Книга: От атомов к древу. Введение в современную науку о жизни
Назад: 5. липиды и мембраны
Дальше: 7. нуклеотиды
6. углеводы
Ну вот и присмотрись к себе, как сидишь ты возле бюро в халате. В руках-то что у тебя? Чашечка кофе? Еще и с сахаром? А думал ли ты, сколько невольников погибло на плантациях, кровью под бичами надсмотрщиков облившись, сколько жизней загублено было и что слез пролито, дабы кофе вот этот к тебе в чашечку попал? Про сахар и упомянуть страшно.Анна Коростелева. Александр Радищев
Углеводы по многим своим свойствам противоположны липидам. Если липиды — самые гидрофобные биологически активные вещества, то углеводы, пожалуй, самые гидрофильные. Значение углеводов для жизни так же огромно, как и значение липидов, хотя функции у них другие. Посмотрим же на них повнимательнее.
Мы уже знаем, что углевод — это спирт, одновременно являющийся или альдегидом, или кетоном. Углеводы бывают довольно разные. Основу любого углевода из тех, что могут заинтересовать нас в этой главе, образует цепочка, состоящая или из пяти, или из шести атомов углерода (см. рис. 6.1). Один из этих атомов углерода входит в состав либо альдегидной группы (если он на конце), либо кетогруппы (если он внутри цепочки). А ко всем остальным атомам углерода присоединены гидроксильные группы, как в спирте. Вот, собственно, и все (см. главу 1). Добавим, что в обиходе углеводы, подходящие под это описание, принято называть сахарами.
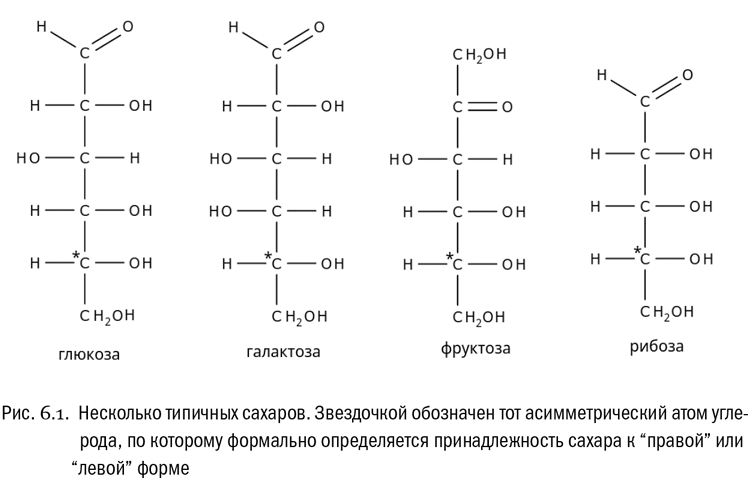
Например, что такое глюкоза? Это сахар с шестью атомами углерода, пятью гидроксильными группами и альдегидной группой. А фруктоза — сахар с шестью атомами углерода, пятью гидроксильными группами и кетогруппой. Причем и глюкоза, и фруктоза имеют формулу C6H12O6. Иначе говоря, это изомеры.
Бывают и пятиуглеродные сахара. Например, рибоза — сахар с пятью атомами углерода, четырьмя гидроксильными группами и альдегидной группой. В отличие от глюкозы и фруктозы, рибоза используется как пищевая добавка относительно редко, хотя в любых продуктах ее все равно полно, потому что биохимическое значение этого вещества колоссально.
А бывают ли сахара с другой длиной углеродных цепочек — например, трех-, четырех-, семи-, восьми- или девятиуглеродные? С точки зрения химии — конечно же, бывают. Иногда они встречаются и в живых организмах: например, семиуглеродные сахара могут быть промежуточными продуктами в синтезе липидов и в некоторых других биохимических процессах. Но в целом биологическое значение этих сахаров не слишком велико, и нас они пока что могут не волновать.
Молекулы сахаров не слишком сложны. Но есть один фактор, делающий их гораздо более разнообразными, чем можно было бы подумать, глядя на обычные графические формулы. Этот фактор — стереоизомерия.
Вновь о правом и левом
Стереоизомерия у углеводов, прямо скажем, такая, что сам черт сломит в ней ногу. Например, если внимательно посмотреть на молекулу глюкозы, легко убедиться, что она содержит целых четыре асимметрических атома углерода (см. главу 4). Четыре, а не один, как в хорошо знакомых нам аминокислотах! А ведь каждый асимметрический атом углерода создает два стереоизомера. Что это означает? Если в молекуле есть второй асимметрический атом, то у каждого из этих стереоизомеров будет еще по два стереоизомера. Если есть третий, то и у каждого из этих двух будет еще по два. И так далее. Следовательно, если молекула содержит четыре асимметрических атома, то стереоизомеров будет 16 (24). Причем по свойствам эти изомеры могут различаться между собой достаточно сильно. Например, галактоза — это шестиуглеродный сахар, совершенно идентичный глюкозе по набору функциональных групп. В галактозе тоже пять гидроксилов и одна альдегидная группа. Ее отличие от глюкозы состоит исключительно в том, что это другой стереоизомер. Между тем галактоза совершенно иначе участвует в обмене веществ, в том числе и у человека. Ферменты, работающие с глюкозой (которая у нас служит важнейшим питательным веществом), для ее усвоения не подходят. Галактоза перерабатывается особым ферментом, служащим только для этой цели. Кстати, его генетически обусловленное отсутствие у некоторых людей бывает причиной серьезного заболевания — галактоземии.
