Книга: От атомов к древу. Введение в современную науку о жизни
Назад: 13. земля и жизнь
Дальше: 15. мир эукариот
14. рождение системы
Моя цель — просто набросать здесь проект систематики китообразных. Я архитектор, а не строитель. Но это — грандиозная задача; простому сортировщику писем в почтовой конторе она не по плечу. Вслепую пробираться вслед за ними на дно морское, шарить руками в неизреченных основах, в плечевом и тазовом поясе самого мира — разве это не жутко?Герман Мелвилл. Моби Дик, или белый кит
Область биологии, занимающаяся разнообразием живых организмов, называется систематикой. Строго говоря, систематика — это абсолютно неустранимый аспект биологического знания как такового. Любое живое существо, изучаемое любым биологом с любыми целями, обязательно имеет какую-нибудь систематическую принадлежность. Другое дело, что есть (и никогда не переведутся) биологи, у которых систематика сама по себе является главной областью интересов. Часто говорят, например, что такой-то человек — величайший знаток систематики жуков-долгоносиков, или хищных динозавров, или сумчатых грибов, или еще кого-нибудь.
Основным понятием систематики является таксон — группа организмов, обладающая заданной степенью однородности. Таксон — это единица системы. Любая система живой природы представляет собой некоторое сочетание таксонов. А любой таксон, в свою очередь, имеет четкие границы. Можно сказать, что природа делится на таксоны примерно так же, как территории континентов делятся на страны и провинции. И об их границах примерно так же спорят — хотя, как правило, и без применения аргументов, опасных для жизни. Все названия таксонов суть имена собственные. Недаром в старых книгах их было принято писать с большой буквы, а латинские названия пишутся так и сейчас. По правилам, заведенным еще в XVIII веке и соблюдаемым по сей день, любой таксон обязательно получает в первую очередь именно латинское название, а уж оно переводится (или не переводится) на современные языки.
Таксоны различаются рангом по возрастанию: вид, род, семейство, отряд, класс, тип, царство, домен. Ранг таксона обозначает его мощность как подмножества: чем выше ранг, тем эта мощность больше. Виды группируются в роды, роды — в семейства, семейства — в отряды, отряды — в классы, классы — в типы, типы — в царства, а царства — в домены. В общем, система живой природы от начала до конца иерархическая. Группы последовательных рангов вложены друг в друга. Например, класс млекопитающих включает в себя отряды грызунов, приматов, рукокрылых, парнокопытных, непарнокопытных, хищных и разные прочие. А отряд хищных, в свою очередь, включает семейства псовых, медвежьих, куньих, енотовых, кошачьих, гиеновых, виверровых и (опять же) некоторые другие. В общем случае таксон может состоять из скольких угодно таксонов подчиненного ранга, в том числе и из одного-единственного. Например, в отряде трубкозубов всего одно семейство, один род и один вид (если не считать ископаемых). Такие “вырожденные” таксоны называются монотипическими. В случае с трубкозубом удивляться этому не приходится: он настолько не похож ни на одно другое современное млекопитающее, что для него пришлось создать собственный отряд, состоящий из одного вида.
Часто бывает, что систематикам не хватает основных рангов и они создают промежуточные: подтип, надкласс, подкласс, надотряд и т.п. Но тут действует простое правило: любой полностью систематизированный организм должен быть обязательно отнесен к какому-нибудь таксону каждого из основных рангов (от вида до домена включительно), а вот промежуточные ранги не обязательны.
Кроме того, в некоторых областях биологии есть особые исторические традиции: например, в ботанике отряд принято называть порядком, а тип — отделом. Сути дела это не меняет.
И наконец, в современной биологии встречается совершенно иной подход к построению системы — безранговый (хотя иерархический принцип и в этом случае строго соблюдается). О таких системах мы поговорим позже, в главе 15.
Биологи всегда стремились создать систему, максимально устойчивую к введению в нее новых объектов или признаков. Такая система называется естественной. Чем меньше меняется система при добавлении новых фактов, тем она естественнее. Идеальная естественная система отличается от любой другой системы тем, что ее невозможно оптимизировать: она уже наилучшим образом отображает отношения между объектами, и любые поправки ее только испортят. Стремление к такой системе, конечно, подобно бесконечному “асимптотическому приближению к абсолютной истине”, математические свойства которого изучали некоторые второстепенные персонажи Стругацких. Тем не менее эта цель по-своему привлекательна. История биологии — по крайней мере, ее систематических областей — это, по сути, и есть сильно растянувшаяся во времени история попыток построить полную естественную систему живых организмов.
Первая такая попытка связана с именем Аристотеля. Это неудивительно: от Аристотеля начинается история большинства естественных наук и уж во всяком случае — биологии и физики. Аристотель вообще уделял очень много внимания исследованиям природы. Этим он отличался от своего учителя Платона, для которого идеалом науки была геометрия и который, насколько можно судить, считал полноценным знанием только то, что хотя бы в принципе можно вывести в уме из набора аксиом. “Естествознание — родная стихия аристотелевской мысли, особенно когда речь идет о живой природе”, — писал историк биологии Валериан Викторович Лункевич. И это не пустая фраза, а совершенно точное наблюдение. Способ мышления Аристотеля был мало приспособлен к математическим абстракциям, зато идеально — к изучению живых организмов, сложных, многообразных и развивающихся. Характерно, что Аристотель не внес никакого научного вклада в математику, хотя, безусловно, хорошо знал ее просто как образованный человек. А вот в зоологии он открыл настолько много, что его с полным правом считают основателем этой науки. Принимался он и за ботанику, но жизни не хватило на все (Аристотель умер в 62 года). И в результате крупнейшим ботаником античности стал его ученик Теофраст.
Система Аристотеля была проста и наглядна. Он разделил живую природу на три ступени: растения (живут, но не движутся), животные (живут и движутся), человек (живет, движется и мыслит). Естественно, при этом он сразу же столкнулся с так называемой проблемой переходных форм, которая всегда возникает при попытке классифицировать непрерывную совокупность по качественным признакам. Дело в том, что античные исследователи знали о существовании организмов, которые (по тогдашним представлениям) неподвижны или малоподвижны, как растения, но могут питаться, как животные. В основном это некоторые морские существа: губки, коралловые и гидроидные полипы, актинии, голотурии. Аристотель предполагал, что в этих “промежуточных” организмах растительная природа смешана с животной. Более поздние античные авторы стали называть их зоофитами, то есть “животными-растениями”. Так что de facto в аристотелевской системе живой природы не три, а четыре категории: человек, животные, зоофиты и растения.
Созданная в середине XVIII века система Карла Линнея принципиально отличается от системы Аристотеля в двух отношениях. Во-первых, она состоит из вложенных друг в друга групп, которым присвоены ранги: царство — класс — отряд — семейство — род — вид (ранги типа и домена были добавлены позже). Система Аристотеля тоже иерархична, но четкого понятия о рангах там нет. Это изобретение Линнея. Животные и растения у Линнея имеют ранг царств.
Во-вторых, человек у Линнея не образует особую категорию, равноправную животным и растениям, а входит в царство животных — отныне и навсегда.
Cистема Линнея претендовала на то, чтобы охватить не одну лишь живую природу, а вообще все объекты во Вселенной. Кроме животных и растений там есть третье царство — минералов. Эта особенность линнеевской системы сейчас интересна только как факт истории культуры. Или, может быть, альтернативной истории науки, которая не осуществилась в нашей реальности.
Справедливости ради надо сказать, что распространение систематики на всю природу (и живую, и неживую) не было личной причудой Карла Линнея, а отражало совершенно нормальное для натуралистов XVIII века отношение к вещам. Мир объектов естественной истории виделся им единым целым. Были, например, авторы, которые добавляли к трем линнеевским царствам четвертое и пятое — царство воды и царство огня. А уж царство минералов признавали все. Причем натуралисты тех времен умудрялись находить переходные формы между всеми тремя общепринятыми царствами — растений, животных и минералов. Они писали не только о зоофитах (“животных-растениях”), но и о литофитах (“растениях-камнях”), и о литозоях (“животных-камнях”); к последним относили, например, коралловых полипов, образующих известковые постройки. Эти взгляды продержались до второй половины XIX века, когда победа эволюционной идеи в биологии и — с другой стороны — стремительное развитие наук о Земле сделали единую классификацию живых и неживых объектов практически бессмысленной.
Главным отличием животных от растений Линней считал активную подвижность. Он писал: “Естественные тела, наделенные всегда одной и той же формой и способностью к перемещению, называются животными; имеющие всегда одну и ту же форму, но не способные перемещаться — растениями; а те, которые имеют разнообразную форму, называются минералами”. Аристотелевских “зоофитов” Линней отнес к царству животных (правда, после долгих колебаний): он понимал, что некоторая подвижность, основанная на чувствительности животного типа, у них все же есть.
Однако проблема переходных форм никуда не делась. Во-первых, есть подвижные растения, в том числе и хищные — вроде росянки, которая ловит насекомых своими листьями. Во-вторых, есть совершенно неподвижные животные — например, губки. Найти у них хоть какие-то сокращающиеся элементы смог только современник Линнея англичанин Джон Эллис: путем тщательных наблюдений он выяснил, что губка может сокращать и расслаблять маленькие отверстия, сквозь которые в ее тело затекает вода. И в-третьих, есть еще и грибы, которые один ботаник, бывший на поколение старше Линнея, в сердцах обозвал произведениями дьявола, призванными нарушать гармонию природы и приводить исследователей в отчаяние (современные специалисты по грибам — микологи — обожают это цитировать). Положение грибов в линнеевской системе было крайне туманным. На первый наивный взгляд это растения. Но у грибов есть особенности, резко отличающие их от типичных растений, — например, независимость от света и отсутствие ветвления (на самом деле микроскопические нити, из которых состоит тело гриба, ветвятся, но Линней этого не знал). Неудивительно, что отнести грибы к растениям Линней решился лишь после больших сомнений, приписав комментарий: “Порядок грибов, к позору ботаники, и поныне представляет собою хаос”.
Проблему переходных форм не раз пытались решить, выделив третье царство живого. Но вот что это должно быть за царство? Тут разные ученые рассуждали по-разному. Ботаник барон Отто фон Мюнхгаузен, современник и хороший знакомый Линнея, в 1766 году предложил создать промежуточное царство — по-немецки Das Mittelreich, куда вошли грибы, а вместе с ними коралловые полипы и другие прикрепленные животные. Это была первая трехцарственная система живой природы, долго не продержавшаяся.
Другую идею опубликовал в 1783 году ботаник Ноэль Мартен Жозеф де Неккер. Он предложил выделить в третье царство только грибы. Это было куда более разумное решение. К 1780-м годам исследователи, в общем, успели убедиться, что кораллы, гидры и губки — не переходные формы между царствами, а просто животные. А вот с грибами ситуация была в точности противоположной. Натуралисты, решившие ими всерьез заняться, обычно довольно быстро приходили к твердому убеждению, что грибы — никак не растения. Неккер был первым из таких натуралистов, но далеко не последним.
