Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 18 Кембрийские горы
Дальше: Глава 20 «Детонаторы кембрийского взрыва»
Глава 19
Спрятаться или убежать?
Итак, появление во второй половине эдиакарского периода крупных многоклеточных организмов, среди которых могли быть и настоящие животные, преобразило океан. Благодаря мягкотелым гигантам – вендобионтам, способным к осмотрофному питанию, содержание растворенного органического вещества понизилось; на мелководье стали зарождаться настоящие кислородные оазисы. Новую среду начали осваивать первые скелетные животные, потеснившие бактериальные рифообразующие сообщества в морях с карбонатным осадконакоплением, которые покрывали Сибирь, Южный Китай, монгольские и казахстанские микроконтиненты, отчасти Лаврентию и Гондвану. Тогда и случился кембрийский взрыв: в течение 25 млн лет общее разнообразие морских животных возросло в несколько раз, а биоминеральный скелет приобрели представители почти всех типов животных, которые существуют до настоящего времени (рис. 17.6). Формирование различного по составу скелета – кремневого, фосфатного, карбонатного (несколько разных минеральных форм) – было сопряжено как с абиотическими явлениями (тектонические циклы Уилсона, парциальное давление углекислого газа), о которых уже говорилось, так и с биотическими (образ жизни конкретных видов животных). Об этом речь пойдет далее.
Причины кембрийского взрыва разбираются в тысячах статей. Десятки, если не сотни предположений выдвигаются, чтобы объяснить это явление самым необычным сочетанием факторов. Например, тем, что расширение Земли способствовало снижению силы притяжения, позволив животным увеличить свою массу за счет скелетных наслоений. Или тем, что учащенная смена полярности магнитных полюсов (15–30 переполюсовок за 1 млн лет против 2–10 за тот же срок в фанерозойском эоне) ослабила магнитное поле планеты и ее мелководные обитатели оказались уязвимыми для ультрафиолетового излучения среднего диапазона (315–280 нм), проникавшего до глубины нескольких метров, для защиты от которого понадобились биоминеральные покровы. Однако вряд ли вендобионты, даже не самые большие, весили меньше самых крупных раннекембрийских моллюсков, а скелеты у многих древних организмов были внутренними – бесполезными для защиты от облучения. Да и решить проблему внешней защиты с помощью органических пигментов было энергетически дешевле, что цианобактерии и водоросли доказали еще в протерозойском эоне. В целом же все гипотезы о кембрийском взрыве можно разделить на три большие группы: молекулярно-генетические, экологические и абиотические. К последним относится и два примера, приведенных выше, но это отнюдь не значит, что все абиотические факторы в равной степени являются надуманными.
Наиболее наглядные свидетельства происходивших в то время реальных событий – это исчезновение ранних доломитов и широкое распространение красноцветных морских отложений (и терригенных, и известковых), уменьшение доли фосфоритов и снижение фонового уровня фосфата, взрывной рост разнообразия и обилия скелетных окаменелостей, появление многочисленных признаков хищничества, а также ископаемых следов, свидетельствующих о развитии сложных поведенческих актов. Все эти изменения указывают на повышение уровня содержания кислорода. Главный окислитель способствовал накоплению растворенного SO42–-иона, препятствовавшего осаждению доломита и фосфатных минералов (рис. 13.2а, в). А живым существам он был необходим для формирования коллагена – белка, без которого невозможно образование сложного, иерархически организованного органического (кутикула членистоногих и головохоботных червей) или биоминерального скелета, а также плотной соединительной ткани. Не случайно эдиакарские скелетные животные селились там, где воды были насыщены кислородом. Необходим кислород и для работы поперечно-полосатых мышц с плотной кровеносной сетью, переносившей обильные красные кровяные тельца, и другими клетками с повышенным количеством митохондрий. Без таких приспособлений невозможно существование активных хищников, которые преследуют добычу или рывком нападают на жертву из засады (из убежища в грунте). Если, например, сидячим фильтраторам, таким как губки, достаточно 0,1–0,8 % кислорода, растворенного в воде, то даже мягкотелым подвижным хищникам требуется на порядок больше (более 1 %). И чем выше уровень кислорода, тем хищники крупнее и разнообразнее.
Важнейшим фактором кембрийского взрыва и было хищничество. Уже магистральные тенденции в эволюции биоминеральных скелетов прямо указывают, что появление все более устойчивых к сдавливанию, скручиванию и растяжению защитных микроструктур и, в свою очередь, особо прочных микроструктур в органах, предназначенных для нападения, является несомненным свидетельством пресловутой «гонки вооружений» (табл. 1). Чем изощреннее становились защитные приспособления (панцири, раковины), тем во все более мощное и крепкое оружие превращались зубы, хватательные щетинки и разного рода «ногочелюсти». Становление многоуровневой защиты хорошо прослеживается на раковинах моллюсков: начиная буквально с первых «лет» кембрийского периода створки становятся не просто толще и многослойнее, но и сами слои приобретают все более и более прочную структуру. А у хищных членистоногих, которые стремились добраться до мягкого содержимого таких раковин, появляются все более сложные «орудия взлома».
