Книга: От атомов к древу. Введение в современную науку о жизни
Назад: 14. рождение системы
Дальше: ЧАСТЬ IV ИСТОРИЯ ЖИЗНИ
15. мир эукариот
Дерево Сфера царствует здесь над другими.Дерево Сфера — это значок беспредельного дерева,Это итог числовых операций.Ум, не ищи ты его посредине деревьев:Он посредине, и сбоку, и здесь, и повсюду.Николай Заболоцкий. Деревья
Область биологической систематики, оперирующая категориями выше типа, называется мегасистематикой. Нечего и говорить, что область эта — исключительно важная. Именно благодаря мегасистематике мы можем представить себе, как в целом, в самом первом приближении устроена живая природа. Крупнейшим мегасистематиком конца XX — начала XXI века можно с полным основанием назвать Томаса Кавалье-Смита. Он сделал в этой области много важных открытий, хотя и ошибок совершил не меньше (но никогда не стеснялся их исправлять, сразу используя для этого появляющиеся новые данные). В любом случае надо учитывать, что современные эволюционные исследования — это всегда коллективный процесс. Над такими проблемами, как построение эволюционного древа эукариот, в наше время работают большие международные группы ученых. Вот общую картину родственных отношений, которую им удалось раскрыть, мы сейчас и рассмотрим. Благо картина эта уже достаточно целостная.
Современная мегасистема эукариот сложилась в первые годы XXI века в основном в результате молекулярно-филогенетических исследований . У этой современной мегасистемы есть три важные особенности.
Во-первых, она выстроена строго по принципам филогенетической систематики — без всяких компромиссов. Это означает, что любой таксон там охватывает свою эволюционную ветвь целиком, вместе со всеми потомками, как бы специализированы они ни были. Сборные группы, выделяемые по образу жизни или по уровню организации, теперь полностью исключены. Это радикально отличает новую систему от большинства систем “классического” толка (в том числе и от тех, что создал и продолжает создавать Кавалье-Смит: он-то никогда не был последовательным кладистом).
Во-вторых, эту систему уже невозможно связать с именем какого-либо одного ученого. “У нее нет единого автора: родство между организмами — это факт, не допускающий авторских трактовок. Ученые, создавшие эту систему, — это не философы, а практики, изучающие структуру отдельных генов и анализирующие полученные данные математизированными методами молекулярной кладистики”.
В-третьих, современная систематика мало интересуется рангами таксонов. Иногда их по старинке еще используют, но в целом сейчас никого всерьез не волнует, считать ли такую-то эволюционную ветвь царством, подцарством, надцарством или субдоменом. Очень часто авторы статей по систематике обходятся вовсе без этих рангов. Мы уже знаем, что в филогенетической систематике это нормально: ранги в ней не обязательны, они сохраняются только там, где это удобно. А в данном случае это скорее неудобно (например, употребление хорошо всем знакомого слова “царство” сразу приведет к тому, что его новое значение начнут сравнивать со старым и это вызовет совершенно ненужную путаницу). Поэтому здесь и далее мы будем для удобства считать, что у таксонов ниже домена, но выше типа рангов нет.
Новая система — действительно совершенно новая. Она не имеет никаких корней в старых двух-, трех-, четырех- или пятицарственных системах и не является преемственной по отношению к ним. Для простоты можно считать, что ее построили вообще с нуля (на самом деле это, разумеется, не так, но понять ее так будет легче).
Новая система, конечно, развивается. Более того, она непрестанно модифицируется по принципу последовательных приближений. “Если раньше ученый долгие годы размышлял над деревом, создаваемым им из опыта, интуиции, вдохновения, то сейчас весомый отчет по гранту ставится выше научной щепетильности, и научное сообщество не осуждает практики выдвижения одним и тем же автором раз за разом противоположных филогенетических гипотез при условии, что для них привлекаются новые данные”. Неудивительно, что филогенетическое древо постоянно обновляется и в разных научных работах можно увидеть его слегка отличающиеся версии. Обновления происходят за счет включения в анализ новых генов, или изучения ранее мало изученных живых существ, или применения более совершенных методов обработки данных. Никаких переворотов, подобных тем, что происходили в конце XX века, это не вызывает (и, скорее всего, уже никогда не вызовет), но поток уточнений, которые часто сопровождаются еще и придумыванием новых названий, вполне может сбить новичка с толку. Лучший выход в этой ситуации — выбрать одну хорошую систему, которая в главных своих чертах будет верна, и уж в ней-то разобраться основательно, а потом, где нужно, модифицировать ее с учетом новых открытий.
В качестве такой “хорошей системы” мы рассмотрим эволюционное древо, которое опубликовали в 2004 году биологи Аластер Симпсон и Эндрю Роджер. Название их статьи — “Реальные “царства” эукариот” (именно так, со словом “царства” в кавычках). Чаще всего эти “царства” называют просто супергруппами. Примем во внимание, что в этой системе кое-что успело заведомо устареть: например, одна из супергрупп Симпсона и Роджера на самом деле, скорее всего, является сборной, и подавляющее большинство современных систематиков ее уже не признает. Так что наш обзор этой системы будет относиться скорее к истории науки (пусть и совсем недавней), чем к ее нынешнему состоянию. Это сделано сознательно, потому что такой порядок изложения представляется здесь более удобным. До современного состояния мегасистематики мы еще доберемся.
Итак, Симпсон и Роджер в своем обзоре 2004 года выделили шесть супергрупп эукариот: Excavata, Rhizaria, Chromalveolata, Plantae, Amoebozoa и Opisthokonta (см. рис. 15.1). Поговорим о них по порядку.
Excavata
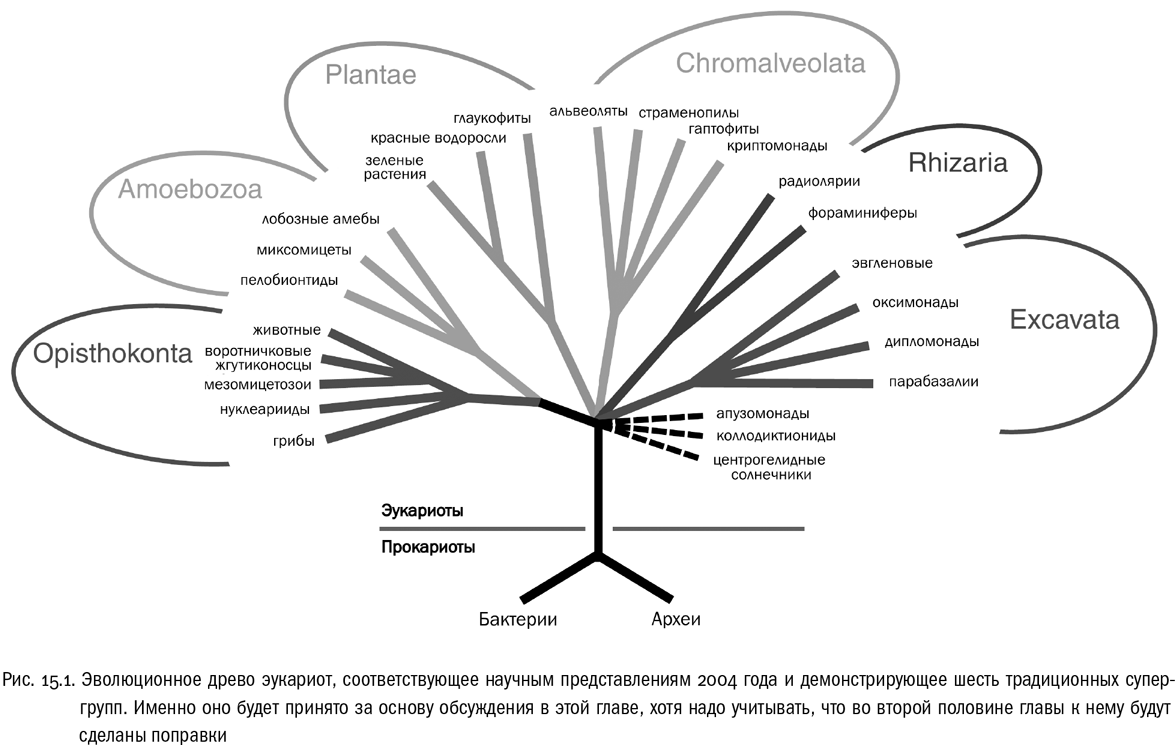
Эта супергруппа состоит в основном из жгутиконосцев — одноклеточных эукариот, у которых главным средством движения служат жгутики (см. рис. 15.2).
Название экскават связано с одной их структурной особенностью: на брюшной стороне клетки (то есть на той, которая обычно обращена к поверхности грунта) находится продольное углубление, со стороны цитоплазмы армированное микротрубочками. Это брюшная ротовая бороздка, через которую жгутиконосец заглатывает пищу (она продолжается в так называемый клеточный рот). Брюшная бороздка есть у многих экскават, но не у всех.
Широко известный представитель экскават — эвглена, попавшая во многие школьные учебники зоологии в роли типичного жгутиконосца. Жгутиков у нее два, но один из них очень короткий и в движении не участвует. Плазматическая мембрана у эвглены подостлана изнутри спиральным каркасом из лент специальных опорных белков, придающим клетке довольно устойчивую форму (в образовании этого каркаса участвуют микротрубочки и даже элементы эндоплазматической сети). Получается упругая оболочка, благодаря которой тело эвглены очень обтекаемо и она хорошо плавает. Но одновременно оболочка затрудняет выпускание ложноножек — и, соответственно, захват пищевых объектов. Поэтому фагоцитоз у эвглены не развит.
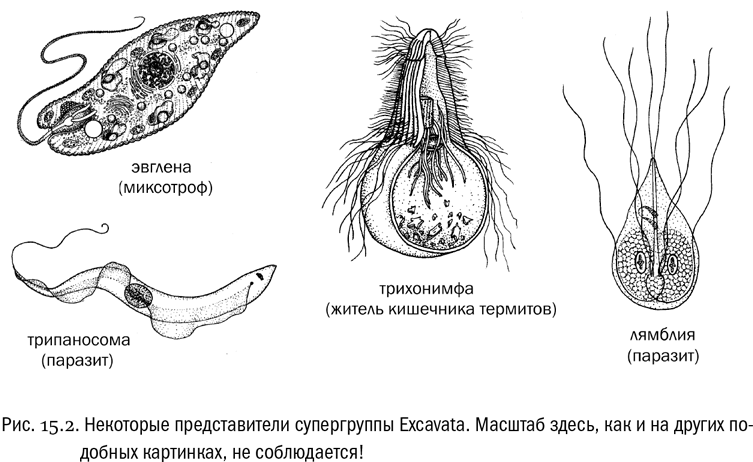
Эвглена — миксотрофный организм. Она прекрасно умеет питаться гетеротрофно, всасывая полезные вещества из раствора (когда эвглен разводят на корм малькам аквариумных рыб, их самих кормят, добавляя в воду мясной бульон). Но она может и фотосинтезировать, потому что у нее есть зеленые хлоропласты. Причем эти хлоропласты произошли от захваченных предком эвглены зеленых водорослей — одноклеточных эукариот из супергруппы Plantae. В свое время эвглену иногда ошибочно сближали с зелеными водорослями, на которые она похожа аппаратом фотосинтеза (и больше ничем). На самом же деле зеленые водоросли родственны не эвглене, а ее хлоропластам.
Близкий родственник эвглены — астазия, жгутиконосец, внешне очень похожий на нее, но бесцветный и питающийся только гетеротрофно. Самое интересное, что у астазии это состояние, скорее всего, вторично: в ее геноме обнаружены гены хлоропластного происхождения. Судя по всему, захват одноклеточной зеленой водоросли, которая стала предком хлоропластов, произошел один раз — у общего предка эвглены и ее родственников. Но с тех пор многие из этих родственников успели потерять хлоропласты и вновь стать гетеротрофными. Потеря хлоропластов произошла не меньше чем в пяти разных ветвях эвгленовых жгутиконосцев, в том числе и у астазии. Благо это несложно: даже у самой эвглены бывают бесхлоропластные популяции, нормально живущие на гетеротрофном питании. Такой вот здесь получился эволюционный зигзаг.
Кроме эвгленовых жгутиконосцев (к которым относится и астазия) в супергруппу экскават входит много других интересных существ. Фотосинтезирующих форм среди них мало, эвглена тут скорее исключение, чем правило. Большинство экскават никогда не имело хлоропластов. Зато среди них есть много гетеротрофных жгутиконосцев, как хищных, так и паразитов. К последним относятся некоторые возбудители инфекционных болезней человека: трипаносомы, лямблии, трихомонады, лейшмании. Все это — паразитические жгутиконосцы из супергруппы Excavata.
Паразитические экскаваты часто имеют причудливую форму клеток. Скорее всего, это связано с тем, что им не нужна особенно высокая маневренность, зато нужен мощный жгутиковый аппарат, чтобы пробиваться сквозь вязкую среду (кровь или содержимое кишечника). Поэтому у них часто увеличивается число жгутиков. Например, лямблия имеет два ядра и восемь жгутиков, ее клетка как бы частично раздвоена. Это далеко не предел, у других жгутиконосцев бывают клетки и посложнее. В то же время непаразитические родственники лямблии живут в толще донных осадков и чаще всего имеют только два жгутика. Такое состояние свойственно большинству хищных жгутиконосцев, которым высокая маневренность как раз нужна. Один жгутик они используют как винт для поступательного движения, другой — как руль.
Некоторые жгутиконосцы-экскаваты, живущие внутри других организмов, приносят им пользу. Например, жгутиконосцы, которые живут в пищеварительном тракте термитов. В отличие от подавляющего большинства животных, термиты могут питаться одной только древесиной. А древесина — это масса растительных клеточных стенок, которые состоят в основном из целлюлозы (см. главу 6). Такой корм очень малопитателен, тем более что у термитов — во всяком случае, у примитивных — нет никакого собственного фермента, способного расщеплять связи между остатками глюкозы в огромной молекуле целлюлозы. Переваривать целлюлозу этим термитам помогают симбиотические жгутиконосцы, живущие у них в кишечнике. Вот эти жгутиконосцы и относятся к супергруппе экскават. Причем жгутиконосцам, в свою очередь, помогают в расщеплении целлюлозы симбиотические бактерии, которыми клетка такого жгутиконосца прямо-таки набита. Так что симбиоз тут “многоэтажный”, по типу русской матрешки.
Типичные обитатели кишечника термитов — крупные жгутиконосцы, называемые гипермастигинами (буквально это значит что-то вроде “сверхжгутиковые”). Такой жгутиконосец может иметь несколько сотен, а то и несколько тысяч жгутиков, расположенных рядами, кругами или спиралями. Ядро у него при этом вполне может быть одно, хотя у некоторых родов много ядер. Неудивительно, что архитектура цитоскелета в таких клетках очень сложная и красивая. Кроме того, у некоторых гипермастигин на поверхности клетки живут еще и спирохеты — подвижные грамположительные бактерии, длинные, тонкие и закрученные в спираль. Они заякориваются на клетке жгутиконосца и несут функцию дополнительных жгутиков. Раньше была популярна гипотеза, что жгутики, собственно, и произошли от таких симбиотических спирохет, но молекулярная биология ее не подтверждает.
Стоит отметить, что жгутиконосцы-гипермастигины иногда живут в кишечнике не только у термитов, но и у тараканов, среди которых тоже есть виды, питающиеся древесиной. Это неудивительно: термиты и тараканы — ближайшие родственники.
Еще один великолепный представитель экскават — Mixotricha paradoxa, живущая в кишечнике одного австралийского термита. Это крупный — до полумиллиметра — одноядерный жгутиконосец, несущий на своем переднем конце не десятки и сотни жгутиков (как у гипермастигин), а всего лишь четыре жгутика, и те малоподвижные. Зато все остальное тело миксотрихи усеяно тысячами симбиотических спирохет, которые бьются вместо ресничек. Причем похоже, что этих спирохет там несколько видов. По подсчетам, на одной миксотрихе может жить примерно 250 000 спирохет. Если убить их антибиотиками, миксотриха тут же теряет подвижность. При этом кроме спирохет на ее поверхности живут еще и другие бактерии, имеющие форму коротких палочек. Эти бактерии сидят на теле миксотрихи там, где прикрепляются спирохеты, как бы заякоривая их. Но и это не все. Помимо “якорных” бактерий и обычных спирохет, работающих как реснички, на теле миксотрихи сидят еще особые крупные спирохеты, относящиеся к другому роду, но тоже участвующие в движении, и какие-то длинные палочковидные бактерии, прикрепляющиеся к ней одним концом. В общем, поверхность миксотрихи подобна густому саду, где процветает никак не меньше четырех совершенно различных разновидностей бактерий-симбионтов. А ведь у нее есть еще и внутренние симбионты, помогающие переваривать целлюлозу. Так что клетка миксотрихи — это целое многовидовое сообщество.
