Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 15 Эмбриональная стадия Земли
Дальше: Глава 17 Два мира – два скелета
Глава 16
Позднеэдиакарский перелом
Несмотря на преимущественно бескислородный – то закисный, то сероводородный – океан, жизнь в нем бурно развивалась. Об этом можно судить не только по фосфатным слепкам организмов из Южного Китая – одного из блоков, расположенных на северной периферии Гондваны. Там же, в формации Доушаньтуо, в самых верхних – черносланцевых – ее слоях встречаются многочисленные крупные сложноорганизованные бентосные (донные) водоросли, известные как биота Мяохе. Если венъянские бентосные многоклеточные водоросли – красные, то здесь представлены в основном зеленые и бурые. Всего в этой биоте известно порядка 20 разных видов. Примерно такое же разнообразие бентосных водорослей, но совершенно других, наблюдается в более древних (600–590 млн лет) черных сланцах Ланьтянь. Самые сложные из них напоминают членистые трубки, и поэтому их часто принимали за остатки древнейших червей. Однако, в отличие от червей, у ланьтяньских ископаемых был прикрепительный диск, а трубка состояла из водорослевых углеводородов. Учитывая, что росли они на дне в бескислородных условиях (Feа/ΣFe>0,38), то это были не просто водоросли, а хемотрофы или организмы, содержавшие хемотрофные (например, серные) бактерии-симбионты.
А в кремнистой нижней части формации Доушаньтуо обнаружены обильные акритархи. Причем, в отличие от более древних, раннеэдиакарские формы значительно крупнее: преобладают сферы 0,6 мм в диаметре против 0,2 мм. Усложняется и поверхность этих водорослевых оболочек, настолько, что среди таких акритарх палеонтологи пытаются распознать цисты животных. В целом в первой половине эдиакарского периода акритархи достигли максимальных размеров за всю свою историю и наибольшего разнообразия для протерозойской ее части. Вторая половина эдиакарского периода выдалась не столь удачной: разнообразие акритарх сократилось, проще стали оболочки, а средняя и максимальная размерность уменьшилась почти на порядок. Возможно, обеднение и упрощение фитопланктона было связано с оскудением фосфатного резервуара: все-таки и масштабы фосфоритонакопления, и фоновый уровень растворенного фосфата начали в то время понижаться (рис. 13.2а). Нуждавшиеся в фосфате бентосные и планктонные водоросли все чаще оказывались на голодном пайке. Или же крупные акритархи стали слишком лакомой добычей для новых одноклеточных врагов. Если потенциальный хищник намного больше жертвы, то оптимальной стратегией выживания являются мелкие размеры: преследование незначительной по размеру добычи энергетически себя не оправдывает (на мезо-кайнозойском этапе развития биосферы это правило скажется на максимальных размерах хищных динозавров и млекопитающих). Зато именно в это время начался расцвет огромных, в масштабах всей протерозойской эры, бентосных организмов, которых обычно именуют «эдиакарскими мягкотелыми», или вендобионтами. «Мягкотелыми» эти организмы можно признать только в том случае, если удастся доказать, что они были животными. А если нет? Скажем, всесветно распространенный белтанеллиформис (Beltanelliformis) дисковидной формы (0,2–9,2 см), которого долгое время считали медузой, сидячим полипом, в крайнем случае – прикрепительным диском других «мягкотелых» (рис. 5.2), оказался колонией бактерий… Выяснилось, что в его органической оболочке совсем нет стеранов – углеводородов, характерных для животных.
Именно к «мягкотелым» организмам принадлежат аспиделла с Восточного Ньюфаундленда, которую долго вообще отказывались принимать за окаменелость, и находки Реджиналда Спригга на Эдиакарских холмах, считавшиеся остатками животных, но кембрийских (рис. 16.1а). Лишь когда к Сприггу в Австралии присоединился большой коллектив палеонтологов и были заново изучены местонахождения Восточного Ньюфаундленда, Англии и Намибии, а советские палеонтологи Борис Сергеевич Соколов и Михаил Александрович Федонкин открыли богатейшие местонахождения подобных органических остатков на Зимнем берегу Белого моря и в украинской Подолии (позднее – на Урале и в Сибири), стало ясно, что вторая половина эдиакарского периода (578–542 млн лет) характеризовалась исключительным своеобразием в отличие от всего предыдущего протерозойского эона, а также от всего последующего фанерозойского. И своеобразие это заключалось в необычных организмах, существовавших только в то время и проживших около 35 млн лет.
