Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 20 «Детонаторы кембрийского взрыва»
Дальше: Глава 22 Строители трофических пирамид
Глава 21
Изучение морских осадков с полным погружением
Роль фильтраторов и биотурбаторов в последующих преобразованиях океана нужно отметить особо. Фильтраторы значительно усилили потребление взвешенной органики. Ведь в отличие от осмотрофов у них есть особые органы, которые ускоряют потребление взвеси: жгутиковые камеры – у губок, мерцательные щупальца – у брахиопод, мшанок и их древних родственников (томмотиид, хиолитов), сифоны – у двустворчатых и некоторых других моллюсков, ветвистые конечности – у ряда членистоногих и аномалокаридид, разветвленные щупики – у сидячих кольчатых червей, амбулакральные щупальца – у прикрепленных иглокожих, жаберный аппарат – у гемихордовых и примитивных хордовых. Правильнее, конечно, разделять собственно тонких фильтраторов и суспензиефагов (от лат. suspensus – парящий, висящий и греч. φάγος – обжора): первые питаются растворенным органическим веществом и взвесью размером с бактерию (0,2–20 мкм) – пико- и нанопланктоном (губки, некоторые брахиоподы), вторые – более крупной добычей – одноклеточными эукариотами, личинками и мелкими беспозвоночными.
К суспензиефагам, например, принадлежали уже не раз упоминавшиеся томмотииды и хиолиты. Обе группы пережили свой расцвет в раннекембрийскую эпоху, но если томмотииды вскоре исчезли, то редкие хиолиты дотянули до пермского периода. Хотя скелеты этих организмов совершенно непохожи (трубки, состоявшие из многочисленных фосфатных склеритов-колпачков, – у томмотиид и известковые конические раковины с крышечкой и двумя тонкими изогнутыми подпорками – у хиолитов), они были родственниками. И внутри прикрепленной к грунту многостворчатой трубки, и в свободно лежащей конической раковине размещался сходный набор органов – U-образно свернутый кишечник и подвижный петлевидный или крыловидный орган, несущий питающие щупальца, покрытые мерцательными ресничками для сбора пищевых частиц. Конечно, для томмотиид строение внутренних органов приходится реконструировать, исходя из их близкого родства с брахиоподами, о котором можно судить по близкому набору микроструктур раковины и наличию осязательных щетинок, торчавших из пор в скелете. Предполагается, что многостворчатые томмотииды были предками двустворчатых брахиопод. От хиолитов иногда остается гораздо больше, в том числе фосфатизированный слепок пищеварительной системы, размещавшейся в конической раковине, и отпечаток крыловидного органа со щупальцами, который располагался прямо под крышечкой и соединялся с глоткой. Кривые скелетные подпорки, также находившиеся в передней части тела, вероятно, помогали разворачиваться по течению, так, чтобы пища сама плыла в рот, и даже ползать. Простенькие, почти прямые следы хиолитов, в конце которых может лежать раковина, вовсе не редкость. И сами хиолиты были настолько обычны в кембрийских морях, что участки ископаемого дна в несколько десятков квадратных метров выглядят буквально вымощенными их раковинами, причем не в один слой, а в несколько (рис. 18.1), а многие хищники набивали желудки этими неповоротливыми созданиями. В ордовикском периоде место примитивных щупальцевых заняли их более высокоорганизованные родственники – брахиоподы и мшанки.
Кембрийские донные фильтраторы, перечисленные выше, превосходили эдиакарских осмотрофов по размерности частиц, которые могли пройти сквозь их фильтрационные сита, и по темпам потребления органического вещества: 90–350 ммоль С/м2 в день, что свойственно современным губкам, против 5–50 ммоль С/м2 в день, исходя из возможностей осмотрофных бактерий (С – углерод, а м2 – площадь фильтрационного сита). Иными словами, за день фильтраторы в среднем потребляют столько же пищи, сколько осмотрофы – за неделю. Растворенное органическое вещество усваивается, входит в состав клеток, которые позднее отмирают и таким образом превращаются в относительно крупные органические частицы, пригодные в пищу суспензиефагам и детритофагам (илоедам). Первые захватывают органические частицы в водной толще, вторые – уже осевшие на дно (детрит). Кроме того, фильтраторы, даже губки, способны формировать из непереваренных остатков, например водорослевых оболочек, покрытые слизью комочки 15–55 мкм величиной, увеличивая тем самым размер пищевых частиц на один или два порядка. Насколько эффективны могут быть обыкновенные губки, можно судить по экосистеме озера Байкал, чистота воды в котором в значительной степени поддерживается этими существами.
