Книга: Что такое жизнь?
Назад: Глава 2 Механизм наследственности
Дальше: Глава 4 Квантово-механическое доказательство
Глава 3
Мутации
Но эти превращенья в свой черед немеркнущими мыслями украсьте.И. В Гёте. Фауст
«Скачкообразные» мутации – инструмент естественного отбора
Изложенные нами общие факты в поддержку стойкости структуры генов, вероятно, слишком привычны, чтобы казаться удивительными либо убедительными. В данной ситуации верна пословица: исключение подтверждает правило. Если бы дети всегда были похожи на родителей, мы бы лишились не только красивых экспериментов, раскрывших нам подробный механизм наследственности, но и великого колоссального эксперимента природы, которая создает виды путем естественного отбора и выживания наиболее приспособленных.
Давайте используем последний важный вопрос в качестве отправной точки для представления значимых фактов – вновь прошу прощения и напоминаю, что я не биолог.
Мы точно знаем, что Дарвин ошибался, считая незначительные, постоянные, случайные вариации, имеющие место даже в самой гомогенной популяции, материалом естественного отбора. Доказано, что они не наследуются. Это важный факт, достойный краткого примера. Если взять урожай чистого сорта ячменя, измерить на всех колосьях длину ости и представить результаты измерений на графике, то получится колоколообразная кривая, как показано на рис. 7, где число колосьев соответствует определенной длине ости. Иными словами, преобладает определенная средняя длина, отклонения от которой в какую-либо сторону происходят с некой частотой.
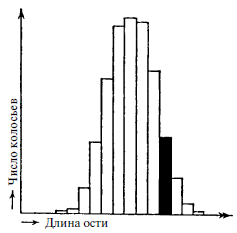
Рис. 7. Статистика длин остей у чистой линии ячменя. Группа, выбранная для посева, показана черным цветом (Это не результаты некоего эксперимента, а теоретический пример)
Теперь выберем группу колосьев (показана черным цветом), чьи ости заметно длиннее среднего и чьего количества достаточно, чтобы высадить их в поле и получить новый урожай. Проделав аналогичные измерения, Дарвин ожидал бы, что кривая сместится вправо. Он ожидал бы путем селекции добиться увеличения средней длины ости. При использовании чистой линии ячменя результат будет другим. Новая статистическая кривая окажется идентичной первой. То же самое повторится, если выбрать колосья с особенно короткой остью. Отбор не оказывает влияния, поскольку незначительные, постоянные вариации не наследуются. Они не определяются структурой наследственного материала и происходят случайно. Однако около сорока лет назад де Фриз обнаружил, что даже в потомстве чистых линий появляется очень небольшое число особей – две-три на десятки тысяч – с малыми, но «скачкообразными» изменениями. Слово «скачкообразный» характеризует не величину изменения, а дискретность, поскольку не обнаруживается промежуточных форм между исходным и измененным состоянием. Де Фриз назвал это мутацией. Дискретность является важным фактом. Она напоминает физику квантовую теорию: между соседними энергетическими уровнями нет промежуточных энергий. Физик метафорически назвал бы мутационную теорию де Фриза «квантовой теорией биологии». Позднее мы увидим, что это определение вовсе не столь метафорично. Мутации действительно происходят за счет квантовых скачков в молекуле гена. Однако квантовой теории исполнилось всего два года, когда в 1902 году де Фриз опубликовал свое открытие. Неудивительно, что лишь следующее поколение ученых обнаружило тесную связь между ними!
Они передаются потомству, а значит, наследуются
Мутации наследуются так же, как исходные, неизмененные признаки. Например, в рассмотренном выше первом урожае ячменя может появиться несколько колосьев с остями, заметно выходящими за рамки изменчивости, показанные на рис. 7, вообще без ости. Они могут быть носителями мутации де Фриза, которая передастся потомству, а значит, все их потомки будут лишены ости.
