зубные пластины. У телодонтов все тело покрывала крупная толстая чешуя, а в глотке в несколько рядов сидели зубы, по виду и расположению напоминавшие акульи; в отличие от конодонтофорид и птераспидоморф, у них появились парные грудные плавники. Значит, эти бесчелюстные могли лучше маневрировать в водной толще. Вполне возможно, что какие-то из этих бесчелюстных «рыб» питались конодонтофоридами (или они сами – друг другом), поскольку сантиметровые копролиты или, скорее, комки отрыжки – регургиталиты (греч. ρήγνϋμι – тошнить и λίθος – камень), набитые конодонтами, в ордовикских слоях не редкость. Современные бесчелюстные – миксины – именно так поедают рыбу, переваривая мягкие ткани и отрыгивая кости. [Позднее, в девонском периоде – около 390 млн лет назад, среди позвоночных произойдет переворот, и вместо разнообразных бесчелюстных, из которых до наших дней доживут лишь миноги да миксины, в морях воцарятся челюстноротые – пластинокожие рыбы и акулы, достигавшие шестиметровой длины (рис. 22.2б).]
Новый виток «гонки вооружений» привел к появлению еще более изощренных способов и средств защиты у потенциальных жертв. Самым простым способом избежать попадания в чью-то пасть была способность стать крупнее этой пасти. По сравнению с кембрийскими предками-предшественниками самые беззащитные донные животные – брахиоподы, брюхоногие моллюски – в среднем выросли в размерах вдвое (с 1 до 2 см), а двустворки – вчетверо (с 0,5 до 2 см) (рис. 22.3), а максимальная величина представителей всей морской фауны в целом увеличилась на два порядка и еще на порядок – к середине девонского периода (в течение 140 млн лет). Интересно, что минимальные размеры в ордовикском периоде, наоборот, на порядок уменьшились. Эти размерные тенденции опять же отражают два основных способа ухода от давления хищников – в большую или меньшую размерную категорию.

Особенно увеличение размерности ордовикских скелетных животных по отношению к раннекембрийским проявляется в большей мощности ракушняков (в среднем с 0,1 до 0,2 м), возросшей доле скелетного материала в известняках (в среднем с 5 до 15 %) и повышении объемов карбонатонакопления в целом (с 10–20 % до 50–80 % от общей мощности разреза). Именно раковины вышеназванных групп и их обломки составили основную массу этого материала.
Поскольку подросли и рифостроящие животные – обызвествленные губки и кораллы, мшанки, мощнее и шире стали рифы (рис. 20.2в). Полуметровые в поперечнике строматопороидеи, похожие на слоистые караваи, и хететиды, чьи скелеты состояли из тонких вертикальных трубочек, пришли на смену вымершим археоциатам, чей диаметр редко превышал 1 см. Как и археоциаты, строматопороидеи и хететиды произошли от обыкновенных губок с кремневыми спикулами, но за ненадобностью либо утратили спикульный скелет, либо сохранили его как легкий каркас для наращивания известковых слоев. А вместо примитивных кембрийских коралломорф появились табуляты со сложными колониями, в основном одиночные, но крупные ругозы, или четырехлучевые кораллы, и еще несколько небольших групп скелетных книдарий. У табулят было по 12 щупалец, у ругоз – разное число, но кратное четырем; у современных кораллов количество этих хватательных органов кратно шести (шестилучевые, или склерактинии) или восьми (восьмилучевые).
При возросших размерах можно было усилить и броню – увеличить толщину раковины, либо наращивая слои одинакового строения, либо за счет добавления новых слоев с иной, более устойчивой к механическим повреждениям микроструктурой. Именно в это время и возникает настоящий перламутр – самая прочная арагонитовая микроструктура (исходя из давления, которое нужно приложить к раковине определенной толщины, чтобы она растрескалась, см. табл. 1). Примечательно, что перламутр независимо возникает в разных группах головоногих, двустворчатых и брюхоногих моллюсков. Их кембрийские предшественники обходились раковинами, образованными не столь надежной призматической микроструктурой или даже рыхлой пластинчато-фиброзной; нередко снаружи створки были покрыты толстым слабоминерализованным слоем органического вещества. Подобные плохо защищенные формы исчезли уже к началу ордовикского периода.
