Книга: От атомов к древу. Введение в современную науку о жизни
Назад: ЧАСТЬ III ДРЕВО ЖИЗНИ
Дальше: 14. рождение системы
13. земля и жизнь
В период Чоу твердеющая масса опускается; образуются вода, огонь, горы, камни, земля — пять элементов, которые получили название пяти стихий. Поэтому и говорят, что земля появилась в период Чоу. Через следующие 5400 лет период Чоу кончается, и с наступлением периода Инь на земле зарождается жизнь.У Чэн-энь. Путешествие на запад
Пять миллиардов лет назад ни Солнца, ни Земли еще не существовало. По современным оценкам, самая точная дата начала образования Солнечной системы — 4,5682 миллиарда лет назад (это определено по возрасту древнейших твердых включений в метеоритах). К тому времени Вселенная уже имела длинную и сложную историю. Мы знаем, что Большой взрыв произошел примерно 13,8 миллиарда лет назад. А это означает, что в момент начала формирования Солнечной системы возраст Вселенной уже насчитывал около 9,2 миллиарда лет. Почти бездонная пропасть исторического прошлого, настоящая “бездна времен”.
К моменту рождения Солнечной системы несколько совершенно разных космических эпох успели сменить друг друга. Началу образования звезд предшествовали так называемые “темные века” (Dark Ages), когда во Вселенной не было никаких светящихся объектов. Первые звезды зажглись не раньше чем через 100 миллионов лет после Большого взрыва. И эти звезды очень сильно отличались от Солнца. Наше Солнце — желтый карлик с ожидаемой продолжительностью жизни порядка 10 миллиардов лет. А первые звезды во Вселенной, судя по расчетам астрономов, были голубыми гигантами с массой, которая могла в 500–1000 раз превосходить массу Солнца, и со светимостью в миллионы раз выше солнечной, но со средней продолжительностью жизни всего лишь три миллиона лет. Планетных систем эти звезды не имели: в древнейшей Вселенной, состоявшей целиком из водорода и гелия, им было бы просто не из чего образоваться.
Когда короткий жизненный цикл звезд первого поколения заканчивался, эти звезды или взрывались в качестве сверхновых, или (если их масса более чем в 250 раз превышала массу Солнца) коллапсировали в черные дыры без взрывов. Некоторые из этих массивных черных дыр потом сливались друг с другом и становились центрами ядер формирующихся галактик. А те звезды, которые становились сверхновыми, при взрывах разбрасывали на огромные расстояния газ и пыль — теперь уже насыщенные новыми химическими элементами, гораздо более тяжелыми, чем водород и гелий. Эти тяжелые элементы могли синтезироваться только в недрах звезды (или даже непосредственно во время ее взрыва). И в туманностях, которые оставались от таких звезд, постепенно началось образование звезд следующих поколений — уже с планетными системами.
Около пяти миллиардов лет назад одна из таких туманностей находилась на окраине галактики Млечный Путь. Вероятно, она могла бы до сих пор оставаться в покое, если бы не близкий взрыв сверхновой звезды . По крайней мере, такова популярная у современных астрономов гипотеза, основанная на особенностях изотопного состава некоторых метеоритов. Ударная волна сверхновой, распространившаяся в безвоздушном пространстве благодаря огромному количеству выброшенного при взрыве звездного материала, заставила наше газопылевое облако потерять устойчивость и коллапсировать. Оно расслоилось, в нем появились сгущения, которые притянули к себе окружающее вещество, и в конце концов вся масса облака стала стягиваться к одному центральному ядру. Это и была растущая звезда. Но одновременно потерявшее устойчивость облако начало вращаться, поэтому возникшая центробежная сила растянула его в диск. В конце концов центральное ядро крутящегося газопылевого диска стало Солнцем, а периферия — планетами.
Планетная система делится на две части так называемой “линией льда”, которая в нашем случае проходит между орбитами Марса и Юпитера. Внутри от “линии льда” летучие вещества (вода, аммиак, метан, углекислый газ, угарный газ) в основном испаряются, а снаружи от нее — конденсируются в ледяные пылинки, огромные массы которых могут входить в состав планет. В результате внутри от “линии льда” образовались твердые планеты земного типа (Меркурий, Венера, Земля, Марс), а снаружи — планеты-гиганты, превосходящие планеты земного типа массой в десятки и сотни раз, но гораздо менее плотные (Юпитер, Сатурн, Уран, Нептун).