Уже в середине раннекембрийской эпохи среди морских членистоногих процветает группа, названная Megacheira (от греч. μέγας – большой и χείρ – рука). «Большеруки» отличались очень развитыми передними конечностями, согнутыми в «локтях», на конце которых могли находиться либо клешни, либо жесткие шипы, либо длинные ловчие хлысты (рис. 19.1). Внешне эти придатки напоминали ногочелюсти современных раков-богомолов. Эти ракообразные, несмотря на небольшие размеры (до 35 см длиной), в стотысячные доли секунды прокалывают или дробят сопоставимую с ними по массе и более крупную добычу, обрушивая удар силой до 1500 Н. (Не каждый боксер-тяжеловес способен бить с такой силой, притом что масса рака в 100 раз меньше.) Различается только положение органов нападения: у появившихся в мезозойскую эру раков-богомолов это грудные ножки, а у мегахейр – передняя пара головного отдела, по положению соответствующая чувствительным усикам-антеннулам их отдаленных родственников. Одна из самых больших мегахейр, найденных в сланце Берджесс, за устрашающие клешнеподобные конечности с внушительными шипами получила имя кутенихела Деппа (Kootenichela deppi – от реки Кутеней, протекающей рядом с местонахождением, греч. χηλή – раздвоенный и в честь актера Джонни Деппа, сыгравшего заглавную роль в фильме «Эдвард Руки-ножницы»). Скопления проломленных панцирей трилобитов в кембрийских слоях могли появиться благодаря таким хищникам.

У других членистоногих – Artiopoda (греч. αρτί – πους – крепконогий) – сразу несколько пар конечностей головного отдела преобразились в клещевидные органы захвата, покрытые многочисленными зазубренными шипами. Особенно мощные пилообразные структуры располагались у артиопод под головой, ближе к основанию конечностей, где за счет мускульных рычагов можно было создать максимальное давление и тут же отправить раздробленную добычу в рот (рис. 19.2). В кишечнике таких животных обнаружены перемолотые раковины хиолитов и панцири трилобитов и мелких двустворчатых членистоногих – брадориид (Bradoriida, впервые найденные в кембрийских слоях полуострова Лабрадор). Агласпидиды (Aglaspidida, от греч. αγλαός – блестящий и ασπίς – щит) обзавелись мощными шипастыми клешнями, напоминавшими педипальпы (хватательные конечности) скорпионов и других хелицеровых, чьими предками они, возможно, были.

