Книга: Сотворение Земли. Как живые организмы создали наш мир
Назад: Глава 14 Доломитовые Альпы
Дальше: Глава 16 Позднеэдиакарский перелом
Глава 15
Эмбриональная стадия Земли
Не только мощные отложения доломитов по всему миру отражают необычность морских обстановок эдиакарского периода. Особое место среди осадочных пород этого интервала и начала следующего, кембрийского, периода занимают фосфориты. Четвертая часть мировых запасов этого ценнейшего сырья для получения удобрений и горючих веществ (в прежние годы оно использовалось также для производства фотохимических реактивов) сформировалась в то время – 635–500 млн лет назад (рис. 13.2а). Содержание фосфата в пересчете на пентоксид фосфора (Р2О5) в месторождениях эдиакарско-раннекембрийского возраста (а их насчитывается более 350) составляет от 15 до 35 %, а запасы пяти самых крупных их них оцениваются более чем в 1010 кг для каждого (промышленными залежами фосфоритов считаются отложения, содержащие более 9 % пентоксида). Две другие эпохи фосфатонакопления приходятся на пермский период и на юрско-четвертичный интервал.
Именно юрские фосфориты в 1829 г. привлекли внимание геолога Уильяма Бакленда на южном побережье Англии. Он писал: «Свидетельство борьбы между собой обитателей нашей планеты, не пресекавшейся из поколения в поколение, – нетленный фосфатный помет, образовавшийся из их переваренных скелетов, который остался замурован в веществе и в фундаменте вечных холмов» – и позднее предположил, что фосфатизированный помет древних существ может использоваться для удобрения быстро истощавшейся почвы. Лишь когда его совету последовали во второй половине XIX – начале XX в. и в Центральной России стали разрабатывать залежи меловых фосфоритовых конкреций (минеральных, обычно округлых стяжений, которые образуются вокруг органических остатков), стало ясно, что масштабы фосфатонакопления вряд ли сопоставимы с деликатным механизмом их образования, предложенным Баклендом.
Советский геохимик Александр Васильевич Казаков в 1937 г. пришел к выводу, что фосфориты формируются иначе – в результате явлений, связанных с океаническим апвеллингом, как называют этот процесс теперь (англ. upwelling – подъем вод). Фосфор, как уже отмечалось в предыдущей главе, – один из важнейших биогенных элементов, без которого невозможно образование органического вещества, поэтому время его пребывания в океане ограничивается 10 000 годами: в виде ортофосфат-иона (РО43–) весь элемент поглощается бактериями и водорослями. Поступает же он только за счет выветривания континентов, главным образом кислых силикатных минералов (потому фосфориты полностью отсутствовали в архее). Далее вместе с органическим веществом фосфат уходит в глубины и при разложении органики вновь возвращается в раствор. В современном океане благодаря органическому «дождю» в год накапливается до 21×103 кг фосфата на 1 км2. Обогащенные фосфатом холодные подкисленные донные воды как часть системы океанических течений поднимаются наверх у западных берегов континентов. Это и есть апвеллинг. В зонах апвеллинга, особенно у берегов Перу – Чили и Намибии, «созревает» новый обильный урожай фитопланктона, на котором развиваются личинки разных животных и т. д. – по всей пищевой цепочке вплоть до крупных моллюсков, ракообразных и рыб. Именно поэтому, хотя зоны апвеллинга занимают всего 12 % площади океана, на них приходится 50 % продуктивности океана, чем пользуется человек. Фосфат же отчасти возвращается на сушу в виде помета морских рыбоядных птиц – гуано, являющегося ценнейшим удобрением.
Не весь растворенный фосфат в зонах апвеллинга потребляется организмами: часть его реагирует с бикарбонатом и оседает в виде минеральных корок и конкреций франколита [упрощенная формула – Ca5[PО4, CO3)3F]. Этот процесс происходит на глубине 300–400 м, на переходе от внешнего шельфа к континентальному склону при насыщении поровых растворов ионами Са2+ и РО43–. Именно к таким глубинам привязаны в большинстве своем фанерозойские залежи фосфоритов. Эдиакарско-кембрийские – другие. Они формировались в весьма мелководных условиях – там, где образовывался доломит, перекатывались валы оолитовых песков, накапливались ракушняки (кембрий), обитали цианобактерии и водоросли или росли строматолиты (рис. 15.1), что, в общем-то, неудивительно, ведь сероводородный слой был тогда намного ближе к поверхности. Все эти отложения тоже стали фосфатными в результате последующей циркуляции минеральных растворов. (Лишь микроструктура раковин и повышенное содержание в них кальция и магния свидетельствуют о первично арагонитовом или магнезиально-кальцитовом составе.) А строматолиты, видимо, сразу фосфатными и росли: в них нередко сохраняются остатки биоматов и бактериальных клеток, которые, как мы уже знаем, могли способствовать садке разных минералов, включая фосфатные.

