Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 16 Позднеэдиакарский перелом
Дальше: Часть III От кембрийского взрыва до Великой ордовикской радиации (540–440 млн лет назад)
Глава 17
Два мира – два скелета
Нелегко найти аналоги в более поздней истории Земли и эдиакарским скелетным ископаемым. Были они относительно небольшими: до 5 см длиной или в диаметре – за одним-единственным исключением: размер намапойкии (Namapoikia) превышал в поперечнике 1 м. У самого распространенного из этих существ – клаудины (Cloudina) – скелет состоял из эксцентрично вложенных друг в друга неправильных известковых воронок с расширявшимися, словно воротники Елизаветинской эпохи, краями (рис. 17.1а). Намапойкия представляла собой бесформенную сотоподобную массу, суворовелла (Suvorovella), вероятно, обызвествленный прикрепительный диск вендобионта (рис. 16.1б), синотубулитес (Sinotubulites) – цилиндрическую раковину с многоугольным сечением из неупорядоченных наслоений арагонита (рис. 17.1в). Все эти древнейшие скелетные организмы тоже исчезли в конце эдиакарского периода и, судя по их необычному строению, не имеют отношения к последующему кембрийскому взрыву: появились они не ранее 550 млн лет назад, а к рубежу 541 млн лет вымерли. По образу жизни все они, вероятно, были прикрепленными фильтраторами, что объясняет их приуроченность к возвышенностям морского дна, где течения сильнее, и если скелеты этих организмов сохранились в прижизненном положении, то все они имеют сходную ориентацию, скорее всего, по направлению преобладающих водных потоков.

Эти организмы, несмотря на огромную разницу в строении, очень похожи друг на друга. Во-первых, состав скелета у них арагонитовый или высокомагнезиально-кальцитовый (Mg-кальцит). Эти две разности карбоната кальция (CaCO3), которые отличаются по строению кристаллической решетки: обычно шестоватые призмы – у первого и ромбоэдры – у второго. Кроме того, в кристаллической решетке арагонита некоторые атомы кальция замещены стронцием, у Mg-кальцита – магнием (не менее 0,004 %). Есть у этих минералов и общая черта: они легко растворимы, и, следовательно, построить из них скелет гораздо проще, чем из менее растворимого кальцита, тем более – из доломита, для разрушения которого нужны достаточно сильные кислоты. Поэтому организмы с доломитовым скелетом никогда не появились. Это означает, что эдиакарские скелетные организмы жили в нейтральной среде с относительно высоким содержанием Mg2+-ионов: при соотношении Mg2+/Са2+>2 формируются преимущественно Mg-кальцитовые и арагонитовые скелеты. Mg2+ также влияет на термодинамическую устойчивость чистого кальцита и тем самым препятствует его кристаллизации, но никак не затрагивает стабильность арагонита, потому организмы с арагонитовыми и Mg-кальцитовыми скелетами сосуществуют.
Во-вторых, внутреннее строение эдиакарских скелетов (микроструктура) очень простое: это мельчайшие гранулы (менее 4 мкм величиной), не имеющие определенной ориентации и образующие очень тонкие стенки (15–60 мкм толщиной). Такая примитивная микроструктура встречается у красных и зеленых обызвествленных водорослей, фораминифер, губок и кораллов, но даже у них скелеты имеют массивное многослойное строение. В-третьих, практически все эти организмы обладают бесскелетными двойниками: у клаудины – конотубус (Conotubus), у синотубулитеса – корумбелла (Corumbella), а у суворовеллы – аспиделла (рис. 16.1, 17.1). Это означает, что при насыщенности среды Са2+– и Mg2+-ионами, а также бикарбонатом они легко строили минеральные оболочки, а нет – так и не надо. Скелет был не убежищем, защищавшим от крупных хищников (они еще не появились), а складом ионных излишков. В тех же морях, где среда долго оставалась насыщенной названными ионами – на карбонатном мелководье Сибири и Намибии, эдиакарские скелетные организмы формировали мощные биогенные отложения: намибийские клаудины создали первый риф, причем 10-метровой высоты, в строительстве которого участвовали только животные (прежде рифы – строматолитовые и тромболитовые – создавали лишь бактерии), а сибирские суворовеллы образовали древнейший в мире слой ракушняка, тоже немаленький – более 2 м мощностью и свыше 1 км протяженностью (рис. 17.2). Все эти рекорды, как и размер самого крупного скелетного животного – намапойкии (более 1 м в поперечнике), были побиты даже не в кембрийском периоде, а 100 млн лет спустя – в ордовикском.
