Книга: От атомов к древу. Введение в современную науку о жизни
Назад: 4. cимметрия
Дальше: 6. углеводы
5. липиды и мембраны
Господь Бог создал объем; поверхности же были изобретены дьяволом.Вольфганг Паули (цитируется по книге Манфреда Шредера “Фракталы, хаос, степенные законы. миниатюры из бесконечного рая”)
Липиды — удивительно разнообразная группа молекул. Они бывают и структурными “кирпичиками”, из которых строятся компоненты клеток, и питательными веществами, и гормонами. В общем, без знакомства с липидами невозможно разобраться в устройстве жизни — по крайней мере, жизни на Земле.
Но тут нас подстерегает затруднение. В отличие, например, от белков или углеводов, липиды не имеют никакой общей формулы. Их определяющее свойство — нерастворимость в воде, то есть гидрофобность (см. главу 2). Липиды — это сборное понятие, объединяющее все гидрофобные биологически активные вещества.
Напомним, что “гидрофобные” фактически значит “неполярные”, то есть включающие много углерода и водорода, но мало кислорода. В таких молекулах преобладают ковалентные неполярные связи, не создающие никаких локальных маленьких электрических зарядов. Поэтому они плохо взаимодействуют с водой, в молекулах которой связи как раз полярны и локальные маленькие заряды есть.
С химической точки зрения липиды бывают очень различны. Например, они вполне могут быть спиртами. Существует спирт, у которого единственная гидроксильная группа присоединена к огромному углеводородному радикалу с несколькими замкнутыми циклами (тремя шестичленными и одним пятичленным) и дополнительной длинной ветвящейся цепочкой. Этот спирт называется холестерином (см. рис. 5.1). Иногда холестерин переименовывают в холестерол — это синонимы. Холестерин очень гидрофобен, он не растворяется в воде и поэтому считается липидом.
Как и многие другие липиды, холестерин — важное питательное вещество. Из-за того, что он нерастворим в воде, он не может переноситься кровью в чистом виде (как, например, глюкоза), а переносится только при помощи специальных белков, образующих с ним комплекс.
Основу молекулы холестерина образует уже упоминавшееся ядро из четырех углеродных колец (трех шестиугольников и одного пятиугольника), которое в случае, если убрать из него двойную связь, будет называться великолепным словом “циклопентанпергидрофенантрен”. Производные циклопентанпергидрофенантрена называются стероидами. Это очень важная группа липидов. К стероидам относятся, например, половые гормоны и гормоны коры надпочечников. Довольно часто стероиды используются и как лекарства (каждый, кто смотрел сериал “Доктор Хаус”, это знает). В организме человека все стероиды синтезируются из холестерина — это одна из причин, почему он нужен нам как питательное вещество.
Другая важная группа липидов — жирные кислоты, то есть карбоновые кислоты с длинными (10–20 атомов углерода и больше) углеводородными “хвостами” (см. рис. 5.1). Чем длиннее “хвост”, тем хуже кислота растворяется в воде. Жирные кислоты, у которых в цепочке больше 12 атомов углерода, принято называть высшими.
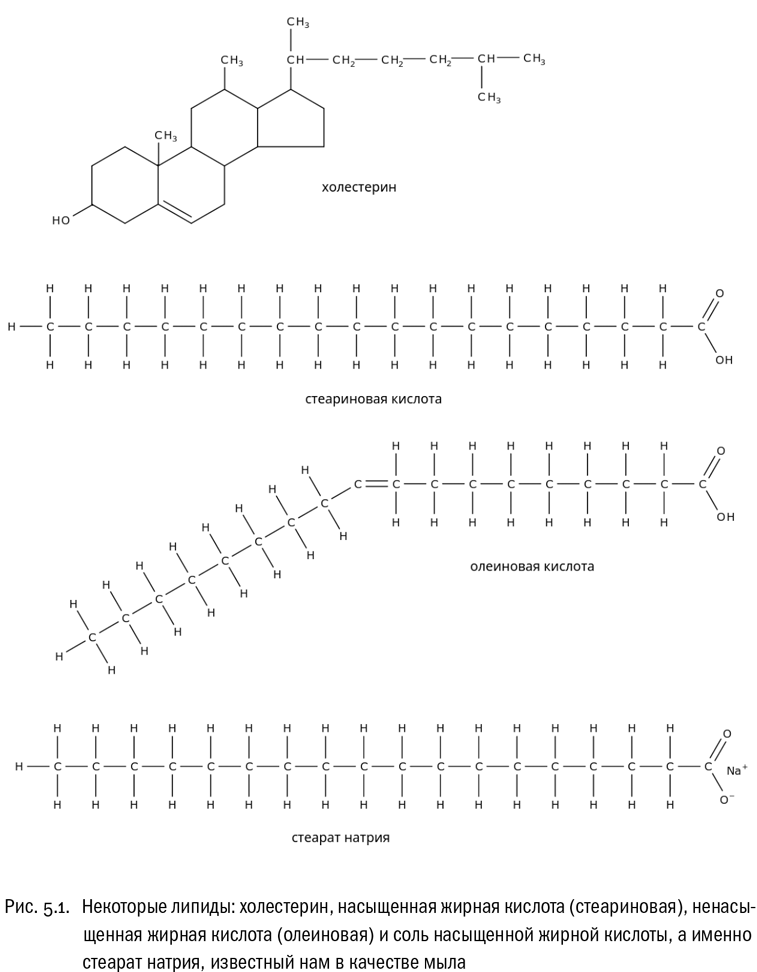
Жирные кислоты бывают насыщенными (без двойных связей в углеводородной цепочке) или ненасыщенными (с двойными связями). У насыщенных жирных кислот “хвосты” прямые, а у ненасыщенных — изогнутые в местах двойных связей. Из-за этого молекула может приобрести причудливую форму, особенно если двойных связей в ней несколько. Насыщенные высшие жирные кислоты при комнатной температуре — твердые вещества, а ненасыщенные — жидкости. Связано это с тем, что молекулы кислот с насыщенными “хвостами”, в которых нет создаваемых двойными связями изломов и изгибов, способны к более компактной упаковке. Особенно много ненасыщенных жирных кислот (и их производных) во всяких растительных маслах. Именно из-за этого, например, подсолнечное масло при комнатной температуре жидкое, в то время как сливочное — твердое.