Таким образом, мутация представляет собой изменение наследия и должна определяться неким изменением наследственного материала. На самом деле, большинство важных экспериментов по скрещиванию, объяснивших нам механизм наследственности, заключались в анализе потомства, полученного скрещиванием согласно разработанной схеме, носителей мутации (во многих случаях – нескольких мутаций) с немутантными особями или с носителями других мутаций. Кроме того, поскольку мутации наследуются, они являются подходящим материалом, с которым может работать естественный отбор, производя новые виды, как описал Дарвин, путем вымирания неприспособленных и выживания сильнейших. Нужно лишь заменить в теории Дарвина «незначительные случайные вариации» на «мутации» (так же, как квантовая теория заменяет «постоянный перенос энергии» «квантовым переходом»). Во всех прочих аспектах теория Дарвина нуждалась в незначительной корректировке, если я правильно понимаю точку зрения большинства биологов. Активно обсуждался вопрос, способствует ли естественному отбору (или даже доминирует ли над ним) явная склонность мутаций происходить в благоприятном направлении. У меня на данный момент нет своей точки зрения, однако следует отметить, что возможность «направленности» мутаций далее не учитывается. Более того, я не могу уделить внимание взаимодействию генов-«переключателей» и «полигенов», как бы ни были они важны для механизма естественного отбора и эволюции.
Локализация, доминантность и рецессивность
Теперь следует рассмотреть фундаментальные факты и понятия, касающиеся мутаций, вновь в немного категоричной форме, не затрагивая напрямую подтверждающие их экспериментальные доказательства.
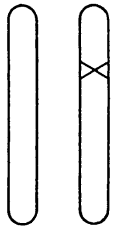
Рис. 8. Гетерозиготный мутант. Крестиком показан ген с мутацией
Разумно предположить, что некую наблюдаемую мутацию вызывает изменение определенного участка одной из хромосом. Так и есть. Важно отметить, что мы точно знаем: это изменение затрагивает лишь одну хромосому, но не соответствующий «локус» на гомологичной хромосоме. Это схематично показано на рис. 8; мутировавший локус обозначен крестиком. Факт, что мутация происходит только в одной хромосоме, можно доказать, скрестив мутантную особь («мутанта») с нормальной. Половина потомства будет обладать мутантным признаком, а другая половина – нормальным. Этого следует ожидать вследствие расхождения двух хромосом у мутанта во время мейоза, как схематично показано на рис. 9. Это «фамильное дерево», на котором каждая особь (из трех последовательных поколений) представлена парой рассматриваемых хромосом. Имейте в виду, что если у мутанта затронуты обе хромосомы, все его потомки получат одинаковое (смешанное) наследие, отличное от родительского.
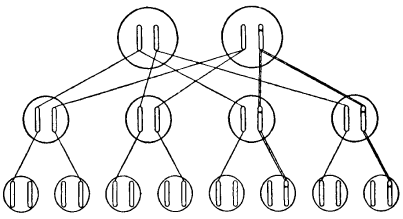
Рис. 9. Наследование мутации. Поперечные прямые линии показывают передачу хромосомы, двойные линии – передачу мутантной хромосомы. Неучтенные хромосомы третьего поколения пришли от партнеров второго поколения, не включенных в схему. Предполагается, что это не родственники, лишенные мутаций
Однако эксперименты в данной области не столь просты, как может показаться. Их затрудняет второй важный фактор, а именно то, что мутации очень часто латентны. Что это означает?
У мутанта две «копии кода» уже не идентичны, а представляют собой два различных «прочтения», или «версии», по крайней мере в одном конкретном месте. Следует отметить, что, как бы заманчиво это ни выглядело, нельзя считать исходную версию «ортодоксальной», а мутантную – «еретической». В принципе мы должны полагать, что они имеют равное право на существование, поскольку нормальные признаки также возникли благодаря мутациям.
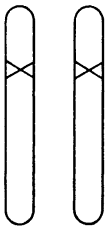
Рис. 10. Гомозиготный мутант, представленный одной четвертью потомства, появившегося путем самооплодотворения гетерозиготного мутанта (рис. 8) либо скрещивания двух гетерозиготных мутантов
В действительности «структура» особи, как правило, определяется той или иной версией, которая может быть нормальной либо мутантной. Определяющую версию называют доминантной, другую – рецессивной. Иными словами, мутация является доминантной или рецессивной в зависимости от того, меняет ли она непосредственно структуру или нет.
Рецессивные мутации встречаются намного чаще доминантных и крайне важны, хотя поначалу никак не проявляют себя. Чтобы повлиять на структуру, они должны присутствовать на обеих хромосомах (см. рис. 10). Подобные особи появляются, когда друг с другом скрещиваются два одинаковых рецессивных мутанта или когда мутант скрещивается сам с собой. Такое бывает у гермафродитных растений и даже случается спонтанно. В данном случае около одной четверти потомков будут носителями двух мутантных хромосом, а значит, у них проявится мутантный признак.
