Книга: От атомов к древу. Введение в современную науку о жизни
Назад: 9. генетическая информация
Дальше: 11. энергия
10. эукариотная клетка
На общедоступном языке мы можем назвать ядро администратором клетки. Два главных свойства роднят его с более знакомыми нам администраторами: оно стремится плодить себе подобных и чрезвычайно успешно отражает все наши попытки узнать, чем же именно оно занимается. Только попытавшись обойтись без него, мы можем наконец убедиться, что оно действительно работает.Дэниел Мазия (цитируется по сборнику “Физики продолжают шутить”)
А сейчас обсудим одну особую группу живых организмов, которая называется эукариотами. Раньше мы уже встречались с этим названием (см. главы 5 и 8). К эукариотам относятся животные, растения, грибы и многие (но не все) одноклеточные существа — такие, как описанные в традиционных школьных учебниках зоологии амебы, эвглены и инфузории. Две другие главные группы клеточных организмов, кроме эукариот, — это бактерии и археи. Причем молекулярные данные показывают, что эукариоты произошли, скорее всего, от архей, а не от бактерий. Архей и бактерий вместе часто называют прокариотами, но надо иметь в виду, что это не название эволюционной ветви, а сборное понятие, образованное методом исключения: вот, мол, есть эукариоты, а есть все прочие.
Самые древние предполагаемые эукариоты, остатки которых удалось найти палеонтологам, имеют возраст 2,2 миллиарда лет. При этом общепризнанный возраст планеты Земля равен 4,6 миллиарда лет, а возраст самых древних предполагаемых следов жизни составляет, по-видимому, 4,1 миллиарда лет (см. главу 16). Первые живые клетки на Земле, несомненно, были прокариотными. И, судя по датам, они оставались такими в течение двух миллиардов лет. Иначе говоря, получается, что на протяжении половины времени своего существования земная жизнь обходилась безо всяких эукариот. В эту эпоху она была чисто прокариотной, то есть бактериальной и архейной.
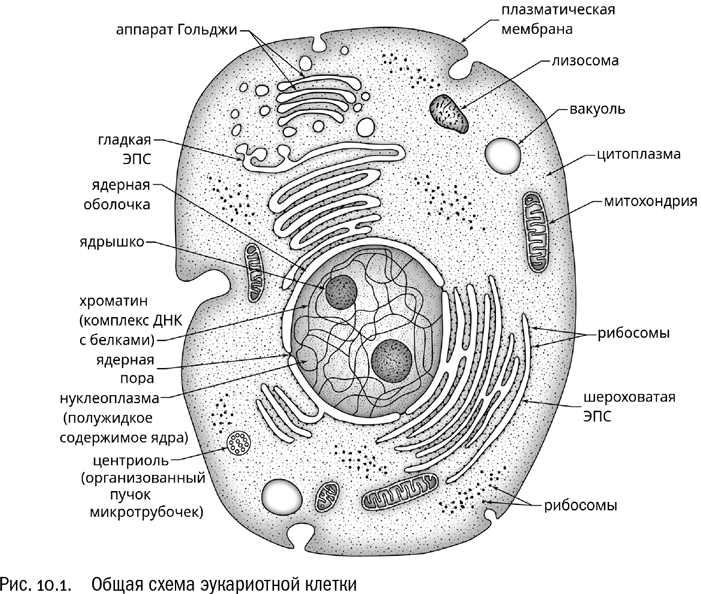
Если кратко просуммировать основные отличия эукариотной клетки от прокариотной, у нас получится примерно следующий список (см. рис. 10.1):
* большой размер клеток: средний одноклеточный эукариот крупнее среднего одноклеточного прокариота примерно в 10 раз по длине и примерно в 1000 раз по объему;* ДНК эукариот всегда заключена в клеточное ядро, окруженное оболочкой из двух мембран;* ДНК эукариот линейна, в отличие от кольцевой ДНК бактерий и архей;* эукариотная клетка пронизана сложной системой внутренних мембранных полостей и пузырьков (эндоплазматическая сеть, аппарат Гольджи, вакуоли);* у эукариот есть система внутриклеточных опорно-двигательных образований, называемая цитоскелетом;* у эукариот широко распространены дополнительные внутриклеточные структуры, окруженные собственными мембранами, — митохондрии (обеспечивают дыхание) и хлоропласты (обеспечивают фотосинтез).
Чтобы познакомиться с эукариотами, мы должны поговорить про эти отличия чуть подробнее.
ЭПС и ядро
Начнем с самого общего. В любой эукариотной клетке впридачу к наружной мембране есть внутренняя система взаимосвязанных мембранных полостей и каналов, которая называется эндоплазматической сетью (ЭПС). Основные функции ЭПС, если описать их буквально парой слов, — синтез и транспорт белков, липидов и некоторых других веществ. Принято различать шероховатую и гладкую ЭПС. На шероховатой ЭПС снаружи сидят рибосомы, а на гладкой — нет. Поскольку единственной функцией рибосом является синтез белка, то и связанная с ними шероховатая ЭПС занимается в первую очередь тем, что накапливает и транспортирует белки. У гладкой ЭПС функции другие, более разнообразные.
Благодаря ЭПС и другим мембранным структурам эукариотная клетка, как принято говорить, очень сильно компартментализована. Это означает, что она разделена на ограниченные мембранами отсеки (компартменты), переход веществ между которыми, как правило, возможен только с помощью специальных транспортных белков. Надо сказать, что это куда более фундаментальная особенность, чем может показаться на первый взгляд. Прокариотную клетку, ни на какие отсеки не разделенную, можно в грубом приближении рассматривать как единый мешок с раствором (пусть и вязким). А вот для эукариотной клетки такое приближение не работает в принципе. Внутри прокариотной клетки ДНК, белки и другие молекулы перемещаются из конца в конец путем простой физической диффузии, как в любом сосуде с обычным водным раствором. В эукариотной клетке такое совершенно невозможно: там практически все молекулы вынуждены добираться от места синтеза до “места работы” с помощью сложных транспортных систем, обеспечивающих направленные переходы из отсека в отсек, часто еще и с затратой энергии. Все это означает, что у эукариотной клетки гораздо выше внутренняя упорядоченность, или, говоря научным языком, ниже энтропия. Справедливо замечено, что с точки зрения физики более фундаментальное отличие трудно придумать.
Специализированное разрастание ЭПС, заключающее в себе почти всю ДНК клетки, называется клеточным ядром (см. рис. 10.2). Пространство между двумя мембранами ядра непосредственно продолжается во внутреннюю полость ЭПС. Во избежание путаницы надо сразу пояснить, что ДНК находится не в этой полости, а снаружи от нее, между цистернами ЭПС, сомкнутыми в шар. Этот шар и есть ядро (хотя бывают и ядра куда более сложной формы). Часть содержимого клетки, находящуюся за пределами ядра, принято называть цитоплазмой.
Транспорт из ядра в цитоплазму и обратно происходит через ядерные поры — отверстия, пронизывающие обе ядерные мембраны и окруженные довольно сложными белковыми комплексами, которые избирательно пропускают разные молекулы в ту или другую сторону. Белки, которые образуют ядерные поры, называются нуклеопоринами.
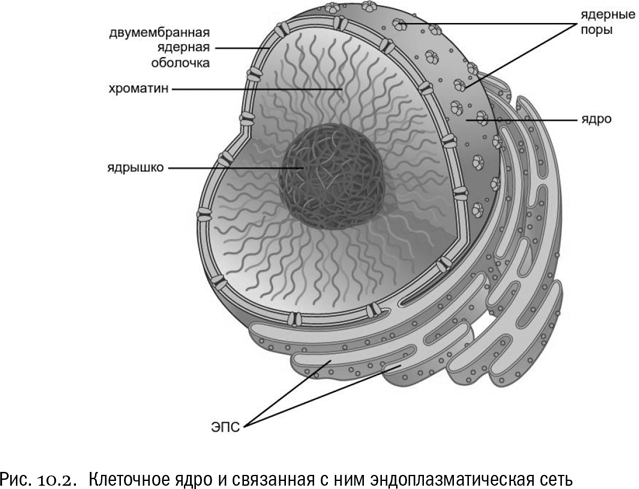
Основное содержимое ядра — ДНК, связанная со специальными белками. Важнейшие, хотя и далеко не единственные, из этих белков — гистоны, на которые ядерная ДНК эукариот буквально наматывается. В гистонах всегда много лизина и аргинина — положительно заряженных аминокислот, заряды которых облегчают взаимодействие с ДНК (ведь ее молекула заряжена, как мы знаем, отрицательно). Кроме того, гистоны часто подвергаются дополнительным химическим модификациям, которые бывают важны для регуляции экспрессии генов.
Совершенно особая часть ядра — ядрышко, в котором экспрессируются рибосомные гены и формируются составные части рибосом. В целые рибосомы они собираются уже в цитоплазме, потому что целая рибосома не пройдет сквозь ядерную пору. По-латыни ядро называется nucleus, а ядрышко — nucleolus.
Клеточное ядро — важнейший признак эукариот. Собственно говоря, “эукариоты” буквально и значит “обладающие настоящим ядром”. У всех современных эукариот ядро полностью сформировано, так что никаких переходных состояний между эукариотами и прокариотами мы не видим. Происхождение клеточного ядра — на сегодняшний день одна из самых больших загадок во всей эволюционной биологии.
Вакуоли и цитоскелет
Любая находящаяся внутри клетки замкнутая полость с жидкостью, ограниченная мембраной, называется вакуолью. Строго говоря, вся эндоплазматическая сеть — это не что иное, как сложная система ветвящихся, сливающихся, переходящих друг в друга вакуолей. Но в любой эукариотной клетке есть еще и множество самостоятельных мелких вакуолей, функции которых очень разнообразны. Они могут служить для переваривания захваченных клеткой пищевых объектов (пищеварительная вакуоль), для собирания и удаления лишней воды (сократительная вакуоль) или просто для транспорта разных веществ.