Тем удивительнее, что признание этих фактов широкими массами ученых задержалось аж на 200 лет. Почти до конца XX века во многих учебниках ботаники грибы по традиции продолжали числиться растениями, хотя к тому времени уже все понимали, что с зелеными растениями у них нет ничего общего. Кроме банальной инерции мышления, никакой серьезной причины тут не найти.
Следующий шаг в развитии системы живой природы был связан с появлением клеточной теории. Согласно этой теории, клетка есть элементарная единица живого. В 1838–1839 годах ботаник Матиас Шлейден и зоолог Теодор Шванн убедительно показали, что это относится и к растениям, и к животным: организмы и тех и других состоят из клеток. Надо сказать, что у Шлейдена и Шванна, безусловно, были предшественники, но авторами клеточной теории по традиции считают именно их, потому что они первыми рискнули провозгласить эту теорию громко и четко — и выиграли. Для нас это важно вот почему. К XIX веку было известно уже довольно много микроскопических живых существ — инфузорий, амеб, жгутиконосцев, радиолярий и других. Их начал исследовать еще великий Антони ван Левенгук, современник Исаака Ньютона. Но долгое время натуралисты считали, что инфузории и амебы — это просто очень маленькие животные, и все тут.
Надо сказать, что животные такого размера действительно существуют (коловратки, например). Но подавляющее большинство микроскопических живых существ отличается от крупных животных гораздо серьезнее. Животные — многоклеточные организмы, а инфузории, амебы, жгутиконосцы и радиолярии — одноклеточные. На то, чтобы понять это, у исследователей ушло полтора века. Первым, кто четко отделил одноклеточные организмы от многоклеточных, был немецкий зоолог Карл фон Зибольд. В 1845 году он выпустил свою “Сравнительную анатомию беспозвоночных”, в которой “одноклеточные животные” (как их с этого момента стали называть) были отнесены к особому таксону Protozoa. В русских учебниках зоологии название Protozoa обычно переводилось как “простейшие”. По рангу это был (у разных авторов) или тип, или подцарство.
Карл фон Зибольд выделил четыре группы простейших: жгутиконосцы, корненожки, инфузории и споровики. Жгутиконосцы — это одноклеточные существа, плавающие с помощью жгутиков. Корненожки передвигаются с помощью ложноножек, временных выростов клетки, которые можно выпускать и втягивать; их типичный представитель — обыкновенная амеба. Инфузории покрыты ресничками (которые устроены точно так же, как жгутики, но их не одна-две, а тысячи) и имеют два клеточных ядра. А споровики — это чисто паразитические простейшие со сложными жизненными циклами; к ним относится, например, возбудитель малярии. Эту классификацию, по тем временам новаторскую, а по нынешним немыслимо архаичную, многие читатели наверняка еще застали: в школьных учебниках зоологии она фигурировала как минимум до 1980-х годов.
Заодно тот же Зибольд предположил, что главный признак, отличающий животных от растений, — не подвижность или чувствительность, а тип питания. Растения питаются автотрофно, животные — гетеротрофно (Зибольд еще не знал этих терминов, но суть различия понимал правильно). Иначе говоря, растения самостоятельно синтезируют сложные молекулы из углекислого газа и воды, а животные получают готовые питательные вещества из тел других организмов (живых или мертвых). Механизмом автотрофного питания у растений служит фотосинтез. Для него нужен зеленый пигмент — хлорофилл, который содержится в специальных частях растительной клетки — хлоропластах. Некоторые растения, в том числе и цветковые (петров крест, подъельник, повилика), лишены хлорофилла и могут питаться только гетеротрофно, но это эволюционно вторично: все они произошли от зеленых предков.
Куда более серьезной проблемой для систематиков стали грибы, которые строго гетеротрофны и не имеют хлорофилла никогда. Чтобы все-таки отнести их к растениям, ботаникам пришлось придумывать для царства растений дополнительные общие признаки: характер роста, способность всасывать питательные вещества всей поверхностью тела и размножение неактивными зачатками (семенами или спорами). Выделить грибы в самостоятельное царство подавляющее большинство биологов еще долго не решалось.
Но сложнее всего было разобраться в одноклеточных организмах. Чем подробнее их изучали, тем больше “переходных форм” среди них обнаруживалось. Например, есть автотрофные одноклеточные, у которых прекрасно выражена активная подвижность, — растения они или животные? Есть одноклеточные, у которых автотрофное питание сочетается с гетеротрофным (таких называют миксотрофами). И наконец, есть одноклеточные, которые могут довольно легко менять тип питания с автотрофного на гетеротрофный или наоборот. Так стало ясно, что в мире одноклеточных организмов провести границу между растениями и животными очень трудно.
В результате в 1860-х годах сразу несколько исследователей предложили объединить всех одноклеточных в собственное царство, отдельное и от растений, и от животных. Знаменитый немецкий биолог-эволюционист Эрнст Геккель в 1866 году назвал это царство Protista (протисты). Трехцарственная система Геккеля выглядела вполне разумно: есть растения, есть животные, а есть протисты, к которым относятся одноклеточные предки как растений, так и животных.
Справедливости ради надо сказать, что первым одноклеточных в самостоятельное царство выделил не Геккель, а английский натуралист Джон Хогг. Еще в 1860 году он предложил создать царство под названием Protoctista (буквально “первично созданные”). В него вошла примитивная (по определению Хогга) жизнь, слабо дифференцированная на растительную и животную. Интересно, что в системе Хогга это царство было не третьим, а четвертым — в придачу к животным, растениям и минералам. Дальше мы будем учитывать только царства живой природы и — соответственно — называть систему из животных, растений и протоктистов (или протистов) трехцарственной, но надо учитывать, что для Хогга и многих натуралистов его времени это было не так.
Протоктисты Хогга и протисты Геккеля — очень близкие понятия, но все же не синонимы. В XX веке название “протисты” обычно использовали исследователи, стремившиеся сузить объем этого царства, а “протоктисты” — исследователи, стремившиеся, наоборот, расширить его, включив туда некоторых примитивных многоклеточных. Например, многоклеточные зеленые водоросли не входили в царство Protista, но входили в царство Protoctista.
Были и другие попытки выделить третье царство, до которых докопались историки науки, но которыми мы не станем утруждать здесь читателя. В общем, по всему видно, что к началу 1860-х годов эта мысль уже носилась в воздухе. Эрнст Геккель просто высказал ее громче и убедительнее всех.
Однако биология продолжала развиваться, и cпустя несколько десятилетий стало ясно, что объединить всех одноклеточных в одно царство невозможно. Трехцарственная система Геккеля создавалась, когда о строении клетки еще было известно очень мало, поэтому она игнорирует такой важнейший признак, как клеточное ядро. У бактерий и синезеленых водорослей (которые в широком смысле тоже бактерии) ядер в клетках нет. У остальных, в том числе и у многих одноклеточных, — есть. В XX веке это было твердо установлено. В результате появились четырехцарственные системы, состоявшие из царств растений, животных, протистов и бактерий. Протистами теперь стали называться одноклеточные или колониальные (но не многоклеточные) существа с клеточными ядрами. На бактерий это название больше никогда не распространяли.
В 1925 году французский протистолог Эдуард Шаттон впервые предложил разделить живые организмы на две крупнейшие группы: эукариоты (с клеточными ядрами) и прокариоты (безъядерные). Шаттон не был теоретиком: он увлеченно изучал одноклеточных паразитов, инфузорий и водоросли, а общими вопросами интересовался мало. Свою классификацию он ввел мимоходом, просто для удобства, и опубликовал ее в виде краткого попутного замечания в работе, которую мало кто читал, кроме других протистологов. Неудивительно, что ее не сразу заметили. К тому же названия, предложенные Шаттоном, пишутся по-разному у разных авторов: или Prokaryota и Eukaryota, или Procaryota и Eucaryota. Первый вариант ближе к правильному написанию греческого слова карион (ядро), а второй — к версии самого Шаттона, который, как часто делают французские биологи, пренебрегал латынью и писал названия “с французским акцентом” — в данном случае Procaryotes и Eucaryotes. Сейчас правильным считается все-таки первый вариант — через k. Для любителей филологической точности можно добавить, что в исконно латинских словах буква k употребляется редко, зато она часто служит маркером проникающих в латынь греческих заимствований, что мы тут и видим.
В конце концов прокариоты и эукариоты получили ранг надцарств. В такой системе было одно царство прокариот (бактерии) и три царства эукариот — протисты, растения и животные. Стало считаться, что система живых организмов начинается именно с деления всего живого на прокариот и эукариот, а все остальные таксоны — всего лишь более мелкие подразделения внутри этих двух гигантских групп.
Тут нужна поправка: речь идет о системе всего живого, кроме вирусов. И прокариоты, и эукариоты состоят из клеток, а вот вирусы — нет. Поэтому в 1965 году зоолог Николай Николаевич Воронцов предложил ввести таксономический ранг империи, более высокий, чем ранг надцарства. В империю доклеточных (Non-Сellulata) входят только вирусы, а империя клеточных (Cellulata) делится на надцарства прокариот и эукариот. Это предложение, хотя и не прошло незамеченным, всеобщей поддержки не получило: большинство биологов в те времена еще сомневались, стоит ли вообще относить вирусы к живым организмам. В 2008 году, когда о вирусах стало известно неизмеримо больше, чем в 1960-е годы, близкую идею высказали французские вирусологи Дидье Рауль и Патрик Фортерр (см. главу 12). Они предложили разделить все живые организмы на рибосомокодирующие и капсид-кодирующие. Здесь и далее мы будем обсуждать исключительно систему рибосомокодирующих организмов, состоящих из клеток. Увы, для вирусов построить единое эволюционное древо просто невозможно — в первую очередь потому, что нет ни одного гена, который был бы общим абсолютно для всех вирусов и который можно было бы считать унаследованным от их общего предка (мы обсуждали это в главе 12). У клеточных организмов такие универсальные гены есть, и их немало.
Широкую известность в тех же 1960-х годах получили идеи американского эколога Роберта Уиттэкера, который заново пересмотрел всеобщую систематику, учитывая последние на тот момент научные достижения. Уиттэкер наконец-то выделил в самостоятельное царство грибы, поэтому царств стало пять: животные, растения, грибы, протисты и бактерии (см. рис. 14.1).