Немного технических терминов
Полагаю, для ясности следует объяснить несколько технических терминов. То, что я назвал версией кода – исходной или мутантной, – называют аллелью. Если версии различаются, как на рис. 8, особь называют гетерозиготной по данному локусу. Если версии одинаковы, как у немутантной особи или в случае, показанном на рис. 10, такие организмы называют гомозиготными. Так, рецессивная аллель влияет на структуру только в гомозиготном состоянии, в то время как доминантная аллель дает одну и ту же структуру и в гомозиготном, и в гетерозиготном состоянии.
Цвет часто доминирует над бесцветностью (или белизной). Например, горох обладает белыми цветками, только когда содержит в обеих соответствующих хромосомах «рецессивную аллель, отвечающую за белизну», то есть когда он «гомозиготен по белизне». Этот признак будет наследоваться, и все потомки данного растения станут цвести белым цветом. Однако присутствие одной «красной аллели» (в то время как другая остается белой – у «гетерозиготы») – равно как и две «красных аллели» (у «гомозиготы») – заставит горох цвести красными цветками. Различие последних двух случаев проявится только в потомстве, когда красные гетерозиготы дадут несколько белых потомков, а красные гомозиготы сохранят признак.
Факт, что две особи могут обладать совершенно одинаковой внешностью, но разной наследственностью, является столь важным, что следует точно разграничить эти понятия. Генетики говорят, что у таких особей одинаковый фенотип, однако разный генотип. Таким образом, вышеизложенное можно суммировать кратким, но в высшей степени техническим утверждением: рецессивная аллель влияет на фенотип лишь в том случае, когда генотип гомозиготен.
Мы будем использовать эти технические термины, однако при необходимости будем напоминать читателю их значение.
Опасное влияние близкородственного скрещивания
В гетерозиготном состоянии рецессивные мутации – неподходящий материал для естественного отбора. Если они вредны, как часто и бывает, то все равно сохраняются в силу своей латентности. Поэтому значительное количество неблагоприятных мутаций может накапливаться, не причиняя непосредственного вреда. Однако они передаются половине потомков – и это имеет большое значение для человека, домашнего скота, птицы и любого другого вида, физическое состояние которого нас заботит. На рис. 9 предполагается, что особь мужского пола, например я сам, несет рецессивную вредную мутацию в гетерозиготном состоянии, то есть она не проявляется. Предположим, у моей жены нет такой мутации. Тогда половина наших детей (второе поколение) окажется носителями мутации – снова гетерозиготной. Если все они опять выберут себе партнеров без мутаций (не показаны на схеме для ясности), около четверти наших внуков будут обладать гетерозиготной мутацией.
Никакого вреда не случится, если только особи с одинаковой мутацией не скрестятся друг с другом. В таком случае, как показывают нехитрые расчеты, четверть их детей будет гомозиготной, и у них неблагоприятная мутация проявится. Помимо самооплодотворения, которое возможно лишь у растений-гермафродитов, наибольшую опасность будет представлять брак между моим сыном и моей дочерью. Каждый из них обладает равными шансами оказаться или не оказаться носителем опасной мутации, и четверть таких кровосмесительных союзов приведет к тому, что у четверти их потомков она проявится. Таким образом, вероятность заболеть у родившегося от подобного союза ребенка окажется 1:16.
Аналогичным образом вероятность заболеть будет составлять 1:64 у потомка союза между двумя моими («чистокровными») внуками, являющимися двоюродными братом и сестрой. Это не слишком высокая вероятность, и второй вариант обычно считают приемлемым. Но не забывайте, что мы рассмотрели последствия лишь одного латентного повреждения у одного партнера предковой пары («я и моя жена»). В действительности они оба скорее всего несут несколько подобных латентных изъянов. Если вы точно знаете, что являетесь обладателем некой мутации, не забывайте, что 1 из 8 ваших кузенов и кузин также может нести ее! Эксперименты на растениях и животных свидетельствуют о том, что, помимо сравнительно редких серьезных пороков, существует множество не столь значительных дефектов, которые в совокупности способны повредить потомку близкородственного скрещивания. Поскольку мы больше не избавляемся от неудачных экземпляров, как это делали суровые спартанцы в горах Тайгет, следует относиться к этим вопросам серьезно, в случае человека, для которого не всегда соблюдается выживание сильнейших, а то и реализуется противоположная ситуация. Противодействующее отбору влияние современного массового убийства здоровых молодых людей всех национальностей перевешивает рассуждения о том, что в более примитивных условиях война могла производить положительный эффект, способствуя выживанию наиболее приспособленного клана.