Как и другие вещества со стереоизомерией, сахара делятся на D- и L-формы — соответственно “правовращающие” и “левовращающие”. Здесь эти слова взяты в кавычки, потому что у сахаров связь формы молекулы и направления вращения поляризованного света на самом деле крайне запутанна. В детали нам вникать нет нужды. Достаточно знать, что химики придумали определенную чисто формальную процедуру, требующую считать сахар D- или L-формой в зависимости от положения радикалов у последнего асимметрического атома в углеродной цепочке. Например, у глюкозы это будет пятый атом, если считать от альдегидной группы.
Глюкоза и галактоза отличаются ориентацией групп у четвертого атома углерода. Они уже не считаются изомерами одного и того же сахара, а носят разные названия. Но при этом и глюкоза, и галактоза, участвующие в биологическом обмене веществ, — это D-изомеры.
В живой природе D-сахара вообще преобладают. Это почти такая же хиральная чистота, как и в случае с аминокислотами. Только у аминокислот преобладают “левые” изомеры, а у сахаров, наоборот, “правые”.
Чем вызвана эта разница? В последнее время появилась вполне убедительная гипотеза, что дело тут как раз в аминокислотах. А точнее — в их каталитическом действии. Напомним, что катализатор — это вещество, ускоряющее химическую реакцию, но само не претерпевающее в ней стойких изменений (см. главу 3). Так вот, существуют многоступенчатые процессы синтеза углеводов, которые катализируются аминокислотами (именно отдельными аминокислотами, а не целыми белками). И показано экспериментально, что L-аминокислоты катализируют синтез шестиуглеродных сахаров таким хитрым образом, что на выходе получается больше D-изомеров этих сахаров, чем L-изомеров. Причем часто намного больше: избыток D-изомера сахара в таких реакциях может достигать нескольких десятков процентов. Проще говоря, “левые” аминокислоты диктуют преобладание “правых” сахаров. И дело тут в чисто химическом механизме синтеза сахаров, в котором аминокислоты принимают участие.
Тогда получается, что никаких загадочных космических причин избытка D-сахаров искать не надо. Если такие причины и действовали — то в основном на аминокислоты (см. об этом главу 4). А что касается сахаров, то здесь живые организмы просто приспособили к делу те молекулы, которые предложила им обычная химия.
О пятиугольниках и шестиугольниках
До сих пор мы по умолчанию предполагали, что молекула сахара представляет собой линейную цепочку. А как же иначе? Но на самом деле в тех условиях, которые господствуют в живом организме, сахара обычно переходят из линейной формы в более энергетически выгодную циклическую. Как это происходит, можно посмотреть на примере глюкозы. Будем иметь в виду, что атомы углерода в глюкозе принято нумеровать, начиная от альдегидной группы. Атом, образующий эту группу, — первый, атом на противоположном конце цепочки — шестой. (Из формулы, кстати, нетрудно видеть, что асимметрическими тут являются второй, третий, четвертый и пятый атомы.)
Итак, переход глюкозы в циклическую форму начинается с того, что в альдегидной группе разрывается одна из двух связей, соединяющих кислород с углеродом (см. рис. 6.2А, Б). В результате у кислорода освобождается валентность, на которую переходит водород от гидроксильной группы предпоследнего (в данном случае пятого) атома углерода (см. рис. 6.2В). Тем самым при первом атоме образуется гидроксильная группа, которой раньше там не было. Теперь в молекуле остается две свободные валентности: у атома углерода бывшей альдегидной группы и у атома кислорода бывшего гидроксила. Они замыкаются друг на друга (см. рис. 6.2Г). Получается цикл. А точнее, кольцо, в котором первый и пятый атомы углерода соединены друг с другом через атом кислорода (–O–). В случае глюкозы это кольцо шестичленное, в него входят пять углеродов и кислород.
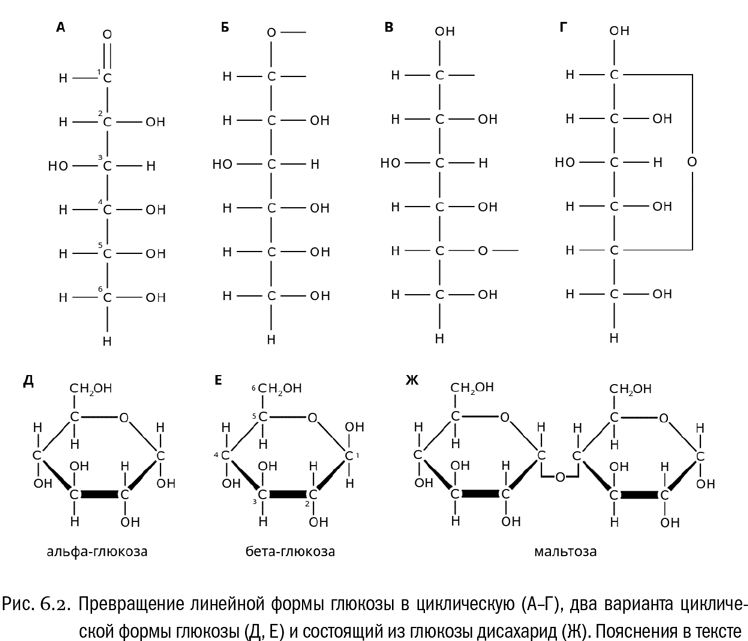
По такому же принципу переходят в циклическую форму и рибоза, и фруктоза, и другие сахара. Правда, и у рибозы, и у фруктозы кольца получаются пятичленными. По этому признаку их легко отличить от глюкозы, особенно если мы видим уже нарисованную кем-нибудь формулу.