Жирные кислоты играют важную физиологическую роль, которая иногда проявляет себя трагически. Например, одна из самых страшных болезней, разрушающих нервную систему, — адренолейкодистрофия — связана именно с нарушением обмена жирных кислот. Дело в том, что жирные кислоты активно используются при синтезе миелина — довольно сложного по составу вещества, образующего оболочку отростков нервных клеток. Строго говоря, миелин — это вообще не единое вещество, а смесь множества разных липидов. А миелиновая оболочка отростков нейронов необходима для нормального проведения по ним нервных импульсов (в физиологические детали этого мы сейчас вдаваться не будем). Так вот, в состав миелина входят жирные кислоты с углеводородными цепями, включающими по 16–20 атомов углерода — это по любым меркам довольно много. Но при адренолейкодистрофии в организме накапливается огромное количество так называемых очень длинноцепочечных жирных кислот (ОДЦЖК), имеющих углеводородную цепь длиной в 24–30 атомов. Вот это уже катастрофа. Очень длинноцепочечные жирные кислоты разрушают миелин, вместо того чтобы нормально в него встраиваться. Результат — расстройство буквально всех функций нервной системы, включая и движения, и чувствительность, и память, и рассудок. Обычно это приводит к смерти в течение нескольких лет. Адренолейкодистрофия — генетическая болезнь. Ее непосредственная причина — выход из строя одного определенного транспортного белка, в норме переносящего ОДЦЖК в те части клеток, где они должны расщепляться. Лечить такое медицина пока что не умеет, хотя можно надеяться, что со временем научится, особенно если о наличии у зачатого младенца гена адренолейкодистрофии будет известно заранее (а это вполне можно обеспечить).
Но вернемся к липидам. Есть липиды, которые являются с точки зрения химии сложными эфирами, то есть продуктами соединения карбоновой кислоты и спирта с общей формулой R1–CO–O–R2 (см. главу 1). Сложный эфир — это уже и не кислота, и не спирт, их свойства в нем взаимно уничтожаются. Сложный эфир, образованный спиртом и кислотой с длинными углеводородными цепями, называется воском. В молекулах восков так много атомов углерода и водорода и так мало атомов кислорода, что в итоге они очень похожи по свойствам на обычные углеводороды. К этой группе веществ относится, например, пчелиный воск, из которого пчелы делают соты.
Кроме того, воск (иной по составу, чем пчелиный) образует основу спермацета — жидкого вещества, находящегося в особом мешке в голове кашалота. Именно из-за спермацетового мешка голова кашалота выглядит прямоугольной, а не вытянутой, как у дельфина, хотя форма черепа у них очень похожая. По современным данным, спермацетовый мешок служит линзой для звуковых волн, с помощью которых кашалот ориентируется в пространстве; особенно это важно на большой глубине, где от зрения толку немного. Каждый, кто читал великий роман “Моби Дик”, знает, что раньше на кашалотов активно охотились ради спермацета, из которого делали ламповое масло, кремы, свечи и некоторые лекарства (например, противоожоговые мази). Сейчас добыча кашалотов, к счастью, запрещена.
Кроме стероидов, жирных кислот и восков есть еще по меньшей мере два типа липидов, без которых в биологии никак не обойтись. Это — жиры и фосфолипиды. С ними мы познакомимся чуть позже.
Детергенты
Жирные кислоты, а вернее их соли, с древних времен используются человеком в качестве моющих средств. Посмотрим, почему это так удобно.
Вот, например, стеариновая кислота: вещество с формулой CH3–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2–CH2––CH2–CH2–CH2–CH2–CH2–COOH, или попросту C17H35COOH. (Последний вариант, конечно, компактнее, но трудно отказать себе в удовольствии хоть раз написать эту формулу в развернутом виде.) Если заменить в этой кислоте атом водорода на атом натрия, получится соль — стеарат натрия C17H35COONa (см. рис. 5.1). В растворе такая соль легко диссоциирует, распадаясь на катион Na+ и анион C17H35COO–, который называется стеарат-ионом.