При этом большая часть воды, находящейся в Солнечной системе, оказалась как раз по внешнюю сторону “линии льда”. Например, ядра комет состоят из водяного льда на 80%, а в составе Урана и Нептуна воды столько, что эти планеты относят к классу ледяных гигантов. Мантия Нептуна, скорее всего, представляет собой океан жидкой воды с примесью аммиака и метана, имеющий непредставимую для нас глубину — около 10 000 километров. Очень вероятно, что и Уран устроен примерно так же. А с другой стороны, гипербазиты, из которых в основном состоит мантия Земли, содержат всего лишь 0,05–0,06% воды. Очевидно, что по сравнению со многими другими объектами Солнечной системы это ничтожно мало.
По счастью, Земля еще и очень активная планета. Ее кора (вместе с верхней мантией) расколота на гигантские литосферные плиты, которые непрерывно движутся. В глубине Земли идут конвективные потоки мантийного вещества, а на ее поверхности есть множество действующих вулканов (причем в некоторые эпохи их было гораздо больше, чем сейчас). А вулканические газы обычно не меньше чем на 75% состоят из водяного пара. Та вода, которая все-таки есть в земной мантии, выходит с этим паром, конденсируется и образует океан. Впрочем, есть вероятность, что значительную часть воды на Землю принесли врезавшиеся в нее льдистые метеориты или кометы. Но, как бы там ни было, без участия вулканов облик нашей планеты был бы совсем другим.
Одной из главных причин высокой тектонической активности Земли, скорее всего, является ее уникальный естественный спутник. Луна всего-навсего в 3,7 раза меньше Земли. Ни у одной другой планеты Солнечной системы нет спутника, настолько огромного относительно нее. У Венеры спутников нет вообще, а спутники Марса имеют ничтожные размеры (27 и 24 километра) и представляют собой небольшие астероиды, захваченные полем тяготения планеты. Правда, в Солнечной системе есть и более крупные спутники, чем Луна, но это спутники планет-гигантов: Ганимед, Титан, Каллисто и Ио. По относительному размеру Луна совершенно необычна (если не учитывать систему Плутон — Харон, которая к планетам не относится). И это очень важно. Поле тяготения Луны, направление которого постоянно меняется из-за ее движения по орбите, создает приливные силы, “накачивающие” дополнительной энергией всевозможные перемещения вещества внутри Земли. Конечно, Луна — не единственная причина тектонической активности Земли, но она значительно усиливает ее, ускоряя в итоге всю эволюцию планеты. В общем, со спутником Земле повезло.
Откуда взялась Луна? Предполагается, что через 50–60 миллионов лет после начала своего формирования молодая Земля столкнулась с другим небесным телом — несуществующей ныне планетой размером примерно с Марс, то есть вдвое меньше Земли . Столкновение выбросило в космос некоторое количество материала земной мантии, который и стал основой формирующейся Луны. Планета, нанесшая удар, развалилась, ее остатки или вошли в состав Луны, или стали метеоритами. Эта гипотеза — ее называют гипотезой великого столкновения, или мегаимпакта (Giant Impact), — порождает много вопросов, но и подтверждений у нее хватает. Например, исследование лунных образцов показывает, что Луна на несколько десятков миллионов лет моложе как Земли, так и других тел Солнечной системы; Земля и Луна почему-то уникально близки друг к другу по изотопному составу (их называют “геохимическими близнецами”); и наконец, Луна близка по типу пород к земной мантии, но, в отличие от Земли, почти лишена железного ядра. Гипотеза мегаимпакта изящно объясняет все эти факты.
Так или иначе Земля — единственная землеподобная планета, которая богата одновременно тектонической энергией и жидкой водой. Причем ей посчастливилось сохранить эти свойства на протяжении нескольких миллиардов лет — в отличие от Марса, где тоже когда-то был водный океан. А ведь именно эти две особенности и есть главные предпосылки возникновения жизни (во всяком случае, жизни земного типа): жидкая вода — потому что она служит растворителем для биохимических реакций, а тектоническая энергия — потому что она может эти реакции запустить. Тектоническая активность Земли не только дает тепло, но и обеспечивает перепады концентраций молекул или ионов (начиная с простейшего иона H+), которые сами по себе становятся источниками энергии.