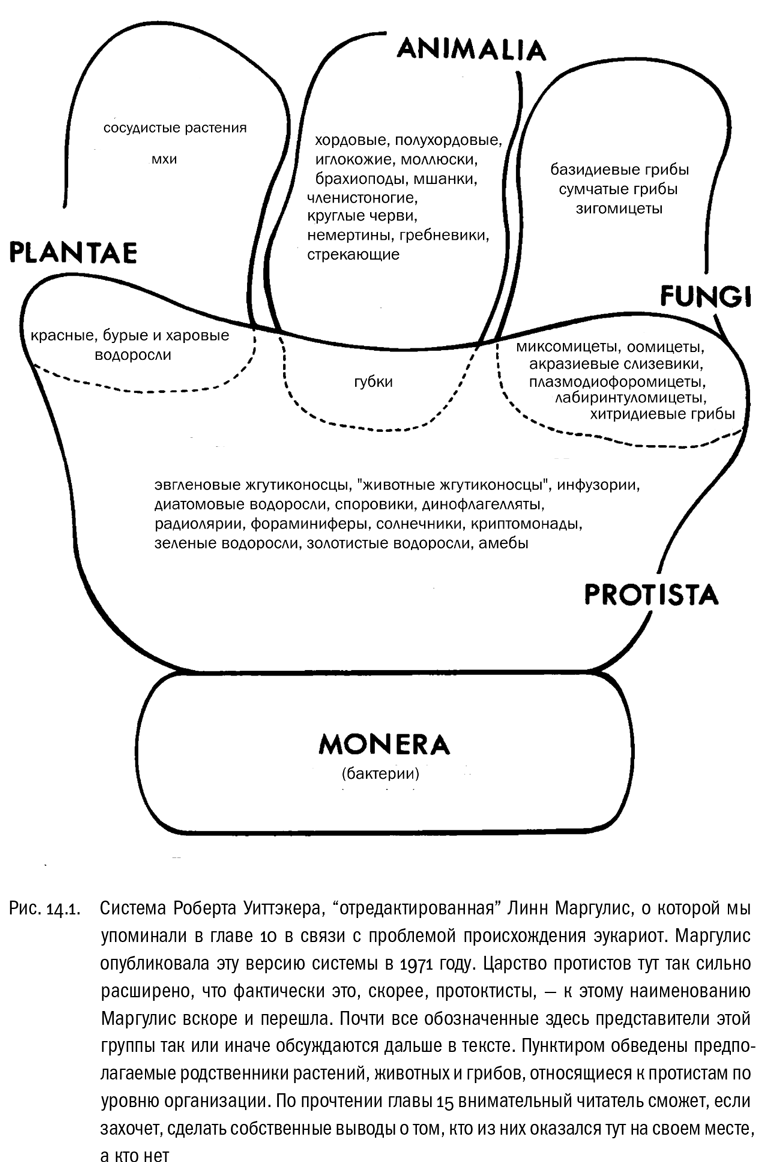
Главной проблемой такой пятицарственной системы оказались границы царства протистов (или протоктистов). Оно было чрезмерно разнородным, и ничего поделать с этим не удавалось никакими силами: решив одну проблему, систематики тем самым тут же создавали другую. Если остальные три царства эукариот хоть как-то соответствовали эволюционным ветвям, то царство протистов охватывало целый эволюционный уровень, в котором ветвей было множество. Фактически это царство было образовано методом исключения: в него заносили всех эукариот, которые не приобрели полноценной многоклеточности, а часто и некоторых многоклеточных с ними за компанию. В итоге реальный статус царства протистов резко отличался от статуса царств растений, животных и грибов. Это бросалось в глаза и не могло не создавать впечатления, что такая система внутренне нелогична.
В 1970-х многие биологи, в том числе и авторы университетских учебников, предпочли вообще ликвидировать царство протистов, сочтя его слишком сборным. Таким образом, царств опять стало четыре — бактерии, растения, животные и грибы. Последние три царства, по идее, должны были включить в себя всех одноклеточных родственников соответствующих организмов. В реальности такой подход привел к чудовищной путанице, которая продлилась до начала XXI века, а кое-где (например, в школьных программах) продолжается и по сей день. Сплошь и рядом на биологических факультетах одни и те же группы одноклеточных существ фигурировали и в курсах ботаники, и в курсах зоологии, причем в совершенно разном статусе. В результате студентам приходилось одновременно учить две противоречащие друг другу системы — зоологическую и ботаническую. Неудивительно, что в глазах большинства биологов это начисто обесценивало систематику как научную дисциплину. Ее стали рассматривать или как второстепенную и прикладную область знания (этакое составление инвентарной описи), или как чистейшую “игру в бисер”, слабо связанную с реальным миром. Бесконечные перестановки одних и тех же групп никого не вдохновляли. В общем, в систематике явно наступил кризис.
Ветви и домены
Кризис систематики был преодолен ценой ее почти полного поглощения другой биологической наукой — филогенетикой. Если систематика может (по крайней мере, в принципе) пользоваться любыми, сколь угодно разнообразными и необычными способами классификации живых организмов, то филогенетика изучает их родство, и только родство. Попросту говоря, филогенетика — это наука о том, кто от кого произошел.
До того как биологи признали эволюцию, фактор родства между организмами при построении систем не учитывался. Вернее, учитывался, но лишь в скрытой форме; то, что сходство организмов может отражать их самые настоящие родственные отношения, в додарвиновскую эпоху признавали очень немногие. Эта идея, для нас совершенно очевидная, тогда казалась слишком смелой и (как это ни странно) слишком упрощенной. Что касается естественных и искусственных систем, то о них стали рассуждать еще во времена Карла Линнея, но вот содержание этих понятий за прошедшие без малого 300 лет сильно поменялось. В XVIII веке естественной начали было считать систему, построенную по как можно большему числу признаков. Иногда оговаривалось, что признаки должны быть “существенными”, но определить, что это такое, толком никто не мог. А в XIX веке возникла идея, что естественная группа организмов — это родственная группа. Именно этому подходу следовал знаменитый немец Эрнст Геккель, когда создавал свою систему из трех царств. Слово “филогенетика” придумал тоже он, и не случайно. Геккель был твердо убежден, что естественная система живых организмов — это система, описывающая ход эволюции.
В XIX веке этой геккелевской идеи было достаточно для продуктивной работы, но в XX веке она стала нуждаться в уточнениях. Поэтому в середине XX века немецкий биолог Вилли Хенниг создал новое научное направление, которое назвал филогенетической систематикой. Современную версию хенниговской филогенетической систематики чаще всего называют кладистикой — от слова “клада”, придуманного на основе греческого корня и обозначающего эволюционную ветвь.
Главное требование филогенетической систематики состоит в том, что соответствие между эволюционным древом и иерархической системой организмов должно быть взаимно-однозначным. Иначе говоря, при правильно выбранном методе для данного эволюционного древа должно быть возможно построить только одну систему. Любой произвол в этом деле исключается.
Чтобы это требование выполнялось, Хеннигу пришлось ввести строгое правило: любая систематическая группа должна включать в себя только одну эволюционную ветвь, причем обязательно целиком. Последняя оговорка, на неискушенный взгляд малозаметная, на самом деле тут важнее всего. Из нее следует, что граница любого таксона может пересекать эволюционное древо только один раз — на “входе” в этот таксон. Вот в этом и состоит главный принцип хенниговской системы (см. рис. 14.2).
Нетрудно видеть, что царство протистов совершенно не соответствует этому условию. Растения, животные и грибы, без сомнения, произошли от разных одноклеточных эукариот. А все одноклеточные эукариоты по определению относятся к протистам. Получается, что граница царства протистов пересекает эволюционное древо как минимум четырежды: первый раз при происхождении протистов от прокариот и еще три раза — при происхождении животных, растений и грибов от протистов. Это типичный, прямо-таки образцовый пример таксона, запрещенного филогенетической систематикой.
А если попытаться классифицировать эукариот так, чтобы граница каждого царства пересекала эволюционное древо только один раз? Тогда система совершенно неминуемо окажется многоцарственной. Это показал в 1974 году английский ботаник Гордон Лидейл. Он честно попытался построить систему эукариот таким образом, чтобы каждое царство действительно заключало в себе ровно одну эволюционную ветвь (согласно тогдашним представлениям о родстве, конечно). В результате получилась система из 18 эукариотных царств, причем 11 из них состояли только из “протистов”.
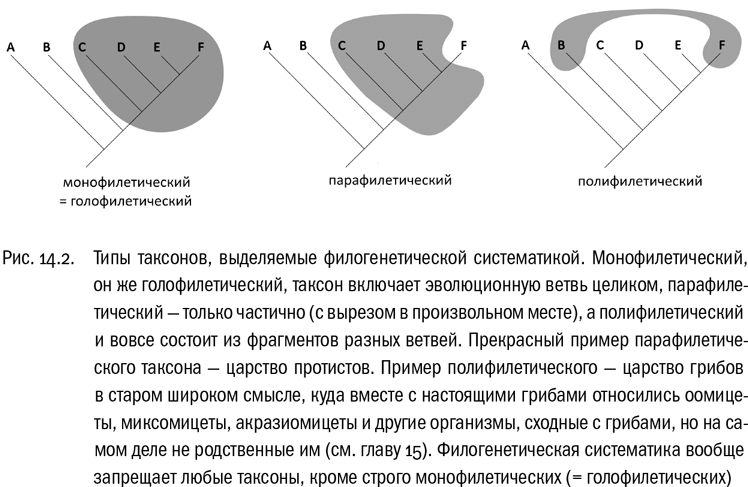
Современники в целом не приняли систему Лидейла всерьез, сочтя ее своего рода интеллектуальным чудачеством. А зря. Она была хотя бы внутренне последовательной. И главное, в 1970-х годах ничего более надежного все равно никто не предложил.
Как раз тогда, когда эти проблемы начали осознаваться, классическая система живой природы получила мощный удар с совершенно неожиданной стороны. Его нанесла молекулярная филогенетика — недавно (на тот момент) возникшая наука, изучающая родственные связи организмов путем прямого чтения аминокислотных последовательностей белков и нуклеотидных последовательностей генов.
Все началось с того, что американский микробиолог Карл Вёзе решил исследовать родственные отношения между разными группами прокариот. Для этого он нашел у разных бактерий (и заодно, для контроля, у нескольких представителей эукариот) один и тот же ген, кодирующий определенную молекулу, входящую в состав рибосом. Это была одна из молекул рибосомной РНК, сокращенно рРНК. Ее точное название — 16S рРНК у прокариот и 18S рРНК у эукариот (буква S тут обозначает некую константу, о которой нам сейчас достаточно знать, что она связана с размером молекулы). Рибосомы есть абсолютно во всех клетках. Их функции у разных организмов ничем не отличаются. И у холерного вибриона, и у крапивы, и у дрожжей, и у человека рибосомы делают только одно — синтезируют белки в точном соответствии с “инструкциями”, записанными на информационных РНК. Тогда логично предположить, что различия между разными организмами в “текстах” генов, обеспечивающих создание самих рибосом, могут быть только случайными. И чем больше этих случайных различий успело накопиться, тем дальше организмы друг от друга (иначе говоря, тем более давно жил их общий предок).
Результат исследования поразил Вёзе. Оказалось, что некоторая часть прокариот настолько сильно отличается от типичных бактерий, что никак не может входить в одно с ними царство. Так была открыта совершенно особая ветвь прокариотной жизни — архебактерии, которых вскоре стали называть просто археями (см. главы 5, 10).