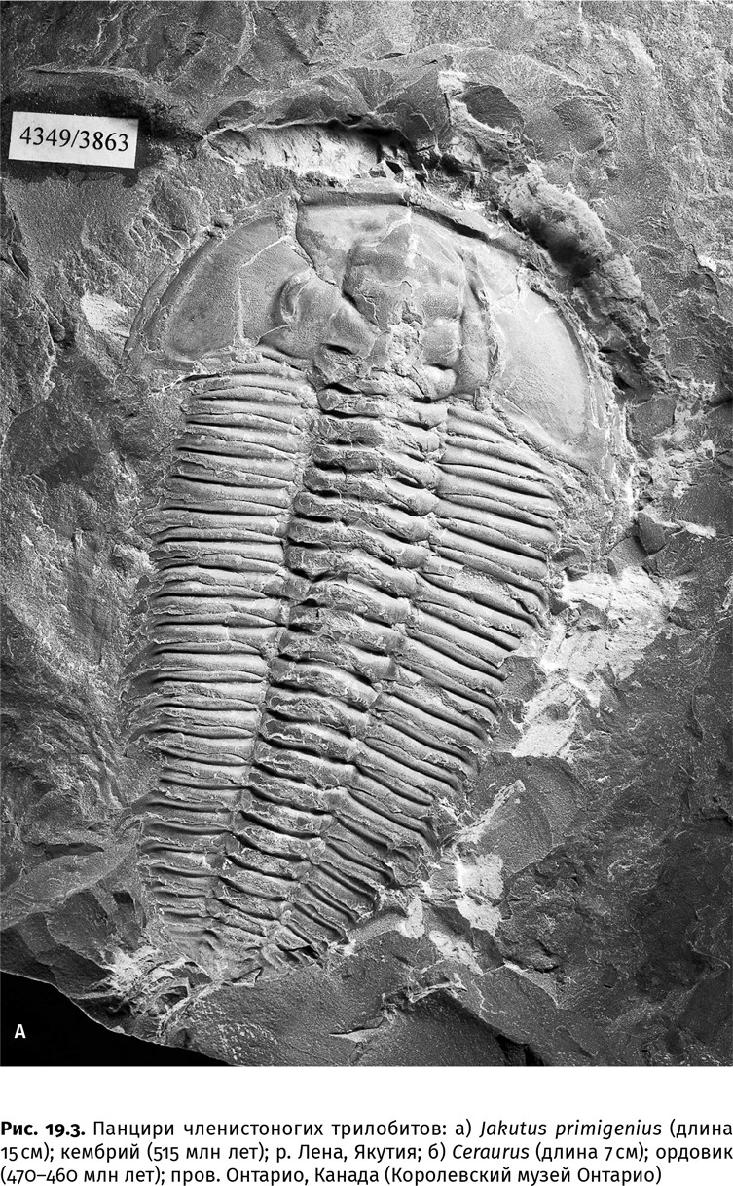
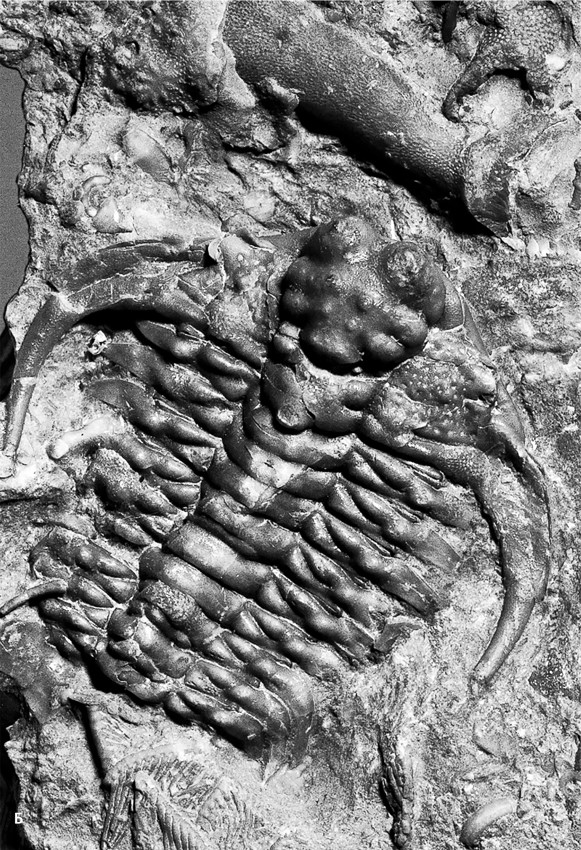
Даже трилобиты (Trilobita, от греч. τρεϊς – три и λοβός – лопасть) – самые распространенные кембрийские членистоногие, у которых все конечности, кроме одной пары антенн, были похожи на ходные ножки благодаря находящимся на них зазубренным жевательным выростам, обращенным навстречу друг другу, сминали в лепешку твердые покровы других животных (рис. 19.3). Их кишечники набиты расплющенными раковинами брахиопод, брадориид, даже спикулами – многолучевыми кремневыми иголками, из которых при жизни состоял скелет стеклянных губок. В том, что кишечный тракт нередко сохраняется, ничего удивительного нет: из-за избытка фосфора в пище, если животное погребено целиком, пищеварительные органы и все их содержимое обрастает фосфатными микрокристаллами. Нередко фосфатизированные кишечные тракты оказываются устойчивее других органов тела и сохраняются как отдельные окаменелости – колалиты (греч. χολάς – кишка и λίθος – камень). Хотя найти хозяина «каменной кишки» – трилобита, другое членистоногое или совсем иное существо – удается редко, можно понять, что большинство кембрийских колалитов принадлежало именно хищникам: они набиты полупереваренными остатками других животных. Фосфатизация сохраняет и пищеварительные железы – парные, обычно дольчатые мешочки, расположенные вдоль средней кишки и являющиеся ее выпячиваниями. Такие железы известны у трилобитов, мегахейр, артиопод и других кембрийских хищников (аномалокаридиды, головохоботные черви). Вероятно, железистые выделения помогали переваривать хитиновые покровы добычи: хотя хитин является полисахаридом, его спиральные, плотно уложенные волокна расщепляются плохо.