Существование в циклических и линейных формах — еще один вид изомерии, на этот раз свойственный только сахарам. После перехода из линейной формы в циклическую гидроксильная группа исчезает при пятом атоме углерода, зато появляется при первом. Но это все равно глюкоза.
Напоследок — еще одна деталь, которая может нам пригодиться. В циклической форме сахара гидроксильные группы могут находиться по разные стороны от плоскости образовавшегося кольца — условно говоря, “над” или “под” кольцом (см. рис. 6.2Д, Е). При этом структура линейной формы глюкозы однозначно определяет ориентацию всех гидроксилов, кроме вновь образующегося при циклизации первого. Подсказка: на изображениях циклической формы глюкозы первый атом углерода, вместе со своим гидроксилом, обычно находится в правом углу. Так вот, если на такой формуле гидроксил оказался под кольцом — значит, это альфа-глюкоза, а если над кольцом — бета-глюкоза. Это еще один особый вид изомерии, уникальный не просто для сахаров, а для их циклических форм. Где и почему он важен в биологии, мы скоро увидим.
Краткий обзор сахаров
Глюкоза важна для живых организмов в первую очередь как источник энергии. Попросту говоря, основное предназначение глюкозы — в том, чтобы распадаться на более мелкие молекулы. Освободившаяся при этом энергия собирается и используется в разных полезных целях.
В нашем организме есть два способа такого распада: брожение и дыхание. Брожение — это неполный распад глюкозы, в результате которого она превращается в две молекулы молочной кислоты C3H6O3. А дыхание — это полный распад до самых простых молекул, а именно до углекислоты (CO2) и воды (H2O). В отличие от брожения, дыхание обязательно требует присутствия свободного кислорода (O2). Зато и энергии дыхание дает намного больше.
В крови человека постоянно есть глюкоза. Падение ее уровня ниже определенной пороговой концентрации ведет к голодной смерти. С другой стороны, устойчиво повышенный уровень глюкозы — это признак такого неприятного заболевания, как сахарный диабет. Но в любом случае надо понимать, что вся эта система — в высшей степени динамическая. Вся глюкоза, находящаяся в крови взрослого человека в любой данный момент, может быть полностью израсходована примерно за 15 минут. Ясно, что ее запас должен непрерывно пополняться. И неудивительно, что именно концентрация глюкозы в крови определяет физиологическое ощущение голода или сытости. В области нашего мозга, называемой гипоталамусом, есть специальные клетки-глюкорецепторы, постоянно измеряющие, сколько глюкозы содержится в омывающей их жидкости. Пищевая мотивация запускается или выключается в зависимости от того, какие сигналы идут от этих клеток.
Кроме того, содержание глюкозы в крови контролируется специальными гормонами. Это инсулин, запускающий захват глюкозы клетками, и глюкагон, который, наоборот, стимулирует высвобождение глюкозы из клеток в кровь. Инсулин понижает концентрацию глюкозы в крови, а глюкагон повышает. И инсулин, и глюкагон — белки. Это самый что ни на есть классический пример сигнальной функции белков, о которой мы говорили в главе 3. И инсулин, и глюкагон синтезируются у человека в поджелудочной железе, уходя оттуда в кровь и распространяясь по всему телу. Недостаток инсулина — одна из главных возможных причин сахарного диабета.
Придумавший метод выделения инсулина и раскрывший тем самым механизм диабета сэр Фредерик Бантинг до сих пор остается самым молодым лауреатом Нобелевской премии по физиологии и медицине — он получил ее в 1923 году в возрасте 32 лет. Интересно, что в жизни самого Бантинга это открытие было лишь эпизодом. Врач по основной специальности, он в разные периоды жизни был неплохим художником и полярным путешественником, занимался ради собственного интереса самыми разными медицинскими проблемами вроде болезней легких и адаптации летчиков к высоте, а еще участвовал в обеих мировых войнах и погиб при крушении военного самолета в 1941 году.
Широко известно (и является чистой правдой), что главный потребитель глюкозы в нашем теле — это мозг. Интересно, что сердце, наоборот, один из самых “всеядных” органов, составляющий в этом плане полную противоположность мозгу. Клетки сердца охотно получают энергию и из жирных кислот, и из аминокислот, и даже из такого отхода, как молочная кислота. Мозг же требует глюкозы, причем в большом количестве и бесперебойно. Как ни парадоксально, именно поэтому работа мозга почти не зависит от режима питания. Дело в том, что энергетическое снабжение мозга — это константа. Оно всегда стоит на максимальной отметке и не может быть снижено ни при каких физиологических условиях: на мозге организм не экономит. Если снабжение мозга глюкозой все-таки падает, это приводит к опаснейшей для жизни ситуации, которая называется гипогликемической комой. Чаще всего она возникает из-за неаккуратного введения слишком большой дозы инсулина — тут уж, что называется, “против лома нет приема”. (В истории медицины бывало, что гипогликемическую кому вызывали специально: это называлось инсулиновым шоком, или инсулинотерапией, и использовалось в XX веке как способ лечения шизофрении — впрочем, с сомнительной эффективностью. В знаменитом фильме “Игры разума” показано, что этим варварским методом пытались лечить математика Джона Нэша.) Но никакой переменой питания столь впечатляющего эффекта добиться нельзя. В частности, это означает, что практически никакая диета для мозга не опасна. Но нельзя забывать, что при этом она вполне может быть опасна для других органов — здесь надо соблюдать осторожность.