У архей много абсолютно уникальных признаков. Например, мы уже знаем, что у бактерий (и у эукариот тоже) клеточные мембраны состоят из сложных эфиров, образованных L-изомером глицерофосфата, а у архей — из простых эфиров, образованных D-изомером глицерофосфата; есть там и другие химические отличия (см. главу 5). Эти типы мембран едва ли могли произойти один от другого. Значит, они возникли независимо. Какой была мембрана у общего предка бактерий и архей — загадка; маловероятно, что ее не было вообще, но она вполне могла быть, например, неорганической. Таким образом, открытие архей сразу же, просто в силу того, что эти существа стали известны, дает нам ценную информацию, проливающую свет на происхождение клеточной жизни. Это хороший пример того, как важен в биологии сравнительный метод.
Не менее поразителен был тот факт, что по ряду признаков археи оказались ближе к эукариотам, чем к бактериям. Например, архейные рибосомы больше похожи на эукариотные, чем на бактериальные. В архейных генах встречаются интроны — некодирующие вставки, характерные в основном для эукариот. В то же время устройство клетки у архей типично прокариотное: ни ядра, ни митохондрий, ни других видимых под микроскопом структур, характерных для эукариот, там нет.
Обдумав все эти данные, Вёзе решил, что деление на прокариот и эукариот попросту устарело. На самом деле множество всех клеточных организмов распадается не на две, а на три главные группы: бактерии, археи и эукариоты. Для этих групп Вёзе предложил новый таксономический ранг — домен (ниже империи, но выше царства). Таким образом, трехцарственную систему Геккеля через 100 лет сменила трехдоменная система Вёзе.
А как же теперь обстоит дело со старой классификацией, делившей клеточные организмы на прокариот и эукариот? А никак. Эти две группы не могут считаться равноправными: их статус совершенно различен. Эукариоты — действительно эволюционная ветвь, происходящая от одного-единственного предка (которым, скорее всего, была архея, вступившая в симбиоз с альфа-протеобактерией). И таксон “эукариоты” включает в себя эту ветвь целиком. Что же касается прокариот... но тут лучше начать с начала, чтобы было яснее.
Начнем с того, что все клеточные формы жизни имеют одного общего предка. Это следует в первую очередь из единства генетического кода, удивительное постоянство которого не спишешь ни на случайность, ни на параллельную эволюцию (она не могла повториться в разных ветвях настолько точно). А если у общего предка всех живых клеток был генетический код, значит, у него был и аппарат трансляции. И вообще он был достаточно сложным созданием. Этого общего предка сейчас принято называть LUCA — сокращение, которое чаще всего расшифровывается как last universal common ancestor (последний универсальный общий предок). Хотя, честно говоря, правильнее был бы вариант last universal cellular ancestor (последний универсальный клеточный предок), который предпочитает Патрик Фортерр. Так или иначе всех потомков “Луки” можно рассматривать как одну гигантскую эволюционную ветвь. В таком случае старая группа прокариот включает в себя лишь часть этой ветви: в нее входят все потомки “Луки”, в какую бы сторону ни пошла их эволюция, но почему-то за исключением одной специализированной веточки, которая называется эукариотами. Филогенетическая систематика категорически запрещает такие таксоны. Вывод: прокариоты — вообще не таксон, а название уровня организации, объединяющего всех тех потомков “Луки”, которые не приобрели уникального плана строения клетки, предусматривающего ядро и другие эукариотные признаки.
Ясно, что при этих условиях прокариоты никак не могут быть равноправны эукариотам. Недаром в системе Вёзе эукариотный домен называется не Eukaryota, а Eukarya. Такое название подчеркивает, что это не одна из двух равноценных групп, а нечто полностью уникальное.
Заодно эта история хорошо показывает, что научные истины никогда не стоит воспринимать как “высеченные в граните”. О том, что живые организмы делятся на прокариот и эукариот, в наше время слышали многие. Но какое место эта классификация реально занимает в истории науки? Давайте посмотрим. Термины “прокариоты” и “эукариоты” были придуманы Эдуардом Шаттоном, который употребил их в нескольких своих работах мимоходом — и все. Этих терминов тогда никто и не узнал, кроме людей, непосредственно общавшихся с Шаттоном; до самого начала 1960-х годов они встречаются в научных статьях очень редко. Кто же, наконец, провозгласил их на весь мир? Это сделали два известных микробиолога — Роджер Станье и Корнелиус ван Ниль. В 1962 году они выпустили обзорную статью под названием “Понятие бактерии”. Вот эта статья получила широкую известность, и система, делящая организмы на прокариот и эукариот, стала общепринятой именно после нее. Но уже в 1977 году Карл Вёзе и его коллеги ясно показали, что реальная структура древа жизни этой системе не соответствует. Таким образом “прокариотно-эукариотная” классификация на самом-то деле продержалась всего лишь 15 лет. До 1962 года о ней мало кто знал, а после 1977-го ее уже можно было с полным основанием объявить устаревшей. Иное дело, что инерция устаревших представлений сама по себе большая сила.
Надо добавить, что в домен архей входит не меньше пяти сильно различающихся групп, вполне заслуживающих ранга царств (эвриархеоты, кренархеоты, таумархеоты, корархеоты и другие, этот список сейчас продолжает пополняться). В домене бактерий подобных групп еще больше. Но выделение такого количества царств прокариот вызвало бы столько ненужной путаницы, что микробиологи предпочитают обходиться без него, переходя к безранговой номенклатуре. Дальше мы увидим, что и к царствам эукариот относится то же самое. В хенниговской филогенетической систематике ранги вообще не обязательны. Если они там сохраняются, то лишь по традиции и для удобства.
Асгард
Итак, есть три эволюционные ветви клеточных организмов — Archaea, Eukarya и Bacteria. Казалось бы, теперь картина ясна. Но, как это постоянно бывает в науке, полученные ответы сразу вызвали к жизни новые вопросы.
Эукариоты произошли от архей. Это твердо установленный факт (симбиотический компонент эукариотной клетки, происхождение которого однозначно бактериальное, мы сейчас временно не учитываем). Но что, собственно, значит “произошли от архей”? Насколько буквально это стоит понимать?
Одно дело, если эукариоты и археи — это две ветви, разошедшиеся от уникального для них общего предка. В этом случае их надо считать сестринскими группами, а вот предками или потомками друг друга называть, строго говоря, нельзя. И совсем другое дело, если эукариоты отпочковались от какой-то из групп архей уже после того, как древо самих архей разветвилось. Тогда эукариоты и вправду окажутся потомками архей в самом точном смысле этого слова. Но одновременно это будет означать, что эукариот надо считать не отдельным доменом, а группой внутри архей — так сказать, их подмножеством. По крайней мере этого в обязательном порядке потребует филогенетическая систематика. Иначе получится, что граница домена архей пересечет эволюционное древо дважды — при происхождении самих архей от “Луки” и при происхождении эукариот от архей; а такое, по современным понятиям, недопустимо (см. рис. 14.3).
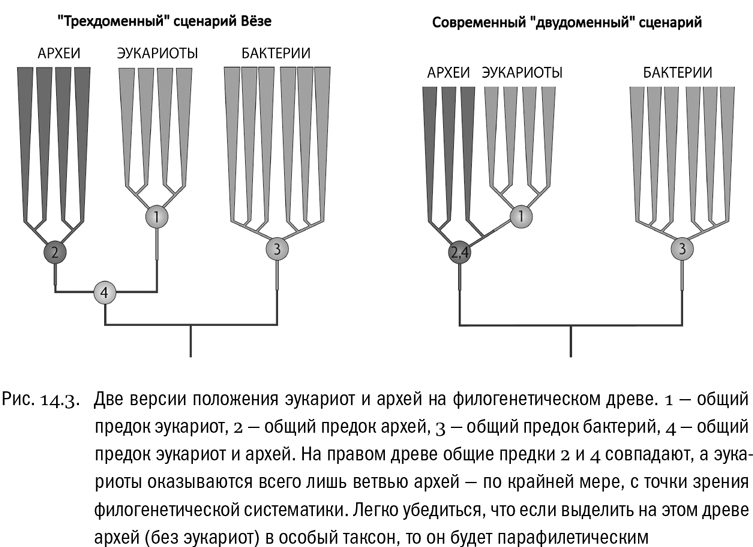
Тем не менее именно эта вторая возможность сейчас и подтверждается. По мере развития генетических исследований становится все яснее, что эукариоты эволюционно ближе к одним археям, чем к другим. И в свою очередь, некоторые археи ближе к эукариотам, чем друг к другу: например, кренархеоты, судя по современным филогенетическим древесам, доводятся куда более близкими родственниками эукариотам, чем каким-нибудь эвриархеотам,. Все это означает, что ветвь эукариот возникла довольно глубоко внутри архейного древа. Попросту говоря, эукариоты — это специализированная веточка архей. А в таком случае на смену трехдоменному древу жизни неизбежно должно прийти двудоменное.
Интересно, что система из трех доменов (бактерии, археи, эукариоты) в любом случае уже продержалась к настоящему моменту дольше, чем система из двух надцарств (прокариоты и эукариоты). Но похоже, что и она оказалась в развитии науки промежуточным этапом. Следующие шаги по обновлению системы уже предпринимаются.
В 2015 году в ходе глубоководных исследований Северного Ледовитого океана были открыты локиархеоты — археи, являющиеся ближайшими родственниками эукариот. Локиархеоты оказались генетически ближе к эукариотам, чем к любым другим археям, известным на тот момент. Название они получили в честь Локи, скандинавского бога огня. У локиархеот есть белки, очень близкие к актину — белку, из которого у эукариот образуются микрофиламенты, важнейшие составные части цитоскелета (см. главу 10). Причем генов актиноподобных белков у них несколько штук. Значит, эволюция этих белков началась достаточно задолго до появления эукариот. Кроме того, у них есть белки, работа которых связана с изменением формы клеточной мембраны и с рециклизацией мембран — процессом, совершенно необходимым клеткам, которые транспортируют вещества в подвижных вакуолях (см. опять же главу 10). Все это означает, что клетка локиархеот имеет много общего с эукариотной.
В придачу к локиархеотам были вскоре открыты еще три группы родственных им архей. Они тоже получили названия в честь скандинавских богов: одинархеоты, торархеоты и хеймдалльархеоты. И наконец, все эти четыре группы вместе были закономерно названы асгардархеотами.
Как это ни удивительно, на момент написания этих строк асгардархеот еще никто никогда не видел своими глазами (вернее, под микроскопом). Но генетического материала от них найдено и прочитано уже столько, что характеристика этих существ выходит достаточно детальной. Например, у одинархеот есть тубулин — белок, из которого у эукариот состоят микротрубочки. И других белков, которые раньше считались уникальными для эукариот, в клетках асгардархеот хватает. При этом асгардархеоты однозначно более близкородственны эукариотам, чем любым другим археям. Иными словами, существует эволюционная ветвь, которая включает асгардархеот, эукариот и больше никого. Более того, похоже, что сами эукариоты — это всего лишь веточка глубоко внутри эволюционного “куста” асгардархеот; например, хеймдалльархеоты генетически гораздо ближе к эукариотам, чем к торархеотам или одинархеотам. Как бы там ни было, эти данные очень здорово подкрепляют новую двудоменную систему жизни.