Термиты, как и другие насекомые, могут очень хорошо сохраняться в ископаемом янтаре (если повезет, конечно).
В янтаре возрастом 20 миллионов лет палеонтологи нашли термита того самого рода, в котором живет миксотриха. И в кишечнике этого термита обнаружены остатки симбиотических одноклеточных, уже тогда связанных со спирохетами. Самое крупное из этих одноклеточных, по всей вероятности, и есть миксотриха.
Для экскават характерны дисковидные кристы митохондрий. Впрочем, многие паразитические экскаваты (и некоторые свободноживущие тоже) митохондрий вообще лишены. Например, ни у лямблии, ни у гипермастигин, ни у миксотрихи митохондрий нет. Но это не признак примитивности, а результат вторичной утраты. Потеря митохондрий — обычное дело у паразитов, живущих в чьем-нибудь кишечнике, или, скажем, у обитателей бескислородного ила. Там, где нет кислорода, митохондрии бесполезны, и они исчезают. Но в таких клетках обычно есть их остатки, сохранившие некоторые метаболические функции, или хотя бы митохондриальные гены, успевшие мигрировать в ядро.
В прошлом митохондрии были абсолютно у всех изученных эукариот. И это, конечно, очень важно для понимания их происхождения. “Эмпирический факт заключается в том, что не существует эукариота, возникшего без предшествующего эндосимбиоза”, — писал в 2002 году крупнейший микробиолог Георгий Александрович Заварзин. И добавлял: “Вероятно, проэукариот, так до сих пор и не найденный, уже был химерой”. Эти выводы совершенно верны, и у них есть эволюционные следствия, о которых мы еще поговорим.
Rhizaria
Типичные представители этой супергруппы — одноклеточные существа с трубчатыми кристами митохондрий и длинными тонкими ложноножками, которые часто бывают нитевидными или шиловидными (см. рис. 15.3).
Самые знаменитые группы ризарий — фораминиферы и радиолярии. Это морские амебоподобные существа со сложными минеральными скелетами. Большинство фораминифер живет на морском дне, и только немногие из них парят в водной толще. Их раковины чаще всего многокамерные, иногда трубчатые, а иногда спиральные (хотя есть и другие формы). “Фораминиферы” буквально значит “дырконесущие”. Называются они так потому, что в раковине фораминиферы есть множество маленьких отверстий, сквозь которые высовываются тонкие ложноножки. Эти ложноножки ветвятся, переплетаются, сливаются и в результате образуют вокруг фораминиферы сложную сеть, которая благодаря разным способам сокращения ложноножек может служить и для питания, и для передвижения.

Особенно сложными формами отличались, например, глоботрунканиды — вымершие фораминиферы, которые жили в конце мезозойской эры. Их раковина не только была многокамерной, но еще имела разнообразные выросты и кили, помогавшие парить в толще воды. Конец мезозоя был эпохой теплого безледникового климата, когда огромные площади занимали мелководные окраинные моря (примерно как нынешние Балтийское и Желтое). Резкая зональность отсутствовала, работающих всепланетными “холодильниками” полярных шапок не было и в помине, поэтому глобальное перемешивание воды было гораздо слабее, чем в современности: массы холодных вод, погружающиеся в глубину и приводящие в движение всю океанскую толщу, тогда просто нигде не возникали. В мезозойском спокойном море взвешенные в воде фораминиферы могли спокойно делить между собой экологические ниши, специализируясь по узким интервалам глубин. К сожалению, глоботрунканиды вымерли вместе с динозаврами.
У радиолярий богатство форм вообще поразительно. В первую очередь это касается их твердых скелетов. Тут можно увидеть вставленные друг в друга дырчатые шары, эллипсоиды, диски, конусы, треножники, короны, многолопастные и шлемовидные фигуры, и все это бывает оснащено длинными иглами, палочками, мечевидными выростами, решетками, перемычками или кольцами. Великолепными формами радиолярий восхищался еще Эрнст Геккель, чьей узкой научной специальностью они как раз и были (а поскольку он к тому же профессионально рисовал, то многими изображениями радиолярий в его исполнении можно и сейчас полюбоваться). Все радиолярии живут в толще морской воды, и сложные формы с выростами нужны им для того, чтобы лучше там парить. Как и у фораминифер, у радиолярий есть сеть переплетающихся ложноножек, которыми они ловят пищевые частицы. Но, кроме того, у них есть особые длинные неветвящиеся ложноножки, которые называются аксоподиями. Внутри каждой аксоподии проходит укрепляющий ее пучок микротрубочек, еще и связанных между собой (в обычных ложноножках ничего подобного не бывает). Аксоподии нужны радиоляриям в основном, опять же, для лучшего парения.
Многообразие ризарий не ограничивается фораминиферами и радиоляриями. Среди них есть типичные жгутиконосцы (в том числе хищные), есть амебы, а есть и паразиты. Например, именно к ризариям относится плазмодиофора, возбудитель так называемой килы капусты. Ее питающаяся стадия — это живущие в клетках капусты многоядерные амебы. Такую жизненную форму принято называть плазмодием.
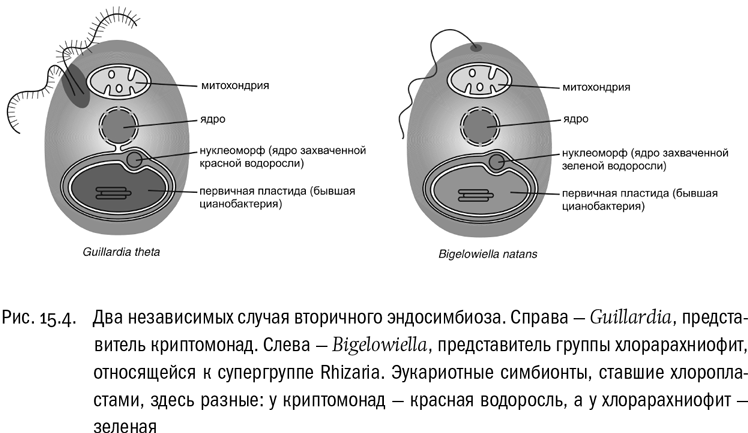
Интересная группа ризарий — хлорарахниофитовые водоросли. В эту группу входят самые настоящие фотосинтезирующие амебы (с характерными для ризарий длинными тонкими ложноножками), хотя у некоторых из них есть и жгутики. Разумеется, хлорарахниофиты имеют хлоропласты, которые в данном случае окружены четырьмя мембранами и представляют собой захваченные одноклеточные зеленые водоросли — эукариоты из супергруппы Plantae. Скоро мы увидим, что ситуация, когда хлоропласты образуются из захваченных эукариот (а не прокариот), встречается в эволюции на самом-то деле очень часто.
“Chromalveolata”
Название этой супергруппы, единственной из шести, в заголовке поставлено в кавычки. Вызвано это тем, что современная систематика признает ее выделенной некорректно — вскоре мы узнаем почему. Тем не менее члены этой группы действительно близки друг к другу, и мы можем для удобства рассмотреть их вместе, а поправки (которых требуют новые данные) внести на следующем этапе: так будет понятнее.
Мы уже не раз говорили о том, что все эукариоты, умеющие фотосинтезировать, получили эту способность благодаря симбиозу. Точнее — эндосимбиозу, один из участников которого живет внутри другого. Эукариотная клетка поглотила прокариотную и “поработила” ее, оставив жить внутри себя в качестве хлоропласта. Именно поэтому хлоропласты до сих пор размножаются делением.
Но возможна и другая ситуация. Эукариотная клетка может проглотить, “поработить” и превратить в хлоропласт другого эукариота, внутри которого уже есть собственный прокариотный симбионт. Это называется вторичным эндосимбиозом. Два таких примера мы уже видели: эвглена и амебы-хлорарахниофиты, хлоропласты которых — это бывшие зеленые водоросли. Но у хромальвеолят это не редкость, а общее явление, определившее эволюцию всей супергруппы. Их хлоропласты — вторичные (см. рис. 15.4). Среди хромальвеолят очень много фотосинтезирующих форм, и все их хлоропласты произошли от захваченных эукариот, а именно от красных водорослей. Окрашены они обычно не в зеленые тона, а в желтовато-бурые. Иногда проглоченный эукариотный симбионт даже сохраняет маленькое ядро, которое в этом случае называется нуклеоморфом. Чаще ядро симбионта исчезает, но сохраняются другие следы вторичного эндосимбиоза в виде дополнительных мембран.
Более того, у одноклеточных хромальвеолят иногда встречается третичный эндосимбиоз: захват эукариотного симбионта, внутри которого на правах хлоропласта живет еще один эукариотный симбионт, и только уже внутри того — прокариотный симбионт, который, собственно, и фотосинтезирует. Если один представитель хромальвеолят захватит другого в качестве хлоропласта, именно так и получится. В случае третичного эндосимбиоза одна клетка объединяет в себе четыре организма: двух разных хромальвеолят, красную водоросль и цианобактерию, ставшую первичным хлоропластом. И это не считая митохондрий.
В группу хромальвеолят (в ее традиционном объеме) входит не меньше семи больших групп преимущественно водных фотосинтезирующих эукариот, то есть водорослей. Это криптомонады, гаптофиты, динофлагелляты, золотистые, желто-зеленые, бурые и диатомовые водоросли. Из этих семи групп четыре (криптомонады, гаптофиты, динофлагелляты и диатомовые) состоят в основном из одноклеточных форм. Колонии они образуют нечасто, а если образуют, то относительно простые. Зато некоторые из этих одноклеточных водорослей живут огромными массами в освещенной зоне океана и играют там примерно ту же роль, что зеленые растения в наземных экосистемах: изо всех сил фотосинтезируют, создавая так называемую первичную продукцию. В первую очередь это относится к динофлагеллятам, диатомовым водорослям и гаптофитам. Последние знамениты тем, что микроскопические известковые пластинки, которыми покрыты снаружи их клетки, образуют основу писчего мела. Воспетые многими поэтами Дуврские белые скалы, что в графстве Кент, сложены в основном из остатков панцирей мельчайших планктонных водорослей-гаптофит, которые когда-то жили в теплых мелководных морях мезозойской эры.
С золотистыми и желто-зелеными водорослями дело обстоит иначе. В этих группах есть обычные жгутиконосцы, есть амебоидные существа, есть гигантские многоядерные клетки, есть колонии, а есть и многоклеточные организмы, нитчатые или пластинчатые. Золотистые и желто-зеленые водоросли — это две совершенно отдельные эволюционные линии. Их предками были одноклеточные жгутиконосцы, хоть и родственные, но все-таки разные. Все этапы усложнения (во многом одни и те же!) эти две линии водорослей прошли независимо друг от друга, хотя и на основе набора общих генов, связанного с близким родством. Это прекрасный пример того, что в эволюционной биологии называется параллелизмом.
Наконец, группа бурых водорослей целиком состоит из многоклеточных организмов. Почти все они живут в морях. И некоторые из них более-менее сравнимы по сложности с хорошо нам знакомыми наземными зелеными растениями — например, ламинария, она же морская капуста. В фантастическом романе Фрэнка Герберта и Билла Рэнсома “Ящик Пандоры” подводные леса из бурых водорослей (их там называют “электрокелпом”) даже обзавелись чем-то вроде собственного коллективного разума.
Кроме всевозможных водорослей среди хромальвеолят есть и много нефотосинтезирующих существ. Есть бесцветные гетеротрофные жгутиконосцы, поедающие бактерий. Есть оомицеты — потомки каких-то морских водорослей, потерявшие хлоропласты и вернувшиеся к гетеротрофности. Они стали осмотрофами (“всасывальщиками”) и приобрели в связи с этим жизненную форму, в точности повторяющую жизненную форму типичных грибов: тело, состоящее из ветвящихся тончайших нитей толщиной в одну клетку. Долгое время их и принимали за грибы. Но к концу XX века выяснилось, что у оомицетов есть признаки, которых у настоящих грибов быть просто не может: клеточная стенка из целлюлозы (а не из хитина), трубчатые кристы митохондрий и подвижная одноклеточная расселительная стадия с двумя жгутиками. Таким образом, сходство оомицетов с грибами — это чистой воды конвергенция, независимое приобретение общих черт, не связанное с родством. В отличие от большинства грибов, оомицеты сохраняют жгутиковые стадии и (как правило) связь с водой. Типичный представитель оомицетов — сапролегния, белесая водная плесень.
Случалось и такое, что оомицеты вмешивались в человеческую историю. Великий голод в Ирландии в 1845–1849 годах произошел из-за фитофторы — оомицета, вызывающего картофельную гниль. Ирландские крестьяне тогда были в массе своей бедны, и картошка составляла значительную долю их рациона. Поэтому, когда практически все посевы картофеля в стране оказались поражены эпидемией фитофторы, это стало настоящей катастрофой. Достаточно сказать, что население Ирландии после нее сократилось примерно на треть в результате как гибели от голода, так и эмиграции в Америку людей, пытавшихся от него спастись.
Очень интересная группа хромальвеолят — опалины. Это одноклеточные, но многоядерные существа, живущие в конечном отделе кишечника амфибий (реже рыб и рептилий). Особого вреда своим хозяевам они, судя по всему, не наносят, так что их можно считать не паразитами, а просто сожителями. Опалины — обычнейшие обитатели клоаки лягушек (клоакой у животных называют расширенную часть задней кишки, в которую впадают выделительные и половые протоки). Клетка опалины может нести несколько тысяч жгутиков, которые в этом случае уже можно называть ресничками. Они расположены рядами и бьются согласованными волнами, давая возможность опалине довольно быстро плавать. Опалины, без сомнения, произошли от обычных одноядерных жгутиконосцев, но по ходу эволюции их клетка постепенно увеличивалась и усложнялась. Мелкие (и, по-видимому, примитивные) опалины имеют всего два клеточных ядра, а у крупных в клетках бывает до 200 ядер и больше. Причем все эти ядра одинаковы — по крайней мере, с виду.
Именно к хромальвеолятам, как выяснилось, относятся и инфузории — одноклеточные существа, известные натуралистам еще с XVII века. Первыми исследователями инфузорий были создатели самых ранних микроскопов — великий натуралист-любитель Антони ван Левенгук и не менее великий физик Христиан Гюйгенс. Инфузорий иначе называют ресничными, потому что их тело, как правило, покрыто несколькими тысячами типичных ресничек. Кроме того, для инфузорий характерна очень сложная клетка с постоянным клеточным ртом и системой внутренних опорных структур. Внешняя форма клетки у них тоже постоянная, часто выразительная и красивая, о чем свидетельствуют названия: инфузория-туфелька, инфузория-трубач. Неудивительно, что биологи довольно долго принимали инфузорий за животных. Тем не менее на самом деле инфузории — самые настоящие одноклеточные, причем даже не многоядерные. Чаще всего в клетке инфузории два ядра: маленький микронуклеус (генеративное ядро, содержащее полный геном и служащее для продолжения рода) и большой макронуклеус (соматическое ядро, содержащее только ту часть генома, что нужна для повседневной жизни, но зато в огромном числе копий). Ядерный дуализм, то есть разделение функций между микронуклеусом и макронуклеусом, — это, пожалуй, самая главная особенность инфузорий как группы.
Сложность устройства инфузорий настолько впечатляет, что некоторые биологи всерьез считали их предками многоклеточных животных. Но это, разумеется, не так. Инфузории тем и интересны, что они нашли свой собственный путь эволюции, альтернативный по отношению к любой многоклеточности.