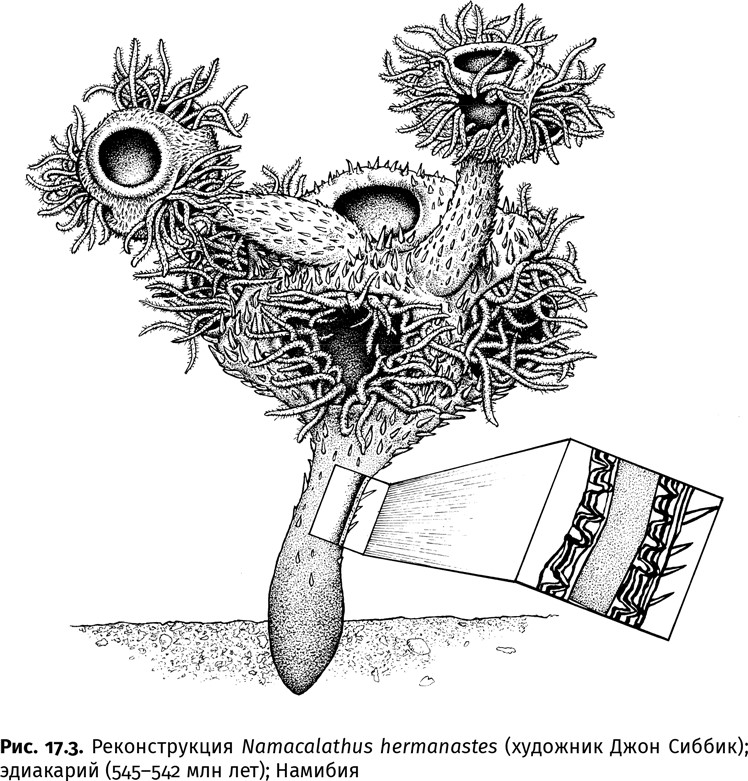
Уже в конце эдиакарского периода появились животные с иным типом скелета: намакалатус (Namacalathus) и анабаритиды (Anabaritida). Намакалатус обладал известковым скелетом в виде пористого правильного шестигранника на ножке (рис. 17.3). Анабаритиды строили свои спирально закрученные конические раковинки с тремя лопастями из аккуратно расположенных в несколько слоев арагонитовых шестоватых кристаллитов. Более толстая по сравнению с современниками (теми же клаудинами) стенка скелета намакалатуса сложена из тонких пластин, которые синхронно выгибались в сторону его наружной поверхности, образуя подобие тонких столбиков. Кроме того, между внешним и внутренним пластинчатыми контурами была прокладка из рыхлого материала, видимо насыщенного органическим веществом. Пластинчатые столбики служили напряженными элементами и вместе с органическим слоем придавали скелету одновременно и прочность, и пластичность.


Сочетание этих двух важных свойств минерального скелета в разных вариациях использовалось животными во всей фанерозойской эволюции. Любая часть скелета (скажем, пластинка многостворчатого моллюска – хитона или чешуя рыбы) – это многослойный, т. е. многоуровневый, элемент защиты. Благодаря различному взаимному расположению, а иногда и минералогическому составу микрокристаллов в такой пластинке образуются микрослои, одни из которых устойчивы к сдавливанию, другие – к скручиванию, третьи – на излом. Внешний слой обычно противостоит растворению. Каждый отдельный микрокристалл одет в органическую оболочку, а расположены микрокристаллы спиральными столбиками – все это усиливает скелет. И сам микрокристалл – это не единое целое, а конструкция из мириад наноразмерных кристаллитов. Такие наногранулы тоже имеют органические оболочки, которые позволяют им вращаться или раздвигаться. Благодаря высокой пластичности и иерархическому строению биокомпозиты – кости и раковины – и оказываются такими прочными: сила нажима или удара гасится органическими оболочками и обратимым смещением как пластин, так и наногранул, а развитие трещин тормозится. Например, арагонитовый перламутр (одна из микроструктур раковины моллюсков, которая состоит из плотно уложенных в несколько слоев шестигранных плоских призм) в 1000 раз прочнее и в 10 раз тверже, чем природные шестоватые кристаллы арагонита (табл. 1).