Кроме того, в эукариотной клетке обычно есть специализированная транспортная система, которая называется аппаратом Гольджи (по имени итальянского биолога Камилло Гольджи). Это стопка крупных плоских вакуолей, от краев которых могут отшнуровываться подвижные маленькие вакуоли. Разные молекулы “упаковываются” в эти маленькие вакуоли и отправляются в них по назначению.
Как правило, аппарат Гольджи развит тем лучше, чем больше веществ клетка должна транспортировать внутри себя и (или) выделять наружу. Например, очень мощный аппарат Гольджи имеют нервные клетки, что и понятно: они должны постоянно и в довольно большом количестве выделять сигнальные вещества — нейротрансмиттеры.
Сложные формы клеток эукариот, а заодно и все виды их внутриклеточной подвижности возможны только благодаря системе внутриклеточных опорных структур, которая называется цитоскелетом. Ни у бактерий, ни у архей цитоскелета нет (по крайней мере, полноценно развитого, насколько мы сейчас знаем).
Цитоскелет эукариот делится на микротрубочки (состоящие из белка тубулина), микрофиламенты (состоящие из белка актина) и промежуточные филаменты (они имеют разнообразный состав). Очень важным свойством цитоскелета является динамическая нестабильность. Это означает, что ни микротрубочки, ни микрофиламенты, ни промежуточные филаменты не образуются раз и навсегда — они могут постоянно собираться и разбираться. Эти процессы требуют затрат энергии, но именно благодаря им эукариотные клетки бывают крупными, сложными по форме, а очень часто и подвижными.
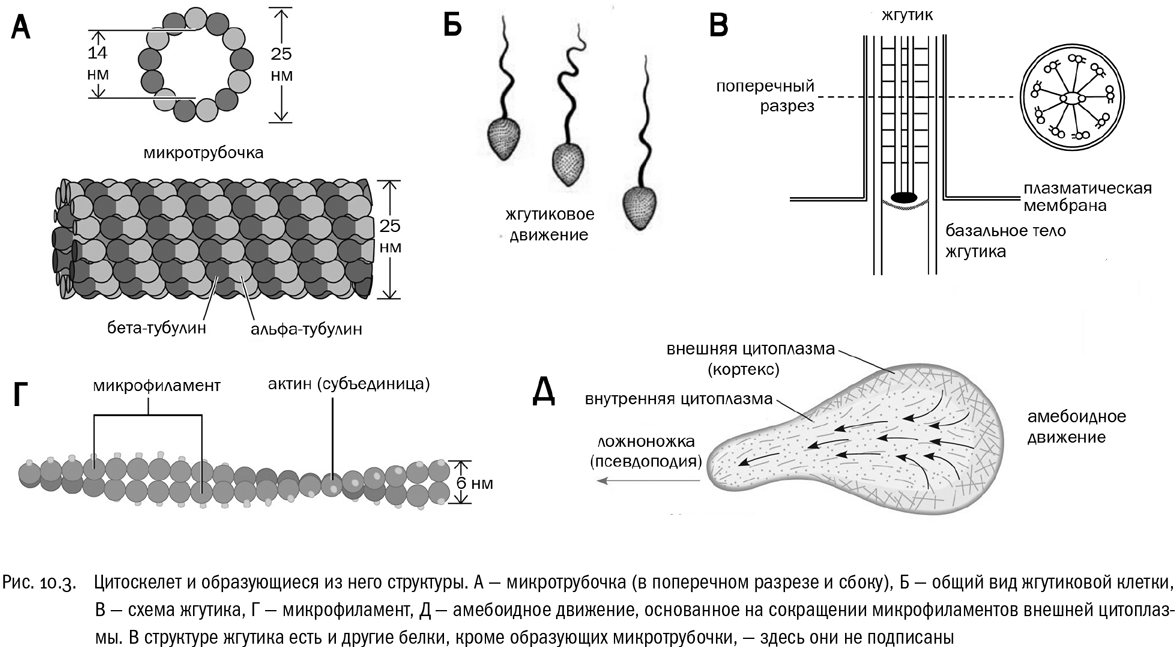
Взглянув на элементы цитоскелета по отдельности, мы обнаружим, что они достаточно сильно отличаются друг от друга. Микротрубочки — это полые цилиндры, собранные из множества молекул белка тубулина (см. рис. 10.3А). Надо заметить, что молекулы тубулина бывают нескольких разных типов, например альфа- и бета-тубулин. Они кодируются отдельными генами и входят в микротрубочки в строго определенных соотношениях. Сборка и разборка микротрубочек питается энергией за счет расщепления ГТФ.
Именно микротрубочки, как правило, придают эукариотным клеткам постоянную форму. Но это далеко не единственная их задача. Микротрубочки служат еще и своего рода “рельсами”, по которым ездят по клетке мелкие транспортные вакуоли. Разумеется, это всегда происходит при участии дополнительных двигательных белков, которые расщепляют АТФ и движутся вдоль микротрубочек за счет высвобожденной энергии. Еще более важно участие микротрубочек в клеточном делении.
И наконец, микротрубочки образуют основу специализированных двигательных структур — жгутиков и ресничек (см. рис. 10.3Б). Эти структуры есть у одноклеточных эукариот, ставших широко известными благодаря попаданию в школьный учебник зоологии, — эвглены (жгутики) и инфузории-туфельки (реснички). Внутри каждого жгутика и каждой реснички проходит пучок микротрубочек, организованный строго определенным образом. Обычно это две самостоятельные микротрубочки в центре и девять сдвоенных микротрубочек по краю, так называемая схема 9+2 (см. рис. 10.3В). С помощью специальных моторных белков эта система микротрубочек совершает ритмичные движения, изгибая жгутик и помогая в итоге двигаться всей клетке.
Микрофиламенты состоят не из тубулина, а из актина, и имеют форму не трубочек, а гораздо более тонких нитей (см. рис. 10.3Г). Они меньше подходят для поддержания постоянной формы клетки, но зато (в отличие от микротрубочек) могут сокращаться. Делают они это с помощью белка миозина, расщепляющего АТФ. Если микротрубочки — это в полном смысле слова внутриклеточный скелет (цитоскелет), то микрофиламенты скорее своего рода внутриклеточная мускулатура (“цитомускулатура”). Правда, надо всегда иметь в виду, что в отличие от скелета и мышц многоклеточного животного тубулиновый цитоскелет и актиновая “цитомускулатура” вовсе не являются образованиями, данными клетке раз и навсегда. Они могут легко собираться и разбираться. Образно говоря, трудно представить животное с настолько пластичным скелетом, чтобы его перестройки на протяжении жизни превращали это животное, например, из крысы в морскую звезду или из змеи в осьминога. А вот цитоскелет такие возможности клетке дает.
Большинство типов клеточной подвижности, свойственных эукариотам, связаны не с микротрубочками, а с микрофиламентами. Утверждение, что микрофиламенты способны сокращаться, строго говоря, неточно. На самом деле длина каждого отдельного микрофиламента при сокращении не меняется. Вместо этого микрофиламенты скользят относительно друг друга по молекулам моторного белка миозина таким образом, что в результате вся клетка или ее часть меняет форму — например, становится короче. Мышечное сокращение устроено именно так. Любая мышечная клетка буквально набита микрофиламентами, которые движутся согласованно. Но, кроме того, микрофиламенты ответственны и за многие другие виды движения клеток. Например, взаимные перемещения микрофиламентов могут вызывать амебоидное движение, при котором клетка постоянно меняет форму, выпуская и втягивая временно существующие выросты — ложноножки (см. рис. 10.3Д). Так ползает амеба и многие похожие на нее одноклеточные существа. А заодно тем же способом ползают внутри организма некоторые клетки тел животных, в том числе и человеческого тела.
Фагоцитоз
Актин, связанный с миозином, кратко называют актино-миозиновым комплексом. Это основа большинства типов движения, на которые способны эукариотные клетки. Самый внешне впечатляющий из них, конечно, мышечное сокращение. Но оно есть только у многоклеточных животных (и то не у всех). Это далеко не самый древний и не самый распространенный способ движения из тех, что имеют подобную природу.
Например, только благодаря актино-миозиновому комплексу возможен такой вид активности, как фагоцитоз — захват клеткой посторонней частицы с ее изоляцией внутри пищеварительной вакуоли и последующим перевариванием (см. рис. 10.4). Открывший фагоцитоз в XIX веке Илья Ильич Мечников сразу понял, что это важнейший механизм иммунитета. Действительно, в организме многоклеточного животного, в том числе и в человеческом, обычно есть подвижные клетки с ложноножками, готовые буквально проглотить и переварить любого подходящего по размеру незваного гостя, например какую-нибудь бактерию. Однако, кроме того, фагоцитоз очень часто используется просто для питания. Одноклеточным эукариотам он вообще только для этого и нужен. Но и многоклеточные животные нередко по старинке усваивают пищу с помощью фагоцитоза. (У человека, правда, фагоцитоз в пищеварении не участвует.)