Возникновение жизни
Есть несколько гипотез, более-менее детально расписывающих вероятные первые химические шаги на пути к жизни . Они отличаются в деталях, но едины в главном. Все эти гипотезы предполагают, что местами зарождения жизни были не открытые водоемы, а микрополости в грунте или минеральных осадках, куда подводилась энергия от горячих источников или от вулканов. Надо сказать, что это не такая уж новость. Например, известный швейцарский биолог Карл фон Нэгели еще в XIX веке писал по поводу зарождения жизни: “Вероятно, это случилось не в открытой воде, а во влажном слое тонкого пористого материала (песка, глины), где совместно действовали молекулярные силы твердых, жидких и газообразных тел”. Вот это мнение сейчас и стало научным мейнстримом. Где возникновение жизни наименее вероятно — так это в водной толще спокойного океана, освещенного солнцем. Там просто нет таких потоков энергии и вещества, которые зарождающаяся жизнь могла бы “оседлать” и перенаправить себе на пользу.
Итак, где-то в воде, пропитывавшей окрестности древних вулканов или горячих источников, начались автокаталитические (то есть самоускоряющиеся) химические реакции, цепочки которых вскоре стали пересекаться за счет общих промежуточных продуктов и замыкаться в циклы. Главные участники этих реакций, скорее всего, были небольшими органическими молекулами, поначалу даже одноуглеродными. Но реакции-то были не простыми. Особенность любой автокаталитической реакции по определению состоит в том, что ее продукт одновременно является катализатором, то есть веществом, ускоряющим ход самой реакции. При условии достаточной сложности реакционной системы (а оно в данном случае наверняка соблюдалось: и реагентов, и продуктов было множество) автокаталитические реакции приобретают свойство саморазвития, потому что в них появляется обратная связь: небольшое изменение механизма реакции влияет на состав ее продуктов, изменение которого, в свою очередь, влияет на механизм — и так шаг за шагом. Спустя какое-то время в системе автокаталитических реакций начали синтезироваться аминокислоты, простейшие углеводы, а там дело дошло и до полимеров — сперва простых, потом посложнее. Наконец, некоторые из этих полимеров “научились” катализировать сначала синтез друг друга (это совсем легко), а потом и воспроизводство самих себя. Иными словами, они стали репликаторами. А с появлением репликаторов автоматически включается дарвиновский механизм естественного отбора, необходимые и достаточные условия для которого — самовоспроизводство, наследственность, изменчивость и конкуренция за субстрат. Все, с этого момента биологическая эволюция запущена.
Можно не сомневаться, что на этих первых этапах жизнь была еще практически незаметной для постороннего наблюдателя (если бы, конечно, он мог тогда существовать). Это легко понять, если вообразить себя инопланетным путешественником, прибывшим пусть даже к самой колыбели земной жизни. Что он увидит? Теплый вулканический грунт, башни пористых осадков на морском дне... И все. Ничего примечательного. Без химического анализа такой путешественник и не понял бы, с чем столкнулся.
Первыми в истории Земли полноценными репликаторами, скорее всего, были молекулы РНК (см. рис. 13.1). Дело в том, что из всех биологически активных молекул только РНК может выполнять сразу все жизненно важные функции: и хранение наследственной информации, и ее копирование, и катализ реакций обмена веществ. Белки и их предшественники, более простые пептиды, никогда таких возможностей не имели. Тем не менее первые пептиды наверняка появились примерно в те же времена, что и первые РНК. Это следует из чисто химических соображений. Дело в том, что синтез РНК довольно сложен, а вот аминокислоты — причем именно альфа-аминокислоты, из которых пептиды обычно состоят, — достаточно легко синтезируются из самых простых молекул, например из угарного газа (CO) и циановодорода (HC≡N), в условиях, примерно соответствующих вероятным условиям в окрестностях древних вулканов. Поэтому существование эволюционного этапа, когда автокаталитические системы состояли бы исключительно из РНК, маловероятно. Скорее всего, эволюция пептидов и РНК была сопряженной всегда, еще со времен их гораздо более простых общих предшественников. Возможно, что дополнительной (в придачу к самокопированию) задачей первых репликаторов как раз и был катализ синтеза пептидов, влиявших на химическую среду таким образом, чтобы эти репликаторы с большей вероятностью могли выжить.