Как ведет себя стеарат-ион в воде? Его заряженная “головка” (–COO–) взаимодействует с водой отлично, а вот образующий большую часть молекулы углеводородный “хвост” — совсем никак. Этот “хвост” — воплощение гидрофобности, и он слишком длинный, чтобы заряженная “головка” могла затянуть его за собой в раствор целиком. Поэтому, если поблизости есть поверхность жидкости, то стеарат-ионы выстроятся по ней так, чтобы “головки” были направлены в воду, а “хвосты” — наружу, в сторону поверхностной пленки. Такое поведение молекул называется поверхностной активностью.
Иное дело, если на пути стеарат-ионов окажется капля какого-нибудь гидрофобного вещества — например, масла или жира. В этом случае стеарат-ионы выстроятся точно по поверхности, разделяющей воду и жир. Их “головки” будут обращены в воду, а “хвосты” погружены в жир. В результате капля жира будет разбита этими “хвостами” на мелкие капельки, которые по отдельности легко смоются водой. Вот почему стеарат-ионы и близкие к ним молекулы хороши в качестве моющих средств. Собственно говоря, именно так они и используются людьми последние несколько тысяч лет. Стеарат натрия — это не что иное, как обычное мыло.
Поверхностно-активные вещества, такие как мыло, часто называют детергентами (от латинского глагола detergere, одно из значений которого — “стирать”). Детергент — это вещество, в молекуле которого один конец растворим в воде, а другой — в липидах или углеводородах. Детергенты бывают анионными (с отрицательно заряженной “головкой”), катионными (с положительно заряженной “головкой”) или неионными (у которых “головка” полярна, но не заряжена). Но в любом случае молекула детергента обязательно включает гидрофильную “головку” и гидрофобный “хвост”. Мыло — это, как уже понятно из описания, типичный анионный детергент.
Великий химик Клод Луи Бертолле говорил, что “грязь — это вещество не на своем месте”. Тут можно уточнить: грязь — это, как правило, гидрофобное вещество не на своем месте. Оно и понятно: гидрофильные вещества вроде сахара без проблем смываются водой, так что никакие дополнительные вещества для их удаления не нужны. А вот для удаления гидрофобных веществ людям, собственно, и пришлось придумать моющие средства.
Бывают и природные аналоги моющих средств. Например, желчные кислоты — полярные производные холестерина, которые вырабатываются у человека печенью и выделяются в двенадцатиперстную кишку. Это самые настоящие детергенты, необходимые в данном случае для того, чтобы разбивать поступающие с пищей капельки жира.
Еще одно красивое название, применимое к детергентам, — амфифильные вещества. Слово “амфифильный” можно буквально перевести как “двояколюбивый”. Оно как раз и обозначает молекулу, одна часть которой “любит” воду, а другая — нет. Это более общий термин, чем “детергент”, буквально означающий все-таки именно “моющее средство” (тем более что далеко не все амфифильные вещества подходят на эту роль).
Возвращаясь к началу этой главы, мы теперь можем уточнить, что липиды на самом деле довольно редко бывают полностью гидрофобными. Чаще они амфифильны. Многие их биологические свойства именно с этим и связаны.
Жиры
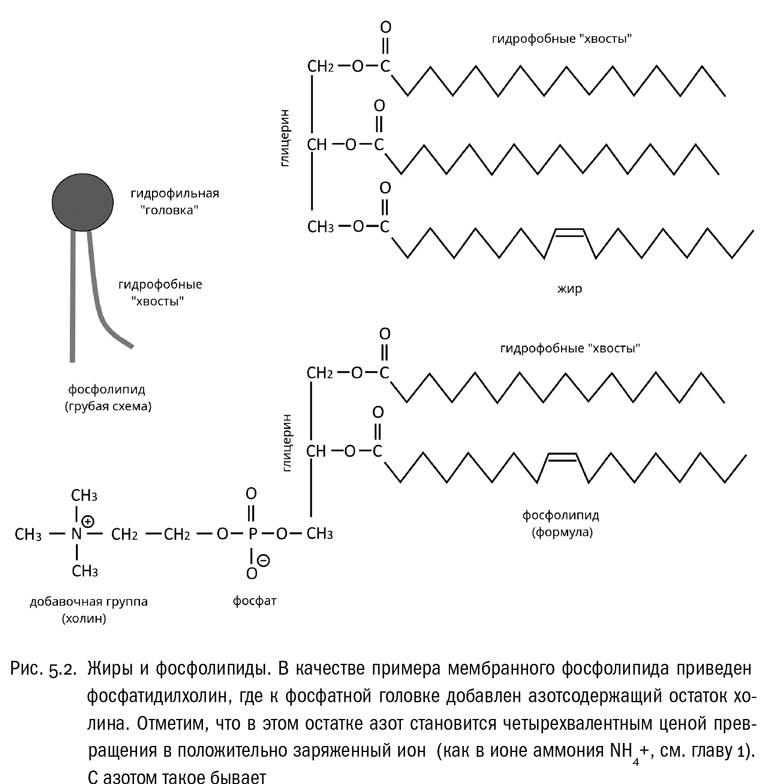
Теперь еще раз вспомним, что любая карбоновая кислота (в том числе и жирная) в принципе может образовать с любым спиртом сложный эфир. При этом от кислоты отщепится –OH, от спирта –H, они образуют воду, а остатки кислоты и спирта замкнутся в единую молекулу со сложноэфирной группой –CO–O– посредине (см. рис. 5.2). Спиртом, участвующим в этой реакции, вполне может оказаться и глицерин, у которого гидроксильных групп целых три (см. главу 1). Сложный эфир глицерина и трех жирных кислот называется жиром. Молекула жира имеет “головку” (остаток глицерина) и сразу три углеводородных “хвоста” (см. рис. 5.2).