Фруктоза — питательный углевод, которого много в растениях, например в сладких ягодах и плодах. Это шестиуглеродный сахар, отличающийся от глюкозы тем, что содержит не альдегидную группу, а кетогруппу. В нашем организме фруктоза частично превращается в глюкозу, а частично усваивается самостоятельно (для этого есть специальные ферменты).
Галактоза — стереоизомер глюкозы, углевод, который человек получает, прежде всего, в составе молочных продуктов. При усвоении нашим организмом галактоза первым делом превращается в глюкозу. Сбой работы соответствующего фермента как раз и вызывает болезнь галактоземию. Впрочем, без внешнего источника галактозы взрослый человек в случае чего может обойтись.
Наконец, два важнейших пятиуглеродных сахара — рибоза (уже нам знакомая) и отличающаяся от нее на один атом кислорода 2-дезоксирибоза. У дезоксирибозы отсутствует одна из гидроксильных групп, вместо нее — просто атом водорода; цифра “2” — номер того атома углерода, при котором эта отсутствующая гидроксильная группа могла бы находиться. Полные названия обоих сахаров — D-рибоза и 2-D-дезоксирибоза, но приставки часто опускаются, если и так понятно, о чем идет речь. Заодно отметим, что атомы углерода в циклических формах рибозы и дезоксирибозы по некоторым историческим причинам принято обозначать не просто цифрами, а цифрами со штрихами — вот так: 2'-дезоксирибоза.
Рибоза и дезоксирибоза нужны живым организмам в первую очередь для того, чтобы строить из них более сложные молекулы, которые называются нуклеотидами и нуклеиновыми кислотами. О них мы поговорим в главах 7 и 8.
Дисахариды
Молекулы сахаров могут создавать друг с другом ковалентные связи. Для этого нужны две гидроксильные группы, принадлежащие двум сахарам. От одной из них отщепляется водород (–H–), другая отщепляется целиком (–OH), эти фрагменты образуют воду (H–O–H), а оставшиеся свободные валентности замыкаются друг на друга, и в результате между остатками сахаров остается мостик с атомом кислорода посредине (–О–). Такая связь называется гликозидной (см. рис. 6.3).
Сложный сахар, состоящий из двух углеводных остатков, соединенных гликозидной связью, называется дисахаридом. А простой сахар, не состоящий из нескольких углеводных остатков, в той же терминологии называют моносахаридом. На данный момент нам знакомо пять моносахаридов: глюкоза, фруктоза, рибоза, дезоксирибоза и галактоза (см. рис. 6.3).
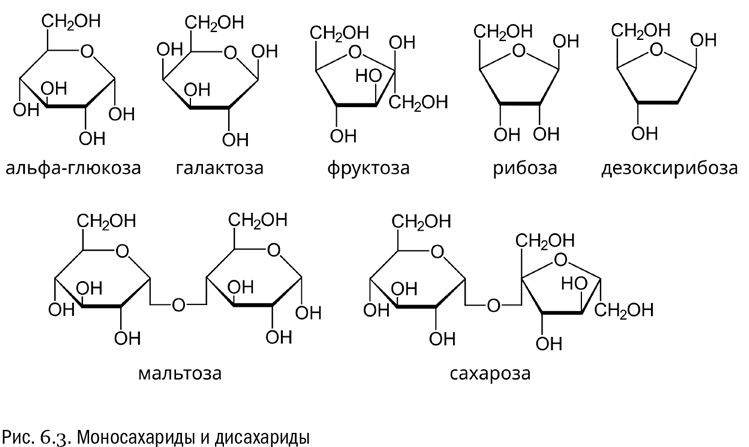
Самый известный дисахарид, с которым человек имеет дело буквально каждый день, — сахароза, состоящая из остатков глюкозы и фруктозы (см. рис. 6.3). Сахароза — это обычный столовый сахар. Получают ее из сахарного тростника, а в странах, где тростник не растет, — в основном из свеклы. Поскольку в сахарозе есть глюкоза, она категорически запрещена людям с сахарным диабетом и не рекомендуется людям с риском такового — в отличие от чистой фруктозы, для усвоения которой не нужен инсулин и которая в принципе допустима даже в диете диабетиков. Фруктоза, кстати, гораздо слаще сахарозы, а та, в свою очередь, слаще чистой глюкозы.
Если соединить гликозидной связью два остатка глюкозы, то получится еще один широко распространенный дисахарид, а именно мальтоза (см. рис. 6.3). Это обычнейшая составная часть любой растительной пищи. Иногда мальтозу называют солодовым сахаром, потому что ее много в зернах: солод — это и есть проросшее зерно. Кстати, слово “солод” родственно слову “сладкий”: например, в сербохорватском языке оно так и пишется “слад”, в чешском и словацком — slad, в польском — słód.
Еще один важный для человека дисахарид — лактоза, состоящая из остатков глюкозы и галактозы. Лактозу называют молочным сахаром, потому что ее действительно очень много в молоке. В отличие от большинства других сахаров, она почти не сладкая. Для усвоения лактозы нужен фермент лактаза, который рвет гликозидную связь, расщепляя лактозу на глюкозу и галактозу. Если этого фермента нет или его не хватает, возникает непереносимость лактозы. Ничего особенно опасного в этом состоянии нет, но таким людям приходится или ограничивать себя в молочной пище, или пить предназначенное специально для них безлактозное молоко (такое обычно есть в современных крупных магазинах).