Многие хромальвеоляты стали паразитами. В первую очередь это группа Apicomplexa (название, буквально означающее что-то вроде “существа со сложным передним концом”). Апикомплексы произошли от жгутиконосцев, и сейчас известно, от каких именно. Но в процессе глубокого приспособления к паразитизму они изменились очень сильно — во многих случаях до неузнаваемости. Именно апикомплексы составляли большую часть сборной группы паразитических эукариот, которых со времен Карла фон Зибольда называли споровиками. К апикомплексам относятся, например, возбудители малярии и токсоплазмоза.
Интересно, что апикомплексы совершенно точно произошли от фотосинтезирующих предков. У этих специализированных паразитов есть апикопласт — остаток вторичного хлоропласта, ставший бесцветным, но сохранивший четыре мембраны и собственную ДНК. Сохранился он потому, что помимо фотосинтеза хлоропласты у предков апикомплексов занимались синтезом жирных кислот и некоторых других полезных веществ. После отказа от фотосинтеза эти метаболические функции никуда не делись, а вместе с ними уцелели и остатки хлоропластов, дающие прямое свидетельство сложного эволюционного пути своих хозяев. В глубокой древности предок апикомплексов был обычным хищным жгутиконосцем. Однажды он проглотил водоросль (скорее всего, красную) и сделал ее своим хлоропластом, а тем самым стал водорослью и сам. Затем произошел отказ от фотосинтеза. Обесцветившийся жгутиконосец вернулся к гетеротрофному питанию и стал в конце концов специализированным паразитом. Однако в соответствии с законом необратимости эволюции в его организме сохранились следы пройденного эволюционного пути — в данном случае не только генетические, но и структурные (апикопласт виден в микроскоп). Когда в 1898 году англичанин Рональд Росс и итальянец Джованни Батиста Грасси раскрыли жизненный цикл возбудителя малярии, никто из них, конечно, не мог и представить, что этот опаснейший микроб окажется потомком безобидной одноклеточной водоросли.
В целом среди хромальвеолят можно выделить две очень крупные эволюционные ветви. Одна из них нам уже знакома — она называется альвеолятами (Alveolata). К этой ветви относятся инфузории, апикомплексы, динофлагелляты. Главная особенность альвеолят — их уже упоминавшаяся оболочка, которая называется пелликулой. Под внешней мембраной у них находится слой плотно сомкнутых маленьких мембранных пузырьков (так называемых альвеол), которые, в свою очередь, подостланы микротрубочками. Это и есть пелликула. Она придает клетке устойчивую форму, а заодно позволяет устанавливать на базе имеющейся оболочки всевозможные дополнения — вроде системы белковых корешков, синхронизирующих у инфузорий биение ресничек, или сложного аппарата проникновения в клетку хозяина, который расположен на конце клетки апикомплексов.
Другая крупнейшая ветвь хромальвеолят называется Stramenopiles. Группу эту выделил в 1989 году протистолог Дэвид Паттерсон. Подвижные клетки страменопилов обычно имеют два жгутика: двигательный, направленный вперед и опушенный, и рулевой, отогнутый назад и гладкий (либо опушенный заметно слабее). Причем волоски, которыми опушен двигательный жгутик, у них имеют особую трубчатую структуру. Такое состояние называется гетероконтным (разножгутиковым). Название страменопилов отражает именно этот признак. Если буквально истолковать вошедшие в это название латинские корни, получится что-то вроде “имеющие солому в волосах”: с соломинками тут сравниваются сидящие на двигательном жгутике волоски. К страменопилам относятся золотистые, желто-зеленые, бурые и диатомовые водоросли, а также оомицеты и опалины. У опалин разножгутиковость исчезает, но они произошли от жгутиконосцев, у которых она точно была.
Есть и такие хромальвеоляты, которые не относятся ни к альвеолятам, ни к страменопилам. Из тех, о ком мы тут говорили, это две группы одноклеточных: криптомонады и гаптофиты. Возможно, это очень древние эволюционные линии.
К большому сожалению, современные генетические данные (причем не такие уж и новые) показывают, что хромальвеоляты, скорее всего, не являются группой, соответствующей требованиям филогенетической систематики. Таксон Chromalveolata, конечно, состоит из родственных друг другу форм, но они объединены в него, по нынешним меркам, слишком произвольно. В одной своей части эта группа получилась сборной, а в другой, наоборот, неполной, с “фигурным вырезом”. Поэтому таксон Chromalveolata на данный момент надо считать устаревшим. О том, как обстоят дела в реальности, мы поговорим чуть ниже.
Plantae
Эта супергруппа состоит в основном из фотосинтезирующих организмов. Она единственная, где нет ни одной крупной эволюционной ветви, освоившей гетеротрофность. Поэтому название Plantae — растения — ей вполне подходит (см. рис. 15.5.).
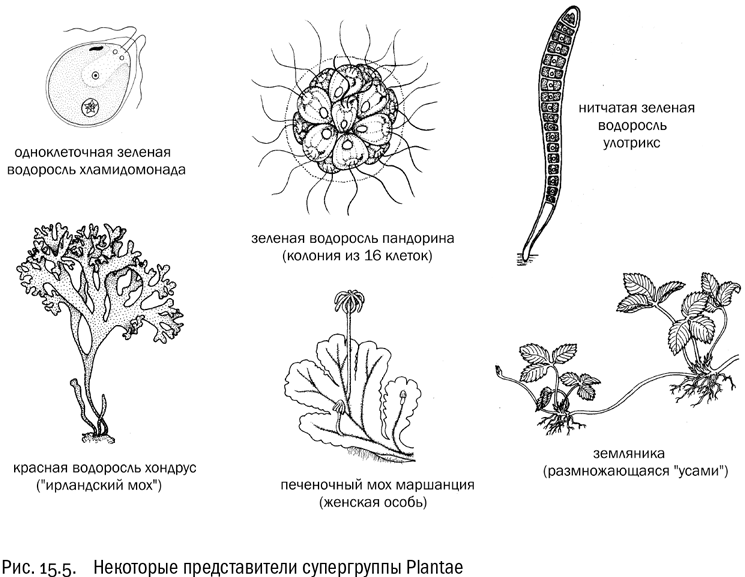
У супергруппы растений есть несколько общих особенностей. Во-первых, это пластинчатые кристы митохондрий (у ризарий и у большинства хромальвеолят они трубчатые). Во-вторых, подвижные клетки у растений чаще всего имеют два одинаковых гладких жгутика, направленных вперед. Такое состояние называется изоконтным (равножгутиковым). Отличный пример равножгутиковости — хламидомонада, одноклеточная зеленая водоросль, описанная в школьных учебниках ботаники.
Есть и третья очень важная особенность. Хлоропласты растений — первичные, то есть возникшие путем первичного эндосимбиоза. Они произошли непосредственно от захваченных прокариот, а именно от цианобактерий. Этим растения резко отличаются, например, от бурых или золотистых водорослей, хлоропласты которых — вторичные, происходящие от эукариотных симбионтов, то есть как раз от захваченных представителей Plantae.
В состав супергруппы растений входит всего три эволюционные ветви. Во-первых, это одна сильно обособленная (и крайне примитивная по некоторым признакам) группа водорослей, которая называется глаукофитами. Они в основном одноклеточные, а если образуют колонии, то очень простые. Во-вторых, это красные водоросли. И в-третьих, это огромная и разнообразная эволюционная ветвь, для которой Томас Кавалье-Смит предложил латинское название Viridiplantae (зеленые растения). В нее входят как наземные зеленые растения, так и их многочисленные родственники-водоросли, крайне разнообразные по уровню организации.
Именно к ветви Viridiplantae принадлежат и самые мелкие, и самые крупные современные эукариоты — по крайней мере, если не учитывать внутриклеточных паразитов (которые бывают очень маленькими) и разрастающиеся грибницы (которые, наоборот, бывают огромными, но все же не являются компактными, истинно многоклеточными телами). Самым мелким свободноживущим эукариотом считается океанская планктонная зеленая водоросль остреококкус: размер ее клетки меньше микрона. А крупнейшие истинно многоклеточные организмы на Земле — это стометровые секвойи и секвойядендроны, самые громадные экземпляры которых получают собственные имена: “Генерал Шерман”, “Гиперион” и т.п.
Первые зеленые растения, несомненно, были одноклеточными жгутиконосцами. Но в ходе своей долгой и бурной эволюции они успели дать огромный спектр жизненных форм. Среди них есть одноклеточные без всяких жгутиков (например, хлорелла), есть шарообразные колонии из нескольких сотен или тысяч жгутиковых клеток (например, вольвокс), есть гигантские многоядерные клетки, есть нитчатые и пластинчатые примитивные многоклеточные, а есть и обладатели довольно сложных ветвящихся тел, как у харовых водорослей. Зеленые водоросли (в широком смысле этого слова) живут в любых водоемах от луж до Мирового океана, а иногда даже и на суше. От кого же из них произошли наземные растения?
Прообразом наземных растений могут считаться или пластинчатые колеохетовые водоросли, или нитчатые водоросли-сцеплянки. Именно к этим двум группам наземные растения ближе всего, судя по генетическим данным . Водоросль, которая была предком наземных растений, жила на мелководье и представляла собой либо однослойную пластинку, либо переплетение нитей. Большой разницы тут нет, потому что эти жизненные формы могут легко переходить друг в друга. Водоросли жили в переходной зоне между водоемом и берегом, которая была в те древние времена чрезвычайно протяженной: тут надо учитывать, что до появления сплошного растительного покрова, удерживающего почву, реки на Земле не имели постоянных берегов, и мелководьем была, по сути, почти вся суша. Есть вполне правдоподобная гипотеза, что “изобретением”, запустившим эволюцию наземных растений, был стволик с коробочкой наверху, поначалу предназначенный исключительно для разбрасывания воздушных спор, чтобы их разносил ветер . Нет сомнений, что этот способ расселения очень эффективен (на современной Земле он позволяет растениям осваивать расположенные за тысячи километров от ближайшей суши вулканические острова). Именно для того, чтобы его обеспечить, могли бы появиться первые механические и проводящие ткани, совершенно ненужные пластинчатой или нитчатой водоросли в воде, но необходимые воздушному стволику. А уж потом наличие этих тканей позволило растениям полностью выйти на сушу и создать наземные жизненные формы. И в результате мы теперь живем в мире, где целые природные зоны определяются наземными растениями: от степных злаков до самых многочисленных деревьев на современной Земле — лиственниц, образующих сибирскую и североамериканскую светлохвойную тайгу.
Главным признаком, отличающим высшие растения от водорослей, традиционно считается архегоний — колбообразный мешочек, в котором созревает яйцеклетка. Это настоящий многоклеточный орган размножения. Причем зародыш организма следующего поколения, уже в свою очередь многоклеточный, какое-то время развивается прямо внутри архегония. Именно с этим связано латинское название высших растений — Embryophyta. К этой группе относятся мхи, плауны, хвощи, псилоты, папоротники, голосеменные (хвойные, саговники, гинкго) и главные звезды современного растительного мира Земли — цветковые. Правда, у голосеменных от архегониев остаются буквально считаные клетки, а у цветковых и вовсе ничего, но тут важно то, что они были у их предков.
Amoebozoa
Супергруппа Amoebozoa — это царство амеб (см. рис. 15.6). Она состоит из существ, которые передвигаются амебоидным способом, выпуская и втягивая ложноножки и постоянно меняя форму клетки. Очень характерна форма самих ложноножек: они не длинные и тонкие, как у ризарий, а широкие, лопастевидные. У некоторых амебозоев одновременно с ложноножками есть и жгутики. Кристы митохондрий у них обычно трубчатые, хлоропластов не бывает. Все амебозои — гетеротрофы. Некоторые из них потеряли митохондрии, потому что живут или в бескислородном иле, или в чьем-нибудь кишечнике (как дизентерийная амеба, например).
Обыкновенная амеба, она же амеба-протей, с которой по традиции начинается школьный учебник зоологии, — довольно типичный представитель амебозоев. Она относится к голым амебам, у которых нет никакого покрова сверх плазматической мембраны. Но бывают и раковинные амебы, причем с очень разнообразной формой и структурой раковинок. И голые, и раковинные амебы могут жить не только в водоемах, но и в почве: находящейся там капиллярной воды им вполне хватает, чтобы оставаться водными существами.
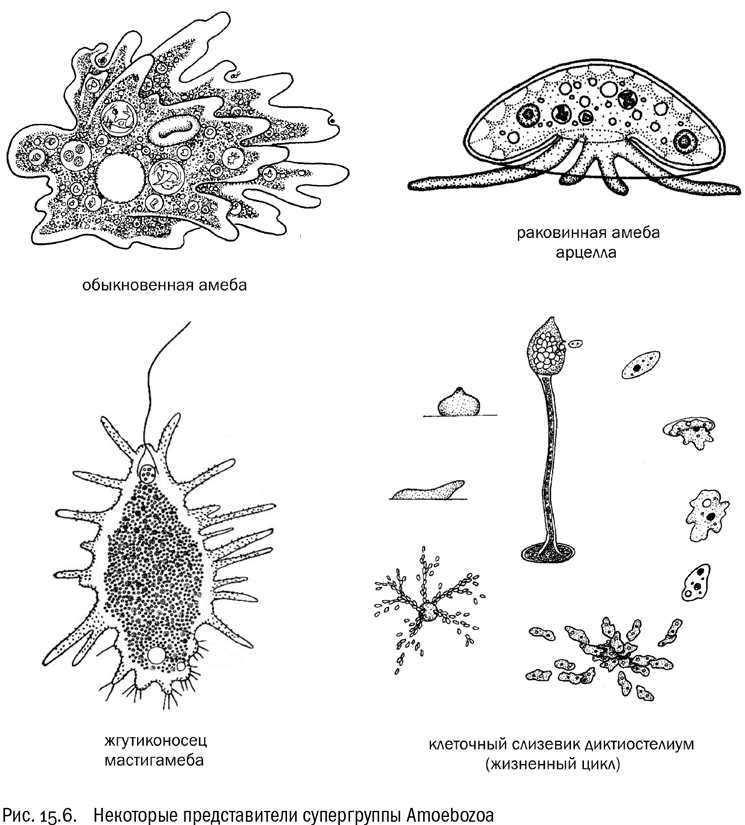
В то же время не будем забывать, что “амеба” (точно так же, как и “жгутиконосец”), — это название не эволюционной ветви, а жизненной формы. Увидев под микроскопом незнакомую амебу, мы, строго говоря, еще ничего не знаем о ее положении в системе. Вполне нормальные амебы, пришедшие к этой жизненную форме совершенно самостоятельно, есть и среди ризарий, и среди экскават. Но амебозои — единственная супергруппа, где этот способ жизни безусловно преобладает.
Существуют амебы, которые в определенный момент своего жизненного цикла приобретают многоклеточность (во всяком случае, примитивную). Большую часть времени они могут ползать поодиночке где-нибудь в почве. Но когда приходит время, эти амебы сползаются вместе и образуют многоклеточное плодовое тело, очень похожее на плодовое тело гриба. В другом случае странствующие амебы сначала образуют плазмодий — гигантскую многоядерную амебу, которая ползает и питается, пока не настанет пора создавать плодовое тело. Потом плодовое тело выбрасывает споры, и все повторяется. Организмы с такими жизненными циклами называются слизевиками.
Жизненная форма слизевика распространена на удивление широко. Например, сейчас мы знаем, что свои собственные слизевики с настоящими плодовыми телами, совершенно независимо освоившие этот образ жизни, есть в супергруппе экскават. Они называются акразиевыми слизевиками, или акразиомицетами. Это еще один яркий пример эволюционной конвергенции. Более того, организмы, близкие по типу жизненного цикла к слизевикам, есть и в группе ризарий (плазмодиофоромицеты), и в группе страменопилов (лабиринтуломицеты). Что же касается “настоящих” слизевиков, которые входят в супергруппу амебозоев, то они называются миксомицетами.
Многие биологи рассматривают слизевиков как прекрасную модель начальных стадий эволюции многоклеточности. Есть даже осторожно высказываемая гипотеза, что примерно такой же жизненный цикл, как у типичных слизевиков, имели предки многоклеточных животных. Главная трудность этой гипотезы в том, что многоклеточные животные относятся к другой супергруппе эукариот — к последней, о которой нам осталось поговорить.