Ядерная интерлюдия
Сделаем отступление.
Открытие асгардархеот, бесспорно, величайшее достижение. Теперь мы знаем точный “адрес”, по которому на эволюционном древе можно найти нашего архейного предка. Значит ли это, что проблема происхождения эукариот решена?
Конечно, нет.
Проблема происхождения эукариот — одна из самых сложных и запутанных во всей эволюционной биологии. Мы уже обсуждали ее в главе 10, но довольно кратко (о некоторых гипотезах там вообще пришлось умолчать). Так что к этой теме не грех вернуться — хотя бы ненадолго.
Итак, чем, собственно, эукариотная клетка отличается от прокариотной? Во-первых, в ней есть сложная система внутренних мембран, образующая эндоплазматическую сеть и тесно связанное с ней ядро — ту самую структуру, которой эукариоты обязаны своим названием. Во-вторых, в ней есть митохондрии — или хотя бы их различимые на молекулярном уровне остатки. В-третьих, в эукариотной клетке перемешаны компоненты, имеющие совершенно разное происхождение — как бактериальное, так и архейное. Любой состоятельный сценарий происхождения эукариот должен объяснить эти три особенности их клеток. Причем желательно все разом — так, чтобы не приходилось вводить дополнительные предположения ad hoc (“для данного случая”).
Собственно говоря, объяснить перечисленные признаки по отдельности было бы гораздо проще. Приобретение митохондрий легко объясняется симбиозом: предок эукариот когда-то “проглотил” бактерию, ставшую предком митохондрий, и оставил ее внутри себя жить. Обилие в эукариотной клетке бактериальных генов и белков столь же легко объясняется горизонтальным переносом генов: предок эукариот жил в каком-то сложном бактериальном сообществе, вот и нахватался генов от соседей. Ну а ядро вполне могло возникнуть из впячивания плазматической мембраны, которое удачно отшнуровалось и замкнулось. Судя по всему, нечто похожее произошло в одной интересной группе грамотрицательных бактерий, которая называется планктомицетами. У них есть довольно сложная система внутренних мембран, окружающая ту область цитоплазмы, где находится ДНК, и у некоторых родов это выглядит почти как клеточное ядро. Вероятно, все внутренние мембраны планктомицетов развились из впячиваний их плазматической мембраны (точнее внутренней из двух мембран, свойственных грамотрицательным бактериям). Самое же любопытное, что планктомицеты — вовсе не родственники эукариот и возникновение аналога ядра наверняка произошло у них совершенно независимо. Но это как раз и убеждает, что такие события в принципе возможны.
Сейчас, однако, нам нужно несколько другое. Нам нужен эволюционный сценарий, который удовлетворительно объяснял бы все главные особенности эукариот — и ядро, и митохондрии, и генетическую “химерность”. Есть ли такой сценарий в нашем распоряжении?
Серьезные попытки его создать, безусловно, налицо. Ближе всего к идеалу сейчас, видимо, подошли знакомые нам из главы 10 гипотезы Евгения Кунина и двоюродных братьев Баумов,. Эти гипотезы отличаются друг от друга, но так или иначе обе они утверждают примерно следующее. Предком эукариот была некая архея, которая обзавелась наружным симбионтом в виде альфа-протеобактерии. Со временем альфа-протеобактерия оказалась окружена выростами архейной клетки, и наружный симбиоз превратился во внутренний. Одновременно с этим (и, вероятно, вследствие этого) возникло ядро. В этом месте гипотезы расходятся. Кунин полагает, что ядро образовалось из слившихся внутри клетки мембранных пузырьков, а Баумы высказали и вовсе парадоксальную идею: ядро эукариот соответствует “телу” архейной клетки, а цитоплазма — ее разросшимся выростам, которые охватили альфа-протеобактерий и слились друг с другом (см. ранее рис. 10.9). В любом случае изоляция архейного генома в ядре оказалась в итоге полезна, чтобы защитить его от чрезмерно активного проникновения генетического материала бактерий-симбионтов. Но такое проникновение все равно на первых порах происходило, и генетическая “химерность” эукариотной клетки объясняется именно этим.
Что ж, авторы этих гипотез сложили пазл почти до конца. Но все-таки лишь “почти” — потому что есть, например, еще такой бросающийся в глаза факт, как различие архейных и бактериальных клеточных мембран. Липиды, из которых состоят эти мембраны, могут создаваться на основе право- или левовращающего изомера глицерофосфата. Архейные мембраны состоят из “правозакрученных” липидов. Бактериальные — из “левозакрученных”. Разница между ними вполне фундаментальна. А теперь спросим себя: почему же в эукариотной клетке все мембраны (и плазматическая, и ядерная, и вакуолярные) относятся к бактериальному типу? Это парадоксальный факт, на первый взгляд вовсе не следующий ни из гипотезы Кунина, ни из гипотезы Баумов. Исходя из этих гипотез, можно было бы скорее ожидать обратного: ведь это архея охватила и поглотила бактерию, а не наоборот. Конечно, объяснить наблюдаемые факты при желании все равно легче легкого: мы же знаем, что большинство генов “домашнего хозяйства” (обеспечивающих всевозможные процессы обмена веществ) в эукариотных геномах имеет бактериальное, а не архейное происхождение; вот продукты этих генов и взяли на себя синтез компонентов мембран, а поскольку “правозакрученные” и “левозакрученные” молекулы в одной мембране лишь ограниченно совместимы, то архейные мембранные липиды в конце концов были полностью вытеснены бактериальными. И все бы хорошо — если не считать, что тут перед нами типичное объяснение ad hoc, не вытекающее само по себе из основной гипотезы, а добавленное специально, чтобы охватить не уложившиеся в нее факты. Хотя само по себе это и не значит, что оно неверно.
Тут нам будет полезно вспомнить историю науки. В старинной астрономии почетное место занимало понятие “спасения явлений” (во многом определявшее всю ее методологию). “Спасти” тот или иной факт значило привести его в согласие с единой теорией, которая, в свою очередь, должна была согласоваться со всеми остальными наблюдаемыми фактами. Иногда эта операция оказывалась очень непростой. Например, докоперниковская астрономия вынуждена была создать ради нее представление о движении планеты по эпициклу — маленькой окружности, центром которой является точка, движущаяся по большой орбите. Астрономический эпицикл стал настоящим символом вспомогательной гипотезы, ниоткуда не вытекающей и нужной лишь затем, чтобы свести концы с концами (то, что мы выше назвали “ad hoc”). В конце концов борьба за наилучшее “спасение явлений” привела к тому, что геоцентрическая система сменилась на гелиоцентрическую, позволившую от эпициклов отказаться. Что ж, европейская наука всегда, еще со времен Пифагора и Платона, руководствовалась идеей, что рациональное объяснение мира должно быть по возможности единым. На то она и европейская.
А не можем ли мы и в вопросе происхождения эукариот, образно говоря, поменять местами Землю и Солнце? Хотя бы из чисто хулиганских побуждений, чтобы посмотреть, что из этого получится. До сих пор мы исходили из идеи, что на заре эволюции эукариот бактерия (вероятно, альфа-протеобактерия) была захвачена и поглощена какой-то археей. А что, если наоборот: архея была захвачена и поглощена какой-то бактерией?
Обратившись к литературе, мы сразу увидим, что это отнюдь не фантазия на пустом месте. В последнее десятилетие XX века подобные гипотезы были удивительно популярны, да и сейчас у них есть осторожные сторонники . Существует целое семейство гипотез, согласно которым клеточное ядро имеет симбиотическое происхождение — от археи, встроившейся в клетку более крупной бактерии и окруженной ее сомкнутыми выростами.
Образцом такой гипотезы можно считать эволюционный сценарий, который опубликовали в 1996 году Радхей Гупта и помогавший ему Брайан Голдинг; у этих авторов есть полная аргументация и нет ничего лишнего. Гупта и Голдинг изучали эволюцию белков, сравнивая их аминокислотные последовательности. И они пришли к выводу, который в основном подтверждается и сегодня: примерно половина эукариотных белков восходит к археям, а другая половина — к грамотрицательным бактериям. Причем это соотношение настолько близко к равному, что классическая теория (согласно которой архейный предок эукариот всего лишь захватил бактериального симбионта как некий “предмет роскоши”) становится неубедительной.
Кроме того, это соотношение трудно объяснить простым горизонтальным переносом генов (даже при том, что интенсивность этого процесса могла быть в древние времена намного выше, чем сейчас). Такой перенос шел бы более-менее равномерно между всеми участниками сообщества. И если бы дело было только в нем, то предок эукариот “нахватался” бы генов от самых разных соседей-бактерий — и грамположительных, и грамотрицательных. Между тем Гупта и Голдинг обнаружили, что у эукариот очень много белков и генов, каким-то образом унаследованных от грамотрицательных бактерий, а вот от грамположительных не нашлось на тот момент ни одного. Конечно, эта избирательность не случайна, вопрос только в ее источнике.
В общем, состав эукариотного генома дает все основания полагать, что при возникновении эукариот клетки археи и грамотрицательной бактерии просто слились (см. рис. 14.4). Получилась единая клетка, сначала унаследовавшая геномы обоих “родителей” — Гупта и Голдинг прямо так их и называют. Но, поскольку два полнофункциональных генома клетке были не нужны, часть каждого из них исчезла, а между оставшимися частями произошло разделение функций. От архейного генома в эукариотной клетке остались в основном “информационные” гены, обеспечивающие работу самого генетического аппарата. А от бактериального генома — в основном “операционные” гены, обеспечивающие обмен веществ. Тогда будет вполне разумно предположить, что в химерной структуре (каковой эукариотная клетка в любом случае несомненно является) от археи произошло ядро, а от бактерии — цитоплазма вместе с наружной мембраной. И получается, что это именно архея проникла внутрь бактерии. Вероятно, сначала она жила во впячивании наружной мембраны бактерии — этаком “заливе”. Потом глубокие складки мембраны бактерии, со всех сторон охватившие архею, сомкнулись друг с другом, образовав внутреннюю мембранную систему — ядро и эндоплазматическую сеть. После этого плазматическая мембрана самой археи стала лишней и исчезла. Фактически от археи остался голый генетический аппарат, заключенный внутри системы бактериальных мембран. Ну а гены самой бактерии, сначала находившиеся снаружи от ядра, постепенно мигрировали в него (молекулярные механизмы, делающие возможной такую миграцию, известны). И получилась нормальная эукариотная клетка.