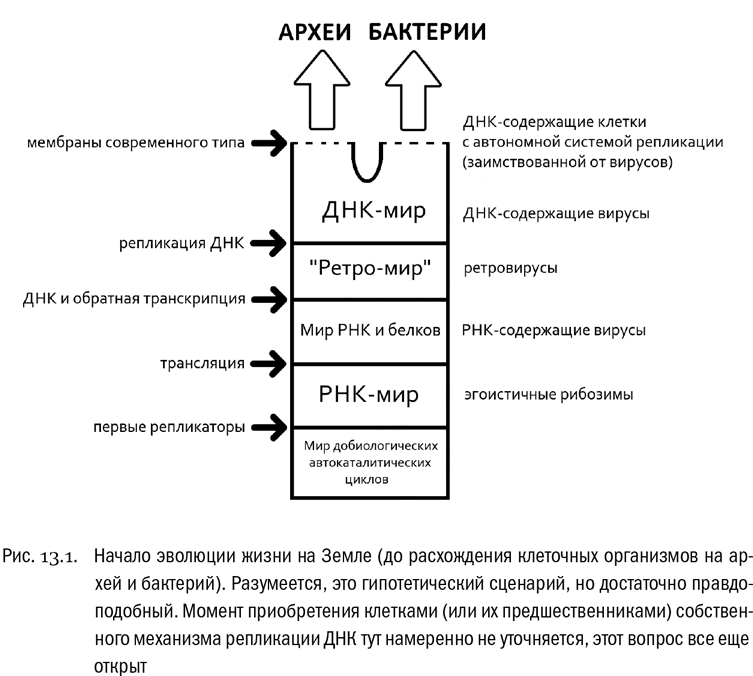
С другой стороны, из современного опыта мы знаем, что белки — более мощные катализаторы, чем РНК, и их возможности в этом плане несравненно разнообразнее. Поэтому неудивительно, что те РНК, которые “научились” катализировать синтез каких-нибудь особых пептидов, получили преимущество в выживании. В результате пептиды (или уже белки?) стали использоваться репликаторами в качестве своего рода молекулярных инструментов, которыми можно было действовать на среду, повышая свои шансы уцелеть и размножиться. Конкурируя друг с другом, РНКовые репликаторы постепенно совершенствовали способность программировать синтез белков, делая это все более и более точно. И в конце концов они “изобрели” механизм трансляции на рибосоме. Этот механизм позволяет запрограммировать всю структуру белка с абсолютной точностью — до каждой аминокислоты. И вот с этого момента возможности живой природы по созданию белков стали буквально безграничными. Заодно появились и первые вирусы — “оппортунистические” репликаторы, которые не стали заводить собственную систему синтеза белка, зато научились паразитировать на чужой.
Следующим важным эволюционным событием был перенос генетической информации с РНК на ДНК. Дело в том, что молекула РНК всем хороша, но вот химическая устойчивость у нее низкая и разрушается она довольно легко. Поэтому длительно хранить на ней генетическую информацию — дело ненадежное. Для этого предпочтителен какой-нибудь другой полимер. Им-то и стала ДНК. Если первые РНК вполне могли синтезироваться спонтанно в неживой природе, то синтез ДНК уже со всей определенностью является “изобретением” живых организмов, и эта молекула с самого начала получила единственную функцию: хранить информацию. Ничего другого она делать не умеет. Одно-единственное преимущество, которое имеет ДНК перед РНК, — ее высокая химическая устойчивость, позволяющая долго и надежно храниться. Для того, кто владеет уникальным “ноу-хау” синтеза каких-нибудь полезных белков, это по-настоящему ценно.
Таким образом, началась эпоха великой перезаписи геномов с РНК на ДНК. В начале этой эпохи на Земле жили РНК-содержащие организмы, которые наверняка уже освоили к тому моменту технологию точного синтеза белка. Иными словами, ДНК появилась эволюционно позже, чем трансляция. Вполне возможно, что генетическая стратегия первых ДНК-содержащих организмов была похожа на генетическую стратегию ретровирусов (см. главу 12). В жизненном цикле вирусов этого типа есть обязательная стадия ретротранскрипции, то есть обратной транскрипции — переноса генетической информации с РНК на ДНК. А вот собственного механизма репликации ДНК у ретровирусов нет. И у клеточных организмов его тоже, скорее всего, вначале не было. Надежные ферменты репликации (они называются ДНК-зависимые ДНК-полимеразы) появились позже. Но уж когда они появились, это дало возможность хранить на ДНК генетическую информацию непрерывно, при необходимости сразу перезаписывая ее с одной молекулы ДНК на другую. И тогда ретротранскрипция стала не нужна.