Нередко в ископаемой летописи встречаются и копролиты (греч. κόπρος – навоз, помет и λίθος – камень) – окаменевшие фекалии. И снова их содержимое указывает не только на отсутствие проблем с пищеварением у кембрийских животных, но и на то, что оставившие их особи были хищниками. Некоторые копролиты достигают 7 см в поперечнике или 20 см в длину и содержат сотни скелетиков хиолитов, брадориид, мелких трилобитов и других членистоногих. Удлиненные плавно-изогнутые копролиты, которые к тому же расположены вертикально в осадке, вероятно, принадлежат головохоботным червям, а округлые – трилобитам-каннибалам или аномалокаридидам.
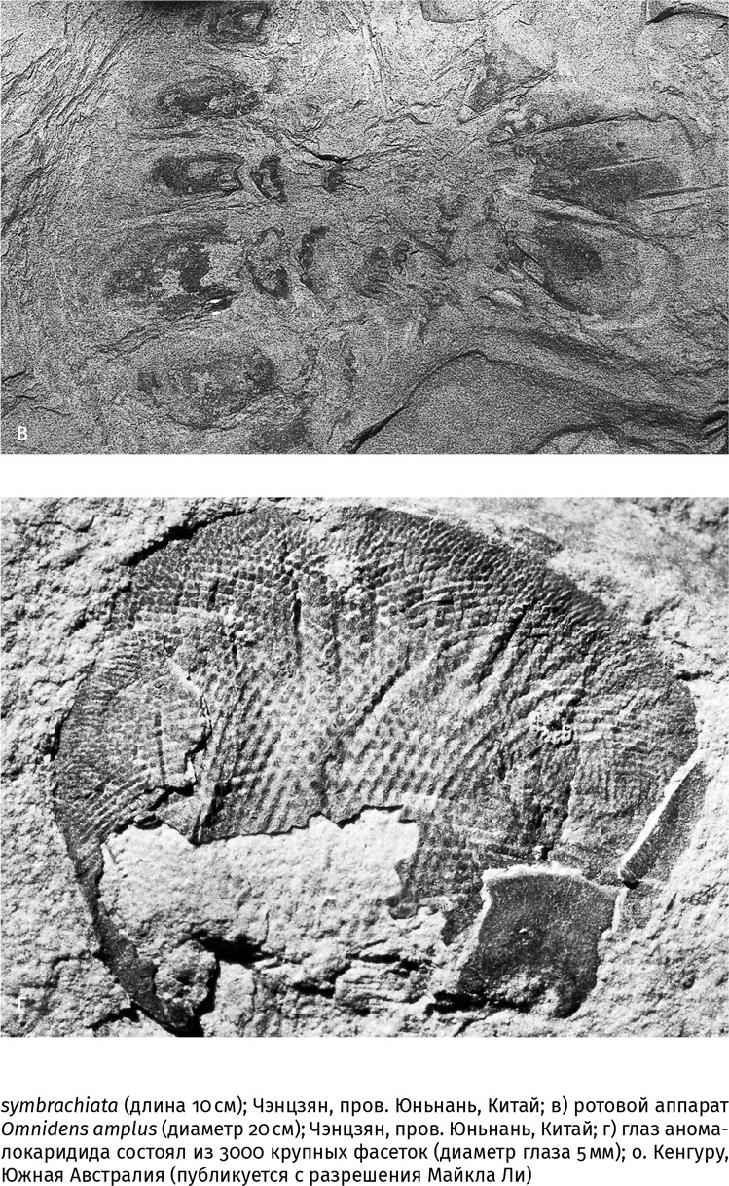
Аномалокаридиды (Anomalocaridida) были самыми большими хищниками и вообще животными кембрийского периода. Они достигали метровой длины и были вооружены двумя крупными членистыми околоротовыми хватательными придатками, за что и получили свое название (греч. ανωμαλία – несоразмерность и καρκίνος – рак). Хотя тело аномалокаридид тоже было членистым, а глаза – сложными, или фасеточными (состояли из многочисленных глазков – омматидиев), они не относились к членистоногим. Ведь других членистых конечностей аномалокаридиды не имели, а их ротовой аппарат представлял собой систему пластинчатых зубов, расположенных кольцом (рис. 19.4). И зубы, и шипастые околоротовые придатки, и огромные стебельчатые глаза (до 3 см в поперечнике) также выдают хищную природу гигантов. 3000 крупных омматидиев, в каждом из которых прекрасно выражена шестигранная линза, и размер линз указывают на то, что аномалокаридиды предпочитали охотиться в сумерках и видели даже при тусклом освещении. Вообще, глаза не хуже содержимого желудка выдают пищевые предпочтения своего хозяина: в раннекембрийском морском сообществе местонахождения Чэнцзян в Южном Китае (520 млн лет), откуда описано более сотни видов животных, среди нехищных видов более 5 % обладали глазами, а среди хищников – свыше 50 %. И все кембрийские членистоногие хищники – мегахейры, артиоподы, трилобиты и другие – были весьма глазасты.