На самом деле жиры стали известны людям гораздо раньше, чем их исходные компоненты. Например, желтый костный мозг, который наверняка извлекали древние люди из трубчатых костей крупных млекопитающих, — это в основном жир.
По опыту мы все знаем, что жир — это вещество животного или растительного происхождения, нерастворимое в воде, жирное на ощупь и оставляющее на бумаге характерные жирные пятна. Жиры, остающиеся при комнатной температуре жидкими, принято называть маслами.
Иногда в разговорах о химическом составе пищи понятие “липиды” для простоты заменяют понятием “жиры”. Теперь мы знаем, что это неточность. Жиры — и вправду ценные питательные вещества, но это далеко не единственные липиды, которые важны в этой роли. Например, холестерин — липид, но никакой не жир.
С участием насыщенных жирных кислот образуются насыщенные жиры, а с участием ненасыщенных кислот, соответственно, ненасыщенные. В растительных маслах гораздо больше ненасыщенных жиров, чем в животных. Хотя в целом и там и там есть и те и другие, отличается только их вклад.
Жиры — очень ценные источники энергии. Молекула жира может дать в два раза больше энергии, чем молекула углевода, имеющая такой же размер. Объясняется это вот чем. Процесс, путем которого мы получаем энергию из питательных веществ, — это, в сущности, окисление, то есть присоединение кислорода ко всем атомам водорода и углерода, до которых можно дотянуться. Все другие связи, образуемые этими атомами, при окислении разрываются, а его конечными продуктами становятся вода (H2O) и углекислота (CO2). Проблема в том, что в молекулах углеводов значительная часть атомов уже соединена с кислородом, так что окислять их дальше некуда (ну, или почти некуда). В молекулах жиров, где есть длинные жирнокислотные “хвосты”, таких атомов гораздо меньше. А потому и энергии из окисления жиров можно извлечь больше.
Кроме того, что жиры энергоемки, они еще и удобны для компактного хранения, поэтому животные (включая человека) часто используют их в качестве запасных веществ. Известно, что организм склонен реагировать на длительный стресс усилением отложения жира — это одна из причин так называемого стресс-индуцированного ожирения. Конечно, это эволюционно обусловленная реакция: с точки зрения нашего организма чем тяжелее и неопределеннее условия жизни, тем выше вероятность того, что запасные вещества в обозримом будущем пригодятся.
Мы уже знаем, что одним из конечных продуктов окисления питательных веществ является вода. Поэтому жировые отложения могут фактически служить запасом не только энергии, но и воды, которая все равно неизбежно выделяется при их переработке. Это особенно важно для пустынных животных вроде верблюдов. Горб верблюда содержит только жир, но при полном окислении этот жир (как и любой другой) превращается в углекислоту и воду. Углекислоту верблюд выдыхает, а воду оставляет в своем теле, чтобы добро не пропадало.
Некоторые тушканчики, тоже живущие в пустынях или полупустынях, запасают жир подобно верблюдам и в тех же целях, но не в горбе, а в хвосте. Они так и называются — толстохвостые тушканчики.
Фосфолипиды
А теперь познакомимся еще с одним важным для нас веществом — фосфорной кислотой, имеющей формулу H3PO4. Структура у нее довольно простая. Мы уже знаем, что валентность фосфора — 5 (см. главу 1). В фосфорной кислоте к атому фосфора присоединены четыре атома кислорода, один двойной связью, а все остальные одинарными. К этим последним присоединены атомы водорода. Когда фосфорная кислота находится в растворе, атомы водорода (вернее, протоны) могут отрываться, превращая фосфорную кислоту в анион. Здесь, как и раньше, надо иметь в виду, что “фосфорная кислота” и “фосфат” (то есть ее анион либо соль) в биохимии почти синонимы. В подавляющем большинстве случаев эти понятия свободно заменяются друг на друга. Очень часто название “фосфорная кислота” заменяют на “фосфат” просто для краткости.
Фосфорная кислота может участвовать в образовании сложных эфиров точно так же, как и карбоновые кислоты (см. рис. 5.2). Сложный эфир глицерина, двух жирных кислот и фосфорной кислоты называется фосфолипидом. Это — исключительно важный для биологии класс соединений. Можно сказать, что фосфолипид — это жир, у которого вместо одного из остатков жирных кислот тем же способом присоединен фосфат. Такая молекула состоит из гидрофильной “головки” (включающей остатки глицерина и фосфата) и двух гидрофобных “хвостов” (жирных кислот). При фосфате бывают еще добавочные боковые цепи, у разных фосфолипидов разные.
Два особенно широко распространенных фосфолипида — фосфатидилхолин и фосфатидилсерин. В фосфатидилхолине дополнительной боковой цепью при фосфате служит холин, небольшая азотсодержащая органическая молекула (см. рис. 5.2Г). А в фосфатидилсерине к фосфату присоединена аминокислота серин. В другие биохимические детали нам тут вдаваться не стоит, общие свойства фосфолипидов все равно гораздо важнее. А состоят они в том, что любой фосфолипид — это ярко выраженная амфифильная молекула, состоящая, если совсем уж попросту, из одной большой гидрофильной “головки” и двух длинных гидрофобных “хвостов”.
