Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 19 Спрятаться или убежать?
Дальше: Глава 21 Изучение морских осадков с полным погружением
Глава 20
«Детонаторы кембрийского взрыва»
Борьба за существование в самом прямом смысле этого классического выражения ускорила темпы эволюции. Так, в древнейших слоях кембрийского периода встречается всего несколько десятков видов ископаемых животных, а в отложениях, которые на 20 млн лет моложе, – свыше 1500. Сравнение темпов преобразования строения панциря у кембрийских и более поздних членистоногих (по 395 признакам) показывает, что в более древние времена внешний вид членистоногих менялся в четыре раза быстрее, чем в более поздние. Поскольку эти изменения привели к появлению разнообразных современных групп, оценивая распределение 62 ядерных генов, кодирующих белки в этих группах, можно определить и скорость молекулярных перестроек: получается, что у кембрийских членистоногих она в 5,5 раза превышала таковую их потомков.
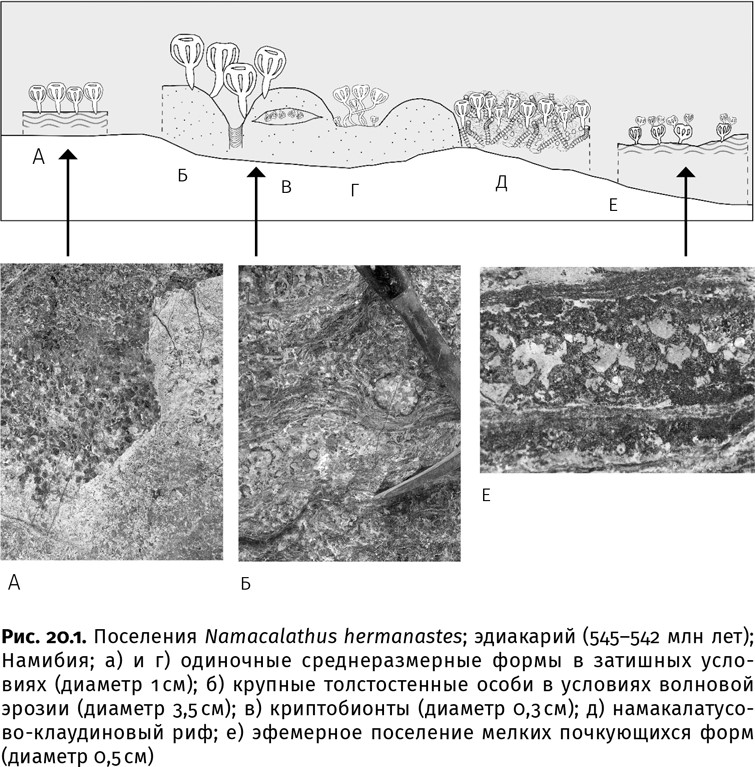
Стремительно росло не только общее число видов. Если сопоставить количество членов одного сообщества, а именно рифового (это, в общем-то, единственное сравнимое место обитания эдиакарских и кембрийских животных-фильтраторов), то эдиакарское сообщество изредка составляли особи трех видов, а обычно – только одного: либо клаудина, либо намакалатус. Правда, индивиды того же намакалатуса не были совершенно идентичны друг другу: на открытых подводных возвышенностях, омываемых сильными течениями, обитали крупные особи (до 35 мм в диаметре) с толстостенным шипастым скелетом; в рифовых полостях – небольшие гладкие формы (до 7 мм); в затишных впадинках, покрытых бактериальными пленками, с непостоянным уровнем аэрации – самые мелкие (до 5 мм) и тонкостенные, причем почковавшиеся (рис. 20.1). В том, что все пригодное для жизни пространство было заполнено генетически одинаковыми, но фенотипически (внешне) разными особями, нет ничего удивительного: свободного пространства с несколько различавшимися условиями было много, видов – мало.
Похожая история повторялась неоднократно. Скажем, 12 000 лет назад, когда ледниковый щит Северной Европы растаял, появилось много новых и необжитых озер. Первая занесенная туда птицами икра одного вида развивалась во взрослых рыб, которые осваивали все озеро и за несколько тысячелетий превращались в 2–4 формы, совершенно непохожие окраской, размерами, пищевыми предпочтениями и поведением. Так, в исландском озере Тингвадлаватн из арктического гольца, в отсутствие конкурентов и несмотря на возможность скрещивания любых разнополых особей, получились крупные рыбоядные хищники, плавающие в толще воды, небольшие придонные всеядные рыбки, мелкие обитатели лавовых расщелин и приповерхностные выедатели планктона.