Крупнейшие эдиакарско-кембрийские месторождения известны в Южном Китае, Вьетнаме, Монголии, Южной Сибири, Казахстане, Индии, Пакистане, Иране, Омане, Западной Африке (от Алжира до Сенегала и Буркина-Фасо), Бразилии, Австралии. Все эти регионы располагались на периферии большого южного материка Гондваны, который состоял из Африки, Индийского субконтинента, Южной Америки, Австралии и Восточной Антарктиды: одни – на континентальном шельфе самой Гондваны, другие – на небольших массивах (микроконтинентах) и даже отдельных подводных горах между Гондваной и Сибирью. Ныне останцы микроконтинентов впаяны в горные массивы Монголии, Казахстана и Южной Сибири. Механизм апвеллинга, конечно, мог бы работать на подводных горах, некоторых микроконтинентах и в отдельных областях Гондваны. Но отнюдь не везде одновременно. Более того, повышенное содержание фосфата, а также скопление фосфоритовых конкреций и наличие тонких фосфатных пластов отмечается и там, где промышленных залежей фосфоритов нет: в Западной Европе (это, впрочем, тоже периферия Гондваны), Балтии (Северная и Восточная Европа), Авалоне (древний континент, включавший самые восточные части Северной Америки, Уэльс и Англию) и Лаврентии (остальная Северная Америка, Шотландия и часть Шпицбергена).
Отчасти своеобразие эдиакарско-кембрийского фосфатонакопления было связано с недавно закончившимися оледенениями: таяние крупных масс льда всегда приводит к сбросу в океан больших объемов биогенных элементов, миллионами лет скапливавшихся во льду в виде пыли. Не могло не сыграть свою роль и формирование Гондваны: в результате коллизии южноамериканских и африканских кратонов 650–590 млн лет назад появился внушительный Панафриканско-Бразильский суперхребет (6000 км длиной при ширине 400–800 км), а затем (580–515 млн лет назад) на стыке Африки с Аравией, Индией и Антарктидой – не менее протяженный Трансгондванский (8000 км длиной и до 1500 км шириной). Образовалось и несколько хребтов поменьше. Высокие темпы роста и одновременно эрозии горных массивов выявляет ползущая вверх изотопная подпись стронция, сопоставимая с аналогичной кайнозойской, отражающей вздымание Гималайско-Тибетского плато. Из продуктов разрушения только Трансгондванского хребта могло получиться 10 × 1010 м3 осадочных пород – достаточно, чтобы покрыть 10-метровым слоем, скажем, площадь современной Сибири. Кислые магматические горные породы, слагавшие ядра этих горных массивов, тоже служили источником фосфата, вымывавшегося в результате их выветривания. Наконец, история с доломитами показывает, что океан был по большей части закисный с низким содержанием сульфатов (рис. 13.2а, в, и). А ведь именно активные сульфаты препятствуют не только осаждению доломита, но и фосфатных минералов: вместо франколита из раствора выпадают гипс и ангидрит, забирающие ионы кальция.
Изучение состава фосфоритов того времени показало, что фосфор попадал на дно не только с органическим веществом (уж больно много фосфора накопилось – столько в органике нет), но и на «железном шаттле». Оксигидроксид железа (FеОOH) связывал ортофосфат-ионы и увлекал их на глубину. В бескислородном водном слое фосфат высвобождался (так как FеОOH реагировал с HS–), концентрировался, и, поскольку этот слой находился недалеко от поверхности, любой подъем вод (т. е. формально опять же апвеллинг), особенно высокий прилив, приводил к насыщению мелководья фосфатом. Поскольку эта зона океана была сероводородной, здесь при окислении сульфида возникал избыток Н+, водный слой подкислялся и фосфатные минералы получали преимущество над карбонатом кальция. И на границе окислительно-восстановительных обстановок росли фосфатные корки, образовывались конкреции и происходила фосфатизация остатков организмов. Кроме того, как показали эксперименты, присутствие таких остатков ускоряло осаждение аморфного фосфата, который позднее кристаллизовался.