И все же кембрийские фильтраторы сильно уступали в активности своим современным собратьям по гильдии. Все они – губки, щупальцевые и прикрепленные иглокожие – фильтруют довольно медленно, являясь пассивными потребителями, в большей степени зависящими от силы и направления внешних течений, чем от работоспособности своих органов. А предки самых активных фильтраторов современности – двустворчатых моллюсков – были еще слишком малы (менее 5 мм). Много ли воды может прокачать через себя такое существо? Разнообразием и обилием они тоже не отличались. Возможно, именно поэтому придонные воды еще не были насыщены кислородом, и любой подъем уровня моря, выносивший на шельф сероводородные воды, приводил к вымиранию организмов на обширных территориях в течение всего кембрийского периода. Об этом же свидетельствуют и мощные отложения черных сланцев с высоким содержанием неокисленного органического вещества, и все еще высокое фоновое содержание фосфата, и особенности изотопной подписи углерода и серы: положительные сдвиги δ13С (до +6‰) и δ34S (до +30‰), связанные с захоронением больших объемов отмершей органики и пирита в бескислородных обстановках (рис. 21.1). (Поскольку органическое вещество при фотосинтезе и при восстановлении сульфата обогащается легкими изотопами этих элементов, в океане, соответственно, остается относительно больше тяжелых, что и выявляет изотопная подпись.) Данные по изотопам серы показывают, что темпы ее поступления в осадок в 50–70 раз могли превосходить скорость осаждения этого элемента в Черном море.

Однако за счет периодических вымираний сообщества постоянно обновлялись, что также положительно повлияло на рост разнообразия организмов. Кроме того, частые заморы тоже способствовали эволюции отдельных групп. Чтобы дышать в почти бескислородных условиях, у трилобитов, брадориид и других членистоногих, обитавших в такой среде, были развитые жаберные ветви конечностей, сильно уплощенное тело и панцирь, пронизанный обширной сетью сосудов (у трилобитов, к примеру, плотность подобной сети в несколько раз превышала таковую у современных мечехвостов). Все эти усовершенствования позволяли быстрее вентилировать органы и получать кислород там, где этот газ присутствовал в следовых количествах.
Очищение верхних слоев океана происходило быстрее. Скелетные и бесскелетные животные начали осваивать толщу воды, и первопроходцами в этом пока еще не изведанном мире оказались мелкие членистоногие – мезозоопланктон (0,2–5 мм величиной), – возникшие к концу раннекембрийской эпохи. Их двуветвистые конечности с жаберной наружной ветвью, усаженной частыми, длинными, перистыми щетинками толщиной несколько мкм, позволяли и дышать, и парить в водной толще, и выцеживать из воды пищевые частицы. То были предки ракообразных, и за счет дальнейшего усложнения придатков, приспособления ножек к разнообразным действиям с пищей и их специализации эти членистоногие в конце концов (к середине кембрийского периода) превратились в настоящих ракообразных (рис. 21.2). Если у раннекембрийских членистоногих особым строением отличается только первая пара конечностей, то у современных высших раков, например у омара, все пары различаются по строению и по функции (и в совокупности этот рак может совершать ими десятки операций). Самые важные эволюционные преобразования конечностей происходили во второй половине кембрийского периода, и в первую очередь у планктонных форм. На каком-то этапе эволюции у планктонных членистоногих должна была возникнуть перитрофная мембрана – особая органическая оболочка, в которую, как в пакеты с мусором, упаковываются собственные фекалии. Это новшество помогло и самим древним ракам – они больше не фильтровали свои же отходы, и всему океану – появился направленный поток органического вещества в придонную область (пеллетный конвейер). Ведь тяжелые упаковки – пеллеты – стремительно идут ко дну, а фекалии, как им и положено, всплывают. Если погружение частиц глинистой или бактериальной размерности занимает годы (0,03–0,05 м в сутки), то пеллеты даже дна океанических впадин достигают за считаные дни – со скоростью 1000 м в сутки. Поскольку речь идет об органическом веществе, то при быстром погружении не вся его масса успевает разложиться бактериями и, следовательно, погребается без окисления, что и требуется для насыщения океана кислородом.