Населяли же они моря всего мира: Восточный Ньюфаундленд и Англия – это кусочки небольшого древнего высокоширотного континента Авалона, Подолия и Беломорский регион (а также Урал) – два края высокоширотной Балтии, Сибирь – отдельный, целиком покрытый теплым мелким морем материк, запад Северной Америки – северный край Лаврентии, а Намибия (как территория Африки), Австралия, а также большая часть Западной Европы, Индия и Южная Америка – области южного суперматерика Гондваны, где-то вблизи северной границы которого располагались китайские блоки (Южный и Северный Китай, Тарим).

Поначалу эдиакарские «мягкотелые», достигавшие 2 м в длину или высоту, были описаны как медузы, моллюски, кольчецы, иглокожие, хотя похожи они на этих животных разве что на художественных реконструкциях. М. А. Федонкин был первым, кто обратил внимание на то, что многие из этих организмов имеют сходный тип сегментации – симметрию скользящего отражения (с полушаговым смещением условно левых сегментов относительно правых вдоль оси тела), не характерную для каких-либо многоклеточных животных (рис. 16.1, 16.2). (Такие формы часто и считают древнейшими червями и членистоногими.) Теперь эти псевдосегменты принято называть фрондлетами. Учитывая это важное отличие эдиакарских мягкотелых от настоящих многоклеточных животных и приуроченность их остатков к отложениям этого периода, немецкий палеонтолог Дольф Зейлахер назвал ископаемые формы со сложным рельефом, напоминающим рисунок стеганого одеяла, вендобионтами (Vendobionta). Это название, как более нейтральное, будет использоваться далее. Причем эдиакарские «медузы» оказались прикрепительными дисками более сложных лопастевидных вендобионтов, сидящих на стебельчатых основаниях.

Сходство вендобионтов между собой и их разница с многоклеточными животными определяются отсутствием ротового и анального отверстий, кишечника, каких-либо зооидов, щупалец или конечностей, а также неограниченным характером роста тела (как у растений или грибов). Причем новые фрондлеты закладывались асимметрично на одной, двух или даже нескольких конечных точках тела; в последнем случае вырастал веерообразный организм (и в этом вендобионты тоже напоминают растения и грибы). Но главное отличие проявляется во внутреннем строении – в пронизывающей все тело системе камер (иногда – каналов), что удалось выявить с помощью лазерного сканирования высокого разрешения и цифрового картирования. Эта система и придает поверхности вендобионтов облик стеганого одеяла. Такое же тонкое камерное строение наблюдается на трехмерных карбонатных слепках вендобионтов.
Камеры – открытые с одного, внешнего, конца, а вся камерная система напоминает фрактал: каждая из них расчленяется на более мелкие структуры, но строго определенным образом – структура второго порядка внешне подобна таковой первого порядка, только в меньшем масштабе, и т. д. (до пяти раз). Такие самоподобные фигуры обладают одним удивительным свойством: их реальная площадь значительно превышает топологическую. Внешний вид обманчив. Момент фрактального удвоения с образованием фрондлетов той же формы, но меньшего размера наступал, когда делящийся фрондлет дорастал до некоторой конечной величины, видимо опять же соответствовавшей определенному соотношению его площади и объема. Расчеты показывают, что фрактальное масштабирование увеличивало соотношение площади к объему в 1,4–3,7 раза, поддерживая его на уровне 1 мм2/1 мм3. Это на целых от одного до четырех порядков ниже, чем у современных гигантских бактерий (например, у названной выше тиомаргариты). Толщина живой ткани, где происходил обмен веществ, при этом могла быть меньше 0,1 мм.
Поскольку бактерии являются осмотрофами (питаются растворенной в водной среде органикой, всасывая ее всей поверхностью), исключительно высокое соотношение площади поверхности тела к его объему позволяло и вендобионтам использовать тот же ресурс. Содержание растворенного органического вещества в эдиакарском океане, очевидно, в два-три раза превышало нынешний уровень, что определяется по изотопной подписи углерода: для углерода в карбонатах эта подпись имеет размашистый росчерк, но при этом соотношение его изотопов в керогенах практически не меняется. Это и означает, что объемы углерода, содержавшегося во взвешенном органическом веществе, намного превышали таковые, зафиксированные в минералах. Да и деться всей этой органике было некуда: чтобы быстро утонуть в плотных слоях воды, частицы должны достигнуть определенной размерности – более 0,5 мм, и, даже попав на дно, они не могли быть сразу погребены в осадке – биотурбаторы (животные, способные перерабатывать осадок) еще не появились (от греч. βίος – жизнь, образ жизни и τυρβάζω – мешать, мутить). С первым же волнением воды частицы, с трудом осевшие за годы, опять всплывали мутным облаком.