Высокая прочность биокомпозитов помогает существенно экономить на толщине скелета, увеличение массы которого в два раза требует троекратного повышения затрат на его перемещение. Основа прочности биокомпозитов, как отмечено выше, в их многоуровневом – иерархическом – строении, благодаря которому каждая степень защиты не просто дублируется, а дублируется многократно: каждый дополнительный слой снижает силу нажима на раковину в 10 раз. При этом сам потенциальный хищник, чтобы прокусить раковину или раздавить ее клешней, на преодоление каждой новой преграды теряет в четыре раза больше энергии.
В эдиакарском периоде, кроме намакалатуса, никто строить иерархические биокомпозиты «не догадался». В кембрийском и последующих периодах совершенствование таких скелетов стало одним из лейтмотивов эволюции. Уже среди древнейших кембрийских ископаемых мы встречаем остатки животных с очень сложным скелетом. Например, кольчатая трубочка кодоситубулуса (Codositubulus) из нижнекембрийских слоев Иберийских гор (Испания) при длине до 5 мм и толщине до 0,04 мм имеет три слоя: гомогенные внешний и внутренний и пластинчатый (псевдоперламутр) средний, призмы которого (5–7 мкм шириной и 2–3 мкм толщиной) состоят из более мелких кристаллитов (рис. 17.4). Все это сохранилось благодаря фосфатизации.

Не менее сложно устроены раковины древнейших моллюсков, которые могли состоять из нескольких структурно разных слоев, например призматического (наружного) и пластинчато-фиброзного (внутреннего). К ордовикскому периоду у моллюсков образуются самые совершенные и прочные перламутровая и перекрещено-пластинчатая микроструктуры. Ныне же в одной раковине, причем не сильно влияя на ее толщину, количество слоев может доходить до пяти: два – гомогенных, два – перекрещено-пластинчатых и призматический.
К сожалению, в естественных условиях, когда скелет после смерти живого существа остается без органических защитных оболочек, арагонит и Mg-кальцит закономерно либо растворяются, либо превращаются в обычный кальцит. Арагонит редко «живет» более 300 млн лет. Однако есть целый ряд признаков, позволяющих судить о присутствии этих минералов в первичном скелете. Иногда остается повышенная концентрация характерных элементов: Mg – в Mg-кальцитовых и Sr – в арагонитовых. О древнейших микроструктурах можно судить благодаря сохранению специфических микроструктур при замещении фосфатными минералами (как в случае с кодоситубулусом) или кремнеземом. Даже если практически ничего исходного в раковине не осталось, относительная сохранность разных микроструктур в одном и том же образце подскажет, что это было: кальцит остается неизменным, реликты изначального строения и присутствие микрокристаллов доломита укажут на исходный Mg-кальцит, а грубозернистая кристаллическая мозаика, заполняющая полости после полного растворения первичного минерала, – на когда-то существовавший арагонит. Поскольку при посмертном преобразовании скелетов они часто покрываются корочками морского цемента такого же состава, как сам скелет (фиброзный – на Mg-кальцитовом, ботриоидальный – на арагоните), это тоже помогает определить первичный состав самого субстрата. Аморфный кремнезем (SiO2 × nH2O), конечно, тоже преобразуется в кристаллический кварц (SiO2), а гидроксилапатит [Ca5(PО4,CO3)3ОН] – во фторапатит [Ca5(PО4,CO3)3F], но в этом случае химический состав минерала существенно не меняется, теряется только вода.
Совокупность этих данных позволяет установить, что в начале раннекембрийской эпохи, несмотря на значительное усложнение скелетных структур у животных, состав скелетов оставался арагонитовым и Mg-кальцитовым. Добавился только третий биоминерал – гидроксилапатит, что объясняется по-прежнему достаточно высоким содержанием ортофосфат-иона в океане. Четвертый биоминерал – кремнезем – использовали лишь относительно простые организмы: в кембрийском периоде это были одноклеточные радиолярии (Radiolaria), шестилучевые и обыкновенные губки (рис. 17.5 г).