Конечно, сами пеллеты из-за их малости и неустойчивости сохраняются в древних отложениях редко, но есть геохимические признаки, по которым можно установить, когда включился пеллетный конвейер: в керогенах второй половины раннекембрийской эпохи возрастает доля водорослевых углеводородов – алгенанов. Это как раз и означает, что водорослевые остатки не успевали полностью разложиться, пока опускались на дно. К отложениям этого же времени приурочены и находки первых ископаемых пеллет, набитых акритархами, которые могли принадлежать планктонным членистоногим. Скопления пеллет, превратившиеся в обычное явление к концу раннекембрийской эпохи, становились затравкой для образования фосфатных линз, как и предполагал Бакленд. Именно в этих линзах и сформировались лагерштетты типа Эрстен со слепками мезозоопланктонных ракообразных и их современников.
Тогда же к плавающим фильтраторам присоединились даже некоторые аномалокаридиды: пользуясь своим господством в океане, они, подобно усатым китам, урезали длину пищевой цепочки – перешли на питание зоопланктоном, т. е. древними ракообразными. Вместо грозных хватательных предротовых придатков у таких аномалокаридид появился свой «китовый ус» – очень часто расположенные, длинные и тонкие щетинки, которыми можно было выцеживать мелкоразмерную пищу.
Одновременно пеллетный конвейер ускорил переброску органического вещества в глубины океана, где животные сидели на голодном пайке. С этого времени донный осадок начал перемешиваться илоедными животными, или детритофагами. Так возникли одни из самых мощных биотурбаторов. Конечно, биотурбаторами были все организмы, которые с той или иной целью ворошили морской грунт, – те, кто в нем прятался, затаивался, охотился, строил ходы, чтобы фильтровать или разводить съедобные грибы и бактерии, но илоедам эта среда сама по себе служила основным источником пищи. Всю жизнь они заглатывали ил и песок и пропускали все это сквозь кишечный тракт, чтобы извлечь оттуда отмершую органику вместе с питавшимися ею бактериями и одноклеточными грибами. Благодаря такому наполнителю нередко сохраняются органы пищеварения в остатках трилобитов и других донных членистоногих. Одни животные всасывали осадок, ползая по поверхности, другие отдались этому делу с головой, погрузившись в мягкий ил и выбираясь оттуда разве что для спаривания и откладки икры. Из-за постоянного изъятия из грунта органического вещества меньше пищи оставалось на долю сульфатвосстанавливающих бактериальных сообществ. Да и насыщение донного осадка кислородом за счет постоянного перемешивания слоев тоже не сулило этим микробам ничего хорошего. Восстановители сульфата должны были уходить все глубже в грунт, и образование фосфоритов и ранних доломитов на границе водной среды и осадка прекратилось (рис. 13.2а, в, ж, з).
Эти события выявляются по двум признакам. Во-первых, начинает расти индекс биотурбации, хотя и немного – до первого или второго уровня (это означает, что первичные осадки перерабатывались не более чем на 10 %). Для эдиакарских отложений индекс биотурбации равнялся нулю: все слои и слойки оставались лежать ровнехонько с момента образования осадка до времени его отвердения, превращения в горную породу. В кембрийском периоде ненарушенными оставались лишь относительно глубоководные отложения, где кислорода еще недоставало, чтобы биотурбаторы смогли приступить к работе. Все прочие первичные поверхности осадконакопления в той или иной степени были покрыты различными следами (рис. 21.3а, б). Но пока еще биотурбаторы как бы делали только первую прикидку: очень редко там, где «наследили» одни животные, по их следам проходили другие, до конца вычищая грунт. Кроме того, кембрийские биотурбаторы не зарывались глубоко (обычно не более чем на 0,03 м) и не образовывали многоярусные «подземные» поселения, где каждый вид обитает в пределах своего яруса в зависимости от способа питания и устойчивости к среде с пониженным содержанием кислорода (рис. 13.2з). Поэтому отдельные кембрийские слои все еще хорошо различимы, хотя их кровля и подошва (верхняя и нижняя поверхность соответственно) утрачивают четкие очертания (рис. 19.10б). Эти седиментологические признаки и соответствуют индексу биотурбации 2-го уровня при средней глубине проникновения 0,002 м (сейчас 0,1 м). Во-вторых, серосодержащие биомаркеры – один из важнейших показателей активности сульфатвосстанавливающих бактерий – становятся менее обильными. Одновременно повышается концентрация ионов SO42–, что определяется по росту показателя Δ34S и частоте изменения значений δ34S (∂δ34S/∂t). Содержание сульфата в морской воде по сравнению с эдиакарским уровнем повысилось примерно в три-четыре раза (с 1–4 × 10–3 ммоль/1 кг до 3–15 × 10–3 ммоль/1 кг; современное – 28,8 × 10–3 ммоль/1 кг) (рис. 13.2в).