Вся поверхность осадка, где жили вендобионты, на нескольких десятках квадратных метров может быть покрыта особями одного и того же вида (на 1 м2 от 30 до 150 и даже до 3000 экземпляров в случае аспиделлы). Причем это не единовременное скопление: сотни слоев осадка той же площади выглядят точно так же – все переполнено некогда живыми организмами. Такие очень плотные поселения, особенно если в них преобладали лопастевидные формы (от них и оставались прикрепительные диски – аспиделлы), создавали эффект «верхового потока» даже в местах со слабым течением. Верховой поток образуется, когда течение упирается в многоуровневую преграду и расслаивается. При этом между соседними слоями, перемещающимися с разной скоростью, возникают завихрения, которые ускоряют движение сопряженного слоя на верхней границе с препятствием. Препятствием и служили плотно росшие вендобионты. В этих условиях в выигрыше оказывались особи, чьи «макушки» достигали наибольшей высоты, если, конечно, они питались осмотрофно.
Биомеханические опыты с пластиковыми моделями лопастевидных стебельчатых вендобионтов в аквариумах показали, что «стеганая» поверхность направляла водный поток вдоль поверхности тела. Возникавшая при этом вибрация лопасти снижала нагрузку потока и одновременно усиливала водообмен с окружающей средой, способствуя осмотрофному питанию. А вот для фильтраторов вибрация – лишняя помеха, да и изолированные друг от друга фрондлеты тоже не способствовали развитию фильтровальной системы (для этого все тело должно быть пронизано системой каналов с закономерным размером сечения на входе и выходе водного потока). А трехлопастным, как некоторые вендобионты, фильтратор вообще быть не может: при таком строении образуются «мертвые зоны», куда течение не доходит.
В свою очередь, вендобионты, перемещавшиеся по дну (впервые такие формы появились около 558 млн лет назад), могли выедать – всасывать всей поверхностью – органическое вещество бактериальных пленок и биоматов: следы-отпечатки оставались именно в тех местах, где организм присасывался к поверхности биопленки. Биопленки могли служить не только источником пищи, но и кислорода: в современных малокислородных лагунах Венесуэлы насыщенность биоматов кислородом в дневное время в четыре раза превышает таковую водной среды. Фрактальная система камер могла помогать и в дыхании при низком уровне кислорода.
Отличались вендобионты не только своеобразным строением тела. Находки плотных поселений одного и того же вида на Восточном Ньюфаундленде, где эти организмы гибли под периодическими пеплопадами и сохранялись в прижизненном положении, выявляют у них необычный тип бесполого размножения. Достигнув определенной размерности (нескольких сантиметров), лежащий на дне вендобионт выпускал вокруг себя многочисленные столоны – трубочки, похожие на «усы-побеги», на которых начинали расти его точные копии, но меньшего размера. На этом развитие колонии не прекращалось: и материнский, и дочерние организмы продолжали увеличиваться за счет добавления новых фрондлетов, и в определенный момент, так и не обретя полной самостоятельности, отпрыски выбрасывали свои столоны, на кончиках которых начинали развиваться очередные копии «главы семейства». Иначе говоря, и колонии вендобионтов напоминали фрактальную структуру.
Следы передвижения эти организмы тоже оставляли очень необычные – дискретные, совершенно непохожие на следы ползания каких-либо животных (рис. 16.3). Но раз вендобионты перемещались, у них должен был развиться определенный аналог мускульной системы.

Все это свидетельствует об особом положении вендобионтов среди многоклеточных организмов. В свою очередь, распространение вендобионтов ниже фотической зоны океана – до глубины 1,5 км – исключает их водорослевую природу, а также наличие фотосимбионтов. Равно не имели они прямого отношения и к грибам или лишайникам, с которыми их иногда сравнивают, поскольку фрактальная система камер, характерная для вендобионтов, не имеет ничего общего с цилиндрическими гифами грибов.