Полисахариды
Гликозидные связи могут соединить между собой не только пару остатков моносахаридов, но и какое угодно их количество. Длинные цепочки моносахаридов, соединенных гликозидными связями, называются или олигосахаридами (если остатков в них больше двух, но не больше десяти), или полисахаридами (если они еще длиннее). Полисахариды — это уже самые настоящие полимеры (см. главу 3). Число остатков сахаров в молекуле полисахарида вполне может измеряться многими тысячами.
Один из самых широко известных полисахаридов — крахмал. Он представляет собой длинную цепь соединенных гликозидными связями остатков глюкозы. Это важнейшее запасное вещество у растений. Крахмала много в любой растительной пище. Он очень легко переваривается, расщепляясь до глюкозы, которая сразу же поступает в кровь. Обычная мука больше чем на 60% состоит из крахмала, а вот белков там, наоборот, довольно мало. Когда доктор Хаус в 20-й серии 6-го сезона обзывает пончики “жареными углеводами”, он подразумевает в первую очередь именно крахмал. Ну и заодно — сопутствующие углеводы вроде мальтозы, которых в пончиках, конечно, тоже предостаточно.
Животный аналог крахмала — гликоген. Это тоже важный запасной углевод. У нас он накапливается в первую очередь в печени. В случае надобности молекулу гликогена можно быстро расщепить до молекул глюкозы, которые тут же уйдут в кровь. Гликоген — это энергетический запас, пригодный к срочному использованию. Этим он отличается от жиров, которые содержат больше энергии, но мобилизуются гораздо медленнее. Впрочем, надо сказать, что расщепление гликогена у нас идет не только при каких-то чрезвычайных нагрузках, но и в состоянии покоя, понемногу, в промежутках между приемами пищи. Это важный способ поддержания концентрации глюкозы в крови на постоянном уровне. Само слово “гликоген” буквально означает “рождающий сладость”.
Итак, мы можем видеть, что полисахариды отличаются от моносахаридов в нескольких отношениях. Во-первых, их молекулы огромны. Во-вторых, они не растворяются в воде (именно поэтому их удобно делать запасными веществами). И в-третьих, они не сладкие. Например, чистый крахмал — это безвкусный порошок. По всем этим причинам называть полисахариды “сахарами” уже как-то не принято. Термин “сахар” обычно относят или к моносахаридам, или к дисахаридам.
Вспомним, что в ходе нашего разговора мы уже встречались с полимерами (см. главу 3). Это были белки. Мономеры белков — аминокислоты — соединены друг с другом пептидными связями. А мономеры полисахаридов — моносахариды — соединены друг с другом гликозидными связями. Серьезное различие здесь в том, что белки не ветвятся. А вот полисахариды вполне могут. “По умолчанию” гликозидная связь образуется между гидроксилами первого и четвертого атомов углерода глюкозы (1-4-гликозидная связь). Тогда из остатков глюкозы получается простая линейная цепочка. Но в глюкозе есть и другие гидроксилы, между которыми образование гликозидной связи тоже возможно. Например, на 1-6-гликозидной связи полимерная цепочка обычно разветвляется (см. рис. 6.4). В гликогене такое ветвление выражено сильнее, чем в крахмале, хотя оно есть и там и там.
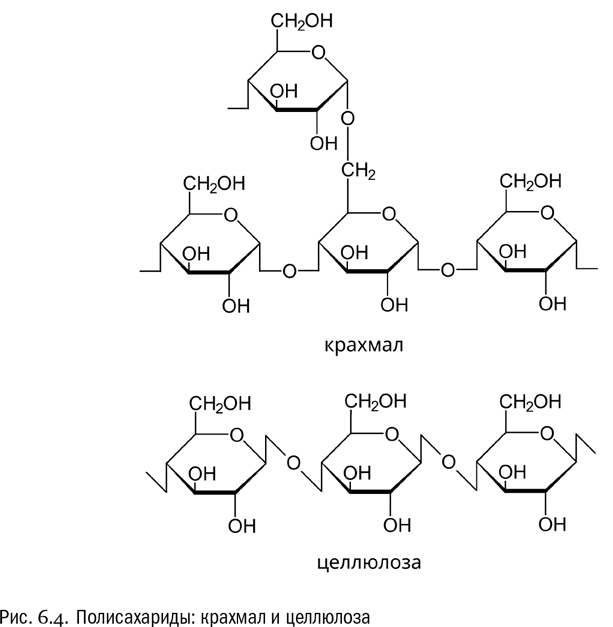
Еще один полимер глюкозы — целлюлоза, или клетчатка. Это плотное волокнистое вещество, образующее основу древесины. В том или ином количестве целлюлоза есть в любой части любого растения. С химической точки зрения целлюлоза принципиально отличается от крахмала тем, что состоит не из альфа-глюкозы, а из бета-глюкозы. При этом возникают так называемые бета-гликозидные связи, отличающиеся от присутствующих в крахмале и гликогене альфа-гликозидных (последние выглядят на схеме молекулы прямыми, а бета-гликозидные — зигзагообразными). Кроме того, молекулы целлюлозы не ветвятся (см. рис. 6.4).