Знакомство с мембраной
Мы уже знаем, что никакие липиды не растворяются в воде. Что же будет, если их с водой все-таки принудительно смешать? Правильно: молекулы липидов начнут слипаться друг с другом своими гидрофобными частями, как бы защищая их от контакта с водой, и это называется гидрофобным взаимодействием. А гидрофильные части молекул будут, наоборот, втягиваться в воду, ориентируясь в сторону ее толщи. Это типичное поведение амфифильных веществ.
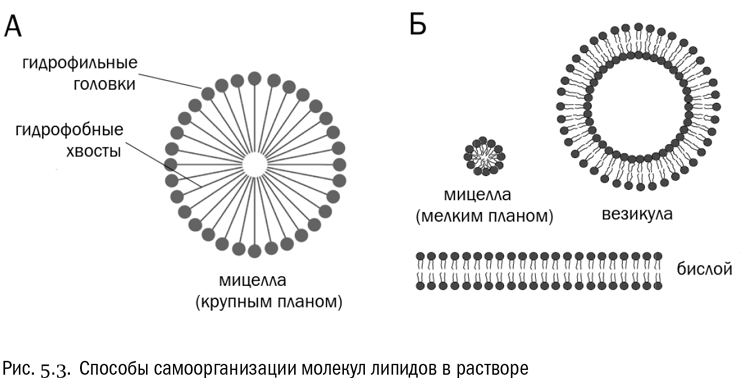
Будем для определенности называть гидрофильную часть любого липида “головкой”, а гидрофобную — “хвостом”. И мы увидим, что при смешивании с водой молекулы липидов могут группироваться тремя способами (см. рис. 5.3):
* мицелла — шарообразное скопление, где “хвосты” обращены внутрь, минимизируя контакт с водой, а “головки” — наружу. Ничего, кроме самих молекул липидов, мицелла не содержит. Мицеллы особенно легко образуются из молекул, где “головка” по диаметру превосходит “хвост”, то есть молекула имеет форму конуса. Таковы, например, молекулы жирных кислот;* бислой, в котором два слоя аккуратно выстроенных молекул липидов обращены “хвостами” друг к другу, а “головками” к водной толще. В этом случае гидрофобные области обоих слоев взаимодействуют друг с другом, не касаясь воды. Бислой легче всего образуется, если ширина “головки” и “хвоста” молекулы (или всех ее “хвостов” вместе) одинакова, то есть молекула цилиндрическая. Именно так устроены молекулы фосфолипидов;* везикула — пузырек, представляющий собой бислой, замкнувшийся в сферу. Такое замыкание происходит довольно легко, потому что края бислоя всегда неустойчивы — ведь гидрофобные “хвосты” там обнажаются. В везикуле же никаких свободных краев больше нет. Внутри везикулы находится полость, заполненная той же водой, что и снаружи.
Итак, жирные кислоты охотнее образуют мицеллы, а фосфолипиды — бислои, замыкающиеся в везикулы. Вот именно таким бислоем и является клеточная мембрана. Она состоит из двух слоев фосфолипидов, обращенных гидрофобными “хвостами” друг к другу. (На самом деле так устроена мембрана не абсолютно всех клеток, а только подавляющего большинства, но про исключения мы поговорим позже.) Как правило, клеточная мембрана не имеет никаких свободных краев, она полностью замкнута. То есть вся клеточная мембрана — это, в некотором смысле, одна сильно разросшаяся везикула.
Повторимся еще раз: типичная клеточная мембрана представляет собой фосфолипидный бислой. Это утверждение, которое могло бы показаться совершенно загадочным несколько страниц назад, сейчас нам уже понятно. И к нему можно добавить еще более очевидную мысль: там, где нет клеточной мембраны, нет вообще никакой клетки. Ведь говорить о клетке можно лишь тогда, когда есть четкая граница, отделяющая внутриклеточное пространство от внешней среды. Таким образом, история возникновения клеточных форм жизни — это в большой степени именно история возникновения липидных мембран.
Знакомство с мембраной (продолжение)
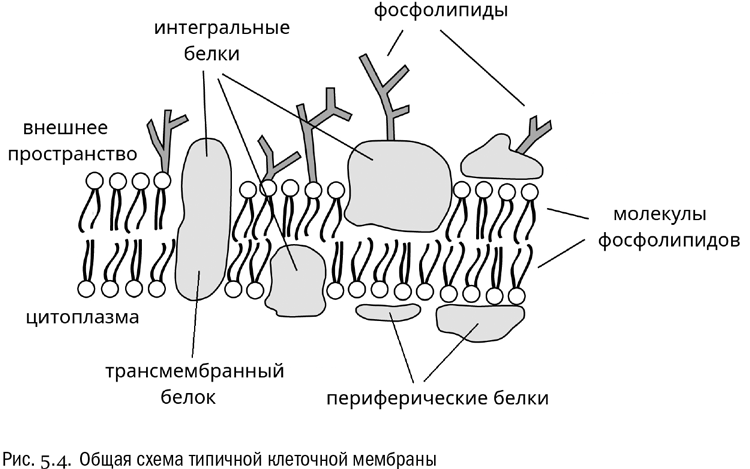
На самом деле биологические мембраны никогда не состоят из одних только фосфолипидов (см. рис. 5.4). Настоящая мембрана — это фосфолипидный бислой со встроенными в него многочисленными белками, которые называются интегральными. Многие (но не все) интегральные белки пронизывает клеточную мембрану насквозь, так что концы белка торчат из нее и внутрь, и наружу. Такие белки называются трансмембранными. Часть интегрального белка, погруженная глубоко в мембрану, всегда богата гидрофобными аминокислотами, такими как валин, лейцин, изолейцин и фенилаланин (см. главу 3). Радикалы этих аминокислот хорошо взаимодействуют с гидрофобными “хвостами” фосфолипидов. Именно благодаря этому интегральный белок и держится в мембране, в которую он вставлен. Очень часто интегральный белок бывает свернут таким образом, что его полипептидная цепочка пронизывает мембрану несколько раз подряд. Отрезками такого белка, непосредственно проходящими сквозь мембрану, чаще всего бывают альфа-спирали или бета-слои, почти целиком состоящие из гидрофобных аминокислот (о том, что такое альфа-спираль и бета-слой, см. опять же главу 3).