Трудно сказать, были ли все вендобионты родственными организмами, т. е. представляли собой раннюю и полностью вымершую группу, близкую и к многоклеточным животным, и к грибам (т. е. к «заднежгутиковым», или Opisthokonta), но не являвшуюся предками ни тех ни других. Некоторые вендобионты, например кимберелла (Kimberella), несколько напоминающие моллюсков, судя по следам питания – глубоким веерообразно расходящимся царапинам на поверхности осадка, – скоблили пищу относительно твердыми зубами. Немного похожие следы оставляют своим зубным аппаратом – радулой – современные моллюски, когда соскребают бактериальные пленки (рис. 16.4). Другие вендобионты (Vendoconularia) – в форме перевернутой пирамидки – обладали шестилучевой симметрией и походили на некоторые древние сидячие медузоподобные организмы.
Не только вендобионты появились на свет и исчезли навсегда во второй половине эдиакарского периода. Еще одну группу составляют сегментированные лентовидные ископаемые, лежащие на поверхности осадка: ненокситес (Nenoxites), палеопасцихнус (Palaeopascichnus), орбизиана (Orbisiana), гаочжиашания (Gaojiashania) (рис. 16.5). Подобно вендобионтам, они распространены практически всесветно и первоначально были описаны как ископаемые следы. Однако ни одно животное не оставляет бессмысленных следовых дорожек: это всегда «продуманный» путь, проделанный либо ради поимки жертвы, либо для того, чтобы не стать оной, либо для того, чтобы выесть органическое вещество оптимальным образом, не возвращаясь на уже обследованный участок, но и ничего не пропуская. Эдиакарские «следы», напротив, петляют так, будто их оставили совершенно безголовые существа. Кроме того, они могли раздваиваться и увеличиваться в размерах – расширяться. Безголовые многоклеточные организмы существуют на самом деле. Только многоклеточными они становятся на определенных стадиях жизненного цикла, а до этого живут и питаются как одноклеточные амебы. Лишь когда все бактерии вокруг съедены (далеко ли уползет одиночная амеба?), они испускают особые химические сигналы. Все ближайшие амебы в ответ на этот молекулярный «зов» объединяются в многоклеточный грибовидный организм, способный пережить неблагоприятные времена и прорасти спорами (из которых развиваются новые амебы), или собираются в единый организм – слизевик (нечто напоминающее очень мелкого и безголового слизня), который может проложить дальний маршрут. Слизевик и оставляет петляющий след. Поскольку эта группа организмов – амебозои – является ближайшими родственниками опистоконтов, заманчиво предположить, что бессистемные эдиакарские петли – именно следы слизевиков. Тем более что подобные, но более простые следы появились уже 1,7 млрд лет назад: достаточно давно, чтобы слизевики, эволюционируя, дали начало грибам и многоклеточным животным. Но слизевики слишком мелкие и живут на суше, хотя и предпочитают влажную среду.


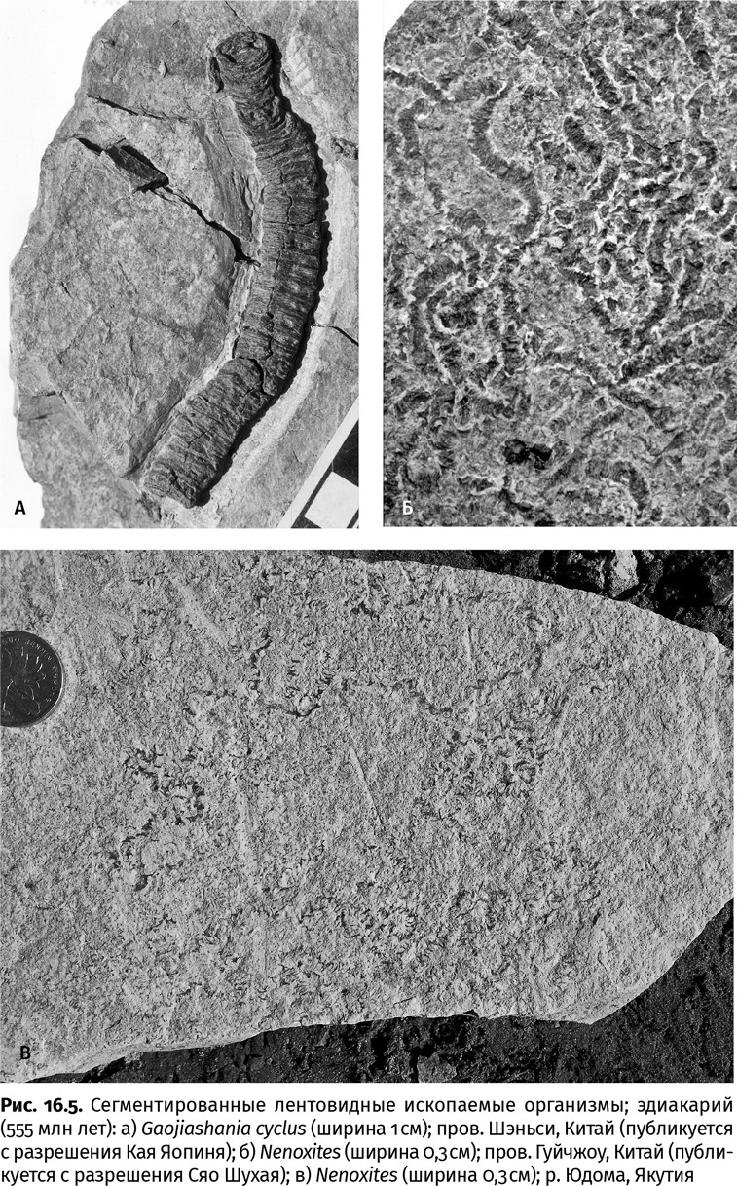