Когда и этих ухищрений оказывалось недостаточно, упрочить раковину можно было с помощью ребер жесткости. Ребристые раковины независимо возникают во многих группах брахиопод и брюхоногих моллюсков. Брахиоподы именно с такой раковиной составляют более половины разнообразия видов этой группы в ордовикских слоях. В меньшей степени ребристость использовалась двустворками и ростроконхиями, которые просто прятались в грунт, выставив оттуда лишь сифоны. (Из всех брахиопод вести скрытный в грунте образ жизни в ордовикском периоде приспособились лишь лингулиды с фосфатной раковиной, и у них до наших дней сохранились гладкие створки.) Ростроконхии (Rostroconchia) – вымершая группа моллюсков с раковиной, внешне похожей на двустворчатую, но развивавшейся из одностворчатого личиночного колпачка. Если у них конструкция раковины была жесткая, то настоящие двустворки достигали плотного смыкания створок благодаря зубному аппарату (система супротивных выступов и ямок на левой и правой створке) и мощным мускулам-замыкателям. Раскрывалась такая раковина при ослаблении мускулов благодаря эластичной связке – лигаменту, которая располагалась в ее макушечной части. Кроме того, воспрепятствовать тому, чтобы хищник без особых усилий мог вытащить живое содержимое из «домика», призваны были суженные устья и плотные завитки раковин у брюхоногих моллюсков и гофрированный передний край створок у брахиопод. C этого времени доля гладких форм в разнообразии моллюсков и брахиопод начинает сокращаться, а среди улиток все меньше становится раковин с открытым устьем и неплотно свернутых.
Дополнительную степень защиты раковине придавали шипы. Эти жесткие колючие выступы опять же увеличивали размер всего животного и могли серьезно ранить хищника. В кембрийском периоде колючими были только виваксииды, сахитиды, их сидячие родственники – ханцеллорииды и некоторые ксенузии, небольшие трилобиты и двустворчатые членистоногие. В ордовикском шипы выросли у очень многих донных и плавающих животных, включая трилобитов, иглокожих, брахиопод и колониальных гемихордовых – граптолитов (рис. 19.3б). Трилобиты, чтобы использовать шипы для защиты более целенаправленно, стали сворачиваться в плотные клубки. При этом шипы, расположенные у них на краях головного и хвостового щитов, а также туловищных сегментов, т. е. в плоскости панциря, «вставали дыбом», как у ежа (сравнивать можно с любым – и морским, и наземным).
Впервые этот способ защиты трилобиты опробовали во второй половине кембрийского периода, но очень немногие (менее 10 % видов), в ордовикском этот нехитрый маневр для спасения от хищника стали использовать уже 17 % видов, в силурийском – 35, а в девонском – 40 %. Чтобы колючий шарик был плотнее, на противоположных краях головного и хвостового щита у трилобитов появились специальные выступы и желобки.
Уберечься от хищников, в том числе биотурбаторов, можно было на твердом дне – хардграунде (англ. hardground). В отличие от естественного скального грунта, твердое дно стало своего рода «рукотворным» образованием. Основными его создателями были иглокожие. Их скелеты – теки (греч. θήκη – вместилище, шкатулка) состоят из множества Mg-кальцитовых пластинок и, быстро распадаясь после смерти животных, образуют обильный известковый песок (рис. 21.4). Поскольку эта разность карбоната кальция легко растворяется, то отдельные пластинки, утрачивая Mg2+-ионы, превращаются в частицы устойчивого кальцита, а свободные катионы реагируют с бикарбонатом, плотно заполняя кальцитом все свободное пространство в россыпи пластинок и преобразуя подвижный осадок в твердое дно. На плотном грунте поселяются новые, еще более многочисленные иглокожие, чьи скелеты тоже со временем распадаются. Этот саморегулирующийся процесс мог длиться десятки тысяч лет.