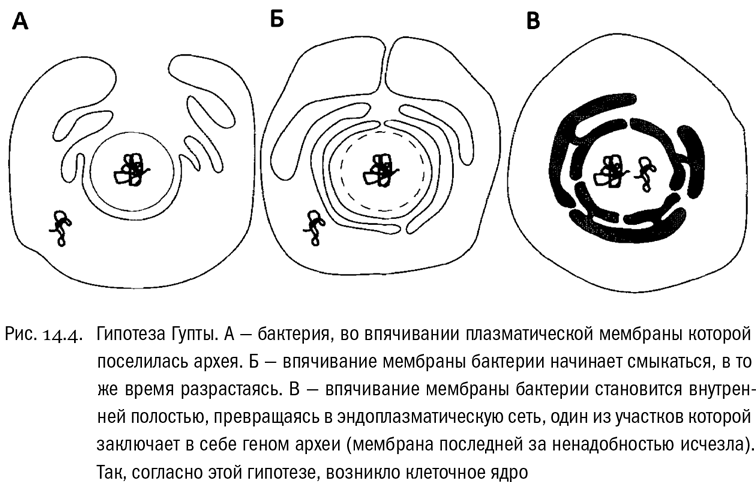
Интересно, что в гипотезе Гупты (назовем ее так для краткости, хотя ее поддерживали и другие ученые) есть кое-что общее с уже знакомой нам гипотезой Баумов. И там и там получается, что остаток исходной архейной клетки находится в основном в эукариотном ядре. Хотя приводящие к этому выводу сценарии прямо-таки полярно противоположны. Гипотеза Баумов, при всем ее бесспорном изяществе, по части состава мембран (архейные vs. бактериальные) дает предсказания, строго обратные тому, что наблюдается в действительности, и выйти из этого положения она может только путем добавления “эпициклов”.
Пурификасьон Лопес-Гарсия и Давид Морейра справедливо замечают, что загадочное превращение архейных мембран в бактериальные остается настоящей ахиллесовой пятой традиционных симбиотических гипотез, согласно которым цитоплазма эукариотной клетки имеет архейное происхождение. А вот если допустить, что происхождение цитоплазмы — бактериальное, то проблема мембран тут же снимается и существующая картина получает простое объяснение.
Слабые места у этой гипотезы тоже есть — куда ж без них. Прежде всего, она никак не объясняет происхождения митохондрий (которые как-никак тоже потомки грамотрицательных бактерий и заведомо являются источником значительной части генов, полученных эукариотами из этой эволюционной ветви). С учетом гипотезы Гупты выходит, что симбионтов, без которых немыслим эукариотный организм, было не два, а три. Сначала клетка археи слилась с клеткой какой-то неведомой грамотрицательной бактерии, а потом образовавшийся гибридный монстр вобрал в себя еще и предка митохондрии.
Может показаться, что такой сложный сценарий, включающий дополнительных участников, нарушает принцип экономии мышления (согласно которому критерий истины состоит в достижении максимума знаний с помощью минимума познавательных средств). Само по себе это не страшно: механизм такого уникального эволюционного события, как возникновение эукариот, не мог быть простым. Хуже другое. Древнейший бактериальный хозяин симбиотической археи (если он существовал) до сих пор неизвестен. На эту тему были кое-какие предположения, но проверить их оказалось очень трудно. Чтобы защитить гипотезу Гупты, приходится допустить, что древнейший бактериальный предок в процессе интеграции в эукариотную клетку изменился до неузнаваемости — в отличие от митохондрий, которые вступили в симбиоз позже и сохранили свой исходный тип обмена веществ. Это, конечно, возможно, но вот тут позиция сторонников гипотезы Гупты становится довольно слабой. Судя по данным современной геномики, ни одна другая группа бактерий не дала такого серьезного вклада в геном эукариот, как альфа-протеобактерии (хотя мелких заимствований от самых разных групп там полным-полно). Между тем альфа-протеобактерии — это и есть та самая группа, в которую входят предки митохондрий. А вот поиск какого-нибудь еще бактериального предка эукариот зашел в тупик. Например, гены дельта-протеобактерий (которых Лопес-Гарсия и Морейра одно время считали теми самыми “древнейшими бактериальными предками”) в эукариотных геномах встречаются, но в количестве, ничем не выделяющемся на фоне многих других источников генетического материала. Если дельта-протеобактерии когда-то и сыграли некую особую роль, то последующая эволюция стерла генетические свидетельства этого.
Конечно, можно предположить, что альфа-протеобактериальных симбионтов было два: древний (предок цитоплазмы) и поздний (предок митохондрии). Но такая гипотеза явно не соответствует критерию “экономичности”, и, чтобы она была принята, потребуются очень сильные свидетельства, которых пока нет. (Тем не менее кое-что на эту тему обсуждается в главе 10.)
На другие козырные аргументы теории Гупты сторонникам теории Баумов тоже есть что ответить. Во-первых, сейчас выяснилось, что не все бактерии, передавшие эукариотам свои гены, были грамотрицательными. Например, гены, кодирующие ферменты гликолиза, эукариоты получили не от грамотрицательных бактерий, а от грамположительных . Правда, от грамотрицательных бактерий взято больше, но это может объясняться просто преобладанием грамотрицательных бактерий в тех природных сообществах, где эукариоты возникли. Кроме того, в последнее время появились свидетельства, что и обычные археи, до всякого происхождения эукариот, уже довольно активно заимствовали гены от бактерий. Выводы отсюда пока неясны, но в любом случае эти факты дополнительно усложняют картину.
Во-вторых, “правозакрученные” и “левозакрученные” молекулы липидов не столь уж несовместимы в одной мембране: оказывается, можно подобрать условия, при которых смешанная мембрана будет вполне стабильна, и это в принципе позволяет допустить, что именно такая мембрана и была у первых эукариот. Тогда замена компонентов мембран по ходу эволюции могла быть и постепенной.
Наконец, еще одно замечание касается стройной схемы, согласно которой архейные гены (информационные) управляют генетическим аппаратом, а бактериальные (операционные) — обменом веществ в цитоплазме. Из этой схемы есть одно важное исключение: цитоскелет. Белки цитоскелета — типично “операционные”, характерные не для ядра, а для цитоплазмы, но уж они-то абсолютно точно унаследованы эукариотами от архей. Тут налицо, если воспользоваться термином из наук о Земле, несогласие, уже давно озадачивавшее исследователей. Конечно, объяснить его так или иначе при желании нетрудно: гены белков цитоскелета могли сохраниться при любом слиянии клеток просто потому, что к тому моменту они были “ноу-хау” архей, полезным на все случаи жизни и не имевшим никаких бактериальных аналогов. Но во всяком случае это еще раз показывает, с какой сложной проблемой мы имеем дело. Остается дождаться, пока исследователи извлекут из морских бездн или еще откуда-нибудь живых асгардархеот: тогда, скорее всего, что-нибудь да прояснится.
“Обозначились смутные контуры”
А теперь вернемся к системе самих эукариот. В последней трети XX века в этой области науки тоже начались интересные события.
Прежде всего, 1970-е стали эпохой переоценки признаков, по которым эукариоты обычно классифицируются. С одной стороны, окончательно выяснилось, что выделять крупные группы эукариот по способу питания, равно как и по составу клеточных стенок или по типу запасных веществ, — дело безнадежное: эти признаки слишком легко меняются в ходе эволюции. С другой же стороны, новым источником данных о строении клеток стала электронная микроскопия. Она позволяет изучать такие структурные детали, которые ни в один световой микроскоп просто не видно. Например, оказалось, что хлоропласты — части клеток, где происходит фотосинтез, — у одних эукариот окружены двумя мембранами, а у других почему-то тремя или даже четырьмя. Не менее интересной оказалась структура митохондрий. Как известно, любая митохондрия имеет две мембраны — наружную и внутреннюю, причем ключевые биохимические процессы, ради которых митохондрия, собственно, и нужна, связаны с ее внутренней мембраной (см. главу 11). Неудивительно, что поверхность этой мембраны увеличивается за счет складок, которые принято называть кристами. Так вот, электронная микроскопия показала, что кристы митохондрий могут иметь разную форму. Они могут быть пластинчатыми, а могут быть трубчатыми. Есть и другие варианты формы крист (например, дисковидные и мешковидные), но они встречаются реже. У большинства эукариот кристы или пластинчатые, или трубчатые. Самое же интересное, что форма крист, судя по всему, обычно очень медленно меняется в эволюции и остается устойчивой внутри крупных групп эукариот. А значит, она может быть хорошим признаком для различения этих групп. Почему бы не попробовать?
Забегая вперед, надо сказать, что сейчас форму крист митохондрий уже не помещают в основу системы хотя бы потому, что некоторая изменчивость внутри групп по этому признаку все же есть. И тем не менее ряд важных вещей по кристам митохондрий понять можно. Чем больше накапливалось данных на эту тему, тем яснее становилось, что все эукариоты распадаются как минимум на две большие группы, связанные только через общий корень. Причем линия раздела между этими группами пролегла довольно неожиданным образом: из классической зоологии и ботаники подобное никак не следовало.
В 1986 году петербургский зоолог Ярослав Игоревич Старобогатов придал этим представлениям максимально четкую форму. Он отважился разделить всех эукариот на таксоны Lamellicristata (с пластинчатыми кристами) и Tubulicristata (с трубчатыми кристами), которые получили ранг надцарств (см. рис. 14.5). Предполагалось, что это два главных эукариотных эволюционных ствола. В группу Lamellicristata вошли зеленые водоросли, высшие растения, эвгленовые жгутиконосцы, воротничковые жгутиконосцы, многоклеточные животные. В группу Tubulicristata — бурые, золотистые и желтозеленые водоросли, инфузории, фораминиферы, радиолярии. В обеих группах были и одноклеточные, и многоклеточные, и автотрофные, и гетеротрофные представители. Теперь они делились не по этим признакам, а исключительно по предполагаемому родству. Так стали прорисовываться контуры совершенно новой системы организмов, резко отличающейся от геккелевской.
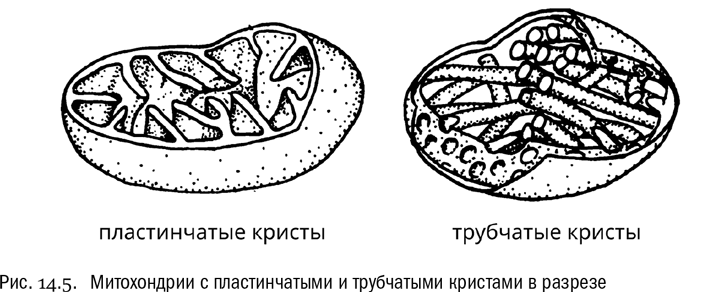
На самом деле в системе Старобогатова было еще и третье надцарство — Akonta (безжгутиковые). В него вошли эукариоты с пластинчатыми кристами митохондрий, на всех стадиях жизненного цикла полностью лишенные жгутиков: красные водоросли и типичные грибы. Это объединение не поддерживается никакими новыми данными, так что обсуждать его нет смысла. Старобогатов был бы гораздо более прав, если бы включил красные водоросли и грибы в группу Lamellicristata. Увы, даже гениальные ученые (а Старобогатов явно был отмечен проблеском гениальности) почти никогда не угадывают правильно все сразу.