Учитывая аномальные размеры аномалокаридид, им приписываются не только самые большие копролиты, но и следы укусов на панцирях крупных трилобитов. Конечно, подобные отметины сохранялись только тогда, когда потенциальной жертве удавалось вырваться, а затем залечить поврежденную часть панциря. Везло тем, кто успевал вовремя заметить опасность и увернуться, лишившись только части хвоста. Важнее другой вывод «палеокриминалистов», основанный на статистической обработке находок: более чем в 60 % случаев пострадавшие лишались части хвостового щита – пигидия или туловищных сегментов с правой стороны. Это свидетельствует об асимметрии в поведении хищника: у него был достаточно развитый мозг, а левая и правая его половины работали независимо друг от друга (как полушария мозга у млекопитающих).
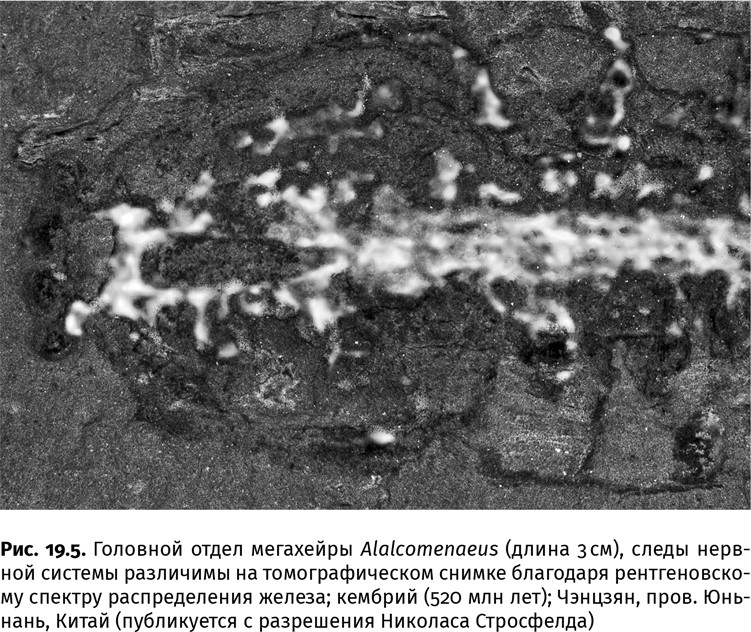
Сложный мозг аномалокаридид и некоторых хищных кембрийских членистоногих иногда тоже можно увидеть (с помощью рентгеновской спектроскопии): если органы пищеварения подвергаются избирательной фосфатизации, то нервная система, правда очень редко, замещается железосодержащими глинистыми минералами, покрывающими остатки органического вещества (рис. 19.5). Этот орган состоял из трех пар крупных нервных узлов: передняя пара иннервировала предротовые хватательные придатки, средняя – большие стебельчатые глаза, а задняя отвечала за пищеварение, движение плавательных лопастей и другие процессы, связанные с органами туловища. Как и глаза, крупный, по кембрийским меркам, мозг принадлежал только хищникам.
Впрочем, у некоторых из них этот орган представлял собой всего лишь окологлоточное нервное кольцо, поскольку, если хватательный орган, составляющий треть тела, то и дело вворачивается внутрь и выворачивается обратно, иначе нервные узлы не расположить. Этот хватательный орган, усаженный гребенчатыми крючьями – глоточными зубами, называется интроверт, или хоботок, а его обладатель – головохоботным червем. Хищные головохоботные черви существуют и сегодня (приапулиды, лорициферы, киноринхи), но они малочисленны и по большей части мелки (несколько миллиметров длиной), а в кембрийских морях составляли существенную часть донных сообществ и размерами могли сравниться с аномалокаридидами. Их уплощенные глинистые отпечатки, трехмерные фосфатные слепки и оставленные ими норки в изобилии встречаются во всех крупных кембрийских местонахождениях окаменелостей (рис. 19.6). Именно головохоботным червям принадлежат большие удлиненные копролиты, набитые раковинками хиолитов. На целых окаменелостях можно в подробностях изучить и содержимое желудка. Головохоботные черви на диете явно не сидели: в их последней трапезе есть место не только хиолитам, но и брахиоподам, трилобитам, агностоидам (Agnostoida, от греч. αγνοέω – не знать: безглазые ракообразные, похожие на трилобитов), брадориидам, виваксиидам и кольчатым червям – любым сидячим и медленно передвигавшимся животным, до которых можно было дотянуться крючковатым хоботком, не вылезая из норки. Сообща они могли съесть и крупное (более 10 см длиной) членистоногое, но, скорее всего, падаль. Даже у этих огромных и скрытных хищников жизнь не была легкой: встречаются копролиты, переполненные фрагментами их шкурок, а по следам охоты трилобитов можно понять, что крупные особи этих членистоногих не боялись вступать в схватку с червями, не уступавшими им в размерах.