Бета-гликозидные связи гораздо прочнее альфа-гликозидных. Они расщепляются только очень немногими ферментами. Например, почти никто из животных, питающихся растениями, не может самостоятельно переваривать целлюлозу. Тем, кто берется ее усваивать, приходится заводить для этой цели микроскопических кишечных сожителей, у которых есть нужный фермент — целлюлаза. В теле человека таких микроорганизмов нет, и он не может переваривать целлюлозу вообще. Правда, она все равно приносит некоторую пользу, придавая пище подходящую консистенцию и стимулируя сокращения кишечника. Но никакой питательной ценности целлюлоза для нас не имеет, в этом смысле она — бесполезный балласт.
Долгое время биологи были уверены, что самостоятельно производить целлюлазу неспособны абсолютно никакие животные. И только в 1998 году выяснилось, что у некоторых термитов все-таки есть целлюлаза “собственного производства”, к синтезу которой не имеют отношения никакие микроорганизмы. А в дальнейшем подобные “встроенные” целлюлазы были найдены у нескольких питающихся древесиной тараканов, жуков, круглых червей и улиток. И по всей вероятности, этот список еще не полон. Отличная иллюстрация того, как рискованны в биологии категоричные утверждения! Слишком уж разнообразны объекты этой науки, сюрпризы исследователям они подбрасывают очень часто.
Клеточные стенки и судьба Земли
Совершенно особый интерес для биологов представляют те полисахариды, которые входят в состав клеточных стенок. Из предыдущей главы мы уже знакомы с клеточными мембранами. Так вот, клеточная стенка — это совсем не то. Клеточной стенкой называется самостоятельная оболочка, находящаяся снаружи от мембраны и заключающая в себе всю клетку целиком. Такое расположение означает, что это, строго говоря, внеклеточная структура — наподобие домика или раковины. Обычно она жесткая и придает клетке постоянные очертания. Клеточная стенка может состоять из полисахаридов (у растений, грибов), из сложных полимеров, в состав которых входят углеводы и аминокислоты (у бактерий) или из белков (у архей). У некоторых организмов, например у животных, клеточных стенок нет вообще. Это делает клетки менее прочными, зато позволяет им легко менять форму.
Клеточные стенки растений — целлюлозные. Растительная клеточная стенка часто бывает гораздо толще клеточной мембраны. Если растение многоклеточное, то между клетками обычно есть плазмодесмы — проходящие сквозь отверстия в клеточных стенках межклеточные “мостики” (см. рис. 6.5А). Через плазмодесмы растительные клетки общаются между собой и обмениваются разными веществами.
Сухое дерево, так же как пробка и другие подобные материалы, представляет собой не что иное, как массу пустых клеточных стенок. Живых клеток там давно уже нет, но их стенки остаются — целлюлоза для этого достаточно прочна. Ее прочности хватает на то, чтобы древесина сохраняла свою структуру буквально тысячелетиями: самое старое деревянное здание мира — японский буддийский храм Хорю-дзи, построенный в VII веке, в эпоху Асука. Кроме того, из целлюлозы, когда-то входившей в растительные клеточные стенки, делают бумагу.
Между прочим, сам термин “клетка” (cell) когда-то ввел в науку английский физик Роберт Гук, который исследовал под микроскопом пробку и увидел в ней характерные маленькие полости. В данном случае это были не живые клетки, а именно пустые целлюлозные клеточные стенки, повторяющие их форму.
На самом деле растительная клеточная стенка вовсе не состоит исключительно из чистой целлюлозы. В нее еще обязательно входят короткие ветвящиеся полимеры, причем включающие не только глюкозу, но и другие моносахариды. Эти полимеры собирательно называются гемицеллюлозами (см. рис. 6.5Б). Есть там и некоторые структурные белки. Целлюлоза вместе с гемицеллюлозами и белками образует сложную объемную сеть, усиленную к тому же водородными связями (см. главу 2). Благо между длинными молекулами целлюлозы, в которых много гидроксильных групп, водородные связи возникают очень легко. Для клеточных стенок растений, а значит, и для древесины это важный источник прочности.