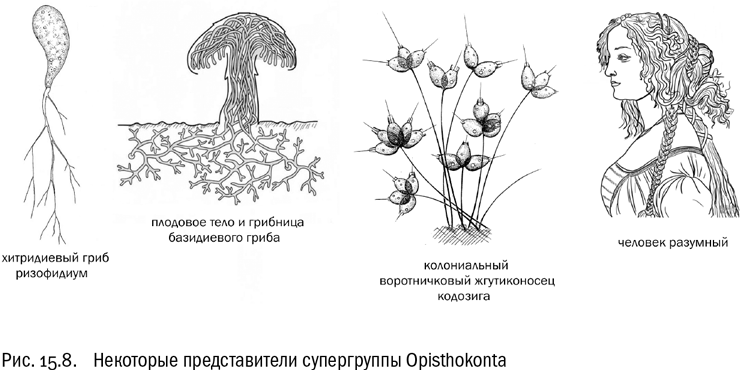
Opisthokonta
В эту супергруппу входят гетеротрофные существа с пластинчатыми кристами митохондрий и жгутиковыми клетками, имеющими единственный жгутик, направленный назад. Мы можем легко догадаться, что такое состояние называется заднежгутиковым, или опистоконтным.
Самое удивительное, что, кроме перечисленных признаков, у разных опистоконтов нет между собой почти ничего общего. По крайней мере, так кажется на первый взгляд. Сильно ли похожи друг на друга воротничковый жгутиконосец, белый гриб и синий кит? А ведь по меркам мегасистемы эукариот они близкие родственники.
На самом деле кое-что общее найти все же можно. Например, во внутреннем ухе позвоночных животных (в том числе и в человеческом) есть чувствительные элементы, которые называются волосковыми клетками (см. рис. 15.7). Вершина такой клетки несет один жгутик (его функция тут опорная) и группу собранных вместе микроворсинок. Если внешняя сила — например, течение жидкости в полости внутреннего уха — сгибает или наклоняет эти микроворсинки, то клетка генерирует электрический сигнал. Это основной принцип работы внутреннего уха. Так вот, давно замечено, что волосковая клетка внутреннего уха напоминает по структуре клетку воротничкового жгутиконосца. Компоненты там те же самые: жгутик и группа микроворсинок на вершине клетки, они только смещены относительно друг друга. И это отнюдь не поверхностное сходство. Недавно исследователи показали, что в волосковых клетках внутреннего уха позвоночных и во внутренних клетках губок, которые называются хоаноцитами, экспрессируется целая группа общих генов. В основном эти гены кодируют белки, связанные с цитоскелетом и участвующие в образовании микроворсинок. А хоаноциты губок — это клетки, в точности похожие на воротничковых жгутиконосцев, уж тут преемственность очевидна. Порядок ветвлений эволюционного древа животных вполне позволяет допустить, что эта преемственность “дотянулась” и до позвоночных.
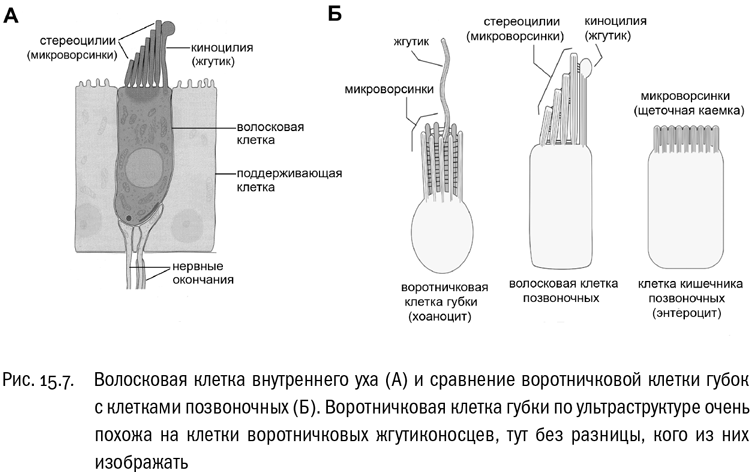
Про сперматозоиды животных мы уже упоминали. Сперматозоид — обладатель типичного жгутика (правда, в некоторых группах животных встречаются безжгутиковые сперматозоиды, но это явно производное состояние). И плывет он жгутиком назад. Это и есть признак, характерный для опистоконтов, но не встречающийся у других эукариот. В этом отношении животные более архаичны, чем грибы, у большинства из которых все жгутиковые стадии исчезли.
Итак, опистоконты — это грибы, животные и воротничковые (см. рис. 15.8). Но не только они. Можно назвать по меньшей мере четыре группы “странных опистоконтов”, не относящихся ни к многоклеточным животным, ни к воротничковым жгутиконосцам, ни к типичным безжгутиковым грибам. Это хитридиевые грибы, нуклеарииды, криптомицеты и мезомицетозои. Все эти группы относительно малоизвестны (во всяком случае, у широкой публики они не на слуху). Но это вовсе не значит, что они неважны для понимания структуры древа жизни.
Хитридиевые грибы, или хитридиомицеты, — это преимущественно водные организмы. Тело примитивного хитридиевого гриба представляет собой фактически амебу: одноядерную или многоядерную, но в любом случае с длинными, тонкими и сильно ветвящимися ложноножками, которые глубоко проникают в питательный субстрат. От таких ложноножек один шаг до грибницы, и неудивительно, что этот шаг был в ходе эволюции легко сделан: у высших хитридиевых налицо уже типичная грибница (амебоидное тело в этом случае покрывается клеточной стенкой и теряет подвижность). Тем не менее у хитридиомицетов, в отличие от других грибов, сохраняются плавающие жгутиковые споры — разумеется, со жгутиком, обращенным назад.
Нуклеарииды — это пресноводные или почвенные амебы с длинными тонкими ложноножками, эволюционно близкие к грибам. Легко представить, что от кого-то вроде них грибы как раз и произошли. Хотя скоро мы увидим, что мнения на эту тему есть разные.
Криптомицеты — одноклеточные организмы, безжгутиковые или с одним жгутиком, лишенные клеточной стенки, близкие, опять же, к грибам и паразитирующие внутри клеток других грибов или водорослей. Их считают примитивными родственниками грибов, которым к тому же удалось необычайно глубоко специализироваться в сторону паразитизма.
Но вот тут есть большая проблема. Кроме криптомицетов существует еще целых две группы специализированных внутриклеточных паразитов, близких к примитивным грибам: афелиды и микроспоридии. Афелиды паразитируют в клетках водорослей, а микроспоридии — в клетках животных, включая и человека. Мы между делом упоминали в главе 12, что микроспоридии умудрились стать не простыми паразитами, а энергетическими: они выкачивают из хозяйской клетки АТФ. Современные молекулярно-биологические исследования приводят к выводу, что криптомицеты, афелиды и микроспоридии составляют единую эволюционную ветвь, сестринскую по отношению ко всем грибам . Для этой единой ветви предложено название Opisthosporidia (опистоспоридии). И что характерно, все ее члены — внутриклеточные паразиты. Не значит ли это, что внутриклеточным паразитом был и общий предок опистоспоридий с грибами? Не обязательно — но не исключено.
Мезомицетозои — группа амебообразных организмов, которые чаще всего паразитируют в каких-нибудь морских или пресноводных многоклеточных животных. Жгутиков у них обычно нет, но есть ложноножки. Питаются они примерно как грибы: осмотрофно, всасывая вещества сквозь клеточную мембрану. Известно, что на некоторых стадиях жизненного цикла у мезомицетозоев бывает клеточная стенка — по-видимому, хитиновая, как и у грибов. Но еще интереснее, что некоторые мезомицетозои образуют в ходе своего размножения колонии покоящихся амеб, удивительно похожие на ранние зародыши многоклеточных животных. Причем показано, что деления клеточных ядер в такой колонии синхронизируются — тоже как у зародышей многоклеточных животных. Это означает, что клетки “зародыша” (или не клетки, а только их ядра, если клетки на этом этапе слиты) делятся не вразнобой, но строго одновременно, так что их число остается степенью двойки: 4, 8, 16, 32.
Мезомицетозои — эволюционные “кузены” многоклеточных животных (в одной статье их прямо так и назвали). “Братья” многоклеточных животных, по генетическим данным, воротничковые жгутиконосцы: это самая близкая к ним группа. Но при этом жизненный цикл воротничковых жгутиконосцев, даже колониальных, слишком прост, чтобы быть для многоклеточных животных предковым. Например, никакого механизма синхронизации клеточных делений при образовании колонии у воротничковых, судя по всему, нет. В этом отношении мезомицетозои более продвинуты. Кстати, у некоторых мезомицетозоев встречаются ультратонкие ложноножки, из которых вполне могли бы получиться микроворсинки “воротничка”.
В 2010 году у мезомицетозоев обнаружили свойственный многоклеточным животным ген, который называется Brachyury. У животных этот ген кодирует один регуляторный белок, активно участвующий в эмбриональном развитии. Например, у позвоночных экспрессия гена Brachyury важна для нормального развития основы осевого скелета — хорды — и прилегающих к ней зачатков (вдаваться в эмбриологические детали мы тут не будем). Возникает естественный вопрос: зачем этот ген существу, у которого нет не то что хорды, а вообще никаких органов? Данных на эту тему пока маловато, но кое-что понять уже можно. Известно, например, что у одного из мезомицетозоев ген Brachyury активен в момент, когда его многоядерный плазмодий распадается на множество одноядерных амеб. Очевидно, он как-то регулирует происходящие во время развития сложные взаимные перемещения клеток — собственно, как и у животных. В любом случае мы знаем, что этот ген очень эволюционно консервативен: его продукт, взятый у одноклеточной амебы, может запустить развитие осевых структур в зародыше лягушки — правда, не с таким успехом, как продукт гена самой лягушки. Но сам факт, что белковые продукты столь специализированных генов амебы и лягушки оказались взаимозаменяемыми, уже поразителен.
Естественно, исследователи заинтересовались вопросом, у кого еще есть ген Brachyury. Выяснилось, что за пределами супергруппы опистоконтов он (равно как и родственные ему гены) не встречается нигде. Зато внутри опистоконтов распространен очень широко. Ген Brachyury есть у всех многоклеточных животных, у самых разных мезомицетозоев, у криптомицетов, у хитридиевых грибов и еще у одной относительно примитивной группы грибов, которая называется зигомицетами. Унаследовать его они могли только от общего предка всех опистоконтов: конфигурация эволюционного древа просто не оставляет других возможностей. А вот у воротничковых жгутиконосцев и у высших грибов гена Brachyury нет. Причем из положения этих групп на древе однозначно следует, что у их предков он когда-то был, но оказался утрачен за ненадобностью.
Итак, ген Brachyury имеется у мезомицетозоев и у многоклеточных животных, но потерян у воротничковых жгутиконосцев (которые, напомним, на эволюционном древе находятся к животным ближе всего). По-видимому, из этого неумолимо следует, что жизненный цикл современных воротничковых жгутиконосцев вторично упрощен. У реального предка многоклеточных животных он наверняка был гораздо сложнее. Увы, мы не знаем точно, каким был этот предок. Эволюционное древо опистоконтов вообще полно провалов: по нему видно, что многие переходные формы давно вымерли и, скорее всего, не уцелели даже в палеонтологической летописи. Что-то (но не все) сохранили от общих предков мезомицетозои, а что-то (но не все) — воротничковые. То, что предок животных сочетал в себе некоторые признаки обеих этих групп, можно утверждать наверняка. Кроме того, в последнее время накапливаются молекулярные данные, позволяющие считать, что мезомицетозои — это не эволюционная ветвь, а эволюционный уровень, объединяющий всех примитивных родственников животных . Тогда прямые предки животных просто неизбежно должны были через этот уровень так или иначе пройти.
Скорее всего, когда-то мезомицетозои (или их близкие родственники) были свободноживущими и населяли море. Но затем их постигла обычная судьба архаистов, переживших свое время: они были вытеснены из большинства местообитаний и сумели уцелеть только ценой глубокой специализации, в данном случае — ухода в паразитизм. Причем вполне вероятно, что вытеснили мезомицетозоев их же собственные потомки. “Эволюция уничтожает свои причины”. Подобные примеры история жизни на Земле знает. Правда, в данном случае мезомицетозои “в отместку” научились на своих потомках паразитировать.
Теперь мы можем подвести некоторые итоги, и прежде всего — понять, как в целом выглядит эволюционное древо супергруппы опистоконтов (см. рис. 15.9). Эта супергруппа состоит из двух больших ветвей: Holomycota (“все грибы”) и Holozoa (“все животные”). К ветви Holomycota относятся нуклеарииды, опистоспоридии, хитридиомицеты и высшие грибы. К ветви Holozoa — мезомицетозои (в широком смысле этого слова), воротничковые жгутиконосцы и многоклеточные животные.
Высшие грибы — группа, эволюция которой связана с сушей. По глубине приспособленности к наземному образу жизни с ними могут сравниться разве что цветковые растения (животные остаются далеко позади). У всех грибов, кроме хитридиевых, жизненный цикл полностью лишен жгутиковых стадий. У них не бывает никаких подвижных половых клеток, а расселяются они с помощью спор, рассчитанных на перенос по воздуху. Поэтому размножение грибов не зависит от наличия жидкой воды. Правда, грибнице все равно нужна для роста высокая влажность, но независимость размножения от воды тем не менее заметно расширяет их экологические возможности. Примерно 98% современных видов грибов относится к группе Dikarya, у которой половой процесс, как правило, сводится к обмену клеточными ядрами между вступившими в контакт нитями разных грибниц. В частности, к этой группе относятся все шляпочные грибы, плодовые тела которых мы называем “грибами” в обыденной жизни.
Раньше грибами считались еще и оомицеты, которые на самом деле, как мы теперь знаем, представляют собой бесхлоропластные разножгутиковые водоросли, относящиеся к группе страменопилов. Среди прочих признаков грибы отличаются от оомицетов способом синтеза лизина — аминокислоты, которая входит в состав белков (см. главу 3). Формула лизина следующая: CH2(NH2)–CH2–CH2–CH2–CH(NH2)–COOH. У большинства эукариот, в том числе и у оомицетов, лизин синтезируется через диаминопимелиновую кислоту (HOOC–CH(NH2)–CH2–CH2–CH2–CH(NH2)–COOH). Это так называемый ДАП-путь. Но у грибов лизин синтезируется другим путем, через альфа-аминоадипиновую кислоту (HOOC–CH2–CH2–CH2–CH(NH2)–COOH). Это так называемый ААА-путь. Из этих двух путей ДАП-путь — намного более распространенный и, судя по всему, более древний. Почему же грибы перешли на ААА-путь? Очевидно, их предки на каком-то эволюционном этапе вообще потеряли способность синтезировать лизин, а потом восстановили ее с нуля. И в соответствии с законом необратимости эволюции восстановление получилось неточным. Но почему же предки грибов разучились синтезировать лизин? Не потому ли, что они были внутриклеточными паразитами и могли получать аминокислоты прямо из клеток хозяев? Это вполне логичное объяснение, но верно ли оно, мы пока не знаем.
Многоклеточные животные — группа, которая по-латыни называется Metazoa. Это единственная во всей живой природе Земли эволюционная ветвь, в которой есть многоклеточные хищники. Обратим внимание, что практически все жизненные формы, о которых мы раньше говорили — колониальные и многоклеточные водоросли, амебы, слизевики, грибоподобные осмотрофы, — возникали в разных группах эукариот по несколько раз. В эволюции это обычное дело. И только многоклеточные хищники составляют тут исключение: у них аналогов нет.
Хищниками, однако, можно назвать далеко не всех многоклеточных животных. Группа Metazoa включает пять эволюционных ветвей: губки, пластинчатые, гребневики, стрекающие и билатерии, они же двусторонне-симметричные (см. рис. 15.10). Губки — это сидячие неподвижные существа, которые проводят жизнь, будучи прикрепленными ко дну или подводным предметам, и питаются за счет фильтрации воды. А единственный представитель пластинчатых — трихоплакс, существо хоть и подвижное, но размером с крупную амебу (порядка миллиметра) и устроенное по меркам многоклеточных животных исключительно просто. Все, на что способны губки и трихоплакс, — это поглощать микроскопические пищевые частицы, которые им попадаются. При таком способе питания многоклеточность хоть и полезна, но не обязательна.