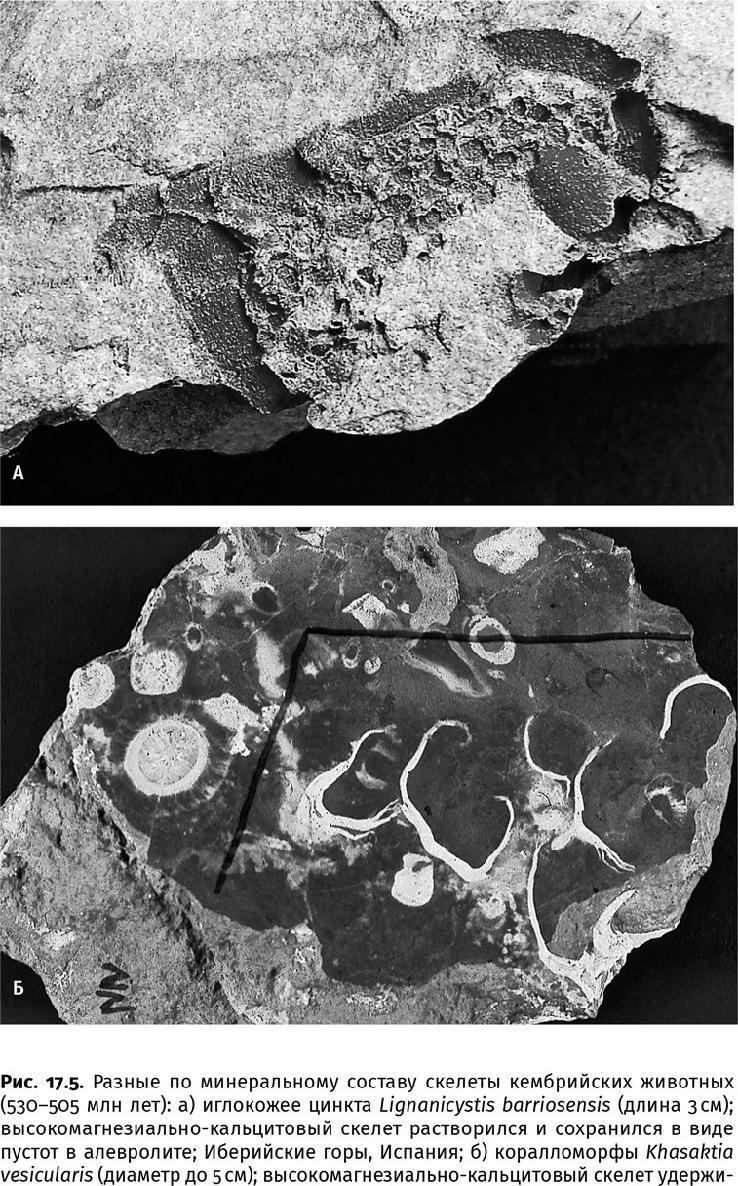
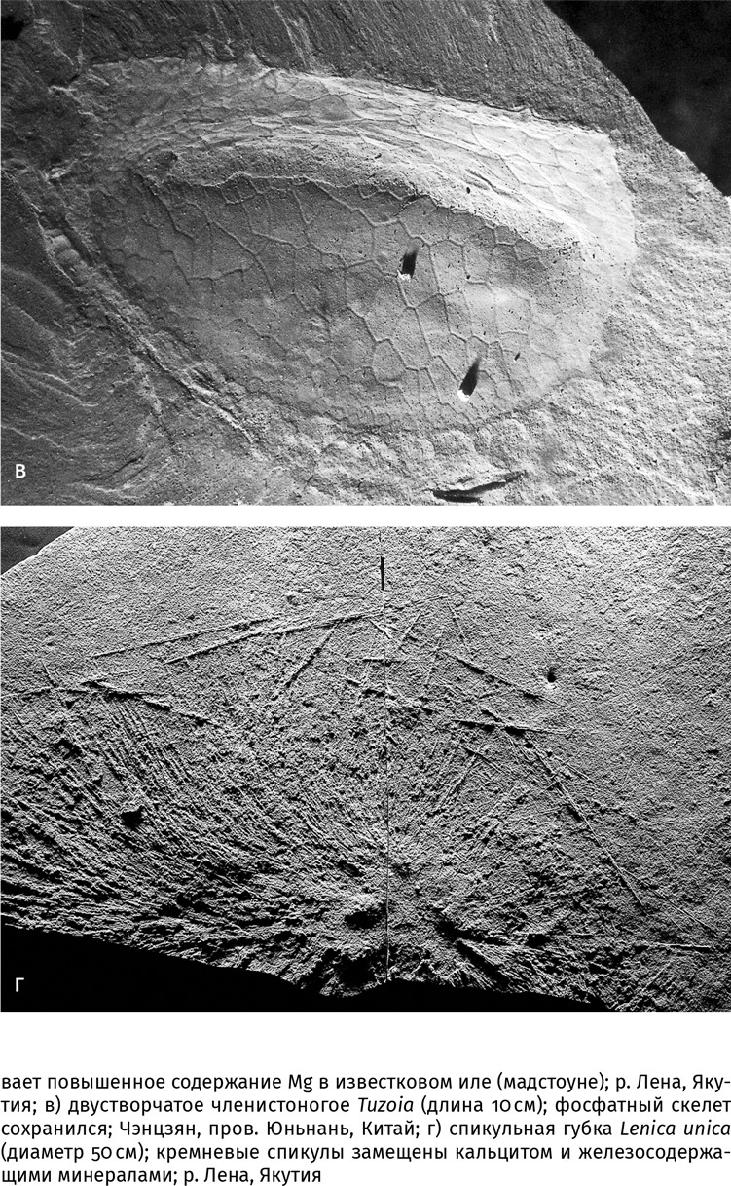
«Выбор» типа минерализации оставался за живыми организмами. Неподвижные сидячие животные [древние губки – археоциаты (Archaeocyatha), коралло- и водорослеподобные организмы] предпочитали массивный, но плохо организованный скелет из Mg-кальцитовых призм, напоминавший корки морского цемента. Подвижные и свободнолежащие донные формы [разные моллюски, хиолиты (Hyolitha) и ряд других вымерших групп] строили раковины из арагонита, компенсируя его небольшую толщину повышенной устойчивостью биокомпозита к физическим нагрузкам. (Формировать массивные арагонитовые скелеты энергетически затратно из-за большей плотности кристаллической решетки – 2950 кг/м3 против 2720 кг/м3 у кальцита, да и незачем: та же плотность обеспечивает его прочность.) Наконец, активные плавающие животные, по большей части хищники (некоторые членистоногие, щетинкочелюстные и хордовые, позднее – настоящие позвоночные), обзавелись фосфатными зубами, челюстями или другими органами захвата добычи.
Дело не только в том, что гидроксилапатит на несколько порядков по твердости превосходит любые структуры из карбоната кальция, для разрушения которого органы, состоящие из гидроксилапатита, и