В результате образовалась самая привычная нам форма жизни: ДНК-содержащая клетка с генетической стратегией “ДНК-РНК-белок”.
Признаемся честно: мы не знаем, когда именно живое вещество разбилось на клетки, отделенные от внешней среды и друг от друга замкнутыми липидными мембранами. Вполне возможно, что это произошло раньше, чем появилась репликация ДНК и исчезла обязательная ретротранскрипция. В таком случае вполне может оказаться, что первые клетки по жизненному циклу напоминали ретровирусы (или даже классические РНК-содержащие вирусы, хотя это менее вероятно). Клеточная мембрана делит весь мир на внутреннюю среду, где химические реакции жестко контролируются геномом, и окружающую среду, где контроль гораздо менее жесткий, требует специальных инструментов (например, выделения из клетки каких-нибудь белков) и где его приходится делить с обладателями других геномов, конкурируя с ними за влияние. Кроме того, клеточная мембрана придает геному целостность, резко ограничивая обмен генетическими элементами с окружающей средой и защищая генетическую систему от генетических паразитов. Только с этого момента приобретает реальный смысл понятие особи, индивидуума, — слово, которое по-латыни значит “неделимый”.
Парадоксальным образом размножаются все живые клетки именно делением. Материнская клетка делится на две дочерние, которые получают достаточно точные копии ее генома. Цепь последовательно делящихся клеток — это цепь прямых, без всяких метафор, предков и потомков. Иногда потомки одной и той же клетки оказываются в разных условиях (или получают разные мутации) и начинают под действием естественного отбора накапливать различия. Тогда мы можем заметить, что линия предков и потомков ветвится.
Первым таким ветвлением было разделение всех клеточных организмов на архей и бактерий. Оно произошло точно раньше, чем появился полноценный механизм репликации ДНК, и наверняка раньше, чем появились клеточные мембраны современного типа. А это означает, что типичные (с нашей точки зрения) клетки, окруженные липидной мембраной и имеющие генетическую стратегию “ДНК-РНК-белок”, с самого начала существовали в виде двух расходящихся эволюционных ветвей.
Так возникло древо жизни.
Arbor vitae
Из всех естественных наук только биология может представить свои объекты в виде эволюционного древа. Связано это с тем, что у живых организмов (и только у них) есть передаваемый по наследству геном, который обеспечивает уникальную преемственность информации и структуры. Белок, возникший миллиард лет назад, вполне может до сих пор сохранить не только свою функцию, но и большую часть аминокислотной последовательности — конечно, если этот белок достаточно важен, чтобы его устойчивость поддерживалась естественным отбором. Предок человека, живший миллиард лет назад, скорее всего, был одноклеточным существом. И тем не менее сохранившаяся преемственность (причем не только чисто генетическая, но и запечатленная в наблюдаемой под микроскопом структуре клеток) не дает забыть о нашем прямом родстве.
Древо жизни — это не воображаемая конструкция или модель, а совершенно реальный объект. Любая его веточка так же реальна, как любая веточка любого дерева, разветвляющегося в пространстве. (Эту фразу написал в одной статье замечательный зоолог и биогеограф Иван Иванович Пузанов, но она выражает настолько очевидную мысль, что ее и в кавычки заключать не хочется: все равно как если бы Пузанов сказал, что Волга впадает в Каспийское море.) Эволюционное древо ничем качественно не отличается от родословной какой-нибудь королевской семьи или линии охотничьих собак. Точнее, различия-то между ними есть (например, очень разным может быть вклад процесса гибридизации), но все они одинаково реальны. Кстати, это одна из причин, почему англичане в середине XIX века довольно легко поняли теорию Дарвина. Англия традиционно была животноводческой страной, ее жители превосходно знали, как составляются родословные породистых коров, собак, голубей и тому подобных одомашненных тварей. Перейти от родословных к эволюционным древесам было нетрудно.