Однако в формировании и современных, и, вероятно, древних фосфоритов существенную роль играли нитчатые серные бактерии, плотные скопления которых покрывают тысячи квадратных километров континентального шельфа на краю окислительной и восстановительной зон. В кислородной среде они запасают для своих нужд NO3– и РО43–, а затем на длительное время перебираются в сероводородную обстановку, где для удержания нитрата и полифосфата используют энергию, полученную при окислении HS– до S0. После отмирания микробов гидролиз бактериального полифосфата ведет к быстрому осаждению франколита, кристаллы которого покрывают и чехлы самих бактерий, и остатки других организмов. Например, поселения современных гигантских бактерий тиомаргарит (Thiomargarita) в зонах апвеллинга способны накопить 0,05 кг фосфора на 1 кг сухого осадка, что соответствует месторождениям с 25 %-ным содержанием фосфата. Правда, это не нитчатая бактерия, а в эдиакарских фосфоритах встречаются именно фосфатизированные нитевидные чехлы с включениями серы.
С усилением аэрации океана в кембрийском периоде отложений фосфоритов становилось все меньше (они практически исчезли около 500 млн лет назад), фоновое содержание фосфата в отложениях сократилось в 8 раз, прекратилась и фосфатизация органических остатков.
Но прежде всеобщая фосфатизация всех и всего вызвала еще одно небывалое явление, которого не было ни до, ни после, – образование несметного числа фосфатных слепков. Эти слепки, своего рода посмертные маски, запечатлели не только внешний вид вымерших организмов, но их внутреннее строение вплоть до отдельных клеток и даже субклеточных структур – ресничек, некоторых органелл. Случилось это благодаря быстрому развитию микрокристаллов (менее 1 мкм длиной) франколита на органическом субстрате, в том числе на клеточных оболочках. В юрских, меловых и кайнозойских отложениях, в третью фазу фосфоритообразования, тоже возникали фосфатные слепки организмов с великолепно сохранившимися деталями тканевого и клеточного строения, но все это были достаточно крупные животные – рыбы, морские ящеры, птерозавры. Их распадающиеся ткани, наверное, и служили источником фосфата, как почти два века назад предположил Бакленд. Размеры же кембрийских и особенно эдиакарских ископаемых редко превышали 1 мм. Это могли быть даже эмбрионы и одноклеточные существа. Зато их число и масса в миллиарды раз превосходили любые скопления фосфатных слепков последующих периодов.
Важнейшие «залежи» таких остатков обнаружены в Южном Китае, Северной Монголии и Восточной Сибири. В раннекембрийских фосфоритах находят немало разных эмбрионов, в основном головохоботных червей и кишечнополостных, встречаются также мелкие членистоногие и их личинки, пятиустки, тихоходки, сохранившие все детали строения вплоть до мельчайших щетинок и пор (менее 1 мкм величиной); от крупных особей остаются только фрагменты конечностей менее 2 мм. Этот тип минерализации организмов получил название «эрстен» – по одному из шведских местонахождений, где был впервые установлен в 1960-е гг.
Самыми древними и интересными являются фосфориты Венъян возрастом 610–600 млн лет, в которых нашли фосфатизированные акритархи, красные водоросли, цисты (крепкие оболочки, в которых некоторые организмы пережидают неблагоприятные условия) и странные многоклеточные микроскопические шарики. Мощный, с десяток метров, пласт фосфоритов, относящийся к формации Доушаньтуо, залегает в горах китайской провинции Гуйчжоу. Пласт, конечно, разрабатывается для производства удобрений и, наверное, был бы весь уже исчерпан, если бы в нем не оказались эти окаменелости. Принадлежность встречающихся там ветвистых агрегатов из округлых телец (несколько миллиметров величиной) к красным водорослям определяется по отчетливому разделению таллома на коровый (внешний) слой и сердцевину, сложенные многочисленными призматическими клетками, а также по структурам, напоминающим карпогоны (женские органы размножения) и тетраспорангии (тетрада – четыре клетки, осуществляющие бесполое размножение) современных водорослей. Эти красные водоросли устроены намного сложнее, чем мезопротерозойские бангиевые, и, вероятно, представляют собой первые флоридеи. Округлые тельца (менее 1 мм размером) с полигональным узором на поверхности, скорее всего, являлись цистами планктонных многоклеточных водорослей: у них есть шипы на поверхности, обеспечивающие плавучесть, а внутри – тысячи мелких клеток, образовавшихся при палинтомическом делении (при таком делении по мере возрастания числа клеток их размер уменьшается). Причем, как у некоторых современных зеленых водорослей, в этих окаменелостях различаются репродуктивные клетки и неспособные к размножению соматические. Последние начинают делиться еще внутри цисты и образуют над новыми клеточными скоплениями свои оболочки. Подобные дочерние колонии специалисты назвали «матрешками».