предназначены (табл. 1). Например, у современных высших раков фосфат сосредоточен в клешнях или ногочелюстях, остальной панцирь – кальцитовый. И это не случайно: при сокращениях поперечно-полосатых мышечных волокон, позволяющих развивать ногочелюстям или всему телу запредельные скорости (скажем, рак-богомол проламывает или прокалывает защиту жертвы за 0,0002 с), в организме вырабатывается молочная кислота. Кислотность внутренней среды возрастает от 4 до 5, и скелет легко бы растворился, будь он известковым. Опыты на форели, которой вживляли в мышцы разные по составу пластинки, показали, что известковые импланты в отличие от фосфатных растворялись, стоило рыбе немного поплавать.
Вновь обратившись к ископаемой летописи, видим, что предки позвоночных были активно плавающими хищниками с фосфатными зубами, а позднее и другими элементами скелета. Их образу жизни мы – потомки этих первых хищников – и обязаны своей слаборастворимой, прочной и в то же время удивительно пластичной внутренней опорой. А скажем, ближайшие скелетные родственники позвоночных – иглокожие – предпочли облегчить себе жизнь, соорудив панцирь из Mg-кальцитовых пластинок. С тех пор и ползают неспешно по дну, не освоив ни суши, ни даже слегка опресненных водоемов.