Впервые морской осадок был потревожен около 550 млн лет назад: по поверхности дна проползли выедатели биопленок, оставившие очень простые, почти прямые следы, такие как гельминтопсис (Helminthopsis) (рис. 21.3а). Необязательно, что они представляли собой многоклеточные организмы. Но уже 541 млн назад – с этого момента и отсчитывается начало кембрийского периода – «днокопатели» стали проникать в грунт, прокладывая субгоризонтальные, подобные фикодесу (Phycodes), и вертикальные, например сколитос (Skolithos), тоннели. 535 млн лет назад впервые наследили своими многочисленными конечностями членистоногие – русофикус (Rusophycus), диплихнитес (Diplichnites), появились и «фермы» по разведению бактерий – сквамодиктион (Squamodictyon) (рис. 21.3б, в). Максимальный диаметр ходов за эти 15 млн лет возрос с менее 1 до 3 см, а средняя глубина проникновения в грунт составила от 0 до 3 см.


Деятельность биотурбаторов сказалась не только на живущих в грунте микробах: резко сократились площади биопленок и биоматов, для развития которых необходима ненарушенная, стабильная поверхность. Лишь на ограниченных участках дна они продолжали существовать, приютив небольшое число кембрийских животных. Вместе с биопленками окончательно пропали и жившие на них и за их счет вендобионты (их разнообразие и обилие сокращалось в последние 10 млн лет эдиакарского периода по мере усиления биотурбации). Со временем в очень сложном положении оказались и животные, которые либо свободно лежали на поверхности осадка, либо заякоривались в нем основанием скелета (или мягкого тела). Из-за превращения стабильной среды обитания в подвижную одни из них оказывались закопаны, а другие повалены. Лишь немногие из этих животных имели органы, помогавшие восстановить положение или хотя бы выбраться из завала. Такой характер влияния биотурбаторов на образ жизни малоподвижных и сидячих животных получил в научной литературе название «эффект бульдозера», понятное и без перевода. Учитывая, что в раннекембрийскую эпоху более половины донных организмов вели «обломовский» образ жизни, «эффект бульдозера» привел к исчезновению некоторых групп. Биотурбаторы не охотились на них, те просто оказались не в том месте не в то время. Скорее всего, именно поэтому вымерли анабаритиды и некоторые другие существа, строившие тонкие трубчатые скелетики, едва торчавшие из осадка. Другие сократили область своего распространения твердыми грунтами. Так поступили археоциаты и сидячие иглокожие: первые практически перестали возводить рифы на мягком субстрате, вторые даже дали начало новым донным поселениям – сообществам твердого грунта (рис. 21.4).

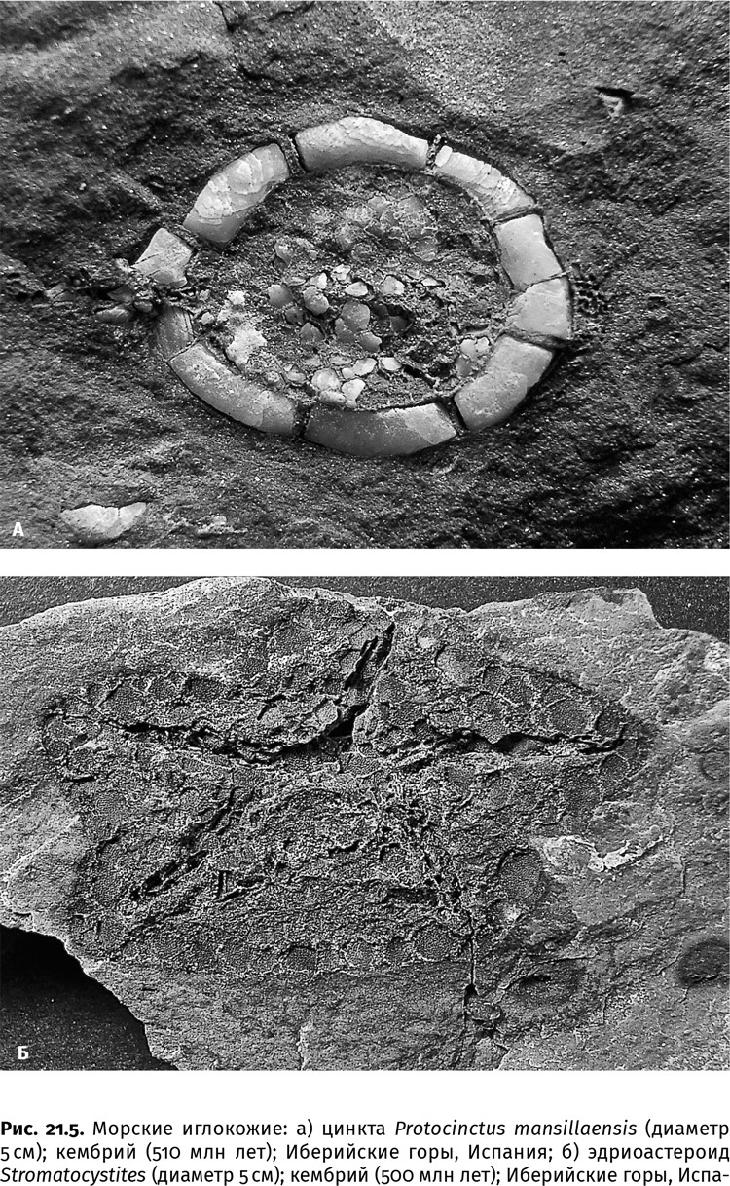