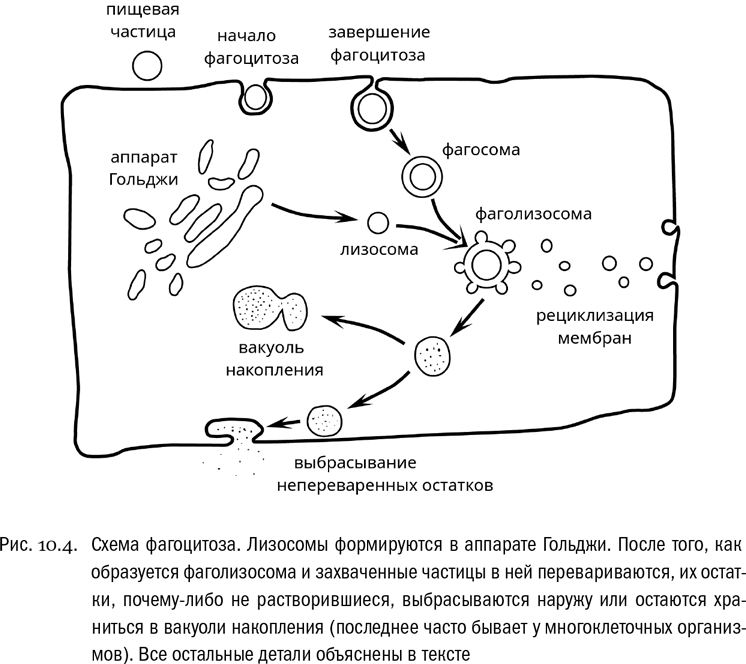
Вакуоль, образующаяся непосредственно в результате фагоцитоза и заключающая в себе проглоченные объекты, называется фагосомой. Она транспортируется цитоскелетом до места слияния с другой вакуолью — лизосомой, содержащей пищеварительные ферменты, которые могут расщепить слишком крупные молекулы до мономеров (например, белки до аминокислот). После слияния вакуолей образуется фаголизосома, в которой захваченные частицы и перевариваются.
При фагоцитозе клетка может потерять за счет включения в фагосому довольно большую часть наружной мембраны, особенно если она “проглотила” что-то очень крупное. Но это ненадолго: по мере того как проглоченная пища переваривается, от фаголизосомы отделяются маленькие вакуольки, которые перемещаются к наружной мембране и встраиваются в нее, чтобы вернуть мембранные липиды обратно. Этот процесс называется рециклизацией мембран. У прокариот такого активного круговорота мембран нет, а вот для эукариотных клеток он очень характерен.
Именно путем фагоцитоза питается, например, попавшая в школьный учебник зоологии обыкновенная амеба. Клетки, активно занимающиеся фагоцитозом, есть и в человеческом теле. Это разновидности белых кровяных клеток (лейкоцитов), которые называются нейтрофилами и моноцитами, а также подвижные клетки рыхлой соединительной ткани — макрофаги. Последние есть во всех органах человека, и в них могут превращаться моноциты, выползающие из кровеносных сосудов. Макрофаги постоянно ползают по организму амебоидным способом, меняя форму клетки и образуя с помощью своего актино-миозинового комплекса временные выросты, то есть ложноножки. А в наружной мембране макрофага при этом сидят специальные белки-рецепторы, которыми он “проверяет” все встречные объекты. Любые клетки, на внешней поверхности которых нет некоторого определенного набора белков и липидов, макрофаг тут же заглатывает. Это довольно эффективный способ борьбы, например, с вредными бактериями. Разумеется, работа макрофага строго регулируется в зависимости от того, какие химические сигналы он получает из внешней среды. В его наружной мембране есть рецепторы, срабатывание которых запускает фагоцитарную активность, а есть и такие, срабатывание которых, наоборот, тормозит ее.
Правда, бактерии тоже сопротивляются клеткам, которые пытаются их съесть. И иногда это сопротивление бывает крайне изощренным. Например, возбудитель проказы — грамположительная бактерия, которую в честь первооткрывателя называют палочкой Хансена, — научился жить аж внутри макрофагов. Как мы знаем из главы 6, грамположительные бактерии отличаются от грамотрицательных отсутствием второй клеточной мембраны, так что их самой наружной оболочкой является толстая клеточная стенка. У палочки Хансена клеточная стенка в основном полисахаридная. Но, кроме того, в ней содержится много очень необычных жирных кислот с длинными разветвленными цепями и трехуглеродными замкнутыми кольцами в них (по этим кольцам формулу такой кислоты сразу можно опознать). Они называются миколовыми кислотами. Молекулы миколовых кислот делают поверхность бактерии чрезвычайно гидрофобной и устойчивой к внешним воздействиям — в том числе и к действию лизосомных пищеварительных ферментов, которые, по идее, должны расщеплять все что угодно. В каком-то смысле миколовые кислоты и есть главная тайна возбудителя проказы. Именно благодаря им палочки Хансена, поглощенные макрофагами, с большим удовольствием живут и размножаются прямо в цитоплазме этих клеток. К счастью, у большинства бактерий таких невероятных биохимических способностей все-таки нет.
Фагоцитоз есть далеко не у всех эукариот. Во-первых, многим из них хватает других способов питания, а во-вторых (и это еще важнее), фагоцитоз несовместим с наличием клеточной стенки. Сквозь клеточную стенку, которая находится снаружи от мембраны и часто бывает довольно толстой, никого проглотить невозможно. А если от клеточной стенки отказаться, это сразу же делает клетку и менее прочной, и менее защищенной. Как раз по этим причинам нет фагоцитоза, например, у зеленых растений — им-то клеточная стенка уж точно нужнее. Но у самых древних эукариот он, скорее всего, был.
Рождение чудовища
Все эти истории рассказываются вот к чему. Мы теперь знаем, что фагоцитоз возможен только при наличии актино-миозинового комплекса. Это чисто эукариотное свойство. У бактерий и архей актино-миозинового комплекса нет, поэтому к фагоцитозу они неспособны. Хищные прокариоты (очень, надо заметить, немногочисленные) всегда меньше своих жертв и являются на самом-то деле скорее паразитами. Такой бактериальный “хищник” вбуравливается в толщу клеточной стенки более крупной бактерии, питается находящимися там белками, липидами и полисахаридами и там же размножается. А вот проглотить свою жертву целиком никакая бактерия не может в принципе.
Это означает, что до появления эукариот — то есть в первые два миллиарда лет истории жизни — на Земле не было настоящих хищников. Самыми крупными и сложными живыми объектами тех времен были строматолиты, подушкообразные многослойные колонии прокариотных синезеленых водорослей (они же цианобактерии). Наработанная ими биомасса в основном просто захоранивалась в морских осадках: поедать и разлагать ее, возвращая в итоге в атмосферу в виде углекислого газа, было некому. Цепи питания были очень короткими и простыми.
Появление хищника, способного к фагоцитозу, мгновенно изменило ситуацию. Адекватным ответом жертвы на давление такого хищника был естественный отбор в сторону увеличения размера, чтобы хищник не смог ее проглотить. Но и хищники стали увеличивать размеры в ответ. Возникла положительная обратная связь, и началась эволюционная гонка вооружений. (Это не метафора, подобные процессы описываются теми же дифференциальными уравнениями, что и гонка вооружений в экономике.) Клетки постепенно становились все более крупными и сложными. И наконец, когда увеличивать размер отдельной клетки стало уже некуда, в ход пошел последний довод эукариот: многоклеточность.
Этот сценарий навел некоторых ученых на мысль, что именно появление цитоскелета, и особенно актино-миозинового комплекса, было тем самым начальным звеном, за которым последовало возникновение всего остального набора эукариотных признаков. Эндоплазматическая сеть, вакуоли, аппарат Гольджи, ядро — все это появилось несколько позже, чтобы структурировать внутренний объем разросшейся громадной клетки. В начале же был цитоскелет, и только он.
Эта гипотеза хороша тем, что поддается проверке фактами. Есть ли основания считать, что цитоскелет действительно появился раньше других признаков эукариот? Да, есть. В последние несколько лет было обнаружено, что белки, очень близкие к актину и тубулину, имеются у некоторых архей. Правда, фагоцитоз эти археи еще не освоили: они умеют создавать в лучшем случае выросты клеток, но не впячивания, а последнее для фагоцитоза необходимо. Однако то, что белки цитоскелета действительно очень древние, теперь ясно.
Итак, если мы посмотрим на предка эукариот с точки зрения бактерии, то увидим невероятного монстра. Лишенный клеточной стенки и постоянной формы тела, он компенсирует это гигантским размером, а главное — направо и налево пожирает своих соседей по сообществу, как Безликий Бог из знаменитого аниме “Унесенные призраками”. В общем, это поистине прокариотный ночной кошмар, nightmare.
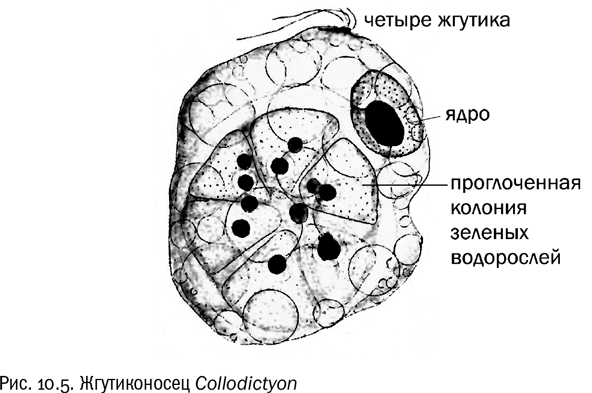
Хорошей иллюстрацией этого эволюционного сценария служит один из самых древних и архаичных эукариот, доживших до современности, — пресноводный жгутиконосец Collodictyon (см. рис. 10.5). Это крупный одноклеточный хищник, обладающий как жгутиками, так и ложноножками. Он склонен питаться очень крупными объектами, вплоть, например, до колоний зеленых водорослей, состоящих из восьми клеток. Причем зеленые водоросли тоже эукариоты, так что клетки у них довольно большие, но коллодиктиона и это не останавливает. Весьма вероятно, что первые эукариоты были на него во многом похожи (см. главу 15). Трудно даже представить, какую революцию в мирном прокариотном сообществе должно было произвести появление такого суперхищника.