Если в эдиакарских отложениях или на окаменелостях той поры не встречается никаких следов хищничества, то кембрийские слои буквально переполнены и остатками самих хищников, и свидетельствами их нападений. К последним наряду с копролитами, колалитами и отметинами зубов относятся следы сверления – небольшие (менее 0,1 мм) сквозные отверстия с ровными краями в раковинах. Конечно, дырки могут появиться и при посмертном переносе створок, когда те трутся друг о друга, песок или камни, но тогда они и расположены случайным образом и правильных очертаний не имеют. Наоборот, следы сверления привязаны к наиболее уязвимым участкам на раковине жертвы – там, где защитные слои тоньше, а сам участок открыт для нападения. Например, у брахиопод-акротретид (Acrotretida) с фосфатной раковиной 95 % пробоин приходится на коническую брюшную створку, а у лингулид (Lingulida) с такой же по составу раковиной – 87 % на плоскую спинную. Кроме того, даже самые изощренные хищники-сверлильщики не всегда успешны: смертельная сквозная рана всегда окружена лунками той же величины – это следы неудачных попыток проникнуть внутрь. Жертвами кембрийских сверлильщиков, кроме брахиопод, были хиолиты, двустворчатые моллюски и улитки. Все эти животные обитали на поверхности осадка, могли целиком спрятаться в раковину и сомкнуть створки или плотно прикрыть вход в нее крышечкой. Однако правило «я в домике» срабатывало далеко не всегда. Кто были кембрийские сверлильщики, установить сложно. Сейчас так охотятся разные брюхоногие и головоногие моллюски, используя острые зубы – радулу – или особый выступ на своей раковине и выделяя сильные органические кислоты, а также круглые черви, протыкающие панцирь жертвы полыми стилетами для того, чтобы впрыснуть секреты пищеварительных желез и высосать уже переваренные ткани.
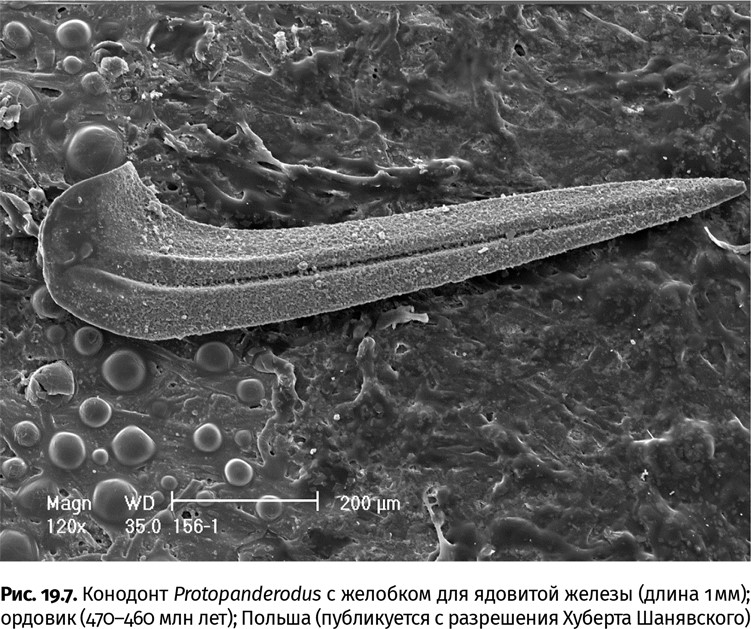
Выходит, от многочисленных хищников никто не мог считать себя защищенным: ни те, кто жил на поверхности дна, ни обитатели грунта, ни те, кто ползал или бегал, ни те, кто плавал. По дну рыскали всевозможные членистоногие с хватательными конечностями, под обманчиво гладкой поверхностью ила затаивались головохоботные черви, в толще воды (пелагиали) добычу поджидали огромные аномалокаридиды, многочисленные щетинкочелюстные и гребневики. Позднее, в среднекембрийскую эпоху, к когорте пелагических хищников присоединились и зубастые позвоночные – конодонтофориды. Конодонт и означает «конический зуб» (лат. conus и dens). Зуб, конечно, был не один: в глотке животного находился сложный аппарат из 7–9 пар (и один непарный элемент) конических, гребенчатых и пластинчатых фосфатных зубов (0,1–2 мм высотой), которые плотно смыкались, дробя угодившую в пасть добычу. По сколам на зубах, типичным для хищников, будь то тираннозавр или лев, и удалось определить пищевые предпочтения конодонтофорид. Тем более что целиком они сохраняются исключительно редко. Находки отпечатков этих животных с большими камерными глазами, V-образными туловищными мускульными блоками и хвостовым плавником подтвердили их принадлежность и к стремительным хищникам, и к позвоночным, эволюция которых как скелетных организмов началась с зубного аппарата. В зубах различается три слоя – эмаль, дентин и кость, также характерные для позвоночных. Не исключено, что некоторые виды конодонтофорид имели ядовитые железы, о чем, опять же, можно судить по зубам с отчетливой ложбинкой – каналом, проводившим яд от железы к укушенной жертве (рис. 19.7). Обладатели таких приспособлений, вероятно, могли обездвиживать добычу, превосходившую их собственные размеры (до 40 см длиной; в кембрийском периоде, правда, на порядок мельче).