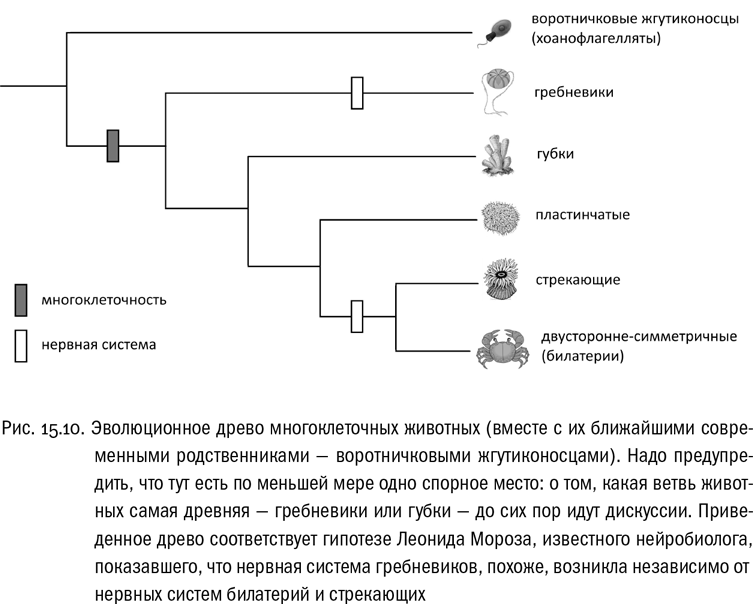
Совершенно иначе обстоит дело у животных, имеющих нервную систему, мышцы и рот. Иногда их называют “настоящими многоклеточными животными”. Они могут активно искать и захватывать крупную добычу, нередко сравнимую по размеру с самим хищником. Скорее всего, выход на эволюционный уровень “настоящих многоклеточных животных” произошел дважды: у гребневиков и у общего предка стрекающих с билатериями. Есть серьезные указания на то, что нервная система была независимо “изобретена” в этих двух эволюционных линиях . То же самое относится и к мускулатуре. У гребневиков она относительно слабая, эти животные плавают в основном за счет биения ресничек. У стрекающих и билатерий тоже может сохраняться ресничное движение (коловратки, плоские черви, многие морские личинки), но обычно оно сочетается с гораздо более эффективным мышечным движением.
В группу стрекающих входят медузы и полипы (гидры, актинии, кораллы). Стадии плавающей медузы и сидячего полипа часто чередуются у них в одном жизненном цикле. Почти все стрекающие — хищники. Признак, давший им название, — стрекательные клетки, которые поражают добычу выбрасывающимися нитями. Симметрия тела у стрекающих лучевая, с центральной осью и несколькими равноценными сторонами. Но есть гипотеза, что лучевая симметрия стрекающих — вторичная, возникшая на основе двусторонней симметрии, которая была у их ползающих предков. Если это верно, то первичная лучевая симметрия есть только у гребневиков.
И наконец, двусторонне-симметричные животные, или билатерии, — это необыкновенно разнообразная эволюционная ветвь, богатая оригинальными планами строения (см. рис. 15.11). В теле билатерий выделяются срединная плоскость, спинная и брюшная сторона, передний и задний концы. Часто оно делится еще и на сегменты. На переднем конце обычно концентрируются органы чувств и разрастаются нервные узлы. Чтобы обеспечивать развитие такого тела в каждом жизненном цикле, у билатерий возникло много новых регуляторных генов. А это, в свою очередь, способствовало бурной эволюции новых форм. Некоторые билатерии стали сидячими и (или) сменили двустороннюю симметрию на лучевую. Некоторые, как садовая улитка, приобрели диссимметрию, охватившую значительную часть их тела. Некоторые до предела уменьшились в размере, став невидимыми невооруженным глазом, но сохранив почти все системы органов. Некоторые перевернулись, так что спинная и брюшная стороны у них поменялись местами. Некоторые удлинились до нескольких сотен сегментов, а другие, наоборот, вовсе отказались от сегментации. Некоторые выработали сложный наружный скелет, а некоторые — еще более сложный внутренний. Изучением этого невероятного богатства форм животных занимается сравнительная анатомия — интереснейшая наука, но погружение в нее, к сожалению, увело бы нас слишком далеко за пределы тем, охваченных этой книгой.
Новые горизонты
Знаменитый американский физик-теоретик Джон Арчибальд Уилер однажды сказал: “Мы живем на острове, окруженном морем нашей неосведомленности. По мере того как растет остров знания, растет и береговая линия соприкосновения с неведомым”.
Конечно, Уилер был прав. Любой научный прорыв решает и закрывает те или иные вопросы, которые раньше были актуальными (и могли мучить исследователей столетиями). Но одновременно он тут же порождает новые вопросы и ставит новые задачи, недоступные прежним поколениям ученых. Причем этих новых вопросов и задач обычно больше, чем прежних. Так в принципе устроена технология познания. Хорошо это или плохо, но мы никогда не дождемся конца науки. Даже если человечество просуществует несколько миллиардов лет и доживет до превращения Солнца в красный гигант, то и тогда любой ученый, скорее всего, найдет для себя не меньше интересных нерешенных вопросов, чем он может найти сейчас. Хотя, разумеется, вопросы эти к тому времени будут другими — невообразимыми для нас.
Достигнутое в начале XXI века установление истинного (в хорошем приближении) эволюционного древа эукариот вписывается в эту тенденцию. Те, кто его установил, после этого и не думали почивать на лаврах. Они оценили достигнутое, увидели, какие новые вопросы оно порождает, и стали искать ответы на эти вопросы, привлекая новые данные (в первую очередь генетические) и организуя работу по принципу последовательных приближений. И эти исследования сейчас вовсю продолжаются. Увы, любая попытка дать их полную сводку превратила бы эту главу в реестр латинских названий, который никто не стал бы читать. В этой области, условно говоря, еще не улеглась пыль. Поэтому сейчас мы ограничимся в основном теми новыми фактами, которые установлены уже более-менее бесспорно и у которых к тому же есть ясный биологический смысл. Ну, и добавим пару поправок, без которых в наше время уж совсем не обойтись.
Первой такой поправкой стало переименование супергруппы растений. Во всемирно известной сводке, которую выпустил в 2005 году большой международный коллектив протистологов, это название было сочтено неудачным по причине его исторически сложившейся двусмысленности. Вместо него предложили название Archaeplastida (“имеющие древние хлоропласты”). Название Plantae в этой версии системы было сохранено только за высшими наземными растениями, то есть за теми, кого иначе называют Embryophyta.
Исчезла ли в результате этих переименований двусмысленность, вопрос спорный. Но так или иначе название “архепластиды” сейчас очень распространено. Смысл его прост: это единственная супергруппа эукариот, где хлоропласты произошли прямо от цианобактерий, а не от каких-нибудь других эукариот со своими собственными хлоропластами внутри.
Есть, впрочем, и такие авторы, которые как ни в чем не бывало продолжают называть архепластид растениями. Право на это у них есть, потому что выбор названий для групп такого ранга не регламентирован никакими строгими правилами. В данном случае это больше вопрос моды, чем правил.
Добавим, что первичные хлоропласты архепластид — особенность чрезвычайно характерная, но все же не уникальная. Раковинная амеба паулинелла, относящаяся к супергруппе ризарий, тоже приобрела первичный хлоропласт, возникший из захваченной цианобактерии, причем сделала это эволюционно очень недавно (скорее всего, порядка 100 миллионов лет назад). Предок архепластид, безусловно, приобрел свои хлоропласты гораздо раньше.
В 2007 году обнаружилась куда более серьезная проблема. Генетические исследования показали, что группа Rhizaria примерно так же близка к группам Alveolata и Stramenopiles, как и эти две группы друг к другу . Это не раз перепроверяли, и результат получался тот же самый. Попросту говоря, ризарии встраиваются внутрь хромальвеолят. Более подробные молекулярные исследования вроде бы показывают, что ризарии генетически ближе к альвеолятам, чем к страменопилам. Есть, однако, и результаты, противоречащие этому. В любом случае тут мы имеем компактный “куст” из трех ветвей. Этот “куст” стали называть SAR (Stramenopiles + Alveolata + Rhizaria).
SAR — единая эволюционная ветвь, к основанию которой примыкают рано обособившиеся, очень древние группы криптомонад и гаптофит. Положение этих групп на эволюционном древе до сих пор неясно, но, во всяком случае, в саму супергруппу SAR их предпочитают не включать. Следует добавить, что одно время криптомонад и гаптофит считали единой эволюционной ветвью, которой дали название Hacrobia, но эта гипотеза так и не стала общепринятой .
Что касается самой группы SAR, то у нее есть и альтернативное название — Harosa.
Главная проблема супергруппы SAR (= Harosa) состоит в том, что ей пока невозможно дать никакую осмысленную общую характеристику. Она выделена исключительно по молекулярно-генетическим данным, и в нее входят организмы, предельно различные по образу жизни и внешнему облику. Разве что кристы митохондрий у них трубчатые, но этот признак никак не является определяющим (он есть, например, у амебозоев, которые не имеют с SAR ничего общего). Как возникла группа SAR? Каким был ее предок? В чем уникальность ее эволюционного пути? Эти вопросы — настоящий вызов для биологов-эволюционистов.
Unikonta и Bikonta
В 2003 году уже знакомый нам Томас Кавалье-Смит, взявшись в очередной раз корректировать свои представления о филогенетическом древе с учетом новых данных, пришел к выводу, что супергруппы опистоконтов и амебозоев связаны тесным родством . По Кавалье-Смиту, амебозои и опистоконты образуют единую самостоятельную эволюционную ветвь. Это подтверждается и сходством жизненных форм (которое здесь налицо), и генетическими исследованиями. Например, только у опистоконтов и амебозоев слиты вместе три гена, кодирующие ферменты синтеза пиримидинов — очень важного для всех живых организмов класса молекул, в который входят цитозин, урацил и тимин (см. главу 7). Этого слияния нет ни у других эукариот, ни у бактерий, ни у архей. По всей вероятности, оно произошло только один раз. Именно по таким уникальным признакам и следует выделять родственные группы.
Эволюционная ветвь, состоящая из опистоконтов и амебозоев, называется Unikonta (одножгутиковые). Связано это с тем, что у них, натурально, один жгутик. По крайней мере, такое состояние в этой группе распространено довольно широко. У других эукариот жгутиков чаще всего два, если не считать случаев, когда один из них редуцирован — как у эвглены, к примеру.
Кавалье-Смит пошел и дальше. Он предположил, что все эукариоты делятся на две эволюционные ветви: Unikonta (одножгутиковые) и Bikonta (двужгутиковые). К униконтам относятся опистоконты и амебозои, а к биконтам — все остальные, включая экскават, хромальвеолят, ризарий и архепластид.
В чем биологический смысл деления эукариот на Unikonta и Bikonta? Кавалье-Смит не был бы Кавалье-Смитом, если бы не попытался ответить и на этот вопрос. Он обратил внимание на то, что все основные группы Unikonta — это исходно обитатели твердых поверхностей (дна или почвы). Они легко переходят в прикрепленное состояние и, даже будучи подвижными, часто питаются, “заякориваясь” ложноножками на субстрате. В общем, их адаптивная зона — преимущественно придонная.
А вот Bikonta перешли от придонного образа жизни к плаванию в толще воды, среди планктона. Они приобрели второй жгутик, повышающий маневренность, и оболочки, основанные на каркасе из микротрубочек (как пелликула альвеолят), которые придают клетке постоянную форму, делая ее обтекаемой. Это позволило биконтам сначала научиться отлично плавать, а потом и захватить фотосинтезирующих симбионтов, ставших хлоропластами (у униконтов их нет). Неудивительно, что жизненные формы биконтов теперь гораздо разнообразнее.
К сожалению, этот изящный сценарий не выдержал проверки фактами. В 2009 году канадские исследователи Эндрю Роджер и Аластер Симпсон (с которыми мы встречались в начале этой главы) провели тщательное сравнение жгутиковых аппаратов разных одноклеточных эукариот и убедительно показали: общий предок всех современных униконтов наверняка имел два жгутика, а не один. В таком случае называть остальных эукариот “биконтами” бессмысленно. К тому же единство эволюционной ветви биконтов не поддерживается большинством генетических исследований. Видимо, от этого таксона надо просто отказаться.
А вот гипотеза о существовании таксона Unikonta проверку выдержала. Молекулярно-генетические данные надежно подтверждают: супергруппы Opisthokonta и Amoebozoa в самом деле образуют единую эволюционную ветвь . Правда, большинство современных исследователей согласны с тем, что старое название этой ветви — одножгутиковые — не отражает исходное для нее состояние признаков. Но во-первых, это всего лишь название, а во-вторых, одножгутиковых существ среди униконтов и вправду очень много. Иногда униконтов даже считают одной супергруппой, а опистоконтов и амебозоев — подразделениями внутри нее (например, Евгений Кунин в книге “Логика случая” именно так описывает систему эукариот). Что ж, с точки зрения филогенетической систематики это по-своему резонно.
Томас Кавалье-Смит, всегда внимательно относящийся к новым фактам, вскоре согласился, что предки как униконтов, так и всех эукариот когда-то имели два жгутика Отсюда следует, что исчезновение второго жгутика у опистоконтов и у многих амебозоев — состояние производное. Тем не менее верным остается то, что адаптивная зона, которую выбрали униконты, — в самом деле придонная, не предполагающая (как правило) освоения водной толщи, зато связанная с поверхностью грунта. Именно поэтому униконты специализировались в основном к жизни в пресных водоемах или в почве. Вот из этой-то группы и вышли миксомицеты, грибы, животные.
В серии относительно недавних работ Кавалье-Смит отказался от деления на Unikonta и Bikonta, зато разделил эукариот на два эволюционных уровня — Eozoa и Neozoa (см. рис. 15.12). Это решение имеет простой биологический смысл. Eozoa — это низшие эукариоты, которых раньше объединяли (а многие авторы объединяют и сейчас) в ветвь экскават. Однако, по мнению Кавалье-Смита, это на самом деле не ветвь, а эволюционный уровень, являющийся исходным для эукариот в целом. К нему принадлежат жгутиконосцы, имеющие два жгутика, — правда, один из них может редуцироваться (как у эвглены), или же, наоборот, их может становиться больше (как у гипермастигин). Кроме того, для примитивных жгутиконосцев-“эозоев” характерна внутренняя оболочка из микротрубочек — тоже как у эвглены. А вот передвижение с помощью ложноножек у них не развито.
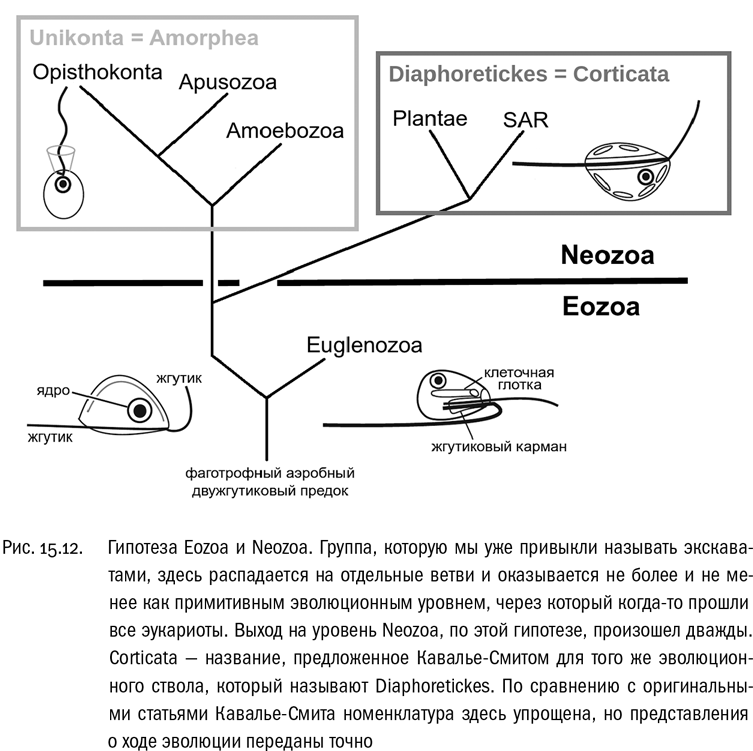
Neozoa, по Кавалье-Смиту, — это высшие эукариоты, у которых возникли два расходящихся магистральных направления эволюции. Одни “неозои” освоили амебоидное движение, которое удобно при донном образе жизни и заодно способствует питанию путем фагоцитоза. Второй жгутик при этом стал не нужен, и во многих эволюционных линиях он исчез. По этому пути пошли униконты. Другие “неозои” сохранили два жгутика, а в придачу приобрели пелликулу и (или) клеточную стенку — структуры, придающие клетке жесткую обтекаемую постоянную форму. Это дало им возможность специализироваться к плаванию в толще воды и к фотосинтезу — последнее, конечно, с помощью захваченных симбионтов. По этому пути пошли SAR и растения, причем последние вообще потеряли способность к фагоцитозу из-за слишком прочной клеточной стенки.