Есть предположение, что гаочжиашания и подобные ей образования при жизни вовсе не лежали на осадке, а колыхались в водной толще, словно водоросли. Очень длинные сегментированные формы действительно известны – это стебли древних морских лилий, достигавшие двухметровой длины. Но такой стебель – очень сложная конструкция, сравнимая по механике с подвесным мостом, и возвышается над субстратом за счет жестких сцепок отдельных элементов друг с другом при поддержке мускульных и нервных волокон. У гаочжиашании ничего подобного нет, ее остатки никогда не накладываются друг на друга в отличие от стеблей умерших морских лилий. Загадка эдиакарских лент остается пока нераспутанной, как и они сами.
Какова бы ни была природа эдиакарских организмов, в том числе вендобионтов и скелетных животных (о них рассказ впереди), жизнь их, как уже отмечалось, протекала в океане с повышенным содержанием растворенного органического вещества и, как следствие, при нехватке кислорода. Этот газ расходовался на окисление обильной органики, что приводило к частым заморам. О низких концентрациях кислорода, собственно, и свидетельствуют мощные доломиты и фосфориты, а также высокое соотношение Feа/ΣFe (>0,38), очень низкие показатели I/(Ca+Mg) и Zn/Fe, малые концентрации Cr, Mo, V, U и сульфатов и изотопные подписи (δ13С и δ34S) (рис. 13.2а – г, и – л). В некоторых морях, например на юге Гондваны (Намибии), вендобионты предпочитали селиться там, где уровень содержания кислорода был хоть немного выше, о чем можно судить по низким значениям соотношения Feа/ΣFe (менее 0,22) во вмещающих отложениях, а также по следам «марганцевого океана». Для формирования слоя с повышенным содержанием растворенного марганца требуется определенная, но низкая концентрация кислорода – субкислородная обстановка (около 0,1 % О2): при таких условиях Mn2+ окислялся до Mn4+, а Ce3+– до нерастворимого Ce4+. (Первичное содержание марганца обнаружить практически невозможно, и об обогащении среды этим элементом судят по отрицательной аномалии в содержании церия, который выводится из среды оксидом марганца – Mn4+О2.) Карбонат марганца не отлагается, поскольку в бескислородной зоне Mn2+ вновь восстанавливается до Mn4+. (Наблюдаются все эти явления опять же в Черном море.) Марганцевый слой служил барьером для сероводорода, не выпуская ядовитый газ наверх:
Mn4+О2 + HS– + НCO3– + 2H+ → Mn2+CO3 + S0 + 2H2О.
Наоборот, в морях Лаврентии (Западная Канада) разнообразные и крупные вендобионты существовали в закисной среде. Вероятно, уровень кислорода не был определяющим фактором в их жизни. Ничего странного в этом нет: митохондрии, для работы которых организм, собственно, и поглощает кислород, при синтезе АТФ создают протонный градиент на внутренней мембране за счет восстановления О2 до H2О. При необходимости они легко заменяют эту реакцию окислением H2S до тиосульфата (S2O32–). И такие митохондрии есть и у животных, и у грибов, т. е. у всех опистоконтов, к которым, несомненно, принадлежали и вендобионты.
Главное, что спокойной, практически растительной жизни эдиакарских организмов не мешали биотурбаторы и хищники. Как только на рубеже эдиакарского и кембрийского периодов влияние последних резко повысилось, а объемы растворенного органического вещества, наоборот, понизились, вендобионты и многие их современники полностью исчезли.