А вот разгадать природу других многоклеточных шариков оказалось непросто. Эти микросферы (обычно 300–650 мкм в поперечнике), состоящие из еще более мелких сфер, были описаны как эмбрионы. Однако внешне так могли бы выглядеть и водоросли, и гигантские серные бактерии – обитатели зон апвеллинга, и даже естественные стяжения фосфатного минерала. С помощью фазово-контрастной синхротронной микротомографии некоторых венъянских микрообъектов в рентгеновском спектре излучения удалось увидеть, что неправильно-эллипсоидные 32-клеточные формы действительно напоминают эмбрионы со следами дробления на микромеры, сосредоточенные у одного полюса, и макромеры, цепочкой расположенные у противоположного полюса, ближе к «брюшной» стороне (рис. 15.2). В эмбрионах двустороннесимметричных животных такая цепочка предшествует закладке кишечника, тогда как микромеры дают начало покровным клеткам. Встречаются и другие возможные эмбрионы: у них на одном полюсе сосредоточены микромеры, образующие почти плоскую «кепочку», а на противоположном расположена пара гораздо более крупных макромеров. Такие эмбрионы благодаря неполному (меробластическому) дроблению дискоидального типа развиваются у разных современных животных, раннее развитие которых протекает через стадию богатого желтком яйца; желток и сосредоточен в макромерах, а микромеры делятся, как бы растекаясь тонким слоем по их поверхности и образуя подобие диска – «кепочки». Нечто, напоминающее желток с многочисленными слоями цитоплазмы, обнаружено и у венъянских эмбрионов.
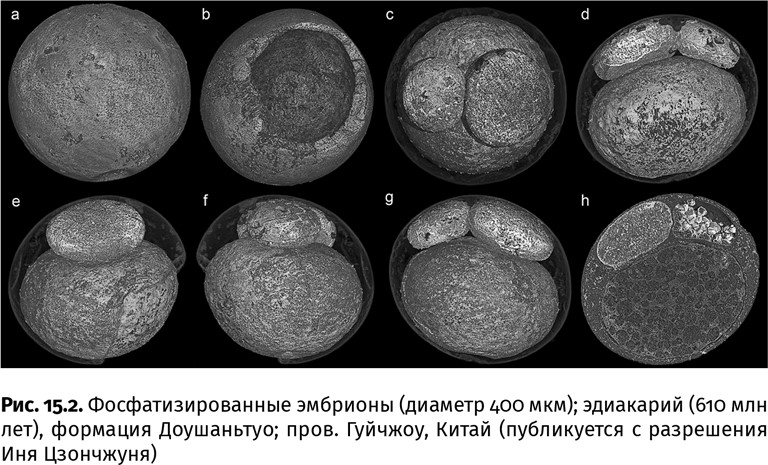
Чьи же это эмбрионы и что их внезапно погубило (иначе бы они не окаменели)? Изучение минералогии микросфер и распределения в них химических элементов показало, что появление некоего подобия клеточных слоев являлось результатом посмертных преобразований минерального вещества. Это исключает принадлежность ископаемых к сложным эмбрионам. Хотя окаменевшие стадии раннего развития в целом напоминают дробление эмбриона у многоклеточных животных, нельзя исключить, что раннеэдиакарские моря населяли их предшественники, жизненный цикл которых на этих ранних стадиях развития и завершался. Кроме того, в последние годы открывают все больше удивительных черт у некоторых одноклеточных организмов (раньше большинство из них считали упростившимися из-за паразитического образа жизни грибами): у этих существ, которых вместе с многоклеточными животными и грибами объединяют в группу Holozoa (от греч. ολος – целый, совершенный и ζωον– животное), на определенных стадиях развития тоже образуется некое подобие эмбрионов, состоящих из нескольких клеток.
Проще ответить на второй вопрос, и то приблизительно: эмбрионы мог погубить именно подъем бескислородных водных масс на мелководье, где плодились организмы. А поскольку в этих водах была повышенная концентрация фосфата, эмбрионы и оказались законсервированными навеки.
Назад: Глава 14 Доломитовые Альпы
Дальше: Глава 16 Позднеэдиакарский перелом