Классификация “Eozoa/Neozoa” не получила большой популярности, потому что она противоречит принципам филогенетической систематики — та категорически запрещает выделение эволюционных уровней в качестве таксонов. Сейчас эта классификация интересна скорее как факт научной мысли, показывающий, что филогенетическая систематика — еще не окончательная истина.
Заодно тут возникает одна частная, но интересная задачка. Мы давно уже знаем, что митохондрии (у всех эукариот) и хлоропласты (у фотосинтезирующих) произошли от симбиотических бактерий. Предками митохондрий были альфа-протеобактерии, а предками первичных хлоропластов — цианобактерии (вторичные хлоропласты, происходящие от поглощенных эукариот с первичными хлоропластами внутри, нас в данном случае не интересуют). И митохондрии, и первичные хлоропласты имеют две мембраны. Логично предположить, что внутренняя мембрана митохондрии или первичного хлоропласта — это плазматическая мембрана захваченной бактерии, а внешняя — мембрана пищеварительной вакуоли ее эукариотного хозяина. Именно это и утверждала классическая теория эндосимбиоза.
Но давайте обратим внимание на то, что предки и митохондрий, и хлоропластов относятся к грамотрицательным бактериям. Это означает, что у них две мембраны — внутренняя и внешняя. После захвата такой бактерии эукариотной клеткой неизбежно образуется конструкция из трех мембран: мембрана пищеварительной вакуоли и две мембраны бактерии. Какая же из них была утрачена? Кавалье-Смит считает, что и в хлоропластах, и в митохондриях наиболее вероятна потеря самой внешней из трех мембран — той, что принадлежала пищеварительной вакуоли хозяина . Ибо потерять ее легче всего: в конце концов, захваченная бактерия просто разорвет эту мембрану, если вздумает внутри нее расти. Ко всему прочему, внешние мембраны митохондрий/хлоропластов и грамотрицательных бактерий совпадают по свойствам: и те и другие устроены как свободное “сито”, проницаемое для максимального числа всевозможных веществ (в отличие как от внутренней мембраны бактерии, так и от внутриклеточных мембран хозяина-эукариота). Так что первая приходящая на ум схема, скорее всего, неверна. И в митохондриях, и в хлоропластах сохранились обе мембраны грамотрицательных бактерий. У одной группы эукариотных водорослей, а именно у глаукофит из супергруппы Plantae, между мембранами хлоропласта сохраняются даже остатки клеточной стенки — пептидогликановой, как бактериям и положено. Кстати, это самое что ни на есть прямое свидетельство происхождения хлоропластов от цианобактерий.
У Томаса Кавалье-Смита есть одна особенность, роднящая его со многими другими хорошими биологами-эволюционистами (начиная, пожалуй, с нашего соотечественника — палеонтолога Владимира Ковалевского). Он прекрасно понимает, что на установлении родственных отношений организмов серьезное изучение эволюции не заканчивается, а начинается. Эволюционная биология не просто наука о том, кто чей предок. В такой же, если не большей мере ее интересует, как и почему произошли те или иные эволюционные события. Например, положение человека на молекулярно-филогенетическом древе сейчас известно абсолютно точно: ближайшая к нему эволюционная ветвь состоит из шимпанзе и бонобо; эти виды разошлись уже после того, как их общий предок отделился от предка человека, и поэтому являются нашими родственниками в строго одинаковой степени. Ну и что? В конце концов, гораздо интереснее узнать, когда и почему человек стал прямоходящим, как при этом менялась его система размножения и социальная структура, для чего увеличился мозг, откуда взялась речь. В общем, выстроить полный эволюционный сценарий, дополняющий тот, который предложил в свое время гениальный Оуэн Лавджой (очень хорошее изложение этого сценария на русском языке можно найти в книге Дональда Джохансона и Мейтленда Иди “Люси”). Разумеется, такие сценарии должны быть проверяемыми, и надо быть готовыми к тому, что иногда они будут опровергаться. Но без них изучать эволюцию было бы просто скучно.
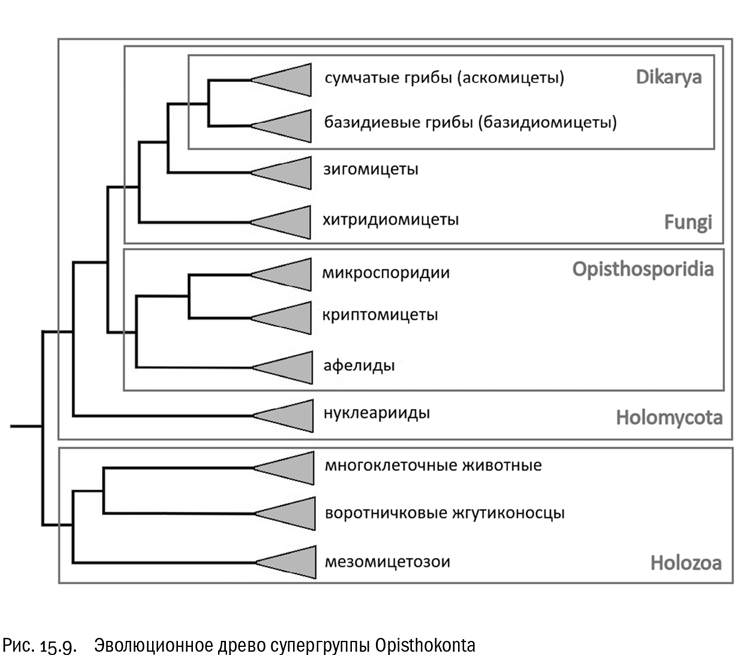
Теперь вернемся к эволюционному древу эукариот и подведем сухие итоги. Исследования последнего десятилетия помимо множества спорных гипотез (которые мы тут не обсуждаем) привели к твердому установлению двух важных фактов. Во-первых, вместо двух равноценных супергрупп — хромальвеолят и ризарий, занимающих на древе соседние места, мы теперь имеем дело с тремя близкими, но самостоятельными ветвями — страменопилов, ризарий и альвеолят, — которые приходится сводить в одну супергруппу: иначе этот пазл не складывается. Во-вторых (и что гораздо более важно), существует эволюционная ветвь Unikonta, состоящая из двух супергрупп — опистоконтов и амебозоев. Подчеркнем, что и опистоконты, и амебозои сами по себе реальны. Просто они вместе образуют ветвь более высокого порядка.
Таким образом, шесть супергрупп эукариот теперь можно свести к четырем: экскаваты, униконты, растения (архепластиды) и SAR.
В капитальной коллективной сводке 2012 года группу Unikonta предложили переименовать в Amorphea. Буквально это значит “бесформенные”: намек на то, что у многих представителей этой группы клетка не имеет постоянной формы. Лучше ли это, чем “одножгутиковые”, — вопрос такой же спорный, как и в случае с архепластидами, но, во всяком случае, в современных публикациях название Amorphea встречается нередко.
А можно ли еще сократить число главных естественных групп эукариот — хотя бы до трех, а лучше до двух? Вероятно, да, но с серьезной оговоркой: чем более древних ветвлений мы коснемся, тем менее надежными будут выводы. В случае с общим эволюционным древом эукариот эта зависимость — очень сильная.
В конце 2000-х появились генетические данные, указывающие, что, скорее всего, эукариоты распадаются на три мегаствола: униконты, экскаваты и растения + SAR . Тогда получается, что растения и SAR образуют единую мегагруппу, в которую входят почти все фотосинтезирующие организмы. Правда, у этой мегагруппы нет уже совсем никаких общих признаков, кроме генетических. Она установлена только благодаря сравнению последовательностей генов, кодирующих несколько сотен разных белков, и математической обработке этих данных. Мегагруппу, состоящую из растений и SAR, предложено называть Diaphoretickes (“разнообразные”), как раз в честь полного отсутствия общих структурных особенностей. Такое вот парадоксальное название. В общем, по этой системе получается, что эукариоты состоят из трех эволюционных мегастволов, названия двух из которых — “бесформенные” и “разнообразные”. Сами эти слова показывают, сколько здесь неясного и насколько это направление исследований открыто для будущего. Не стоит к тому же забывать, что есть и третий мегаствол — экскаваты, уже знакомая нам группа, занимающая на древе особое положение.
Древо и взрыв
Сумма данных, которыми мы сейчас располагаем, не оставляет сомнений, что общий предок современных эукариот уже был полноценной эукариотной клеткой. У него было ядро, эндоплазматическая сеть, аппарат Гольджи, микротрубочки, микрофиламенты, митохондрии и жгутики. Был сложный метаболизм, обеспеченный хорошо знакомым биохимикам набором ферментов. Создается впечатление, что эукариотная клетка во всем ее великолепии появилась в эволюции жизни на Земле сразу в готовом виде. Сразу заметим, что это впечатление может быть и ложным, связанным просто с тем, что переходные формы вымерли, а палеонтологических данных не хватает (по первым шагам эволюции эукариот они вообще отсутствуют). Но так или иначе у общего предка всех современных эукариот полный набор перечисленных атрибутов уже точно был.
Что же произошло дальше с его потомками? Вот здесь структура эволюционного древа вполне может кое-что нам подсказать.
Начнем с того, что есть несколько четко установленных эволюционных стволов эукариот, существование которых бесспорно. Если исходить из общепринятых гипотез и пренебречь некоторыми мелочами, таких стволов можно насчитать девять: опистоконты, амебозои, архепластиды, страменопилы, ризарии, альвеоляты, гаптофиты, криптомонады и экскаваты. Современные генетические данные наводят на мысль, что все эти эволюционные стволы разошлись от некой очень компактной группы предков почти одновременно. И с тех пор они остаются устойчивыми, хотя, конечно, продолжают давать новые формы каждый внутри себя. Иными словами, получается, что в истории эукариот был период чрезвычайно быстрой начальной дивергенции, после которого новые крупные эволюционные ветви уже не возникали. Все расхождения ветвей первого-второго порядка произошли в самом начале эволюции эукариот за такой короткий промежуток времени, что практически наложились друг на друга. Возможно, как раз поэтому выделить главные эволюционные стволы эукариот относительно легко, а вот определить степени их родства между собой очень трудно.
Эту идею нельзя назвать новой. На самом рубеже XX–XXI веков французский биолог Эрве Филипп, поддержанный группой коллег, высказал предположение, что трудность установления родственных связей между крупными ветвями эукариот не является следствием недостатка данных, а отражает реальное событие — “эукариотный Большой взрыв” . Филипп считал, что триггером бурной эволюции эукариот стало увеличение содержания кислорода в атмосфере, вызвавшее симбиоз с предками митохондрий. Появление первого эукариота, обладавшего митохондриями, создало принципиально новую адаптивную зону, которую его потомки начали стремительно осваивать, эволюционируя во все возможные стороны. Вот тогда-то все главные ветви эукариот и возникли. На эволюционном древе соответствующий момент выглядит как мультифуркация — распад одной линии сразу на несколько (см. рис. 15.13). И это не артефакт, связанный с недостатком данных, а отражение реальных особенностей эволюционного процесса в то время и в том месте. Численные прикидки здесь таковы: эукариоты существуют на Земле никак не меньше двух миллиардов лет, но все их основные эволюционные ветви, вероятно, возникли за первые десятки миллионов лет, и уж во всяком случае — не более чем за первые 100 миллионов лет.
В общем, “сейчас представляется, что все известные линии эукариот, дожившие до современности, возникли за промежуток времени, гораздо более короткий, чем время, прошедшее с тех пор. А отсюда следует важный вывод: ни одна современная группа эукариот не может быть значительно примитивнее других”.
Когда стало понятно, что построить филогенетическое древо эукариот все же удается, иные исследователи сочли было это мнение Эрве Филиппа старомодным заблуждением. Но не исключено, что тут они поспешили с выводами.
Во-первых, эволюционный процесс не может идти совершенно одинаково во все времена и во всех таксонах. Существует такое понятие, как “эволюция эволюции”. А возникновение эукариотной клетки — это одно из самых уникальных событий во всей истории жизни на Земле, и нет ничего удивительного, если в его временной окрестности эволюция шла в необычном (с нашей точки зрения) режиме.
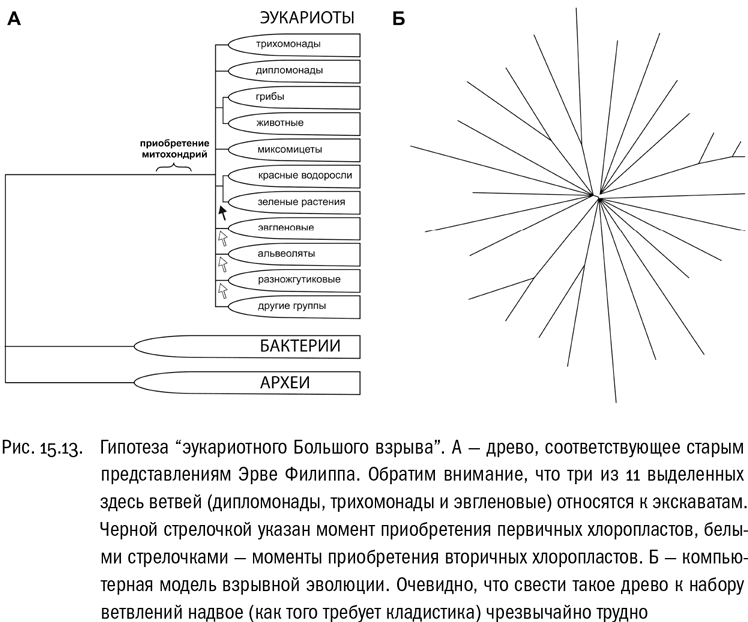
Во-вторых, необычность этого режима все-таки не стоит преувеличивать. Ситуация, когда за короткий промежуток времени один предок дает множество эволюционных ветвей, давно известна биологам и особенно палеонтологам: она называется взрывной эволюцией (explosive evolution) . Хорошо изученные примеры таких событий есть, например, в эволюции млекопитающих. Ничто не мешало подобному “взрыву” произойти и в начале эволюции эукариот. Напротив, к этому были все предпосылки. И тут не надо допускать никаких неизвестных факторов, все вполне укладывается в классическую эволюционную теорию.
Если “эукариотный Большой взрыв” действительно имел место, то нам, как ни странно, не столь уж и важно знать, в каком именно порядке там расходились главные эволюционные стволы. В любом случае первые члены этих стволов почти ничем не отличались друг от друга: при эволюции путем дивергенции иначе и быть не может. А вот что по-настоящему интересно — это кто из них к чему приспосабливался и как они потом делили между собой адаптивные зоны. Обо всем этом мы пока знаем очень мало. Но, видимо, узнаем и больше: установленное эволюционное древо дает для этого неплохую основу.
“Микроцарства”
Настало время сделать очередной шаг вперед. Мы уже успели привыкнуть к тому, что эукариоты делятся на пять-шесть супергрупп. И даже если границы этих супергрупп несколько перекраиваются, общая картина остается более-менее той же.
Проблема в том, что существует несколько небольших групп эукариот, которые ни в какие супергруппы включить не удается (по крайней мере, без серьезных трудностей, натяжек и споров). Мы уже упоминали две такие группы — это гаптофиты и криптомонады. Но есть и другие, менее известные, зато еще более загадочные.