Жертвы, чтобы избежать такой участи, не только наращивали биоминеральную защиту, которая к тому же не всегда помогает против опытных сверлильщиков и взломщиков. Одной из важных стратегий было бегство. Если в первой половине кембрийского периода сидячие, прикрепленные и малоподвижные, свободно лежащие животные составляли более половины населения морей, то в середине периода 70 % видов на месте никогда уже не оставалось – ползали, ходили, плавали. Во второй половине кембрийского периода даже некоторые неподвижные организмы научились «ходить». Например, ребристые брахиоподы-нисусииды прикреплялись к шипам ползающих примитивных моллюсков виваксиид, тоже ребристым, и таким образом сразу обретали и защиту, и покровительственную «окраску», и средство передвижения, а еще и доступ к новым источникам органической взвеси, которой они питались.
Многие приобрели защитную окраску или в случае необходимости прятались в укрытие. Хотя покровные пигменты животных той поры неизвестны, не только они дают представление о том, как внешне выглядело то или иное животное. Панцири и чешуйки у многих мелких членистоногих, моллюсков и кольчатых червей были испещрены тончайшими параллельными ребрышками: под преломленными в водной среде лучами солнца этот микрорельеф срабатывал как дифракционная решетка, превращая своих хозяев в радужно-яркие, но расплывчатые, мельтешащие пятна непонятного размера. А именно оценка размера и направления движения – основные параметры, по которым хищник определяет, куда направляется жертва и по зубам ли она ему вообще.
Членистоногие, головохоботные черви и аномалокаридиды, даже самые большие и хищные из них, на определенных стадиях жизненного цикла, чтобы расти дальше, должны были менять жесткий внешний панцирь – линять. В этот момент они становились очень уязвимыми и пережидали в убежище, пока новый панцирь затвердеет – вновь пропитается карбонатом или фосфатом. Иногда там их накрывал подводный оползень, и сотни миллионов лет спустя палеонтологи находят целые стаи одноразмерных трилобитов и других членистоногих, когда-то спрятавшихся в больших пустых раковинах хиолитов или в чьих-то норках. (Одноразмерными эти существа были, чтобы те, кто побольше, не съел тех, кто поменьше, воспользовавшись удобным случаем.) Коллективная синхронная смена покровов, о которой свидетельствуют массовые захоронения линочных шкурок членистоногих и головохоботных червей, тоже была выходом из почти безвыходной ситуации: не все хищники отваживаются напасть на большие скопления животных, даже если легко расправляются с одиночными особями того же вида.
Многие животные прятались на протяжении всей своей жизни. Брахиоподы, например, обживались среди минеральных спикул-иголок крупных губок, которым они явно отдавали предпочтение (50–60 % особей в поселении). А к концу кембрийского периода появились первые отшельники – животные, использовавшие чужие раковины в качестве переносного блиндажа. Сейчас тема отшельничества целиком принадлежит десятиногим высшим ракам. Во времена, о которых идет речь в этой главе, даже их предки еще на свет не появились, но по характерным следам тонких членистых ножек, с одного края перекрытых ребристыми отпечатками волочившейся раковины, можно определить, что жилье было выбрано на «вторичном рынке»: никакие членистоногие раковин-башенок не строят.