Тут мы видим сюжет, интересный с точки зрения общих закономерностей биологической эволюции. На первый взгляд кажется очевидным, что хищник всегда зависит от жертвы (например, его численность напрямую зависит от численности жертв: чем больше зайцев, тем больше рысей). И это действительно так — в экологическом масштабе времени, то есть на коротких временных отрезках, где эволюцией самих хищников и жертв можно пренебречь. А вот в эволюционном масштабе времени картина сплошь и рядом меняется на обратную: хищник начинает диктовать жертве условия, задавая направление ее эволюции. Это называется эффектом опережающей специализации хищника. Появление первых эукариот, пожалуй, самый яркий пример этого эффекта во всей истории Земли (во всяком случае, до появления нового сверхуниверсального суперхищника — человека). При этом эукариоты моментально начали охотиться и друг на друга. Некоторых из них на время спасла многоклеточность, дававшая крупный размер и тем самым какую-никакую защиту. Судя по всему, эукариотным хищникам было гораздо сложнее стать многоклеточными, чем эукариотам с растительным типом питания (см. главы 15, 16). Отсюда — множество эволюционных ветвей многоклеточных водорослей (красные, бурые, золотистые, желтозеленые, зеленые, харовые). Многоклеточность послужила для них своего рода “убежищем” — хоть и временным, но время это было долгим. Эффект опережающей специализации хищника вообще часто работает ускорителем эволюции, порождающим новые формы живых организмов.
Симбиогенез
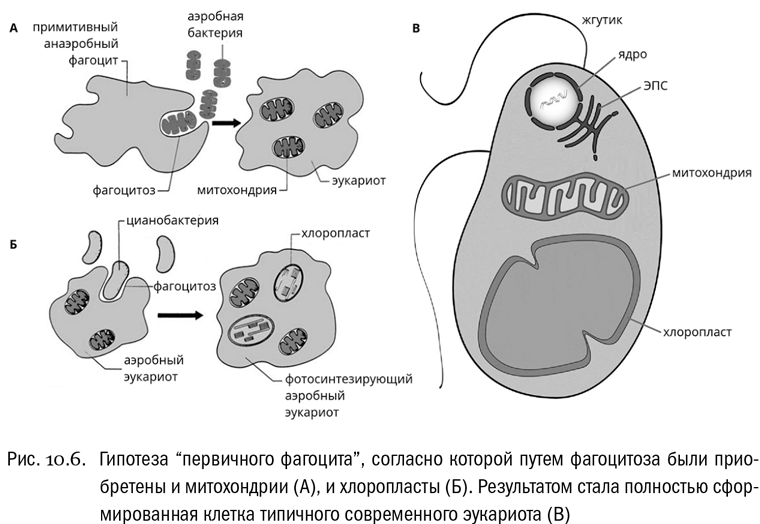
Обычно считается, что именно путем фагоцитоза были приобретены такие клеточные структуры эукариот, как митохондрии и хлоропласты. Напомним, что митохондрии осуществляют дыхание, то есть распад глюкозы до углекислого газа и воды с получением химической энергии, запасаемой в виде АТФ. Митохондрии есть у подавляющего большинства эукариот. Что касается хлоропластов, их задача — фотосинтез, то есть синтез глюкозы из углекислого газа и воды с использованием энергии света. Хлоропласты есть у многих эукариот, но далеко не у всех. Тем, у кого они есть, для питания в принципе достаточно углекислого газа и света, без дополнительных источников энергии они могут обойтись.
Сейчас является общепринятой следующая гипотеза. И митохондрии, и хлоропласты — это потомки бактерий, которые когда-то были поглощены эукариотной клеткой и остались в ней жить, снабжая своего хозяина полезными продуктами обмена веществ. Митохондрии в первую очередь поставляют хозяйской клетке АТФ, хлоропласты — глюкозу. Если это верно, значит, мы видим тут типичный пример симбиоза, то есть взаимовыгодного сожительства разных организмов. Симбиоз, один из участников которого живет внутри другого, называется эндосимбиозом. А теорию, согласно которой многие важные признаки эукариот возникли в результате симбиоза, принято называть симбиогенетической или просто теорией симбиогенеза.
В пользу симбиогенетической теории есть ряд очень веских доводов. Во-первых, и митохондрии, и хлоропласты могут сохранять собственную ДНК. Причем эта ДНК, как правило, кольцевая, как у бактерий. Во-вторых, и хлоропласты, и особенно митохондрии могут сохранять собственный аппарат синтеза белка со своими рибосомами, причем это (опять же) рибосомы бактериального типа: они меньше эукариотных и несколько отличаются от них по строению. Таким образом получается, что в эукариотной клетке действуют одновременно два типа рибосом: обычные эукариотные рибосомы в цитоплазме и бактериальные рибосомы в хлоропластах и митохондриях. В-третьих, ни митохондрии, ни хлоропласты никогда не образуются de novo, то есть с нуля. Они самостоятельно размножаются делением, примерно как обычные свободноживущие бактерии. В-четвертых, оболочка митохондрий и хлоропластов всегда состоит минимум из двух мембран (а не из одной, как у простых вакуолей). Логично предположить, что внешняя мембрана митохондрии или хлоропласта — это мембрана пищеварительной вакуоли хозяина, а внутренняя — мембрана самой проглоченной бактерии.
Идея симбиогенеза “витала в воздухе” еще с конца XIX века, мелькая в самых разных научных работах. Но первым, кто сформулировал ее совершенно четко и во всеуслышание, был русский ботаник Константин Сергеевич Мережковский (старший брат знаменитого писателя Дмитрия Мережковского). Его первая статья на эту тему вышла в 1905 году. Мережковский был уверен, что хлоропласты высших растений и большинства водорослей — это не результат дифференциации цитоплазмы материнской клетки, а внешние по отношению к ней симбионты, имеющие независимое происхождение. Причем предками хлоропластов он считал прокариотные синезеленые водоросли (цианобактерии), как это потом и подтвердилось.
Из биологов первой половины XX века теорией симбиогенеза не заинтересовался почти никто. Слишком уж она была смелой и (по тогдашним меркам) необычной. Например, американский биолог Эдмунд Уилсон, автор самого авторитетного в первой половине XX века учебника по биологии клетки, высказывался о теории симбиогенеза как о чистой фантастике. “Дальнейший полет воображения Мережковского, — писал Уилсон, — приводит его к допущению, что зеленые растения возникли от симбиоза бесцветных ядросодержащих клеток и мельчайших синезеленых водорослей, из которых последние дали начало хлоропластам. Без сомнения, многим такие спекуляции могут показаться слишком фантастичными, чтобы о них можно было упоминать теперь в приличном обществе биологов”.
Второе рождение теории симбиогенеза было связано с работами выдающейся женщины — американского биолога Линн Маргулис. Она настаивала на симбиотическом происхождении не только хлоропластов, но и митохондрий, и некоторых других клеточных структур, например жгутиков. Последнее, как мы сейчас понимаем, неверно. Но зато симбиотическое происхождение митохондрий и хлоропластов с тех пор подтверждено очень надежно. На данный момент это скорее твердо установленный факт, чем гипотеза. Союз предков митохондрий и хлоропластов с эукариотами, безусловно, настоящий симбиоз. Бывшие бактерии снабжают своего хозяина полезными веществами, получая взамен стабильные условия и защиту от других хищников.
Известно даже, какие именно это были бактерии. Митохондрии, скорее всего, произошли от пурпурных альфа-протеобактерий, а хлоропласты — от уже упоминавшихся цианобактерий, они же синезеленые водоросли. В этом пункте Мережковский оказался совершенно прав.
Митохондрии, несомненно, появились гораздо раньше, чем хлоропласты. Во многих эволюционных ветвях эукариот (в том числе и в нашей) никаких хлоропластов просто нет и, судя по всему, никогда не было. С митохондриями ситуация совершенно иная. На данный момент можно твердо сказать, что практически у всех современных эукариот есть или собственно митохондрии, или их маленькие остатки, сохранившие часть биохимических функций, или — на худой конец — отдельные митохондриальные гены, успевшие когда-то мигрировать в ядро и встроиться в ядерный геном. Это означает, что у общего предка всех современных эукариот митохондрии уже были. Их отсутствие (по крайней мере, у современных эукариот — не забудем про эту оговорку) бывает только вторичным, то есть результатом утраты.
Приобретение митохондрий — единственный доступный для эукариот способ быть аэробными, то есть получать энергию с помощью кислорода. Предки современных эукариот стали аэробными только после того, как приобрели митохондрии. Если же какой-нибудь эукариот теряет митохондрии, то он, наоборот, становится анаэробным, приспособленным жить в бескислородной среде. В этом случае кислород бесполезен, кислородное дыхание невозможно, а глюкоза распадается только до молочной кислоты или этилового спирта, что дает в несколько раз меньше полезной энергии.
Между тем потеря митохондрий может случаться у эукариот очень легко (по эволюционным меркам, конечно). Обычно это происходит при переходе к жизни в бескислородных условиях, где митохондрии бесполезны. Причем такое бывает не только с одноклеточными эукариотами, но и с многоклеточными. Бескислородные условия, в которых митохондрии не нужны, — это чаще всего или чей-нибудь кишечник, или некоторые типы морских и пресноводных донных отложений. Например, митохондрий нет у некоторых представителей лорицифер, очень своеобразных мелких морских червей с выворачивающимся хоботом и пластинчатым панцирем. И вероятно, они не одни такие. В случае с этими многоклеточными животными уж точно нет никаких сомнений, что митохондрии у них когда-то были, а исчезли вторично, за ненадобностью.
Отдельный интересный сюжет, связанный с митохондриями, касается вариабельности генетического кода. Когда генетический код только расшифровали, считалось, что он абсолютно единый и всеобщий для всех живых существ Земли — от вируса до слона. Это почти так и есть, но не совсем. Митохондриальные геномы как раз одно из тех мест, где в генетическом коде встречаются отклонения, причем достаточно многочисленные и разные у разных эукариот. Например, генетический код митохондрий человека отличается от “базового” генетического кода, действующего у того же человека в ядерном геноме, значениями четырех кодонов. Кодон УГА, в “базовом” коде являющийся стоп-кодоном, в митохондриях соответствует триптофану, а кодон АУА, в “базовом” коде соответствующий изолейцину, здесь кодирует метионин — и так далее. Для клетки в целом это некритично: поскольку митохондрия имеет собственный аппарат синтеза белка, то на работу ядерных генов изменения в ее генетическом коде никак не влияют. Просто получается, что в одной и той же клетке одновременно функционируют два разных генетических кода — ядерный и митохондриальный.