Вот всего одна наглядная иллюстрация. В 1938 году южноафриканский ихтиолог Джеймс Смит сделал поразительное открытие. Он обнаружил, что “кистеперые рыбы вымерли не совсем”. Действительно, кистеперые рыбы — сборная группа, в которую входит открытая Смитом латимерия, — близки к эволюционным корням наземных позвоночных, но до 1938 года они были известны только из палеонтологической летописи. Считалось, что они вымерли даже раньше, чем динозавры. Поэтому современная кистеперая рыба была потрясающей находкой. Но проблема состояла в том, что латимерия была одна-единственная, и поймать второй экземпляр никак не удавалось. Поиски мест, где живет латимерия, заняли много лет и шли очень трудно (эта захватывающая история прекрасно изложена самим Смитом в книжке “Старина-четвероног”). В какой-то момент Смит обратился за помощью к премьер-министру Южной Африки — известному политику Даниэлю Франсуа Малану, объяснив ему по телефону, почему это так важно. И Малан помог. Он выделил самолет и решил все проблемы с зарубежными визами, которые были нужны для срочного полета за пойманной рыбой. Когда рыбу доставили в Кейптаун, Малан спросил Смита: “Вы хотите сказать, что и мы некогда выглядели таким образом?” Смит несколько растерялся и отделался шуткой. Спустя некоторое время известный английский биолог Джулиан Хаксли сказал по этому поводу, что правильным ответом было бы: “Вообще говоря, да!” И он был абсолютно прав. Самым верным ответом тут было бы простое “ДА”. Правда, латимерия в действительности не предок наземных позвоночных, она, скорее, их эволюционная “кузина”. Но и самые что ни на есть прямые предки, судя по всему, выглядели достаточно похоже на нее — неспециалист не отличил бы.
Древо жизни — это совокупность целых организмов (а не только их геномов). Графически оно в самом деле похоже на дерево, ветви которого, правда, могут иногда и сливаться. Пример такого слияния — объединение организмов древней археи и древней альфа-протеобактерии, которое дало начало эволюционной ветви эукариот (см. главу 10). В этом месте древо жизни замкнулось в кольцо. Но полное слияние эволюционных ветвей происходит все-таки редко (гораздо реже, чем обмен отдельными генами). Основной способ биологической эволюции — дивергенция, то есть расхождение ветвей. Этот вывод в целом сохраняет силу со времен Чарльза Дарвина, который описал вероятную общую структуру эволюционных древес, и Эрнста Геккеля, который первым всерьез попытался построить истинное эволюционное древо всего живого.
Конечно, эволюционные деревья, помещаемые в научных работах, бывают несовершенными — точно так же, как несовершенны старые карты мира. На доколумбовых картах нет Америки, а на картах XVIII века еще начисто отсутствует Антарктида. Но уж тут-то ясно, что отсутствие этих континентов на картах не имеет никакой фундаментальной причины: их просто еще не успели открыть. Вот так же обстоит дело и с неточностями эволюционных древес, которые рисуют ученые (и заодно со спорами о том, какая версия древа более правильна, — а они часто бывают очень бурными).
Оглянувшись на эволюцию Вселенной от Большого взрыва до возникновения на Земле животных и растений, мы сразу увидим, что генетическое родство — это совершенно особый тип отношений, по самой своей сути возможный только между живыми объектами. Здесь есть одно исключение — языки, для которых лингвисты тоже устанавливают самое настоящее родство. Родословные деревья языков очень похожи на родословные деревья живых организмов (они и строятся близкими способами, с применением сходных математических аппаратов). Но это не столь уж удивительно, поскольку любой человеческий язык — в конечном счете явление живой природы, часть “расширенного фенотипа” вида Homo sapiens. Так что исключение на самом деле подтверждает правило. Нигде в неживой природе нет ничего подобного. Бесполезно сравнивать по признаку родства две молекулы, два кристалла, две звезды или два вулкана (если только не понимать “родство” совсем уж метафорически). Зато для любой пары живых существ, сколь угодно далеких друг от друга, определить родственные отношения хотя бы в первом приближении нетрудно. Неудивительно, что именно на родстве в первую очередь и основана биологическая система.
Назад: ЧАСТЬ III ДРЕВО ЖИЗНИ
Дальше: 14. рождение системы