Интегральных белков очень много. Судя по современным молекулярно-биологическим данным, у большинства живых организмов от 20 до 30% всех белков — это интегральные мембранные белки. Неудивительно, что их функции разнообразны. Но чаще всего интегральные белки бывают или рецепторными (принимают сигналы из внешней среды и передают их внутрь клетки), или транспортными (переносят те или иные молекулы с одной стороны мембраны на другую). В типичной клетке человеческого тела есть, как правило, примерно 100 разных интегральных белков. Каких именно — во многом зависит от того, что это за клетка.
Очень важное свойство биологических мембран — избирательная проницаемость. Только очень немногие вещества могут проходить сквозь клеточную мембрану относительно свободно. Это или вода, молекулы которой совсем маленькие, или гидрофобные соединения вроде стероидных гормонов, которые легко “растворяются” в гидрофобном слое мембраны. Для всех остальных молекул требуются специальные переносчики. Например, молекулы углеводов — крупные и гидрофильные, поэтому самостоятельно пройти через мембрану они не могут. А поскольку клетке углеводы все-таки нужны, то существует целая группа трансмембранных белков, занимающихся их переносом. Особенно разнообразны транспортеры глюкозы — углевода, служащего у нас главным источником энергии (см. главу 6).
Более того, в конце XX века выяснилось, что специальные переносчики через мембрану есть даже для воды. Это интегральные белки, которые называются аквапоринами. Правда, перенос воды сквозь мембрану может идти и сам по себе, но аквапорины, во-первых, значительно облегчают его, а во-вторых, позволяют регулировать. Например, у человека есть не меньше 13 белков-аквапоринов, отличающихся друг от друга деталями функционирования. На такой важной задаче, как транспорт воды, эволюция не экономит.
Кроме интегральных белков есть еще и периферические, которые не пронизывают мембрану насквозь, а “прилипают” к ней только с одной стороны, нековалентно связываясь или с интегральными белками, или с головками фосфолипидов. Обычно периферические белки держатся или на ионных, или на водородных связях. Функции этих белков могут заключаться, например, в передаче сигнала от интегральных белков-рецепторов внутрь клетки. Неудивительно, что разных периферических белков довольно много.
С липидной частью мембран тоже не все так просто. Прежде всего, в нее входят не только фосфолипиды. Например, мембраны клеток животных содержат еще и холестерин, причем в довольно большом количестве. В растениях холестерина нет (поэтому встречающаяся на бутылках растительного масла надпись “без холестерина” — чистая правда), но в состав их мембран входят другие гидрофобные спирты, близкие к нему по структуре. Есть и еще несколько классов мембранных липидов, которые мы тут не обсуждаем, но которые тем не менее часто оказываются важными и для клеточной биологии, и даже для медицины.
Клеточная мембрана интересна тем, что для ее образования не нужно никаких ковалентных связей (которые в обычной химии как-никак главные). Она целиком держится на нековалентных взаимодействиях, в первую очередь, конечно, гидрофобных. Входящие в мембрану молекулы фосфолипидов, как правило, ничем не закреплены — они могут дрейфовать по своему слою, как в жидкости. Модель мембраны, учитывающая эти ее свойства, была в свое время достаточно красноречиво названа жидкостно-мозаичной. В живых системах вообще очень многое держится не на сильных связях (ковалентных), а на слабых (гидрофобных или водородных) — в следующих главах мы еще не раз это увидим.
Пенорожденная
Мы теперь знаем, что молекулу, в которой есть гидрофильная и гидрофобная части, для краткости называют амфифильной. При смешивании амфифильного вещества с водой оно может спонтанно собраться в мицеллы, а может и в везикулы (это зависит как от природы самого вещества, так и от физических условий). Если получаются везикулы — значит, молекулы в них выстроились в бислой, очень похожий на липидный бислой клеточных мембран. Можно экспериментально подобрать условия, в которых возникновение таких везикул ускорится. При этом искусственные везикулы могут “расти”, избирательно пропуская сквозь мембрану и удерживая в себе разные вещества, а могут и “делиться” наподобие клеток. Из-за того что липидные мембраны избирательно проницаемы, внутри везикул возникает среда, отличающаяся от окружающего раствора, в которой могут идти собственные химические реакции. В общем, получается, что простая самоорганизация смешанных с водой липидов внезапно дает свойства, привычные для живых систем: рост, размножение, обмен веществ, поддержание внутренней среды.