Известный протистолог Ян Павловский метко назвал эти загадочные группы “микроцарствами”. Обсудить их все тут нет никакой возможности, иначе эта глава превратится в справочник. Краткое перечисление тем более не выход: те, кто всерьез заинтересуется темой, легко найдут его в научных статьях, а всех остальных множество мудреных латинских названий скорее отпугнет. Поэтому, чтобы получить какое-то представление о сути дела, но в то же время не запутаться, мы сейчас кратко поговорим всего о трех живых организмах, явно относящихся к “микроцарствам”. В двух случаях это будут отдельные роды, а в третьем — группа из нескольких родов: бревиата, коллодиктион и апузомонады (см. рис. 15.14).
Бревиата — это одноклеточное амебообразное существо со жгутиком, живущее в солоноватых водоемах. Жгутик у бревиаты один и направлен вперед, в отличие от опистоконтов, у которых он тоже один, но направлен назад. При этом второй жгутик у нее, безусловно, когда-то был: электронная микроскопия обнаружила в клетке его остатки . Наряду со жгутиком у бревиаты есть и ложноножки — вполне типичные, как у самой настоящей амебы. Положение бревиаты на эволюционном древе остается спорным. Достаточно правдоподобно выглядит версия, что это самый примитивный современный представитель супергруппы амебозоев. Тогда получается, что эволюционная ветвь, которую образует бревиата, — сестринская по отношению ко всем остальным амебозоям, вместе взятым. Но иногда бревиату вообще не включают в супергруппу амебозоев, оставляя за ее пределами. И возможно, это разумнее, учитывая, что есть и такие результаты, которые больше сближают бревиату с опистоконтами. В любом случае получается, что бревиата примыкает к самому основанию какой-то из этих двух смежных ветвей. А что, если в лице бревиаты перед нами и вовсе третья ветвь, имеющая тот же ранг, что опистоконты с амебозоями, и возникшая одновременно с ними? Такая гипотеза труднопроверяема (и потому непопулярна), но с точки зрения эволюционной теории в ней нет ничего невероятного.
Интересно, что бревиата предпочитает жить в воде с очень низким содержанием кислорода. Поэтому у нее нет митохондрий, хотя есть митохондриальные гены, успевшие мигрировать в ядро. Учитывая, насколько бревиата примитивна (то есть насколько древней эволюционной ветвью она является), вполне можно было бы предположить, что митохондрий у нее никогда и не было. К сожалению, это опровергается твердо установленными генетическими фактами, против которых не пойдешь. Природа далеко не всегда так экономична, как от нее этого ждут люди. Бревиата — в самом деле очень древнее существо, но митохондрии она тем не менее успела когда-то приобрести, а потом утратить.
Апузомонады — жгутиконосцы, которые живут в морских донных отложениях, на дне пресных водоемов, а часто и в почве: капельно-жидкой воды для них там вполне достаточно. Это отличный пример жизненной стратегии, типичной для “одножгутиковых” в классическом понимании Кавалье-Смита. Жгутиков у апузомонад тем не менее два. Они не несут никаких волосков, но направлены в разные стороны: один — вперед, а другой загнут и тянется вдоль тела клетки (поэтому под микроскопом его бывает плохо видно). Жгутиконосец-апузомонада скользит по грунту, работая жгутиками, а на “брюшной” — то есть обращенной к субстрату — стороне он образует ложноножки, которыми захватывает бактерий. Группа эта относительно немногочисленная, но в то же время широко распространенная. Благодаря особенностям своего образа жизни апузомонады легко преодолевают экологические барьеры как между пресными водами и морем, так и между любыми водоемами и почвой. Поэтому весьма вероятно, что именно апузомонады (или какие-то похожие на них существа) первыми среди эукариот вышли на сушу.
Положение этой группы на древе долго было неопределенным. Скорее всего, апузомонады все же примыкают к опистоконтам . Однако ни в группу Holomycota, ни в группу Holozoa ветвь апузомонад не входит — она отошла от ствола опистоконтов раньше, чем эти группы разделились.
Кроме того, в последние несколько лет появились молекулярные данные, позволяющие объединить бревиат, апузомонад и опистоконтов в единую группу, которая получила название Obazoa. Это название образовано от сокращений: O — Opisthokonta, B — Breviatea, A — Apusomonadida. Если это подтвердится, значит, Unikonta (они же Amorphea) распадаются на ветви Amoebozoa и Obazoa. Такая версия мегасистемы уже вошла в некоторые современные руководства.
Интересен вопрос о кристах митохондрий. У апузомонад кристы трубчатые, как у амебозоев. Но у другой сходной с ними группы жгутиконосцев — анкиромонад — кристы пластинчатые, как у опистоконтов. Палеонтологи прекрасно знают, что признаки, обычно характеризующие крупные эволюционные стволы, могут быть очень изменчивыми на ранних этапах эволюции, когда эти стволы еще не разошлись или разошлись едва-едва. Вот такой случай наблюдается и тут.
И наконец, коллодиктион. Мы уже вскользь упоминали это существо в главе 10, но сейчас есть несомненный повод вспомнить о нем еще раз. Коллодиктион был открыт еще во времена Чарльза Дарвина. Однако понимать, насколько особое место он занимает в живой природе, биологи стали только к началу XXI века . Итак, коллодиктион — это довольно крупный пресноводный жгутиконосец. У него четыре одинаковых жгутика, направленных вперед. Кроме того, у него есть ложноножки, причем широкие, лопастевидные, как у амебозоев. На “брюшной” стороне, обычно обращенной к грунту, у коллодиктиона находится ротовая бороздка, армированная со стороны цитоплазмы корешками из микротрубочек. Этот признак мы уже встречали у экскават, которые именно ему и обязаны своим названием. Кристы митохондрий у коллодиктиона трубчатые. По образу жизни он — хищник, причем склонный питаться довольно крупными объектами. Чаще всего это эукариотные зеленые водоросли, иногда одноклеточные, а иногда и колониальные — вплоть до восьмиклеточных колоний (см. рис. 10.5, на котором это зафиксировано). Причем проглоченные водоросли коллодиктион переваривает настолько медленно, что они, по-видимому, успевают у него поработать в качестве хлоропластов — прекрасная модель ранних этапов симбиотической эволюции.
Что же это за существо такое? Здесь нас ожидает сюрприз — впрочем, не такой уж и большой, если учесть все то, что мы к данному моменту уже знаем. Молекулярные исследования показывают, что коллодиктиона нельзя без натяжек включить ни в одну супергруппу эукариот . Это особая эволюционная ветвь — ранний выброс “эукариотного Большого взрыва”. Бревиату, апузомонад и некоторые другие группы тоже можно назвать такими “ранними выбросами”, но коллодиктион, возможно, самый древний из них.
Задумаемся. Очень похоже, что ветвей эукариот, соответствующих по рангу традиционным супергруппам, на самом деле куда больше, чем пять или шесть (см. рис. 15.15). Просто далеко не все из них достигли большого разнообразия. И те, кто его не достиг, статуса супергрупп не удостоились. Хотя объективно, по положению на древе, они вполне могут быть равноправны таковым (как, например, бревиаты равноправны амебозоям).
Коллодиктион — организм, настолько близкий к общему предку всех современных эукариот, насколько это вообще возможно. Его единственный явно производный признак — четыре жгутика. По всей видимости, у общего предка всех современных эукариот жгутиков было два. Но мы знаем, что увеличение числа жгутиков много раз происходило в эволюции экскават (к которым коллодиктион относительно близок), и он просто оказался еще одной группой, независимо от других пошедшей по этому пути. Скорее всего, в его случае это связано с тем, что коллодиктион за свою долгую историю успел специализироваться на питании сверхкрупной добычей — другими эукариотами. Это вызвало увеличение размера, а оно потребовало дополнительных органов движения. И неудивительно, что это относительно недавняя специализация: ведь во времена самых первых эукариот никаких других эукариот, которыми можно было бы питаться, еще просто не существовало в природе.
Состояние всех остальных признаков у коллодиктиона примерно такое, какого и можно было бы ожидать от древнейшего эукариота. Но история со жгутиками здесь очень показательна. Каким бы древним коллодиктион ни был, он, прежде всего, является реальным живым существом, а вовсе не ожившей схемой. А жить — значит специализироваться.
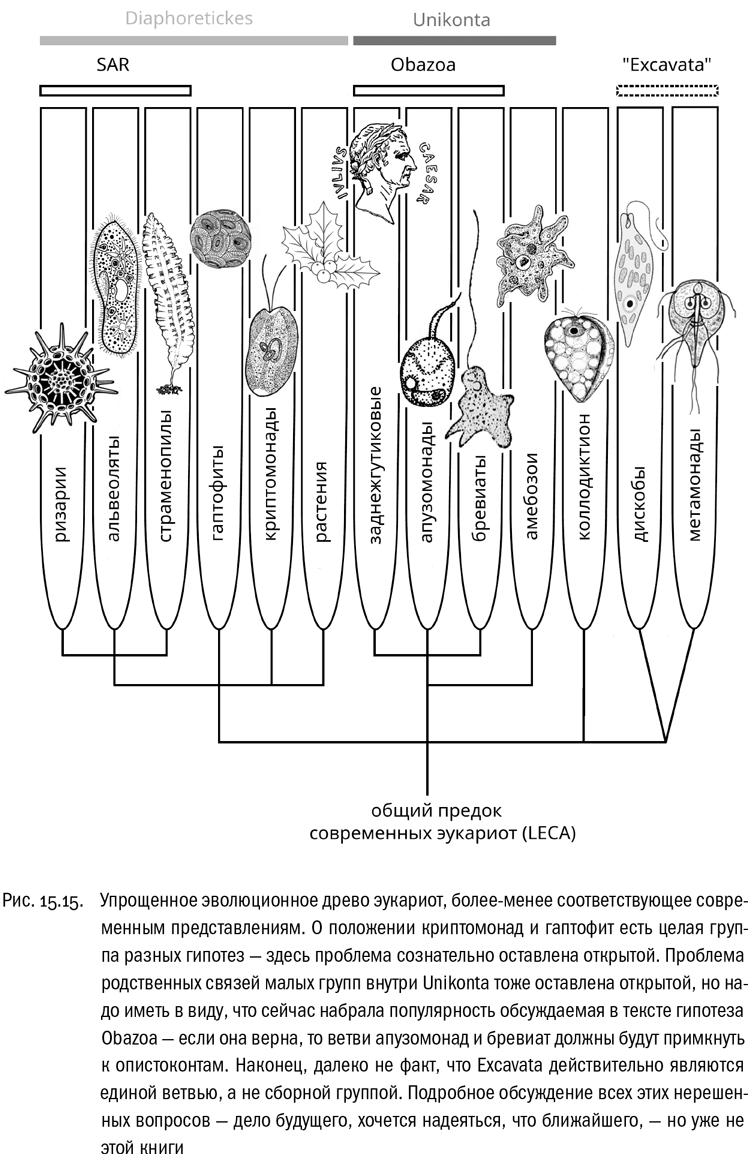
Ну а теперь — торжественный момент. Зная все, что нам сейчас известно, мы можем легко представить себе реального общего предка всех современных эукариот и достаточно подробно описать его. Безусловно, это был жгутиконосец. Одновременно со жгутиками он, скорее всего, имел ложноножки — примерно того же типа, что у обыкновенной амебы, но менее развитые и используемые только для захвата пищи, а не для передвижения. Жгутиков у него было два (без всяких волосков на них). Кристы митохондрий, скорее всего, были трубчатыми, с меньшей вероятностью — пластинчатыми, но в любом случае их форма легко менялась от вида к виду: на том эволюционном этапе этот признак еще не устоялся. Образ жизни был придонным: жгутиконосец передвигался в основном у поверхности грунта, лишь изредка поднимаясь в толщу воды, поэтому в его клетке дифференцировались “спинная” и “брюшная” стороны. На “брюшной” стороне, скорее всего, была продольная ротовая бороздка — та самая, которая сохранилась в супергруппе экскават и благодаря которой эта супергруппа получила свое название. И конечно, наш предок был хищником — в той мере, в какой в тогдашнем мире вообще было возможно хоть какое-то хищничество. Он питался всеми, кого мог поймать, и занимал самую вершину пищевой пирамиды.
Обычному человеку может быть нелегко совместить с наглядными представлениями о мире тот факт, что его личный прямой предок — ничуть не менее прямой, чем дед или прадед, — когда-то, пару миллиардов лет назад, был одноклеточным жгутиконосцем, похожим на коллодиктиона. Но, судя по всему, это чистая правда. Как сказал герой известной пьесы Григория Горина: “Это гораздо больше, чем факт. Так оно и было на самом деле”.
Многоклеточность
Вся биология когда-то началась с изучения многоклеточных организмов. Это совершенно естественно. “Люди, львы, орлы и куропатки, рогатые олени, гуси, пауки, молчаливые рыбы, обитавшие в воде, морские звезды и те, которых нельзя было видеть глазом”, — легко заметить, что в этом классическом списке, порожденном трепетным подсознанием героини Чехова, девять наименований из десяти относятся к многоклеточным существам, которые были объектами науки еще во времена Аристотеля. Изучение природы всегда начинается с очевидного, а к менее очевидному переходит лишь постепенно.
Давайте еще раз взглянем на знакомое нам теперь эволюционное древо эукариот. Перед нами великое множество эволюционных ветвей, большинство из которых — это бросается в глаза — состоит из одних только одноклеточных организмов, как правило, и в самом деле невидимых простым глазом: их удалось открыть только после изобретения микроскопа. Несколько меньше — ветвей, в которых есть колониальные формы, и совсем уж немного тех, где возникает многоклеточность в самом полном смысле этого слова. Например, многоклеточные животные — это всего лишь одна из нескольких сотен (без преувеличения) эволюционных ветвей аналогичного ранга.
Что же интересного в таком редком явлении, как многоклеточность? Какой, собственно, вклад в картину мира она дает? И в конце концов, достойна ли она вообще глубокого фундаментального интереса? Может быть, значение исследований многоклеточных организмов в основном прикладное — для медицины, ветеринарии, сельского хозяйства и тому подобных отраслей?
Обойти эти вопросы было бы интеллектуальной нечестностью. К счастью, ответ на них достаточно очевиден. Любой многоклеточный организм предоставляет нам для изучения множество интереснейших явлений, которых ни у каких одноклеточных просто-напросто не существует. Это программирование зародышевого развития, в ходе которого тысячи клеток должны располагаться в пространстве заданным образом, обновляя это расположение с каждым очередным клеточным делением; работа генных сетей, тончайшим образом регулирующих экспрессию разных генов в разных клетках; клеточная дифференцировка; гормональная сигнализация; регуляция обмена веществ на уровне целого организма; и наконец, работа сложнейшего из всех известных нам природных объектов — человеческого мозга. Неудивительно, что огромная доля биологов занималась (и, вероятно, всегда будет заниматься) именно этими вопросами. Можно смело утверждать, что даже если бы во Вселенной существовал всего лишь один вид многоклеточных организмов, притом не имеющий никакого практического значения, то и тогда он заслужил бы огромное внимание биологов, к какой бы форме жизни сами эти биологи ни относились. Им было бы ясно, что они столкнулись с принципиально новым способом организации живой материи, который грех не попытаться изучить как следует. В похожей ситуации оказались, например, герои лемовского “Соляриса”. Организм, который они обнаружили, был единственным в своем роде и не имел для Земли никакого практического значения, но тем не менее он сразу вызвал к жизни целую область науки — соляристику. Лем прекрасно понимал психологию ученых, в реальности все так и было бы.
Кроме того, появление многоклеточных организмов самым серьезным образом изменило облик Земли как планеты. Они повлияли и на состав атмосферы, и на ландшафты, и на глобальные круговороты разных химических элементов (в первую очередь углерода), и на климат. Без многоклеточной жизни Земля выглядела бы совершенно иначе.
Итак, многоклеточность — это важно. Увы, компактно обсудить ее у нас сейчас не получится. Дело в том, что способы реализации многоклеточности в разных группах организмов слишком различны, чтобы о них можно было хотя бы в самом общем приближении рассказать как о едином целом. Этим занимаются специальные науки — зоология, ботаника, альгология (наука о водорослях) и микология (наука о грибах), — погрузиться в которые в этой книге нет никакой возможности: как сказал классик, никто не обнимет необъятного. К счастью, и о многоклеточных животных, и о многоклеточных растениях, и даже о грибах источников информации для желающих хватает (о грибах, например, есть книга “Занимательная микология”, которую написал один из самых блестящих профессоров биофака МГУ — Юрий Таричанович Дьяков). А мы тут поневоле ограничимся общими моментами.