Скорее всего, изменения в генетическом коде митохондрий стали накапливаться уже после того, как митохондрии окончательно стали внутриклеточными симбионтами. Со временем большая часть митохондриальных генов перешла в ядро, а оставшийся митохондриальный геном стал таким маленьким, что сбой его работы (который неизбежен при любом изменении генетического кода) перестал быть непременно смертоносным. Надо учитывать, что митохондриальные гены, мигрировавшие в ядро и вошедшие в состав ядерного генома, экспрессируются там, где они теперь находятся, то есть в ядре. А их белковые продукты (даже если эти белки нужны только для митохондрий) синтезируются на обычных эукариотных рибосомах, находящихся в цитоплазме, и внутрь митохондрий попадают уже оттуда, с помощью специальных переносчиков.
После того как в митохондриальном генетическом коде накопилось некоторое количество изменений, митохондриальные белки стало уже невозможно правильно синтезировать на эукариотных рибосомах. Вот тогда-то миграция митохондриальных генов в ядро и прекратилась. Если бы не это, митохондрии, вероятно, в конце концов совсем потеряли бы свой генетический аппарат, и тогда разгадать их симбиотическое происхождение было бы гораздо труднее.
И наконец, еще один важнейший факт состоит в том, что — как мы сейчас знаем — эукариоты эволюционно близки не к бактериям, а к археям. Это выяснилось еще тогда, когда археи как отдельная эволюционная ветвь были только открыты (см. главу 5). Скорее всего, эукариоты прямо от них и произошли. А вот предки митохондрий и хлоропластов, наоборот, не археи, а бактерии. Тут произошло довольно редкое событие, а именно слияние совершенно разных эволюционных ветвей.
“Первичный фагоцит”
Факты, которые мы теперь знаем, легко встраиваются в следующий довольно простой эволюционный сценарий (см. рис. 10.6). Когда-то в далекой древности (вероятно, два с лишним миллиарда лет назад) некая бесцветная архея приобрела сложный цитоскелет, включающий актино-миозиновый комплекс. Клеточную стенку она, наоборот, потеряла. Получился “первичный фагоцит” — подвижная клетка, способная активно ползать, выпуская ложноножки, и фагоцитировать (то есть заглатывать целиком) любые более мелкие объекты, в том числе и клетки бактерий. Это оказалось очень выгодным способом питания. Но такое питание непременно предполагает, что хищник должен быть крупнее жертвы. И действительно, современные эукариотные одноклеточные хищники — амебы и жгутиконосцы — по линейным размерам примерно в десять раз превосходят бактерии, которыми они питаются.
Оговорка про линейные размеры тут очень важна. Если увеличить линейный размер клетки в десять раз (обычный порядок разницы между эукариотами и прокариотами), то — по законам геометрии — ее объем увеличится примерно в 1000 раз, с пропорциональным ростом нагрузки на все внутриклеточные системы синтеза и транспорта. Вот для того, чтобы структурировать огромный внутренний объем, у предков эукариот и образовалась система внутренних мембран, включающая ЭПС, аппарат Гольджи, множество вакуолей и ядро. Возникший в результате организм представлял собой что-то вроде крупной бесцветной хищной амебы. Ничего особенного, подобные эукариоты есть и сейчас, разве что они нередко обладают еще и жгутиками в придачу к ложноножкам.
Питание путем фагоцитоза предопределило возможность появления у эукариот внутриклеточных сожителей, то есть, по-научному говоря, симбионтов. Для прокариот, лишенных способности заглатывать кого бы то ни было, приобретение симбионтов таким способом невозможно. А вот амебоидный эукариот может легко превратить некоторые захваченные клетки из пищи в симбионтов, если “примет решение” не переваривать их, а предоставить им жить в хозяйской клетке, делясь с хозяином полезными продуктами обмена веществ. В современных научных статьях такой захват эукариотами чужих клеток часто буквально называют порабощением (enslavement). Именно путем такого “порабощения” эукариотная клетка и приобрела сначала митохондрии, а потом хлоропласты.
К сожалению, на самом деле все не так просто. Познание природы имеет свои закономерности: как правило, чем больше мы узнаем, тем больше возникает новых вопросов, причем таких, которые на предыдущем уровне знания просто не могли бы никому прийти в голову. И проблема происхождения эукариот может послужить отличной иллюстрацией того, что так бывает очень часто. Мы сейчас увидим, что есть как минимум два соображения, заново затуманивающих, казалось бы, предельно ясно описанную картинку.
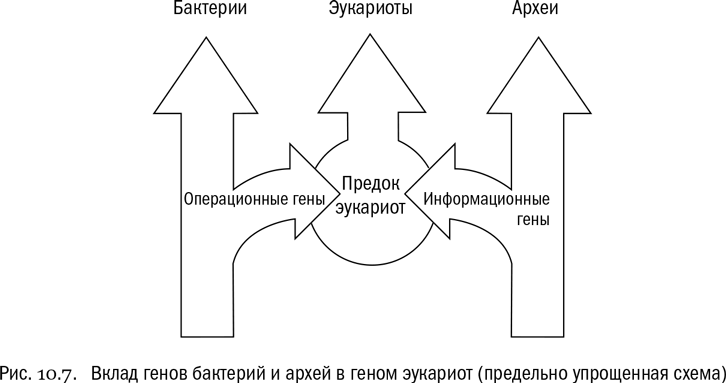
Прежде всего, в ядерных геномах эукариот обнаружено много генов бактериального происхождения, причем — что самое интересное — полученных не только от предков митохондрий и хлоропластов, но и от других групп бактерий, которые внутри эукариотных клеток никогда не жили. Напомним еще раз, что центральная часть эукариотной клетки, включающая ядро, несомненно, произошла от клетки какой-то древней археи (а не бактерии). Между тем подробный анализ эукариотных белков и генов, которые их кодируют, показывает следующее. Непосредственно от архей эукариоты унаследовали в первую очередь белки, работа которых связана с передачей генетической информации, — например, белки транскрипции, трансляции и репликации. А вот белки, обеспечивающие обмен веществ в цитоплазме, оказались в большой степени унаследованными от бактерий, причем от разных (см. рис. 10.7). Сильно упрощая, можно сказать, что “информационные” гены, ответственные за работу самого генома, эукариоты получили в основном от архей, а вот “операционные” гены, ответственные за питание клетки и ее взаимодействие с внешней средой, — в основном от бактерий.
Судя по всему, эти данные невозможно объяснить без введения нового для нас понятия — горизонтальный перенос генов. Вообще говоря, то, что одновременно живущие и часто вовсе не родственные друг другу живые организмы более или менее постоянно обмениваются между собой генами “по горизонтали”, — это на сегодняшний день общепризнанный факт. Причем перенос может происходить как в результате прямого захвата клеткой обрывков ДНК из внешней среды, так и более сложными способами — например, с помощью РНК-содержащих вирусов, которые приносят с собой копии некоторых чужеродных генов и встраивают их в геном хозяина посредством обратной транскрипции (см. главу 9). У прокариот горизонтальный перенос генов распространен настолько широко, что любое эволюционное древо, построенное по последовательностям большого набора прокариотных генов, будет на самом-то деле представлять собой не древо, а сеть. То, что предков эукариот это тоже коснулось, само по себе совершенно неудивительно. Но интенсивность горизонтального переноса генов там, где жил предок эукариот, должна была быть чрезвычайно высокой. Вероятно, это было какое-то сложное многовидовое сообщество, в котором нахвататься соседской ДНК было легче, чем не нахвататься.
Второе соображение вот какое. При взгляде на эволюционное древо у непредвзятого наблюдателя невольно складывается довольно стойкое впечатление, что эволюционная история митохондрий и хлоропластов была принципиально различна. Хлоропласты в самом деле достаточно легко приобретаются путем обычного фагоцитоза, этому есть прямые свидетельства. А вот в отношении митохондрий таких свидетельств нет. Хлоропласты совершенно достоверно приобретались разными эукариотами много раз. Митохондрии — только один раз, общим предком всех современных эукариот. Мы даже и представить себе не можем, как должен был бы выглядеть эукариот, никогда не имевший митохондрий.
Интересно, а были ли такие эукариоты вообще? Собранные факты как-то уж слишком упорно наводят на мысль, что митохондрии (но не хлоропласты!) — это древнейший признак эукариот как таковых. Возможно, не менее древний, чем ЭПС и ядро. И вполне вероятно, что приобретение митохондрий произошло каким-то особым способом, отличающимся от обычного фагоцитоза, — недаром же во всей дальнейшей эволюции больше ни разу не случилось ничего подобного.