В любом случае система Старобогатова (причем именно та ее часть, которая успешно выдержала проверку временем) ведет к одному серьезному выводу, который легко будет понять всякому, кто хоть немного помнит обычный школьный учебник зоологии. Первая часть этого учебника знакомит читателя с такими чудесными созданиями, как обыкновенная амеба, инфузория-туфелька и малярийный плазмодий, иногда дополняя рассказ о них упоминаниями всевозможных сувоек, раковинных амеб, фораминифер и радиолярий. Так вот, все только что названные существа имеют митохондрии с трубчатыми кристами. А это уже само по себе означает, что они относятся к совсем иным эволюционным ветвям, чем многоклеточные животные. Инфузория-туфелька, активно плавающий поедатель бактерий, действительно похожа на животных образом жизни, но эволюционно она к ним ничуть не ближе, чем, например, какое-нибудь зеленое растение. Называть таких существ “одноклеточными животными” попросту неправильно.
Вновь забегая вперед, добавим, что понятие “одноклеточные животные” вообще вышло из употребления, причем уже довольно давно. Поэтому с этого момента мы будем считать, что “многоклеточные животные” и просто “животные” — строгие синонимы. Никаких животных, кроме многоклеточных, просто нет на свете.
Ради справедливости укажем, что есть и такие одноклеточные представители “фауны школьного учебника”, у которых кристы дисковидные (или дисковидные в сочетании с трубчатыми). Это некоторые жгутиконосцы: фотосинтезирующая эвглена, хищный бодо и вызывающая сонную болезнь трипаносома. От многоклеточных животных (у которых кристы, напомним, пластинчатые) эти существа так же далеки, как и инфузории.
В то же время есть как минимум две относительно большие эволюционные ветви эукариот, которые действительно родственны многоклеточным животным. Это всем известные грибы и не столь знаменитые, но зато еще более близкие к животным воротничковые жгутиконосцы. И те и другие имеют пластинчатые кристы митохондрий и гетеротрофный обмен веществ. Надо заметить, что это достаточно “сильное” сочетание признаков, которое встречается лишь у немногих эукариот (по крайней мере, если учитывать крупные ветви, а не количество видов). Однако во всем остальном грибы и воротничковые жгутиконосцы мало похожи друг на друга.
Грибы — это преимущественно наземные организмы, специализирующиеся на осмотрофном питании (см. главу 10). Попросту говоря, “осмотрофы” значит “всасывальщики”. Тело типичного гриба представляет собой систему очень тонких — толщиной в одну клетку — нитей с огромной суммарной поверхностью, сквозь которую всасываются питательные вещества. В просторечии эта система нитей называется грибницей. Нити грибницы одеты хитиносодержащей клеточной стенкой, которая придает им прочность, но затрудняет любое активное изменение формы клеток (см. главу 5). Фагоцитоза у грибов нет. У большинства из них нет и жгутиков, причем ни на каких стадиях жизненного цикла (в отличие от многоклеточных животных, у которых очень часто бывают жгутиковые сперматозоиды).
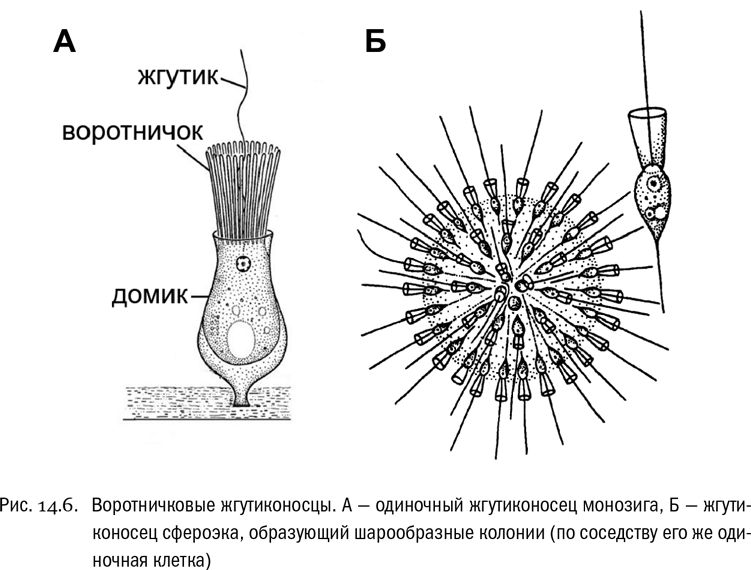
Воротничковые жгутиконосцы — это водные существа, одноклеточные или образующие колонии (см. рис. 14.6). Клетка воротничкового жгутиконосца имеет один жгутик, окруженный характерным “воротничком” — кольцевым выростом, облегчающим ловлю пищевых объектов. Под электронным микроскопом видно, что этот “воротничок” на самом деле состоит из множества длинных тонких выростов клетки, укрепленных проходящими внутри микрофиламентами и как бы склеенных вместе при помощи специальных белков. Эти тонкие выросты называются микроворсинками. Воротничковые жгутиконосцы подвижны. Они могут плавать, а могут и вести прикрепленный образ жизни, но в этом случае они постоянно работают жгутиком, подгоняя к себе воду вместе со взвешенными в ней пищевыми частицами. Питаются они путем фагоцитоза, поглощая эти частицы. По-научному такое питание называют фаготрофным, противопоставляя его осмотрофному.
Воротничковых жгутиконосцев открыли в XIX веке. И тут же было замечено, что клетки, очень похожие на них, входят в состав тел некоторых многоклеточных животных, а именно губок. Это, конечно, не случайность. Современные генетические данные надежно подтверждают сделанный еще полтора века назад вывод: воротничковые жгутиконосцы — самые близкие современные родственники многоклеточных животных. Получается, что это едва ли не единственные “одноклеточные животные”, для которых такое название могло бы иметь хоть какой-то реальный смысл. Но при этом они-то как раз в школьный учебник зоологии и не попали.
Вклад Кавалье-Смита
В 1981 году английский протистолог Томас Кавалье-Смит предложил версию системы эукариот, в которой фигурировали шесть царств.
Царство Fungi. Грибы. Организмы с осмотрофным питанием, пластинчатыми кристами, хитиновой клеточной стенкой, лишенные каких бы то ни было жгутиковых стадий (за исключением самых примитивных форм, выделяемых в особое подцарство).
В остальных пяти царствах жгутиковые клетки сохраняются — если и не у всех представителей, то у многих.
Царство Animalia. Животные. Организмы с фаготрофным питанием, пластинчатыми кристами и активной подвижностью. В это царство входят многоклеточные животные и воротничковые жгутиконосцы.
Царство Plantae. Растения. Фотосинтезирующие организмы с пластинчатыми кристами. Хлоропласты окружены двумя мембранами. Фагоцитоз отсутствует. Одноклеточные, колониальные или многоклеточные формы. В это царство входят красные водоросли, зеленые водоросли, высшие растения.
Царство Chromista. Оно было впервые выделено самим Кавалье-Смитом в этой самой работе. Фотосинтезирующие организмы с трубчатыми кристами (за исключением одной группы одноклеточных). Хлоропласты окружены тремя или четырьмя мембранами. У многих представителей есть фагоцитоз. Одноклеточные, колониальные или многоклеточные формы. В это царство входят бурые, золотистые и желтозеленые водоросли, а также несколько довольно разнообразных групп одноклеточных жгутиконосцев.
Царство Protozoa. Одноклеточные гетеротрофные или (реже) миксотрофные организмы с трубчатыми кристами. Питаются путем фагоцитоза. Передвигаются с помощью жгутиков, ресничек или ложноножек. К этому царству относятся инфузории, амебы, радиолярии, фораминиферы, солнечники, споровики и несколько групп жгутиконосцев. Это и есть “одноклеточные животные” в том (устаревшем) значении, которое раньше встречалось в учебниках.
Царство Euglenozoa. Эвгленовые жгутиконосцы. Одноклеточные формы, плавающие с помощью жгутиков. Некоторые строго гетеротрофны (например, трипаносома и бодо), некоторые способны к фотосинтезу и являются типичными миксотрофами (например, эвглена), а некоторые даже строго автотрофны (например, колациум и трахеломонас). Хлоропласты, если они есть, окружены тремя мембранами. Кристы митохондрий не пластинчатые и не трубчатые, а дисковидные. Относительно немногочисленное царство, состоящее из одних жгутиконосцев, но достаточно сильно обособленное, чтобы его не следовало ни с кем объединять.
Что здесь можно увидеть? Во-первых, как мы сейчас понимаем, эта система не свободна от ошибок (например, некоторых паразитических жгутиконосцев, вошедших в царство “протозоев”, на самом деле правильнее было бы сблизить с “эвгленозоями”). Еще бы — в 1981-м! Скорее стоит удивиться, что серьезных ошибок там не так уж и много. Кроме того, к чести Томаса Кавалье-Смита, надо сказать, что он никогда не отстаивал свои гипотезы любой ценой. Он вполне сознательно рассматривает их как ряд последовательных приближений, описывающих эволюцию все более и более точно, но ни в коем случае не окончательных. Поэтому он всегда легко пересматривал свои предположения, включая в них новые факты, а иногда и сразу формулировал их так, чтобы облегчить последующий пересмотр. Характерно, что в статье 1981 года (а это одна из десятков его работ на подобные темы) Кавалье-Смит на самом-то деле предложил сразу несколько возможных вариантов системы эукариот, с числом царств от пяти до девяти. Например, в отношении красных и зеленых водорослей он тогда колебался — относятся ли они к одному царству или к разным. Поскольку эта книга — все же не труд по истории науки, то во всех подобных случаях тут выбраны версии, наиболее соответствующие тем гипотезам, которые потом подтвердились.
Второе, что бросается в глаза: Кавалье-Смит сохраняет невероятно архаичный таксон Protozoa (предложенный, как мы помним, еще в середине XIX века. Карлом Зибольдом), но при этом радикальным образом меняет его состав и значение. Он исключает из этого таксона всех родственников животных, зато оставляет в нем существ с трубчатыми кристами митохондрий, которые не имеют к животным ни малейшего отношения. В таких случаях для таксонов обычно придумывают новые названия, но Кавалье-Смит, всегда склонный к парадоксам, делать этого не стал.
Кроме того, в той же статье 1981 года Кавалье-Смит высказал два важных предположения:
1 Возможно, царства Fungi и Animalia образуют единую эволюционную ветвь. Во-первых, между ними есть некоторое биохимическое сходство: например, и у грибов, и у животных широко распространен хитин. Во-вторых, самостоятельно плавающие жгутиковые клетки грибов (“низших”, у которых жгутики сохраняются), воротничковых жгутиконосцев и многоклеточных животных имеют один жгутик, направленный назад. У других эукариот такое состояние не встречается.