Многие (моллюски, брахиоподы, хиолиты), чтобы противостоять «эффекту бульдозера», наращивали размеры. Крупных животных и закопать было непросто, да и сами они могли выползти на поверхность. И если в раннекембрийскую эпоху до 90 % видов и особей в донных сообществах могли просто расположиться на мягком иле, то уже в среднекембрийскую эпоху их доля не превышала 60 % и продолжала сокращаться (рис. 21.5). Биотурбаторы не оставили в покое даже мертвых: в результате их распространения на внешнюю часть шельфа, в эти «святые» для науки места, морские лагерштетты, характерные для кембрийских слоев, исчезли; уже в ордовикских отложениях они встречаются крайне редко и приурочены к холодноводным обстановкам. Так случилось, потому что при рыхлении осадка в него поступают кислород и сульфат, что приостанавливает геохимические процессы, связанные с фосфатизацией и пиритизацией мягких тканей, с образованием глинистых слепков остатков животных.
Кривая разнообразия ископаемых следов – ихнофоссилий (от греч. Ϊχνος – след и лат. fossilis – вырытый), «авторы» которых в той или иной степени были биотурбаторами, повторяет кривую разнообразия скелетных животных (рис. 13.2е, ж). Конечно, в более сглаженном варианте, поскольку сидячие и малоподвижные организмы следов не оставляли, а подвижные могли следовать одним и тем же определенным поведенческим актам. Скажем, очень похожие двусторонне-симметричные дорожки с отпечатками конечностей – диплихнитес – могли оставлять трилобиты, ракообразные, а также многощетинковые черви и донные позвоночные с парными плавниками (рис. 21.6). Зато летопись ихнофоссилий неоценима для изучения эволюции поведения. Если во всех эдиакарских отложениях с трудом набирается от семи до девяти разных поведенческих актов, то уже в основании нижнекембрийского отдела их насчитывается более 40, а вскоре добавляется еще полтора десятка (рис. 13.2ж). Да, разносторонностью поведения раннекембрийские животные еще не отличались, но темпы роста разнообразия ихнофоссилий (а значит, поведенческих актов) были просто феноменальными. В истории Земли такое больше никогда не повторялось. Уход в грунт, конечно, тоже был одним из способов укрыться от хищников, но если бы основной задачей животных было спрятаться, вряд ли бы поведенческие акты оказались столь многообразны. Важную роль сыграл «исследовательский инстинкт»: многие типичные раннекембрийские следовые дорожки – например, фикодес, трептихнус (Treptichnus), олдхамия (Oldhamia) – это следы повторных проб грунта через равные промежутки в разных направлениях, т. е. поиск.


Значение биотурбаторов в преобразовании донных сообществ кембрийского периода было столь велико, что заслужило собственное имя (и не одно) – агрономическая или субстратная революция. В первом случае подчеркивается геохимический эффект биотурбаторов (прежде биоматы плотно запечатывали осадок, препятствуя доступу кислорода), во втором отмечается и механическая составляющая этого эффекта – перекапывание грунта.
Две экологические группировки животных – фильтраторы (донные и пелагические, населявшие толщу воды) и биотурбаторы – были не менее важными «детонаторами кембрийского взрыва», чем хищники. Но не стоит забывать, что именно хищники вынудили многих животных искать убежище в грунте и, таким образом, превратиться в биотурбаторов. А затем, начав охоту в новой среде, и сами хищники отчасти стали биотурбаторами. И все эти три группировки сделали океан, от поверхности до самого дна и даже несколько глубже, совершенно иным – более прозрачным, насыщенным кислородом и населенным всевозможными животными.