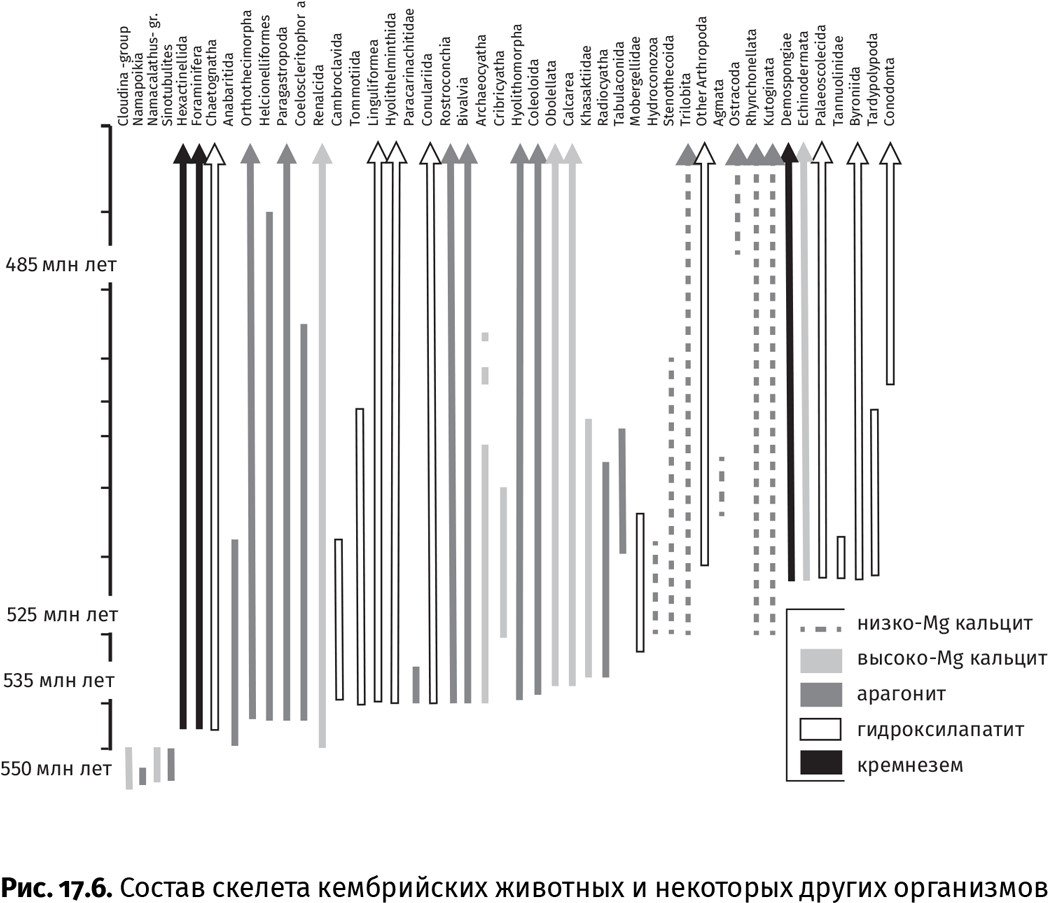
Лишь к концу раннекембрийской эпохи появились животные со скелетом из обычного кальцита (рис. 17.6). То было не просто добавление еще одной минеральной разности к предыдущим, а, по сути, смена парадигм: одни животные с арагонитовым и Mg-кальцитовым скелетом заменили его на кальцитовый (некоторые моллюски), другие, весьма обильные до той поры (анабаритиды, археоциаты), полностью вымерли, и им на смену пришли новые группы с кальцитовым скелетом [трилобиты (Trilobita), «замковые» брахиоподы], третьи, видимо просто избавились от минерального скелета (с губками и кораллами такое периодически происходит). Литологические (состав первичных морских цементов, оолитов и солей) и геохимические данные (соотношение Са2+/Mg2+-ионов) подсказывают, что в это время повысилось содержание углекислого газа в атмосфере и увеличился объем срединно-океанических хребтов. Поскольку воздушная оболочка Земли тесно взаимодействует с жидкой, то при росте содержания в атмосфере CO2 океан также насыщается этим газом. Реагируя с водой, газ образовывает нестойкую угольную кислоту, которая быстро распадается на ионы водорода и бикарбоната, а последний – на ионы карбоната и водорода:
CO2 + H2O ↔ H2CO3 ↔ H+ + HCO3– ↔ 2H+ + CO32–.