Для начала представим себе простенькую зеленую водоросль с двумя жгутиками — вроде хламидомонады, той самой, которая описана в школьном учебнике ботаники. Она, безусловно, одноклеточная. Но если она поделится, а потом еще раз поделится и образовавшиеся клетки почему-либо не разойдутся, то мы получим колонию из четырех клеток — а значит, уже многоклеточное существо, по крайней мере сугубо номинально. И такие четырехклеточные колонии реально существуют (зеленая водоросль гониум, например). Если делений будет больше, то образуются колонии из 8, 16, 32, 64 клеток: все эти примеры тоже реальны. При этом экспериментально установлено, что грань между колонией, где все клетки одинаковы и обладают равными возможностями, и организмом, где они начинают необратимо специализироваться, в данном случае проходит на уровне 32-клеточной водоросли эудорины. “Любая из 32 клеток Eudorina elegans также способна дать новую колонию. Однако у 32-клеточных Eudorina illinoisensis четыре передние клетки более мелкие, они уже не могут участвовать в размножении, представляя дифференцированную и не способную к инициальной роли часть колониальной особи”. Именно здесь мы видим рубеж между колонией, члены которой потенциально могут разойтись и жить независимо, и организмом, откуда обратного хода нет. Но означает ли это, что эудорина находится на одной ступени организации с метасеквойей, пшеницей, слоном или человеком? А если нет, то в чем между ними разница?
Известный американский протистолог Джон Корлисс решил, что разбираться в этом нужно, основываясь на понятии ткани . По общепринятому определению, ткань — это система клеток и межклеточного вещества, объединенных происхождением, строением и выполняемыми функциями. Так вот, Корлисс предложил считать, что протистом называется любой эукариотный организм, имеющий не более одной ткани. Тогда получается, что к протистам относятся все одноклеточные... и некоторые многоклеточные тоже. Например, любая нитчатая водоросль, хоть она и многоклеточная, должна считаться протистом, потому что никакого разнообразия тканей у нее нет.
В то же время определение Корлисса четко указывает границу, за которой отнести живое существо к протистам уже при всем желании нельзя. Например, у многоклеточного протиста не может быть никакой сосудистой системы, потому что она по самой своей природе требует наличия хотя бы двух тканей: той, из которой состоят сосуды, и той, которая их окружает и через них снабжается.
Организмы, имеющие не менее двух полноценных тканей, Корлисс вполне логично назвал многотканевыми. Например, все многоклеточные животные бесспорно попадают в эту категорию. У любого уважающего себя животного, даже если у него нет нервной системы и мышц, есть хотя бы две ткани: эпителиальная (покровная) и соединительная, занимающая внутренний объем и богатая межклеточным веществом. И той и другой необходимы межклеточные контакты, о которых мы упоминали в конце главы 10. Для эпителиев особенно важны плотные контакты и десмосомы, а для соединительной ткани — фокальные контакты, которые позволяют клеткам закрепляться на волокнах межклеточного вещества.
По мнению Корлисса, на уровень многотканевых организмов вышли всего две эволюционные ветви: многоклеточные животные (Metazoa) и высшие растения (Embryophyta). Все остальные до полноценной многотканевости, грубо говоря, недотянули. Ближе всех к ней подошли некоторые водоросли — бурые, красные, а также харовые, которые близки к наземным растениям и раньше считались их прямыми предками. Но Корлисс все равно считает их всех протистами, несмотря на то что длина некоторых бурых водорослей достигает 60 метров. У грибов многотканевость сомнительна, и по классификации Корлисса грибы — тоже протисты. Они потенциально способны выйти на многотканевый уровень, но им это просто не нужно. Есть одна группа грибов — лабульбениомицеты, паразиты насекомых, — в теле которых все-таки возникла практически настоящая тканевая организация. Клетки делятся там в разных направлениях и образуют целую трехмерную структуру, хоть и мелкую (доли миллиметра). Но в мире грибов это исключение, а не правило.
На грани перехода к многотканевости находятся некоторые слизевики, входящие в супергруппу амебозоев. Тут есть примеры, когда в плодовом теле возникает настоящая покровная ткань, причем устроенная поразительно похоже на эпителий животных . Более того, некоторые мембранные белки, обеспечивающие межклеточные контакты, у слизевиков те же самые, что и у животных. Взглянув на эволюционное древо, мы видим, что слизевики и животные могли унаследовать эти белки только от общего предка всей мегагруппы Unikonta. Значит, молекулярная “машинерия”, позволяющая в принципе создать многоклеточность, у этого предка — очень древнего! — уже была на месте.
В высшей степени замечательно, что собственные слизевики нашлись не только в супергруппе амебозоев (куда входит большая их часть), но и в группе опистоконтов. Это фонтикула — с виду вполне типичный слизевик, у которого питающаяся стадия состоит из множества одиночных амеб, а для образования спор служит плодовое тело . Но типичные слизевики принадлежат к амебозоям. А вот фонтикула, как показала молекулярная генетика, вовсе к ним не относится. Она относится к супергруппе опистоконтов и занимает там место в основании “грибного” эволюционного ствола Holomycota — рядом с амебами-нуклеариидами, на которых ее амебоидная стадия, собственно, и похожа . Это означает, что в супергруппе опистоконтов (в которую, напомним, входят не только грибы, но и животные) была самостоятельно, хотя и на общей с амебозоями молекулярной основе, выработана жизненная форма слизевика. И как ни странно, это вполне ожидаемо. Слизевики независимо возникали в разных группах эукариот не меньше семи раз. Единственная из традиционных супергрупп эукариот, в которой нет собственных слизевиков, — это растения (архепластиды). Во всех остальных супергруппах они есть. Как видим, жизненная форма слизевиков почему-то оказалась среди эукариот очень “популярной”. И мы не можем исключить, что через такую жизненную форму когда-то прошли предки многоклеточных животных.
История со слизевиками показывает нам еще одну важную вещь. Многоклеточный и даже многотканевый организм вовсе не обязан быть таким на протяжении всей своей жизни. У слизевиков и, если уж на то пошло, у грибов многоклеточные плодовые тела образуются только на коротком отрезке жизненного цикла. К тому же они никак не участвуют в питании, а служат только для размножения. В общем-то эти организмы вполне могли бы обойтись и без плодовых тел, если бы им не требовалось разбрасывать споры (и многие грибы действительно обходятся).
Теперь посмотрим под тем же углом зрения на эволюцию растений. Мы уже говорили о гипотезе, согласно которой предками наземных растений были пластинчатые зеленые водоросли, напоминающие современные колеохетовые (см. рис. 15.16А). Если эта гипотеза верна, то тело первого наземного растения должно было состоять из двух частей: фотосинтезирующая зеленая пластинка, погруженная в воду либо растущая на влажном грунте, и образующийся в период размножения воздушный стволик с коробочкой для разбрасывания спор (см. рис. 15.16Б–Д). Вот только этот стволик на первых порах и обладал многотканевой организацией. Основной части растения — зеленой пластинке, погруженной во влажную среду, — она была не нужна.
Не просматривается ли тут закономерность? Насколько мы можем судить, и у слизевиков, и у грибов, и у растений многотканевая организация начинает формироваться на временных непитающихся стадиях. И только потом (у растений) многотканевость распространяется почти на весь жизненный цикл, порождая хорошо нам знакомые жизненные формы трав, деревьев, кустарников. Заметим, что, например, у слизевиков ничего подобного не произошло. Правда, у некоторых из них бывают стадии ползающих многоядерных амеб, но там ни на какую многотканевость нет и намека. Плодовые же тела служат для размножения, и ни для чего больше.
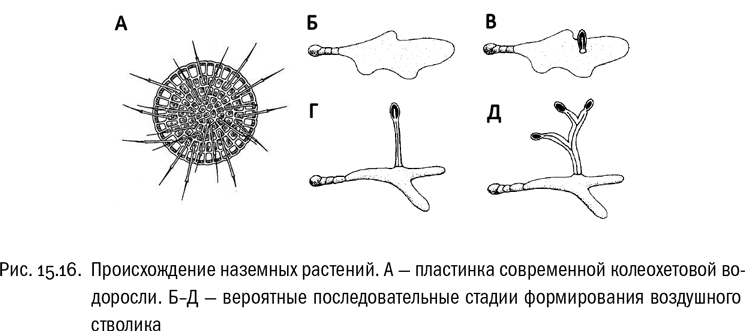
Еще один фрагмент мозаики мы получим, обратившись к достижениям современной молекулярной биологии. Есть целая группа генов и белков, которые сначала считались уникальными для многоклеточных животных, но затем были обнаружены у воротничковых жгутиконосцев и мезомицетозоев — иначе говоря, у одноклеточных и колониальных представителей эволюционной ветви “животных в широком смысле”, которая называется Holozoa. Про один из таких генов мы уже говорили — это ген Brachyury, участвующий у позвоночных в развитии осевых структур. Но есть и другие: например, гены, кодирующие мембранные белки, обычно связанные у животных с межклеточными плотными контактами (теми самыми, что важны для эпителиальной ткани). Или тирозинкиназы — ферменты, переносящие фосфат от АТФ на остаток аминокислоты тирозина в белке-мишени (о том, что это за процесс, см. главу 7). Эти ферменты служат важными внутриклеточными посредниками в ходе сигнальных взаимодействий, которые — опять же — особенно важны для многоклеточных животных. Причем все перечисленные гены распространены как минимум по всей группе Holozoa, а некоторые и шире.
Какой же отсюда следует вывод? По всей вероятности, жизненный цикл общего предка всех Holozoa уже был достаточно сложным и включал в себя многоклеточную стадию — не обязательно питающуюся, вполне вероятно, что покоящуюся или расселительную. Именно на этой сугубо временной стадии, скорее всего, и была первоначально “обкатана” многотканевость, которая затем, в одной специализированной эволюционной ветви, распространилась почти на весь жизненный цикл (см. рис. 15.17). И в результате возник один из сложнейших и красивейших объектов в известной нам Вселенной: животный организм, внутреннюю структуру которого изучает сравнительная анатомия.
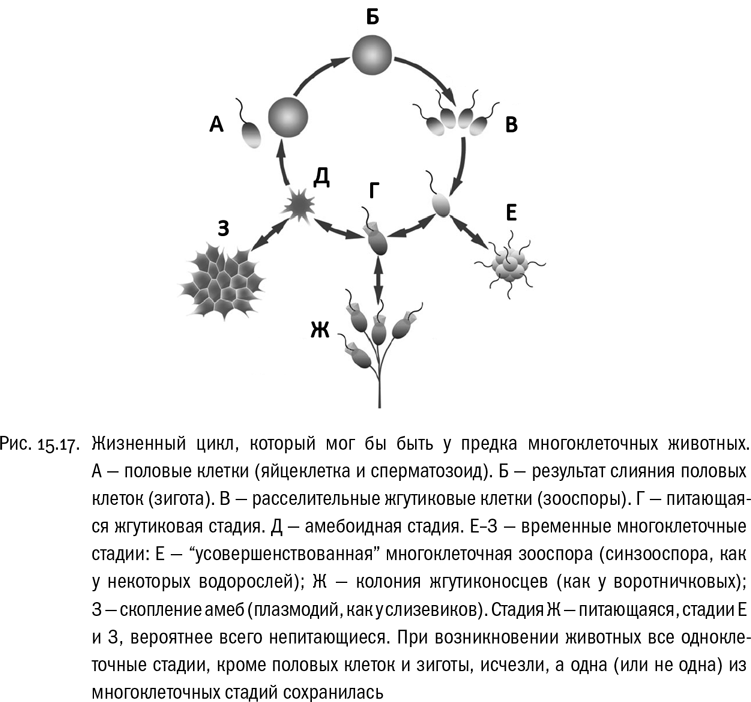
Подвижные в подвижном
Животные — это многоклеточные организмы, образ жизни которых определяется неразрывной связью питания с движением. Любое животное или активно движется в среде, или столь же активно движет среду относительно себя — например, гонит воду бьющимися ресничками, попутно извлекая из нее питательные частицы. Иногда то и другое совмещается. Тогда мы видим фильтраторов, способных к активному движению, среди которых есть наши очень близкие (по меркам эволюционного древа) родственники: например, ланцетник и головастики некоторых лягушек. А вот обратный случай — неподвижное животное в неподвижной среде — почти невозможен. “Почти” — потому что тут, как и обычно в биологии, можно найти исключения: есть крайне специализированные паразиты, которые живут внутри других животных, а сами по образу жизни приближаются к грибам (хотя и у них есть подвижные личинки). Но это — отличный пример исключения, подтверждающего правило. Как сказал один хороший московский профессор зоологии, “во всех тех случаях, когда подвижность теряется, животные теряют свой образ и подобие”[142]. Типичному многоклеточному животному больше, чем любому другому живому существу, подходит девиз корабля капитана Немо: Mobilis in mobile — “Подвижное в подвижном”. Кстати, и свое название этот корабль получил в честь животного — головоногого моллюска наутилуса, свободно плавающего в океанской толще.
Именно с совершенством двигательной системы, скорее всего, связан эволюционный успех одной из самых разнообразных групп животных — типа хордовых (см. рис. 15.18). Этот тип не является самым многочисленным в животном царстве. Есть как минимум два типа, превосходящие его по числу видов: членистоногие и моллюски. Тем не менее разнообразие формы тела и образа жизни у хордовых огромно, а в экологических нишах, требующих крупного размера, они просто вне конкуренции. Почему? Тут стоит обратить внимание на тот самый признак, который дал типу хордовых название. Их “визитная карточка” — хорда, или спинная струна, тянущаяся вдоль большей части тела. Она связана с массивной сегментированной мускулатурой, которая позволяет животному двигаться, волнообразно изгибаясь (хорда при этих изгибах благодаря своей упругости не дает телу потерять форму). Получается очень эффективный двигательный аппарат, который иногда называют миохордальным комплексом. Никаких других уникальных преимуществ, кроме миохордального комплекса, у ранних хордовых, насколько можно судить, не было. Их эволюционный успех вызван необыкновенно удачной конструкцией опорно-двигательной системы, которая, с одной стороны, дала им возможность стать со временем лучшими в мире хищниками, увеличив размер и заняв самую вершину пищевой пирамиды, а с другой — создала потенциал для бурной эволюции на структурном уровне. Выражаясь современным языком, план строения хордовых имеет очень высокую эволюционируемость (evolvability)[143]. Но при этом даже на самых неожиданных эволюционных поворотах он не разрушается, а остается самим собой, сохраняя глубокое единство общей структуры — то, что великий сравнительный анатом XIX века Ричард Оуэн назвал архетипом позвоночных[144]. Сочетание высокой эволюционируемости с фундаментальной устойчивостью плана строения проходит красной нитью сквозь всю эволюцию хордовых животных, начавшуюся свыше 500 миллионов лет назад со скромного, длиной в несколько сантиметров, плавающего фильтратора — “подвижного в подвижном”. Как мы знаем, именно на этом эволюционном пути в конце концов возникли общество и разум.
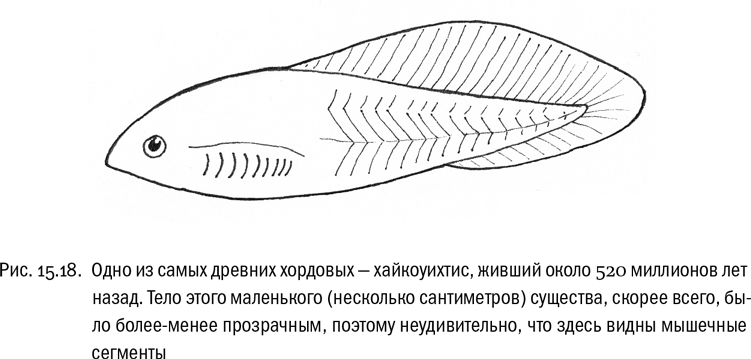
Назад: 14. рождение системы
Дальше: ЧАСТЬ IV ИСТОРИЯ ЖИЗНИ