В раннекембрийских рифах той же площади (1 м2), где в эдиакарском периоде жил один вид, обитали уже от 7 до 17 видов. (Существовали и поселения из одного-двух видов, но исключительно в неблагоприятных обстановках – при осолонении, на периодически пересыхающем мелководье, при частых извержениях близлежащих вулканов.) Подобно намакалатусу, это тоже были фильтраторы, но другие – археоциатовые губки, почти вымершие к концу раннекембрийской эпохи (рис. 20.2а, б). Причем разные виды этих губок играли в сообществах строго определенные роли: одни могли поселиться на мягком подвижном субстрате и, разрастаясь за счет почкования, построить небольшой жесткий каркас и стабилизировать грунт, на котором закреплялись другие виды, а другие занимали полости в каркасе. Виды археоциат различались также по своим предпочтениям к местообитаниям с более сильными, турбулентными или слабыми, плавными течениями. Если же «сталкивались интересы» двух видов, претендовавших на сходную нишу, один старался вытеснить другой, обрастая конкурента и замуровывая его дополнительными скелетными слоями. Поскольку пористый скелет выполнял у археоциат функцию первичного фильтра, дальнейшая эволюция группы пошла по пути создания все более дробных ниш фильтрования за счет варьирования размеров и формы пор. Всего за 20 млн лет десяток первичных видов с просто устроенным скелетом превратился в сотни сложно организованных фильтраторов.
Сходные процессы происходили и в других кембрийских сообществах со всеми группами организмов. В итоге за первые 20–25 млн лет кембрийского периода выросло среднее разнообразие видов внутри сообществ, но в гораздо большей степени увеличилось многообразие самих сообществ. Причем общий рост числа кембрийских видов происходил в большей степени за счет второго из этих факторов – в течение раннекембрийской эпохи морские животные осваивали все новые и новые местообитания и, распределяя между собой функции все более дробным образом, создали гигантскую мозаику сообществ, каждое из которых состояло из относительно небольшого числа видов – в пределах 50. Позднее добавился третий фактор – изоляция отдельных мелководных бассейнов по мере удаления друг от друга континентов: в морях каждого из них – Сибири, Лаврентии, Восточной и Западной Гондваны – начали формироваться сообщества только из местных видов. Космополитных – повсеместно распространенных – видов в раннекембрийскую эпоху практически не существовало, чем это время разительно отличается от всех последующих эпох.


Побочным продуктом «гонки вооружений» и одновременно показателем ее ускорения служат ракушняки – известняки и фосфориты, сложенные в основном из раковин и их обломков, спаянных карбонатом или фосфатом, который выщелачивается из скелетов при перемыве и переотложении первичного материала. В эдиакарских отложениях ракушняки были большой редкостью, а их формирование, как в случае суворовеллового пласта, было обусловлено геохимическими особенностями среды. Кембрийские ракушняки накапливались именно благодаря существованию и отмиранию огромного числа скелетных животных (рис. 18.1). Конечно, мощность и протяженность таких слоев зависели от количества скелетных животных, их размеров и устойчивости раковин к растворению. Поэтому нижнекембрийские ракушняки имеют вид ограниченных по площади линз толщиной не более 0,15 м (обычно менее 0,1 м), но с появлением крупных трилобитов и брахиопод с кальцитовыми скелетами они быстро увеличиваются по мощности до полуметра и становятся одной из самых распространенных карбонатных пород. Растет и содержание в них скелетного материала: с 5 % и менее – в кембрийских отложениях, до 15 % – в ордовикских. Это также означает, что существенные потоки ионов кальция и бикарбоната в океане с этого времени стали регулироваться обменом веществ у животных, а изъятие Са2+, растворенного в океане, ускорилось. То же касается фосфат-иона: мелкие лингулидные брахиоподы с гидроксилапатитовой раковиной благодаря своей биомассе к концу кембрийского периода становятся своего рода накопителем фосфата. Значительные месторождения фосфоритов на территории современной Прибалтики целиком образовались из раковинок этих двустворчатых щупальцевых.
Какие же факторы «сдетонировали кембрийский взрыв» – молекулярно-генетические, экологические или абиотические? Все. Повышение уровня кислорода создало условия для появления крупных многоклеточных животных, в том числе хищников. Хищники вызвали «гонку вооружений». «Гонка вооружений» предопределила ускорение молекулярно-генетических перестроек организмов, выразившееся в появлении разнообразных скелетных и других структур, формировании и развитии нервной системы и совершенствовании поведенческих актов (рис. 13.2е, ж). Но ведь и аэрация – насыщение кислородом – водной толщи происходила благодаря животным – осмотрофам, которые положили начало этому процессу в эдиакарское время. В кембрийском периоде аэрации начали активно способствовать животные, представлявшие новые трофические гильдии, – фильтраторы, суспензиефаги и биотурбаторы.
Назад: Глава 19 Спрятаться или убежать?
Дальше: Глава 21 Изучение морских осадков с полным погружением