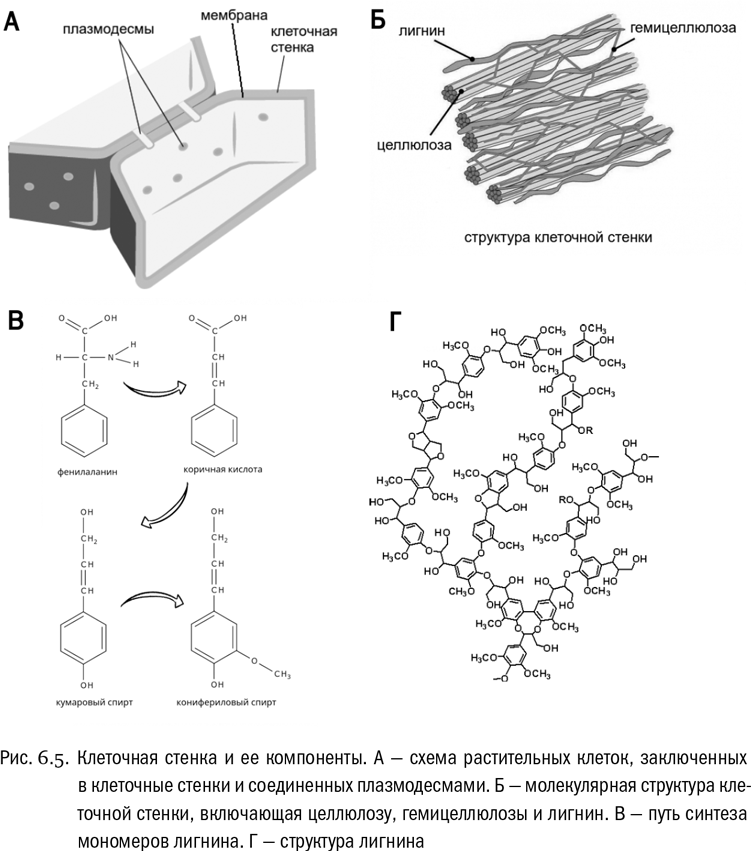
Ну а с точки зрения истории жизни на Земле самая интересная составляющая клеточной стенки растений — это лигнин. Очень своеобразное вещество, у которого нет никакой единой химической формулы. Строго говоря, его и самостоятельным веществом-то нельзя считать. Лигнин — это не углевод. Это сложный полимер, “сшитый” из нескольких разновидностей спиртов. У всех этих спиртов есть более-менее длинные углеводородные цепочки, включающие ароматические ядра (см. главу 1). И все они синтезируются из аминокислоты фенилаланина, которая превращается сначала в коричную кислоту — замечательное вещество, входящее в состав масла корицы, — а потом уже в разнообразные спирты, обычно имеющие две или три гидроксильные группы (см. рис. 6.5В). В лигнине эти спирты сшиваются между собой ковалентными связями в самых разных направлениях, буквально вдоль и поперек, так что получается запутанная сетка (см. рис. 6.5Г).
Образование лигнина — уникальный признак сосудистых растений, то есть папоротников, плаунов, хвощей, хвойных и цветковых. Это эволюционное “изобретение”, сделанное только после выхода растений на сушу, и то далеко не сразу. Дело в том, что лигнин благодаря своей особой структуре придает клеточным стенкам огромную механическую прочность. Он необходим, чтобы сделать ствол наземного растения высоким, вплоть до многометрового, и создать транспортную систему из микроскопических трубочек, качающую воду на всю эту высоту. А отсюда следует, что именно с “изобретением” биосинтеза лигнина связано одно из величайших событий, поменявших облик Земли, — появление лесов (см. главу 17).
Кроме того, появление лигнина сильно повлияло на глобальный, то есть общепланетный, круговорот углерода. Дело в том, что лигнин с его разнообразными мономерами и перепутанными химическими связями исключительно неподатлив к действию ферментов. Поэтому растительной тканью, в которой много лигнина, почти невозможно питаться. Из всех земных живых организмов эффективно разлагать лигнин “научились” только грибы, причем не все и не сразу. Именно они и стали главными разрушителями стволов мертвых деревьев. До этого вся огромная биомасса лигнифицированной древесины просто захоранивалась как есть, создавая залежи каменного угля. В честь этих залежей получил название целый геологический период — каменноугольный, или карбон.
Карбоновые леса непрерывно вели фотосинтез и выделяли в атмосферу огромное, немыслимое ни в какие более ранние эпохи количество кислорода. Мы знаем, что свободный кислород (O2) нужен для дыхания, то есть для полного окисления питательных веществ. Таким питательным веществом могла бы служить и древесина погибших деревьев. Но в карбоновом периоде эффективные деструкторы еще не возникли, поэтому перерабатывать древесину было некому. Стволы деревьев просто захоранивались, и заключенный в них углерод уходил из экологического круговорота вовсе. А живые деревья тем временем продолжали выделять кислород, который накапливался в атмосфере. В результате атмосферная концентрация кислорода достигла уникальной в истории Земли цифры 35%. Как известно, современная атмосфера Земли содержит “всего” 21% кислорода. На самом деле по космическим меркам и это невероятно много, но в карбоне было в полтора раза больше. А дело тут именно в том, что огромная биомасса стволов деревьев в карбоне не съедалась никакими живыми существами. В отличие от современной ситуации, когда упавшие стволы измельчаются насекомыми, перерабатываются грибами и в итоге их углеродные соединения окисляются дыханием до углекислоты (CO2). При этом расходуется кислород, а углекислота выдыхается и уходит в атмосферу.
До той биомассы, которая успела захорониться в виде каменного угля раньше, чем возникли эффективные деструкторы, живая природа смогла добраться только с появлением человека, который неутомимо откапывает каменный уголь и жжет его, используя в качестве топлива. Будем иметь в виду, что процессы дыхания и горения описываются строго одним и тем же суммарным уравнением:
C6H12O6 (глюкоза) + 6O2 → 6CO2 + 6H2O
Одна молекула глюкозы взаимодействует с шестью молекулами кислорода, давая в итоге шесть молекул углекислого газа и шесть молекул воды. С точки зрения интересов жизни на Земле главное тут — высвобождение углерода в виде углекислого газа. А уж фотосинтезирующие организмы (то есть растения) могут, захватив этот углекислый газ, синтезировать из него гораздо более сложные углеродные соединения, пригодные для построения тел живых существ. В этом плане влияние человека на общепланетный круговорот углерода скорее положительно. Огромная масса углерода, которая сотни миллионов лет была “заперта” в пластах каменного угля, благодаря нашим шахтам, паровозам и тепловым электростанциям вновь пошла в дело.