Избыток протонов подкисляет морскую воду. Если, скажем, содержание в атмосфере двуокиси углерода повысится в два раза по отношению к доиндустриальному уровню (0,03 %) при современной температуре поверхностных вод (25 °C) и солености (35‰), то водородный показатель (pH) упадет с 8,16 до 7,92, а кислотность водной среды, соответственно, повысится. Вполне достаточно, чтобы насыщенность океана по отношению к арагониту понизилась с 4,2 до 2,8. Насыщенность, Ω, рассчитывается по формуле Ω = [Ca2+] [CO32–]/K’, где K’ – объем растворяющегося минерала. Поскольку содержание в океане Ca2+ обычно в 20–30 раз превышает таковое CO32–, наиболее значимый количественный показатель здесь именно относительное изменение уровня CO32–. При повышении этого уровня арагонитовые, а также Mg-кальцитовые скелеты, если они плохо изолированы органическими оболочками, начинают растворяться еще при жизни своих хозяев. Впервые эпоха «массового растворения», связанная через парниковый эффект с наступлением теплой эры, случилась во второй половине кембрийского периода (и повторилась на рубеже пермского и триасового периодов). Коснулись эти события в основном водорослей, губок и кораллов, у которых скелеты не имеют достаточной степени защиты от повышенной кислотности.
Что касается влияния срединно-океанических хребтов, то чем больше их протяженность – а этот показатель зависит от числа континентов, – тем больше выделяется базальтовой лавы в рифтовых расщелинах. Взаимодействуя с морской водой, свежий базальт поглощает Mg2+-ионы, и в самой воде остается больше Са2+-ионов. В итоге преимущество опять же получают организмы с кальцитовым скелетом. Оба явления – рост срединно-океанических хребтов и повышение уровня углекислого газа – взаимосвязаны: разрастаясь, подводные хребты уменьшают емкость океанических впадин и тем самым выталкивают излишки воды на окраины континентов. Площадь суши сокращается, а вместе с ней и выходы горных пород с высоким содержанием кремнезема, на выветривание которых расходовался бы углекислый газ. Двуокись углерода накапливается в атмосфере все в больших объемах и растворяется в океане. При этом за счет гидротермальной активности в океан поступает больше ионов Cl–, Na+, и Ca2+, а SO42– и Mg2+ поглощаются новообразованной базальтовой лавой. Эти изменения можно наблюдать непосредственно, изучая состав капелек древней морской воды, запечатанных в кристаллах соли. И, судя по этим капелькам, содержание в океане кальция выросло с конца эдиакарского периода к середине раннекембрийской эпохи в три раза. Одновременно среди морских эвапоритов (солей) вместо сульфатов магния (например, кизерита – MgSO4 × H2O) начинают накапливаться калийсодержащие хлориды (такие, как сильвин – KCl и сильвинит – NaCl × KCl), что тоже наблюдается в середине раннекембрийской эпохи. Наоборот, в неогеновом периоде океан вновь изменился на арагонитовый, среди эвапоритов стали преобладать сульфаты, а температура на планете начала падать. В эту эпоху мы живем и сейчас.
Конечно, организмы строят скелет и из других минералов – без малого из сотни разновидностей (наиболее распространенные приведены в табл. 2). Но все другие варианты биоминерализации используются в исключительных случаях: например, сульфиды серы – пирит и грейгит (FeS2 и Fe3S4) – только улитками, обитающими вблизи гидротермальных источников срединно-океанических хребтов, черных курильщиков, которые выделяют большие объемы растворенного железа и серы (рис. 17.7). В любом случае в ход идут самые распространенные в земной коре, атмосфере и океане элементы, кроме алюминия, который организмы (люди) научились использовать только в XIX в.

Конечно, биоминералы используются и в других целях. Так, нанокристаллы магнетита служат у самых разных организмов (от бактерий до китов) в составе особых органов – магнитосом – для ориентации в пространстве с помощью характеристик магнитного поля Земли. Целестин или барит в виде звездчатых микрогранул помогают сцифоидным медузам и харовым водорослям определить направление силы тяжести: первым – чтобы быстрее вернуться в естественное положение после шторма, вторым – чтобы направить рост питающих ризоидов прямо в бескислородный слой осадка.

Однако только обладатели карбонатных, фосфатных и кремневых скелетов стали породообразующей силой, масштабы действия которой сопоставимы с силами геологическими.
Назад: Глава 16 Позднеэдиакарский перелом
Дальше: Часть III От кембрийского взрыва до Великой ордовикской радиации (540–440 млн лет назад)