“Кунинский сценарий”
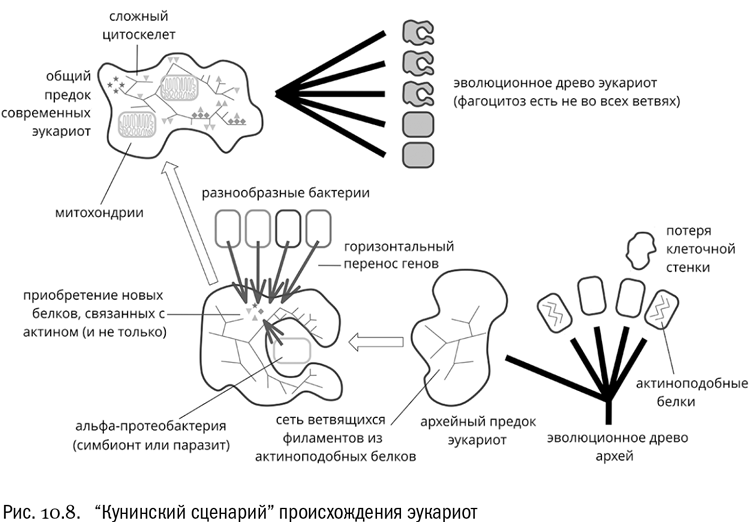
И тут возникает парадоксальная гипотеза. По мнению биоинформатика Евгения Викторовича Кунина, “эукариогенез был инициирован эндосимбиозом, а система внутренних мембран, включающая ядро, развилась как защита против инвазий бактериальной ДНК”. На всякий случай это высказывание надо пояснить. Мудреным словом “эукариогенез” тут назван сам процесс приобретения эукариотных признаков. Эндосимбиоз — это, как мы уже знаем, такой симбиоз, один из участников которого живет внутри другого. Ну а “инвазия” значит просто “вторжение”. Итак, Кунин фактически считает, что появление митохондрий было не следствием, а причиной превращения древней археи в эукариота (см. рис. 10.8).
Новый сценарий предполагает примерно следующую последовательность событий:
1 Предком эукариот была древняя архея, лишенная клеточной стенки, зато имевшая зачатки актинового цитоскелета. Это позволило архейной клетке стать крупной и приобрести сложную форму.
2 В складках наружной мембраны крупной археи стали поселяться разнообразные бактерии, которые паразитировали на ней или просто находили удобное убежище.
3 Близость этих бактерий создала поток чужеродных генов, от которого архее надо было защищаться, чтобы ее собственный геном не разладился. Именно защищаясь от чрезмерного проникновения генетического материала бактерий-паразитов, архея выработала сложную систему внутренних мембран, усовершенствовала внутриклеточный транспорт и цитоскелет. А паразитизм в конце концов превратился в симбиоз.
4 Один из бактериальных сожителей оказался настолько полезным в плане получения энергии, что архея навсегда замкнула его внутри собственной клетки, превратив из внешнего симбионта во внутреннего. Так появились митохондрии.
5 Фагоцитоз возник только в самом конце этих событий, как побочный эффект от усложнения цитоскелета и появления системы рециклизации мембран (последнее совершенно необходимо при постоянном обороте временных мембранных пузырьков, с которым связана работа ЭПС и аппарата Гольджи).
“Кунинский сценарий” имеет несколько красивых подтверждений. Например, известны современные морские археи с гигантскими клетками сложной формы, которые действительно покрыты снаружи симбиотическими бактериями. Причем эти внешние симбионты — не кто-нибудь, а протеобактерии, дальние родственники тех самых пурпурных бактерий, от которых произошли митохондрии. Кроме того, сравнительный анализ белковых последовательностей показывает, что актиновый цитоскелет, скорее всего, сначала был неподвижным. Белки, позволяющие актиновым микрофиламентам еще и сокращаться, появились заметно позже. А это означает, что начаться прямо с фагоцитоза эволюция эукариот не могла.
Споры вокруг митохондрий
На самом деле сейчас конкурируют по меньшей мере два сценария происхождения эукариот: “раннемитохондриальный” (mito-early) и “позднемитохондриальный” (mito-late). Первый предполагает, что появление митохондрий было очень ранним событием, может быть даже запустившим эукариогенез как таковой. Второй гораздо ближе к уже рассмотренным классическим представлениям: сначала сформировалась полноценная эукариотная клетка с ядром, прочими мембранными структурами и фагоцитозом, а потом она проглотила будущую митохондрию и сделала ее своим симбионтом.
В последние несколько лет “раннемитохондриальная” гипотеза успела если не утвердиться, то, во всяком случае, набрать большую популярность. И, как и следовало ожидать, ее сразу начали подвергать скептическим проверкам. Например, недавняя работа испанских биоинформатиков показала, что эукариотные белки делятся по уровню древности на три группы.
К самой древней группе относятся белки архейного происхождения, в основном ядерные и (или) выполняющие функции, связанные с передачей генетической информации. Эти белки унаследованы прямо от архейного предка основной части эукариотной клетки, включающей ядро и цитоплазму. (Иногда эту часть клетки называют “ядерно-цитоплазматический компонент”, ЯЦК.)
Более молодые белки имеют бактериальное происхождение и локализуются в цитоплазме, но не в ядре. Их функции часто связаны с мембранными структурами — эндоплазматической сетью (ЭПС) и аппаратом Гольджи. Причем вновь показано, что эти белки были заимствованы не от одной группы бактерий, а от нескольких разных — вероятнее всего, путем горизонтального переноса генов, когда архейный предок жил в составе какой-то сложной многовидовой колонии. Пока что, как видим, все соответствует модели Кунина.
А вот третья группа белков — самая молодая — происходит от альфа-протеобактерий и локализуется в митохондриях. Несомненно, они были приобретены вместе с бактериальными симбионтами, которые этими самыми митохондриями стали. Вот это уже противоречит модели Кунина, которая, совсем наоборот, предполагает, что симбиоз с предками митохондрий был “спусковым крючком эукариотизации”: сначала появились они, а потом уже ЭПС, ядро и все остальное. А тут получается, что субъектом, который приобрел митохондрии, была достаточно сложная клетка, уже являвшаяся генетической химерой, то есть совмещавшая в себе архейные и бактериальные гены.
Тем не менее считать, что модель Кунина опровергнута, пока не стоит. Ведь если архейная клетка к моменту захвата митохондрий уже имела множество бактериальных генов, то возникает вопрос: откуда она этих генов набралась? А как раз модель Кунина дает на этот вопрос очень внятный ответ. Кунин считает, что предком эукариот была неподвижная архея, имевшая зачатки цитоскелета и в связи с этим создавшая крупную “ветвящуюся” клетку со множеством складок внешней поверхности. В этих-то складках и поселились многочисленные бактерии — как альфа-протеобактерии, так и разные другие. Альфа-протеобактерии в дальнейшем были поглощены архейной клеткой и стали митохондриями — вот это, скорее всего, и запустило новый этап миграции их генов в ядро. Все остальные бактерии не были заключены внутри архейной клетки и эндосимбионтами не стали (скорее всего, потому, что архейной клетке это было не столь выгодно), но своими генами все равно успели щедро поделиться.
При всем этом не исключен (во всяком случае, не может считаться опровергнутым) и такой сценарий, согласно которому альфа-протеобактериальных симбионтов было два — древний и более поздний, причем митохондрии произошли от последнего. Эту гипотезу мы подробно обсудим в главе 14.
Предок, вывернутый наизнанку
Дальнейшим развитием “кунинского сценария” стала еще более парадоксальная гипотеза, согласно которой основная часть предковой архейной клетки соответствует ядру, и только ему. Цитоплазма же, по этой гипотезе, произошла от слившихся между собой внешних выростов архейной клетки, которые поначалу служили для обмена веществами с наружными симбионтами — предками митохондрий (см. рис. 10.9). Эту гипотезу предложили в 2014 году два американских биолога, двоюродные братья Дэвид и Базз Баумы .
Гипотеза Баумов сразу объясняет, почему от архей у эукариот остались в основном белки, связанные с генетическими процессами. Да потому, что остаток собственно архейной клетки — это ядро! Кроме того, из новой гипотезы следует, что эндоплазматическая сеть — это на самом деле участок внешнего пространства, охваченный сомкнувшимися разрастаниями первичной клетки. Тогда получается, что внутри эндоплазматической сети могут найтись остатки архейной клеточной стенки. И действительно, там обнаружены белки, очень похожие на такие остатки. Это так называемые N-гликозилированные белки, в которых к боковым цепям аминокислоты аспарагина через атом азота присоединены остатки сахаров. То, что эти белки найдены у эукариот не на поверхности клетки, а глубоко в полости ЭПС, в самом деле заставляет очень серьезно задуматься.
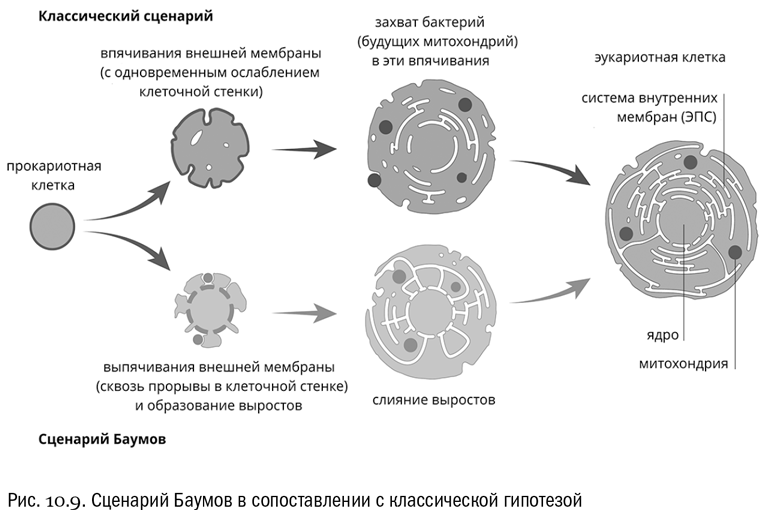
Ну а происхождение митохондрий в новой версии уж точно выглядит совершенно не связанным с фагоцитозом. Хлоропласты — иное дело, но они и приобретены были намного позже. Впрочем, в любом случае надо иметь в виду, что если само по себе симбиотическое происхождение митохондрий — твердо установленный факт, то все подробности того, как это случилось, относятся к области гипотез, которые пока что проверяются. В главе 15 мы еще вернемся к этой теме.
Дела генетические
Между прокариотами и эукариотами есть несколько серьезных отличий, касающихся не столько устройства клетки (которое можно воочию увидеть под микроскопом), сколько принципов функционирования генетического аппарата. Перечислим важнейшие из этих отличий очень кратко.