Из чего состояли первые мембраны? Вполне возможно, что поначалу их молекулярные компоненты были гораздо более простыми, чем фосфолипиды. В экспериментах химикам удавалось получать самые настоящие везикулы из смеси карбоновых кислот и сложных углеводородов, найденных в знаменитом Мурчисонском метеорите (см. о нем в главе 3). Древнейшие мембраны в принципе могли бы состоять, например, из жирных кислот. И действительно, заставить жирные кислоты с “хвостами” длиной примерно в 10 атомов углерода собраться в везикулы в искусственных условиях вполне можно. Тогда почему бы и не в природе? Правда, такие мембраны будут не слишком устойчивыми.
Подводя итог, тут можно сказать две вещи. С одной стороны, можно быть уверенным, что по части мембранной организации между живой и неживой природой нет никакого глубокого разрыва. Биохимическая эволюция вполне могла начаться с простой однослойной мицеллы и прийти к двуслойной везикуле, постепенно обретающей возможность сначала расти, потом делиться, а потом и захватывать крупные молекулы, в том числе способные нести генетическую информацию. Почти все промежуточные шаги на этом пути не только легко вообразить, но и можно уже сейчас воспроизвести экспериментально.
С другой же стороны, здесь, как это очень часто бывает в современной науке, обретенная ясность сразу же порождает новые вопросы. Мы совершенно не знаем, в какой именно момент были “изобретены” первые биологические мембраны и из каких молекул они поначалу состояли. Мембраны из жирных кислот слишком непрочны. Может быть, вначале были другие соединения, не такие сложные, как фосфолипиды, но обладавшие близкими свойствами? И если да — то что это были за соединения? Откуда они брались, в каких были изомерных формах? Как вышло, что фосфолипиды заняли их место? Подобных вопросов можно задать очень много. А когда на них найдутся ответы, перед исследователями наверняка встанут новые вопросы, которых мы сейчас еще даже не можем себе представить. Это — нормальный процесс познания.
Есть по крайней мере одно свойство жизни, совершенно невозможное без мембран (во всяком случае, в современных земных условиях). Это свойство — дискретность.
Мы привыкли к тому, что живое вещество разделено на маленькие самостоятельные порции, которые традиционно, со времен великих ученых XVII–XIX веков, называются клетками. Но обязательно ли любая жизнь должна состоять из клеток? Мы этого не знаем. Станислав Лем в “Солярисе” попытался вообразить иную жизнь — недискретную, охватывающую единым живым океаном целую планету. Конечно, это — фантазия. Но у нас нет никаких серьезных оснований считать, что она абсолютно нереалистична.
Так или иначе в истории жизни на Земле дискретность возникла, и притом очень рано. Сначала появились клетки, а потом и сложенные из них многоклеточные организмы, вплоть до дубов, кашалотов и людей. “Ключевая роль детергентов в формировании дискретных особей (в том числе и прекрасных) вполне соответствует представлению о возникновении Венеры (Афродиты) из пены морской”, — пишет по этому поводу известный биофизик Симон Эльевич Шноль. Слово “детергенты” тут употреблено как синоним “амфифильных веществ” (или “поверхностно-активных веществ”, если кому-нибудь такое название больше нравится). Сама же идея вполне актуальна. Ведь пена возникает там, где есть много амфифильных веществ — таких, как мыло, — которые легко образуют мицеллы, везикулы и вообще любые пузырьки. А принцип деления живой материи на клетки — точно такой же. Клеточная жизнь основана на явлении поверхностной активности. И древний миф здесь неожиданно точно иллюстрирует эволюционную реальность.
У корней древа жизни
Вся живая природа Земли делится на две части: организмы, состоящие из клеток, и вирусы. Вирусы мы тут пока не обсуждаем (о них пойдет речь в главе 12). А вот разнообразием клеточных организмов сейчас самое время поинтересоваться: мы как раз дошли до тех признаков, по которым они серьезно отличаются друг от друга.
В конце XX века микробиолог Карл Вёзе показал, что все клеточные организмы распадаются на три главные эволюционные ветви: эукариоты, бактерии и археи. Об этом открытии и его последствиях мы более подробно поговорим в главе 14, где будет обсуждаться система живой природы. Здесь нам достаточно самой простой вводной информации. Эукариоты — это обладатели клеточного ядра, среди которых есть и многоклеточные (животные, растения, грибы), и одноклеточные (амебы, жгутиконосцы, инфузории). Пока не был изобретен микроскоп, натуралисты волей-неволей ограничивались изучением одних лишь эукариот. Бактерии, в отличие от эукариот, не имеют клеточного ядра, и клетка их в целом устроена гораздо проще. Кроме того, они не бывают многоклеточными. Микроскопический мир бесчисленных бактерий, населяющих все природные среды, какие только есть на Земле, знаком ученым с XVIII века. И наконец, археи — это одноклеточные, не имеющие ядра и по устройству клетки сходные с бактериями, но сильно отличающиеся от них молекулярно-биологически. Многие (но далеко не все) археи живут в экстремальных средах — например, в почти кипящей воде или в растворах с высокой кислотностью.