Массовыми обитателями твердого дна, кроме иглокожих, стали мшанки, брахиоподы и трубчатые родственники этих щупальцевых – корнулитиды (Cornulitida), прятавшиеся в мелких изогнутых известковых рожках, а также известковые водоросли, некоторые граптолиты и сидячие медузы с пирамидальным фосфатным скелетом – конуляты (Conulata). (Даже на мягком грунте эти существа искали твердую опору, сотнями облепляя обломки раковин наутилоидов и панцирей трилобитов.) Основное же население составили иглокожие, создавшие огромное разнообразие прикрепленных форм – на длинных и коротких стебельках или небольших каблучках прирастания. Из всего этого царства стебельчатых фильтраторов, достигавших разнообразия в сотни родов, до наших дней дожили лишь реликтовые морские лилии, и те прячутся в глубинах океана. В отличие от кембрийских предков, ордовикские иглокожие не лежали на дне, а плотно прирастали к нему, буквально прикипали стебельками (рис. 21.4). И если в раннекембрийскую эпоху этот тип животных был представлен исключительно свободно лежащими обитателями мягких илов, то все новые группы, появившиеся в ордовикском периоде, были приспособлены к жизни исключительно на твердом грунте. А сообщество мягкого субстрата практически изничтожили биотурбаторы, индекс активности которых в ордовикском периоде увеличился с 1–2-го до 3–4-го уровня: они могли перелопатить от 10–40 % до 40–60 % всех новообразованных осадков. Это означает, что слоистые отложения из-за постоянной переработки превращались в пятнистые, с едва различимой первичной слоистостью. Однако до нынешнего 6-го уровня им было еще очень далеко.
В сообществе твердого дна очень типичны были вторичные обрастатели. Они селились не на дне, а на раковинах и теках других организмов, причем часто еще живых. Некоторые морские лилии предвосхитили образ жизни современных лиан: они обвивали стебельками своих родственниц и так приподнимались над субстратом. А конуляты, мшанки и корнулитиды предпочитали всю жизнь проводить на раковинах других существ, положив начало еще одной экологической группировке – обрастателей. Корнулитиды благодаря небольшим размерам умещались между ребрами на раковинах брахиопод или между руками морских лилий, откуда их было вообще не достать. Иногда мелкие обрастатели (губки, брахиоподы, мшанки, корнулитиды) могли облюбовать раковину живой улитки – так формировалось сообщество твердого дна, перемещавшееся вместе с хозяином раковины-субстрата.
Чтобы отодрать сросшихся с твердым дном скелетных животных, хищникам приходилось прилагать гораздо больше усилий, чем при схватывании животного той же размерности на поверхности мягкого ила. Для этого офиоцистиоидеи (Ophiocistioidea) и морские ежи обзавелись мощными зубами, хелицеровые – клешневидными конечностями, а головоногие моллюски – щупальцами с присосками и крепкими клювами. Возросло и число хищников-сверлильщиков: им было где развернуться среди недвижимых обитателей твердого дна. Но об их деятельности приходится в основном судить по количеству продырявленных раковин других существ, а также по размеру дырок – некоторые пробоины достигали 6 см в диаметре. Здесь же зародилась еще одна экологическая группировка – биоэродеры (от греч. βιοτή – образ жизни и лат. e-rodo – разъедать), которые скребли и сверлили именно твердое дно чаще в поисках пищи, иногда – укрытия. Отчасти следы биоэрозии можно отождествить с работой губок, двустворок и морских ежей, но точно установить принадлежность большинства ископаемых биоэродеров крайне сложно.
Продуценты – водоросли – тоже не оставались неизменными: в кембрийском периоде среди них преобладали простые мелкие трубчатые формы, в ордовикском-силурийском на их место пришли крупные ветвистые виды, более устойчивые к выеданию, а затем и обызвествленные водоросли с прочным скелетом.