Во-первых, прокариоты и эукариоты различаются способом “упаковки” генов в геном. Молекулу ДНК, заключающую в себе весь геном либо его существенную часть и связанную тем или иным способом со специальными белками, принято называть хромосомой. У прокариот хромосома чаще всего одна, а у эукариот почти всегда несколько, и они линейные, а не кольцевые.
Во-вторых, у эукариот полностью разобщены в пространстве процессы транскрипции и трансляции. При наличии ядра иначе и быть не может. У прокариот вполне возможна ситуация, когда на торчащий “хвост” информационной РНК, синтез которой еще продолжается, сразу же садится рибосома и начинает трансляцию. У эукариот такое полностью исключено. Транскрипция идет только в ядре, трансляция — только в цитоплазме. Даже те белки, которые в силу своих функций используются исключительно внутри ядра (например, гистоны), у эукариот синтезируются в цитоплазме и потом переправляются в ядро через ядерные поры.
В-третьих, между окончанием транскрипции и началом трансляции иРНК эукариот проходит через достаточно сложное созревание (процессинг), в ходе которого она химически модифицируется. Самая важная из этих модификаций называется кэпированием и относится к 5'-концу РНК (см. рис. 10.10А). Вот тут нам хорошо бы кое-что вспомнить. “Нормальная” — то есть прокариотная — иРНК оканчивается на 5'-конце нуклеозидтрифосфатом: это неудивительно, поскольку именно нуклеозидтрифосфаты служат исходными “кирпичиками”, из которых РНК собирается (см. главу 9). У эукариот к этому концевому нуклеозидтрифосфату присоединяется особый гуаниновый нуклеотид, в котором гуанин помечен дополнительной метильной группой. Причем присоединяется он через очень экзотическую, не используемую больше нигде 5'-5'-трифосфатную связь, то есть как бы задом наперед по отношению к нормальному расположению нуклеотидов в РНК. Вот этот добавочный перевернутый модифицированный нуклеотид и называется кэпом. Его функция исключительно сигнальная: без кэпа, например, эукариотная рибосома не может узнать иРНК и начать трансляцию. Ни у каких прокариот нет ничего подобного, и зачем это нужно эукариотам — никогда толком не было понятно.
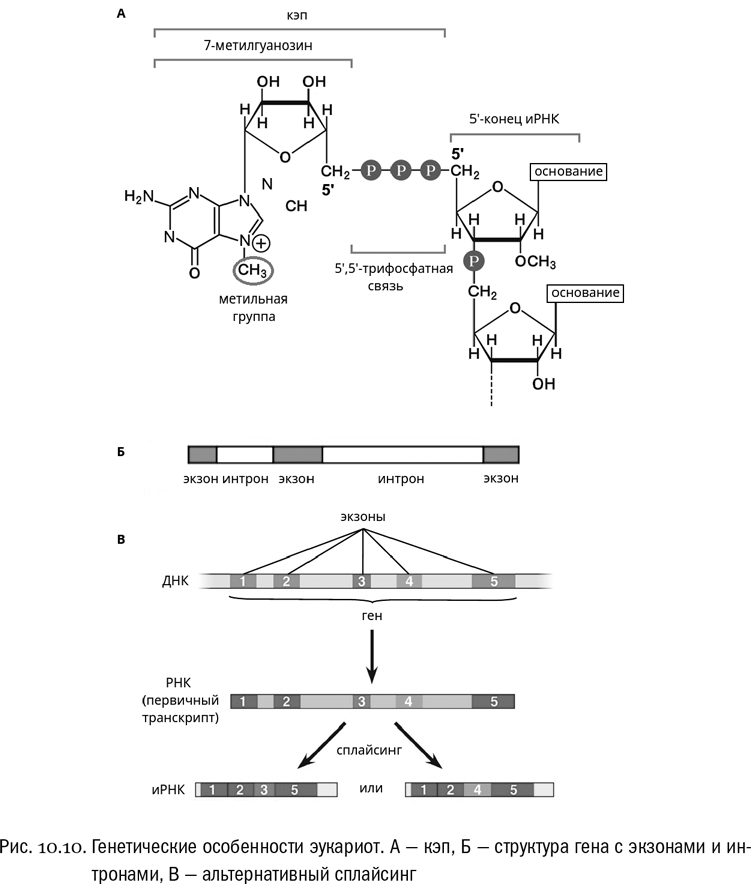
В-четвертых, в эукариотных генах, как правило, полно некодирующих вставок, которые называются интронами. Это бессмысленные отрезки ДНК, не кодирующие никаких полезных аминокислотных последовательностей и расположенные прямо внутри гена. В типичном эукариотном гене кодирующие участки (экзоны) чередуются с некодирующими — интронами — таким образом, что последние как бы разбивают кодирующую последовательность на части. В одном гене вполне могут быть десятки и сотни интронов, причем обычно они длиннее экзонов (см. рис. 10.10Б). Представим себе литературный текст, где аккуратно построенные фразы в случайных местах разрываются длинными последовательностями букв, лишенными знаков препинания и какого бы то ни было намека на смысл. Вот примерно так выглядят интроны в гене. При транскрипции вся последовательность гена (и экзоны, и интроны) переписывается на иРНК целиком, но в ходе процессинга интроны вырезаются, экзоны сшиваются и получается гораздо более короткая зрелая иРНК, состоящая из одних экзонов. Ее уже можно транслировать. Сам процесс вырезания интронов из РНК называется сплайсингом. Это еще одна составная часть процессинга, в придачу к кэпированию.
Эти особенности генетического аппарата было бы неплохо как-то объяснить. Причем в идеале — все сразу, ведь породивший их эволюционный процесс был единым. К сожалению, такого единого объяснения пока нет, но начать можно и по отдельности. Попробуем.
Интроны и ядро
Проще всего, как это ни странно, с интронами. Ключ к разгадке тут может дать вот какой факт. У прокариот интроны тоже встречаются, но в основном в генах, кодирующих рибосомные или транспортные РНК. Почему? А потому, что эти РНК не транслируются. Они проходят процессинг (вернее, сплайсинг), а потом используются по назначению как есть, образуя рибосомы или транспортируя аминокислоты. А вот процессинг информационных РНК у прокариот почти невозможен по уже названной причине: эти РНК очень часто начинают транслироваться до завершения полной транскрипции, еще буквально вися на ДНК, с которой транскрипция идет. Резать и сшивать их некогда и негде. Именно поэтому в генах, кодирующих белки, у прокариот интроны не накапливаются. Они вынужденно сразу удаляются естественным отбором — иначе белки будут получаться ни к чему не пригодными, ведь сплайсинга-то нет, а значит, не только в иРНК, но и в белке останутся большие бессмысленные куски.
Разделение процессов транскрипции и трансляции, вызванное появлением ядра, сняло это ограничение. Поэтому у эукариот интроны стали стремительно “размножаться”, а сплайсинг стал совершенно необходимым этапом подготовки эукариотной иРНК к трансляции.
Заодно это привело к тому, что появился дополнительный способ регуляции экспрессии генов — альтернативный сплайсинг (см. рис. 10.10В). Например, если в некотором гене есть пять экзонов (1, 2, 3, 4 и 5), то в одном случае можно транслировать все пять из них, во втором — только четыре (скажем, 1, 2, 4 и 5), а в третьем — тоже только четыре, но в другом составе (скажем, 1, 2, 3 и 5). Во втором и третьем случаях ненужные экзоны будут вырезаны вместе с интронами. И таким образом, манипулируя ферментами сплайсинга, можно будет получать с одного гена три разных белка. В реальности таких белков, кодируемых одним и тем же геном, может быть гораздо больше — вплоть до многих тысяч. Количество экзонов в эукариотных генах вполне допускает такое число комбинаций. И иногда это бывает очень полезно (например, для белков иммунной системы позвоночных, которым в силу их функций нужна высочайшая изменчивость).
Таким образом, в данном случае усложнение и самого генома, и механизма его работы оказалось побочным эффектом главной, “титульной” особенности эукариот — появления ядра. Откуда же оно все-таки взялось?
В 2001 году австралийский биолог Филип Белл и японский биолог Масахару Такемура почти одновременно предложили вирусную теорию происхождения ядра . Вирусы — это неклеточная и чисто паразитическая форма жизни. У вируса есть гены, но нет собственного аппарата синтеза белка. Генетическая информация у разных вирусов может записываться как на РНК, так и на ДНК. Так вот, известно, что некоторые ДНК-содержащие вирусы (например, вирус оспы) имеют оболочку из двух липидных мембран, очень похожую на клеточное ядро. Там есть даже аналоги ядерных пор. ДНК у таких вирусов линейная — тоже как в эукариотном ядре. И что самое интересное, у них есть собственный аппарат транскрипции (но не трансляции), и этот вирусный аппарат транскрипции включает механизм обязательного кэпирования иРНК — опять же устроенный так же, как у эукариот. В принципе нет ничего невероятного в предположении, что эукариоты получили этот механизм от каких-то ДНК-содержащих вирусов путем горизонтального переноса генов. Но Белл и Такемура пошли дальше. Они предположили, что все ядро целиком произошло от крупного вируса, который вселился в будущую эукариотную клетку, превратился из паразита в постоянную внутриклеточную структуру и постепенно включил в себя почти весь хозяйский геном. Эта теория хороша тем, что она логично объясняет сразу несколько особенностей эукариотной клетки: двумембранное ядро, разобщение транскрипции и трансляции, линейность ДНК, кэпирование.
Правда, популярность вирусной теории происхождения ядра, взлетевшая было несколько лет назад, сейчас снижается. То, что в ядерном аппарате эукариот есть кое-какие белки вирусного происхождения, — установленный факт. Но вот вирусное происхождение всего ядра — это совсем иное дело. К тому же вирусная теория происхождения ядра решительно противоречит уже обсуждавшейся нами теории Баумов, согласно которой ядро произошло от клетки археи, а цитоплазма — от ее сомкнувшихся выростов. Тут уж придется выбрать или одно, или другое.