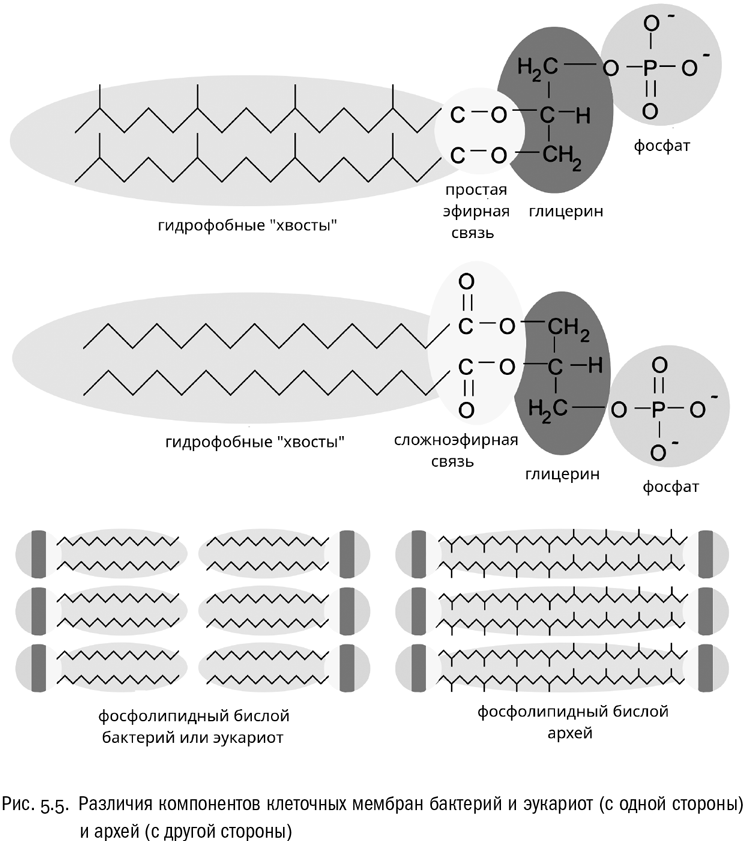
Одна из самых поразительных особенностей архей, отличающая их и от бактерий, и от эукариот, касается устройства клеточной мембраны. До открытия архей считалось, что мембрана, состоящая из двух слоев фосфолипидов, абсолютно универсальна и является всеобщим свойством клеточных организмов. Исследования архей начисто опровергли это представление. Сравнение химических компонентов эукариотных, бактериальных и архейных клеточных мембран показывает следующее (см. рис. 5.5):
* Архейные мембранные липиды представляют собой не сложные эфиры с общей формулой R1–CO–O–R2, а простые эфиры с общей формулой R1–O–R2 (см. главу 1). Надо заметить, что основу типичного мембранного липида в любом случае образует глицерофосфат, то есть сложный эфир глицерина и фосфорной кислоты. Но вот жирные “хвосты” у архей присоединены к нему совсем не так, как у всех остальных: не сложными эфирными связями, а простыми.* Углеводородные гидрофобные цепи мембранных липидов у архей ветвятся за счет множества торчащих в стороны метильных групп (–CH3). Ни бактериям, ни эукариотам это не свойственно.* Самое поразительное: у некоторых родов архей (и только у них) мембрана не двуслойная. Вместо бислоя она представляет собой единственный слой из молекул с двумя гидрофильными головками и длинной гидрофобной цепью между ними. Такие липиды иногда называют биполярными.
Как это объяснить? Биохимики считают, что все перечисленные химические особенности мембран полезны для жизни в экстремальных условиях — например, при высокой температуре или высокой кислотности. А мы уже знаем, что многие современные археи как раз в таких условиях и живут. Значит, налицо просто далеко зашедшее приспособление?
Увы, не все так просто. У архейных мембран есть еще одна важнейшая особенность. Дело в том, что у бактерий с эукариотами и у архей для синтеза мембранных липидов используются разные стереоизомеры глицерофосфата. У большинства живых организмов в мембраны входит L-глицерофосфат, но у архей — почему-то D-глицерофосфат. И вот это уже гораздо труднее объяснить приспособлением к каким бы то ни было внешним условиям. Мы ведь знаем, что на “обычные” физико-химические свойства веществ стереоизомерия практически не влияет. С точки зрения выживания при высокой температуре, кислотности или солености абсолютно неважно, какой стереоизомер глицерофосфата выбран для мембран. К тому же показано, что мембрана, включающая оба изомера одного и того же фосфолипида, будет физически неустойчивой, — то есть переходные состояния тут маловероятны. А ферменты, взаимодействующие с разными стереоизомерами мембранных липидов, отличаются друг от друга настолько сильно, что проще всего предположить их совершенно независимое происхождение.
Как может выглядеть эволюционный сценарий, сводящий воедино все эти факты? Тут допустимы самые смелые предположения. Может быть, общие предки бактерий, эукариот и архей вообще не имели никакой мембраны, то есть еще не были клетками? Или мембрана у них была, но не липидная, а неорганическая, например железо-серная? С другой стороны, современные генетические данные позволяют считать, что у общего предка всех клеточных форм жизни уже было несколько интегральных белков, приспособленных к работе в мембране и бесполезных без нее. Тогда получается, что какая-то мембрана там все же была. В любом случае этот узел еще далеко не распутан.
Назад: 4. cимметрия
Дальше: 6. углеводы