Мы уже мимоходом упомянули, что бывают и другие, нецеллюлозные типы клеточных стенок. Еще один чрезвычайно распространенный в природе полисахарид — хитин, входящий в состав клеточных стенок грибов (наряду с полимерами глюкозы, которые там тоже есть). Кроме того, хитина много в наружных покровах некоторых животных, например насекомых, ракообразных и паукообразных. И грибов, и насекомых на Земле очень много. Потому и общая масса хитина на планете получается гигантской. Хитин — полимер, во многом похожий на целлюлозу. Он состоит из остатков бета-глюкозы, но только модифицированных. Дело в том, что хитин — это азотсодержащий полисахарид. Его мономером является, строго говоря, не сама глюкоза, а ацетилглюкозамин — производное глюкозы, где к одному из атомов углерода вместо гидроксила присоединена аминоацетильная группа –NH–CO–CH3.
Наконец, клеточные стенки бактерий состоят из еще более сложных азотсодержащих производных глюкозы, к которым дополнительно ковалентно “пришиты” цепочки аминокислот. Такой многокомпонентный полимер называется пептидогликаном. Самое интересное, что в состав пептидогликанов входят не только L-, но и D-аминокислоты. Это именно тот случай, когда D-аминокислоты в живых организмах все-таки присутствуют. В состав белков они, конечно, не входят и здесь, но в состав других соединений — в конце концов, почему бы и нет.
Мир, окрашенный по Граму
В 1884 году датский микробиолог Ганс Христиан Грам опубликовал новый метод окрашивания бактерий. Основой метода было применение сочетания органических красителей, главный из которых родствен по структуре обычным аминам (см. главу 1). Тут надо сказать, что окрашивание — это важно. Без окрашивания под микроскопом, как правило, толком ничего не рассмотреть. К тому же окрашивание должно быть стойким — чтобы не смывалось спиртами и другими растворителями при изготовлении препаратов, и, по возможности, дифференциальным — чтобы не красило все сплошь, ведь тогда в объекте, опять же, будет не разобраться. В общем, окраска объектов для микроскопии — это целая наука. В XIX веке, когда многое делалось наугад, изобретение нового красителя требовало как отличного знания химии, так и незаурядной интуиции.
Азотсодержащий краситель, предложенный Грамом, прекрасно действовал на бактерий. Но не на всех. Одних он исправно окрашивал в стойкий синий цвет, а на других почему-то вообще не держался — при промывке препарата они обесцвечивались. Так появилось разделение бактерий на грамположительных и грамотрицательных.
Умерший в 1938 году Ганс Христиан Грам, возможно, и сам не успел вполне осознать, насколько важную вещь он открыл. Обнаруженное им разделение бактерий по типу окрашивания оказалось признаком фундаментальнейших различий в строении клетки (см. рис. 6.6). У грамположительных бактерий снаружи от мембраны находится толстая пептидогликановая клеточная стенка. В этом плане их клетка более-менее похожа, скажем, на растительную, не считая того, что материал клеточной стенки другой. У грамотрицательных бактерий дело обстоит совершенно иначе. Их наружная оболочка включает две полноценные билипидные мембраны с тонкой пептидогликановой клеточной стенкой, расположенной между ними. Клеточная стенка грамотрицательных бактерий заключена между наружной и внутренней клеточными мембранами, как начинка сэндвича. Так не устроены никакие другие клетки.
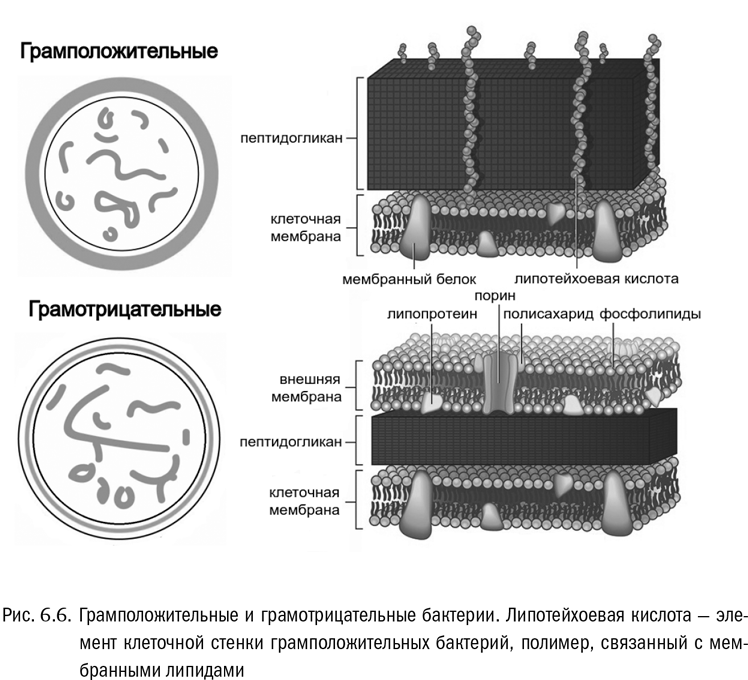
Есть гипотеза, что первые на Земле живые организмы были именно грамотрицательными бактериями, и только у их потомков вторая — наружная — мембрана исчезла. К сожалению, эта красивая идея слабо поддерживается молекулярно-биологическими данными, поэтому сейчас она не слишком популярна. Но независимо от того, верна она или нет, эволюционный зигзаг тут получился очень занятный.
Назад: 5. липиды и мембраны
Дальше: 7. нуклеотиды