В любом случае надо честно признать, что происхождение клеточного ядра на данный момент неизвестно. Если с митохондриями дело хотя бы в общих чертах ясное, то с ядром о такой степени ясности пока что и мечтать не приходится. Загадка его происхождения не решена. Хотя и можно надеяться, что она решится в ближайшее время: биология сейчас развивается быстро.
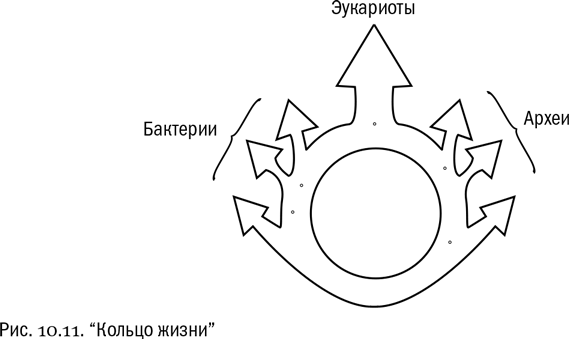
Независимо от того, какая теория происхождения ядра правильна, мы можем совершенно точно сказать, что эукариотная клетка — это химерная структура, “собранная” из составных частей нескольких неродственных организмов. Если бы обитатели Земли три миллиарда лет назад могли мыслить, эукариотная клетка, скорее всего, была бы для них таким же нелепым и невероятным созданием, как для нас — самое фантастическое чудовище из древней мифологии, вроде той же химеры, василиска или уж вовсе невероятного мирмиколеона. А с другой стороны, именно на примере с возникновением эукариот мы прекрасно видим, что ветви эволюционного древа могут не только расходиться, но и сливаться. В этом месте древо жизни превращается в “кольцо жизни” (см. рис. 10.11).
Фундамент многоклеточности
У эукариот есть две особенности, очень облегчающие всевозможные межклеточные взаимодействия. Это наличие цитоскелета и — в некоторых группах — полная потеря клеточной стенки. Скорее всего, именно благодаря своему цитоскелету эукариоты, в отличие от прокариот, относительно легко становятся многоклеточными. Без цитоскелета слишком трудно сориентировать делящиеся клетки в пространстве таким образом, чтобы они создали упорядоченную трехмерную структуру.
Но и становиться многоклеточными можно по-разному. Есть многоклеточные эукариоты, которые питаются путем фотосинтеза, то есть синтезируя глюкозу из воды и углекислоты с использованием энергии солнечного света. Такое питание называется фототрофным. Многоклеточных фототрофов принято называть растениями (в широком смысле этого слова). Жизненная форма растений возникала в эволюции эукариот несколько раз совершенно независимо, на основе разных эволюционных ветвей. Примерами тому служат зеленые наземные растения, а также красные, бурые, золотистые и другие водоросли.
Другие многоклеточные эукариоты фотосинтезировать не умеют. Некоторые из них питаются, выделяя во внешнюю среду пищеварительные ферменты, расщепляющие крупные молекулы до мелких, и всасывая потом эти мелкие молекулы сквозь свою клеточную мембрану. Такое питание называется осмотрофным, то есть всасывательным. Оно требует огромной относительной поверхности тела, поэтому многоклеточность у таких эукариот на самом деле очень условная. Как правило, большая часть их тела на протяжении большинства стадий жизненного цикла представляет собой систему тонких — толщиной в одну клетку — ветвящихся нитей, которые пронизывают субстрат (например, почву) и всасывают оттуда питательные вещества. По-настоящему сложными у таких организмов бывают только органы размножения. Самые типичные осмотрофы — это грибы. Жизненная форма гриба возникала как минимум два раза независимо в разных эволюционных ветвях эукариот (у настоящих грибов и у так называемых ложных грибов, или оомицетов).
И наконец, есть эукариоты, которые ухитрились совместить многоклеточность с фаготрофным питанием, основанным на захвате пищевых частиц отдельными клетками путем фагоцитоза. Многоклеточные фаготрофы — это животные. Насколько мы знаем, жизненная форма животных возникла за всю историю Земли только один раз, в единственной эволюционной ветви.
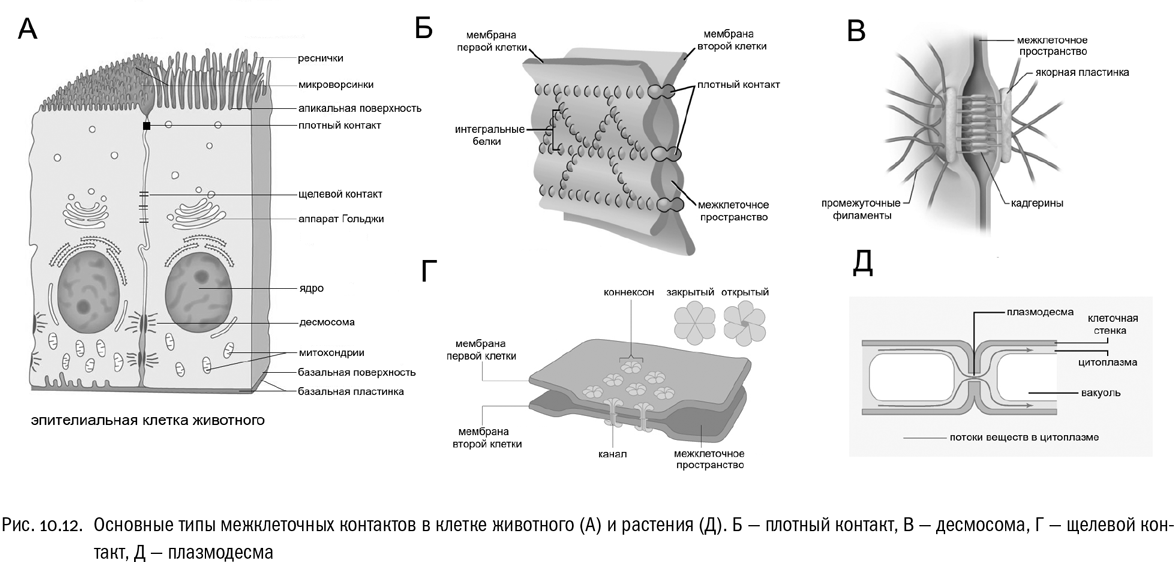
Как раз потому, что у животных нет клеточной стенки, разнообразие способов межклеточных взаимодействий у них особенно велико (см. рис. 10.12А–Г). Есть по меньшей мере четыре типа межклеточных контактов, очень важных для животных:
* плотные контакты, в которых мембраны соседних клеток предельно сближены и сшиты особыми интегральными белками (клаудинами и окклюдинами). Система таких контактов может объединить клетки в непроницаемый слой, что и требуется животным довольно часто. Со стороны цитоплазмы белки плотных контактов всегда связаны с актиновыми микрофиламентами;* десмосомы — контакты, обеспечивающие прочное механическое сцепление клеток. Интегральные белки, создающие такое сцепление, называются кадгеринами (они довольно разнообразны). Десмосомы особенно важны в тех тканях, которые часто подвергаются давлению или растяжению. Со стороны цитоплазмы белки десмосом связаны с сетью промежуточных филаментов, которая тоже достаточно прочна;* щелевые контакты, сквозь которые клетки обмениваются молекулами и ионами, плывущими по цитоплазме. Такой контакт представляет собой группу ультрамикроскопических белковых “трубочек” (правильнее говоря, каналов), пронизывающих мембраны сразу двух соседних клеток. Эти каналы могут открываться и закрываться. Состоят они из белка коннексина;* фокальные контакты, которые, строго говоря, нельзя назвать межклеточными. Они соединяют клетки с внеклеточными структурами — обычно белковыми волокнами или перепонками. Благодаря фокальным контактам клетки могут ползать, менять форму, а могут и сидеть на перепонке постоянно. Со стороны клетки фокальный контакт образуют молекулы белка интегрина, прикрепленные к актиновым микрофиламентам.
Надо обязательно добавить, что это описание, во-первых, сильно упрощено и, во-вторых, строго говоря, относится только к позвоночным животным (включая человека). У других животных межклеточные контакты могут быть устроены немного иначе, а главное — они могут состоять из других белков. Но по функциям они все равно будут аналогичны.
Кроме того, у многих животных есть и более сложные типы межклеточных контактов — например, синапсы, через которые передаются сигналы между нервными клетками. Но разговор о них уже выходит за пределы тем, охваченных этой книгой.
У растений система межклеточных контактов устроена совершенно по-другому. Главная особенность, отличающая растительную клетку от животной, — это наличие клеточной стенки, причем довольно толстой (см. главу 6). Из-за нее между клетками крайне затруднены любые другие контакты, кроме пронизывающих клеточные стенки цитоплазматических мостиков — плазмодесм (см. рис. 10.12Д). Через них могут передаваться любые вещества. В плазмодесмах мембраны соседних клеток сливаются (чего никогда не бывает в щелевых контактах животных), и их цитоплазма становится единой. Наличие системы плазмодесм означает, что все клетки данного растения слиты в единое “соклетие” — как говорят ботаники, симпласт. Это обычная ситуация для наземных зеленых растений.
Межклеточные взаимодействия — в некотором смысле самое главное, что происходит в любом многоклеточном организме. “Элементная база” этих взаимодействий, состоящая из белков с многообразными функциями, очень богата и способна быстро эволюционировать. И в то же время корни этого явления очень древние. Например, такие белки, как кадгерины и интегрины, совершенно точно есть уже у одноклеточных родственников животных. Это и есть молекулярный фундамент многоклеточности — явления, породившего самые большие и сложные живые организмы на Земле.
Назад: 9. генетическая информация
Дальше: 11. энергия