2 Возможно, единую эволюционную ветвь образуют также царства Chromista и Protozoa. Во всяком случае, только в этих двух группах распространено сочетание трубчатых крист митохондрий со способностью питаться путем фагоцитоза.
3 Рассмотренная система Кавалье-Смита — не слишком обобщенная (как система Уиттэкера, где почти все царства на поверку оказались сборными), но и не слишком дробная (как система Лидейла, где статус царства придается каждой крупной ветви). Самое же главное, что эта система абсолютно последовательно строится по родству, или, во всяком случае, по достаточно хорошо обоснованным гипотезам о родстве. Теперь контуры истинного эволюционного древа эукариот стали наконец видны, пусть пока еще и смутно.
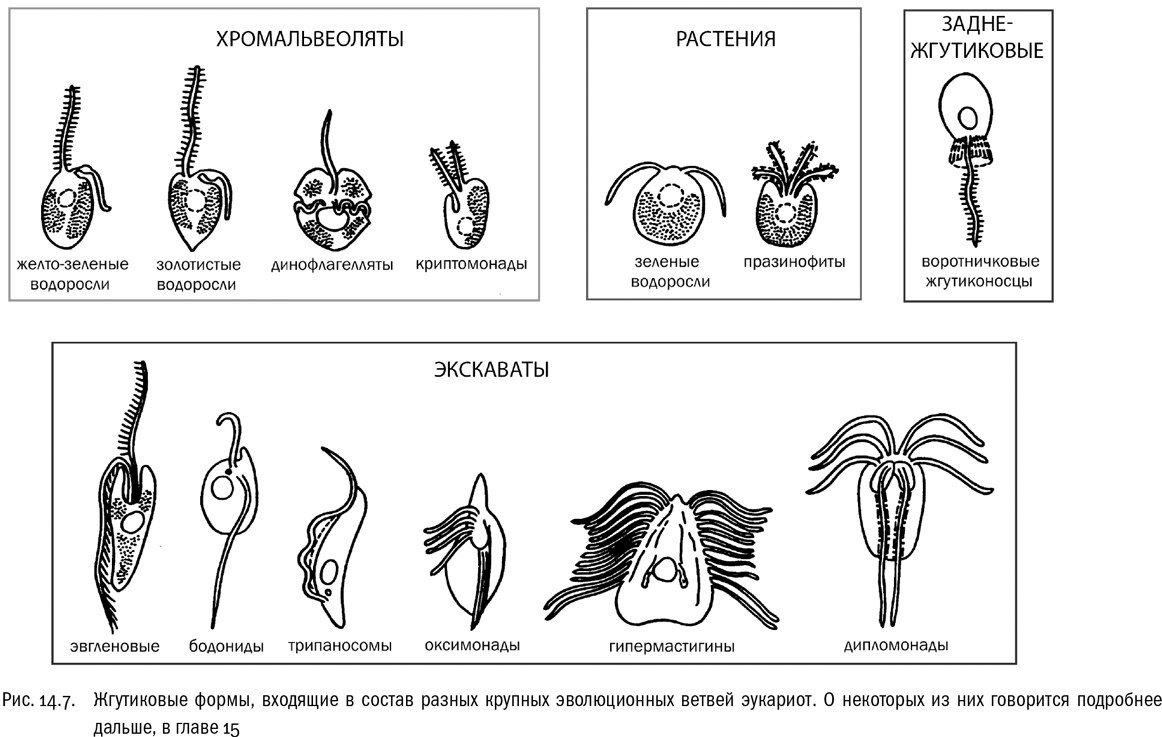
В 1987 году произошло историческое событие: Кавалье-Смит формально выделил эволюционную ветвь, в которую входят воротничковые жгутиконосцы, многоклеточные животные и грибы. Эта группа получила название Opisthokonta (заднежгутиковые). Чтобы понять смысл этого названия, еще раз вспомним, что очень многие эукариотные клетки имеют жгутик — нитевидный вырост, в котором проходит пучок микротрубочек, обычно организованных по схеме 9+2 (девять сдвоенных микротрубочек по краю и две одиночные в центре). Работа жгутика обеспечивает движение клетки в воде. Если жгутиков очень много и они бьются согласованными волнами, то их называют ресничками. Эти структуры уникальны для эукариот. Жгутики бактерий не имеют с эукариотными жгутиками ничего общего, кроме названия. А тех эукариот, у которых жгутик служит основным средством движения, принято называть жгутиконосцами — это слово мы уже не раз встречали (см. рис. 14.7).
Жгутик, вызывающий поступательное движение клетки, можно уподобить винту корабля или самолета, хотя, в отличие от винта (и от бактериального жгутика), он не вращается, а изгибается из стороны в сторону — примерно как рыбий плавник. Тем не менее эукариотный жгутик, как и винт, может быть тянущим (тогда он направлен вперед), а может быть толкающим (тогда он направлен назад). Например, у самолетов бывают винты обоих типов, хотя толкающие встречаются заметно реже. Пример самолета с толкающим кормовым винтом — немецкий истребитель “Хеншель” Hs.P75, разработанный во время Второй мировой войны, но так и не пущенный в производство. (Между прочим, очень похожий на него самолет увековечил японский режиссер Мамору Осии в своем знаменитом полнометражном аниме “Небесные скитальцы”.) Возвращаясь к жгутиковым клеткам, можно сказать, что у них ситуация примерно такая же, как в винтовой авиации: для большинства эукариот характерны тянущие жгутики, а вот толкающие — это скорее нечто необычное. Обладатели “кормовых” толкающих жгутиков — воротничковые жгутиконосцы, жгутиковые стадии примитивных хитридиевых грибов и сперматозоиды многоклеточных животных. Вот эти группы и называются заднежгутиковыми (опистоконтами). Другие общие признаки опистоконтов — пластинчатые кристы митохондрий и строгая гетеротрофность. Кавалье-Смит выделил эту группу абсолютно правильно: ее реальность надежно подтвердили дальнейшие исследования, в первую очередь молекулярно-генетические. Хотя у самого Кавалье-Смита молекулярных данных такого рода в ту пору не было (а если и были, то настолько противоречивые, что это лишь сбивало с толку ). Кроме воротничковых жгутиконосцев, грибов и животных к опистоконтам относится еще несколько небольших групп одноклеточных и колониальных существ, родственных или грибам, или животным.
В 1991 году Кавалье-Смит выделил еще одну группу эукариот, получившую название Alveolata. Это одноклеточные организмы с трубчатыми кристами митохондрий, имеющие дополнительные общие признаки на уровне тонкой структуры клеток. Под наружной мембраной у них находятся многочисленные плоские пузырьки, которые называются альвеолами. Слой альвеол, укрепленный микротрубочками, образует внутреннюю оболочку клетки — пелликулу. К альвеолятам относятся инфузории, некоторые гетеротрофные жгутиконосцы с их паразитическими родственниками (включая малярийного плазмодия) и одна группа одноклеточных водорослей — динофлагелляты. Самое интересное, что эти существа в общем-то совсем не похожи друг на друга ни по внешнему облику, ни по образу жизни. Предполагая, что они образуют одну ветвь, Кавалье-Смит основывался в первую очередь на сходстве внутриклеточных структур, невидимых без электронного микроскопа, и оказался опять прав. (В более старой системе 1981 года эти организмы входили в царство Protozoa, но оно, по мнению подавляющего большинства современных авторов, оказалось все же сборным.)
Между тем в биологии наступила эпоха молекулярной филогенетики. Сама эта наука родилась значительно раньше. Но именно в последнее десятилетие XX века исследования нуклеотидных последовательностей генов начали ставить на поток. Излюбленным объектом молекулярных филогенетиков в тот период были гены, кодирующие рибосомную РНК: она невелика, информативна, и ее изучение уже привело Карла Вёзе к колоссальному успеху. Работы генетиков, сравнивавших последовательности рРНК разных эукариот, сразу подтвердили реальность групп Opisthokonta и Alveolata. А дальше последовал еще более серьезный вывод. Появились указания на то, что группы Chromista и Alveolata образуют единую эволюционную ветвь, многочисленную и разнообразную, но состоящую в основном из организмов с трубчатыми кристами митохондрий. Эта ветвь получила название Chromalveolata.
Хромальвеоляты просто поражают своим разнообразием. Среди них есть одноклеточные, колониальные и многоклеточные фотосинтезирующие существа (всевозможные водоросли), есть плавающие одноклеточные хищники (инфузории, хищные жгутиконосцы), есть крайне специализированные паразиты (споровики, к которым относится малярийный плазмодий), а есть и многоклеточные осмотрофы, точные аналоги грибов по внешнему облику и образу жизни. В общем, одних только хромальвеолят наверняка хватило бы, чтобы обильно заселить эукариотами целую планету, создав на ней сложные и разнообразные экосистемы со своими собственными растениями, фаготрофами, паразитами и поедателями мертвой органики.
Вот кого среди хромальвеолят нет, так это многоклеточных хищников, аналогов животных. При всем их огромном разнообразии такую жизненную форму им “изобрести” не удалось. Судя по всему, живой природе Земли было намного сложнее создать многоклеточных животных, чем многоклеточные растения (мы еще поговорим про это в последней части книги).
Заодно молекулярные исследования подтвердили единство эволюционной ветви Plantae — в том объеме, в каком ее выделил Кавалье-Смит. В эту группу входят все зеленые растения, от одноклеточных водорослей до цветковых, и красные водоросли в придачу.
Тут возникает один интересный чисто эволюционный момент. Общий предок эволюционных стволов Plantae и Chromalveolata мог быть только одноклеточным: это следует из конфигурации древа совершенно однозначно. Значит, входящие в эти два ствола многоклеточные водоросли стали таковыми независимо друг от друга. Если многоклеточные животные возникли в эволюции (насколько мы сейчас знаем) всего однажды, то многоклеточные растения совершенно точно возникали несколько раз. Это стало окончательно ясно еще в первой половине XX века, после работ выдающегося ботаника Адольфа Пашера: он показал, что есть несколько совершенно самостоятельных эволюционных линий водорослей, каждая из которых начинается с одноклеточных организмов и постепенно достигает многоклеточности. Например, многоклеточные зеленые и красные водоросли принадлежат к разным эволюционным линиям, начинающимся от разных одноклеточных предков — и это несмотря на то, что в системе, из которой мы сейчас исходим, обе линии входят в группу Plantae. Можно точно сказать, что многоклеточность растительного типа, пусть и примитивная, возникала не менее трех раз в эволюционной ветви Plantae (у красных, зеленых и харовых водорослей) и по меньшей мере столько же раз в эволюционной ветви Chromalveolata (у бурых, золотистых и желто-зеленых водорослей). А скорее всего, таких событий было еще больше.
Назад: 13. земля и жизнь
Дальше: 15. мир эукариот