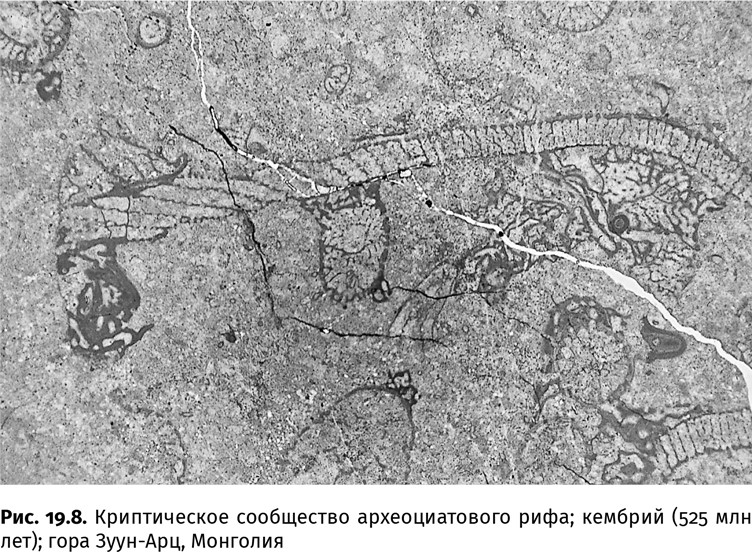
В рифах можно было обойтись и без чужой раковины. Некоторые существа на всю жизнь забивались в полости и даже небольшие трещины. То были криптобионты (от греч. κρύπτω – скрывать и βιοτή – образ жизни). Кембрийскими криптобионтами стали мелкие археоциаты и кораллоподобные создания, гирляндами свисавшие со свода; моллюски, ползавшие по мягкому известковому илу; какие-то червеобразные организмы, рывшиеся в нем; а также губки, сверлившие стенки полостей, чтобы укрыться в убежище внутри убежища (рис. 19.8). Если в начале раннекембрийской эпохи посещения полостей были случайными и в них селились те же виды, что и на открытой поверхности, то спустя несколько миллионов лет там уже появились постояльцы – виды, которые за пределами схронов никогда не встречались. Настоящие криптобионты. К таким постояльцам относится и арахностега (Arachnostega), точнее – тот, кто оставил эти мелкие (субмиллиметровые) ходы в осадке, наполнявшем опустевшие раковины хиолитов и даже перевернутые панцири трилобитов.
Если в кембрийских рифовых полостях обитали исключительно мелкие особи, то в мягком иле или подвижном песке на открытых пространствах морского дна могли поселиться животные любой размерности. Одни из них, например трилобиты, зарывались в грунт на время отдыха; их убежища распознаются по отпечаткам многочисленных членистых ножек. Засадные хищники – головохоботные черви – были прописаны там постоянно, выползая из U-образной норки, на стенках которой отчетливо пропечатывается рельефная поверхность червя, чтобы построить новую. Судя по способу передвижения таких червей за счет гидравлического нагнетания и перераспределения полостной жидкости в хоботке и туловище, они успешно продвигались в грунте, но были почти беспомощны на его поверхности. «Подковерную» жизнь освоили и некоторые моллюски: они оставляли длинные петляющие следовые дорожки с валиками по бокам и волнообразными отпечатками мускулистой ноги вдоль оси следа; по всей дорожке обычно тянется зигзагообразный валик – это грунт, прошедший сквозь кишечный тракт, где из него было извлечено все органическое вещество (рис. 19.9).

Подвижные известковые илы в спокойных обстановках, к примеру с подветренной стороны рифа, колонизировали какие-то членистоногие. Они устраивали вертикальные колодцы метровой глубины и 2–3 см в диаметре на близком расстоянии друг от друга и укрепляли стенки норок своими пеллетами – округлыми фекальными комочками, упакованными в особую мембрану, которая придавала прочность этому строительному материалу и стенкам норки. (Именно пеллеты и выявляют принадлежность хозяев норок к членистоногим.) Так грунт стабилизировался, а дополнительные горизонтальные туннели, соединявшие колодцы, превращали всю систему, занимавшую десятки и сотни квадратных метров, в гигантский фильтровальный аппарат, куда устремлялись потоки воды со всеми ее обитателями (рис. 19.10в). Членистоногие-фильтраторы могли даже не показывать и носа из своих убежищ, разве что антенны. Угадать хозяев всех разнообразных следов, появившихся в морском грунте кембрийского периода, очень сложно, хотя такие же следы существуют и в наши дни. Однако почти неотличимые ходы и дорожки оставляют очень разные животные. Так, вертикальные системы туннелей строят и креветки, и некоторые виды рыб, и морские ежи. Только то обстоятельство, что из всех этих животных в раннекембрийскую эпоху появились лишь членистоногие (но не креветки и вообще не ракообразные), позволяет отождествить колодцы с деятельностью последних.
Еще одним способом защиты была забота о своем потомстве – о самых маленьких и уязвимых. Брадорииды не откладывали икру, а вынашивали ее под панцирем, на специально приспособленных для ее прикрепления трех последних парах ходных конечностей. А у некоторых трилобитов на панцире обнаружены вздутия, очень похожие на выводковые сумки современных раков.