Общие и исторические комментарии
Удивителен факт, что рецессивная аллель в гетерозиготном состоянии полностью подавляется доминантной и не дает видимого эффекта. Следует отметить, что из этого правила есть исключения. Если скрестить гомозиготный белый львиный зев с гомозиготным малиновым, все потомки первого поколения будут обладать «промежуточным» цветом, то есть розовым, а не малиновым, как можно было бы ожидать. Намного более важным примером одновременного проявления двух аллелей являются группы крови, но мы не будем здесь на них останавливаться. Не удивлюсь, если в конце концов выяснится, что рецессивность обладает уровнями и ее проявления зависят от чувствительности тестов, которые мы используем для анализа «фенотипа».
Уместно вспомнить раннюю историю генетики. Суть теории, закон наследования последующими поколениями признаков, по которым различаются родители, и – что особенно важно – разграничение рецессивных и доминантных признаков сформулировал всемирно известный аббат-августинец Грегор Мендель (1822–1884). Он ничего не знал о мутациях и хромосомах. В своем монастырском саду в Брюнне (Брно) Мендель проводил эксперименты на горохе: сажал различные разновидности, скрещивал и наблюдал за потомками в 1, 2, 3-м… поколениях. Можно сказать, он экспериментировал с мутантами, предоставленными ему природой. В 1886 году Мендель опубликовал результаты в «Трудах Брюннского общества естествоиспытателей». Никто не обращал особого внимания на хобби аббата – и уж точно никто и представить не мог, что в XX веке его открытие станет путеводной звездой для совершенно нового научного направления, одного из самых интересных в наши дни. Его статью забыли. Лишь в 1900 году о ней одновременно вспомнили Корренс (Берлин), де Фриз (Амстердам) и Чермак (Вена).
Необходимость редкости мутации
До настоящего момента мы в основном говорили о вредоносных мутациях, которые, возможно, являются самыми многочисленными; однако следует отметить, что встречаются и полезные мутации. Если спонтанная мутация есть небольшой шажок в развитии вида, то складывается впечатление, что определенные изменения «подбираются» весьма произвольным образом, рискуя причинить вред, в случае чего они тут же уничтожаются. Из этого следует важный вывод. Чтобы являться подходящим материалом для естественного отбора, мутации должны происходить редко – что и наблюдается в действительности. Если бы они случались так часто, что существовала бы вероятность появления у одной особи, скажем, десятка мутаций, вредоносные, как правило, преобладали бы над полезными, и вид не улучшался бы в результате отбора, а оставался бы прежним или вымирал бы. Умеренный консерватизм, являющийся следствием высокой стабильности генов, играет существенную роль. Можно провести аналогию с работой крупной производственной установки на фабрике. Чтобы выработать лучшие методы, необходимо применять инновации, в том числе неопробованные. Но чтобы установить, улучшит или ухудшит инновация работу, нужно пробовать их по одной, не изменяя другие детали механизма.
Мутации, вызванные рентгеновскими лучами
Теперь рассмотрим блистательный цикл генетических исследований, которые внесут наиболее значимый вклад в наш анализ.
Процент мутаций у потомства, так называемую частоту мутаций, можно повысить в сравнении с низкой природной частотой мутаций, облучив родителей рентгеновскими или гамма-лучами. Полученные таким образом мутации ничем (кроме количества) не отличаются от спонтанных, и складывается впечатление, что рентгеновские лучи также могут вызывать «природные» мутации. У дрозофилы многие особые мутации спонтанно происходят снова и снова в больших культурах; их местоположение картировали на хромосоме и дали им специальные названия. Более того, были обнаружены «множественные аллели», то есть две и более различных «версий» и «прочтений» – помимо нормальной, немутантной – одного участка хромосомного кода. Это означает существование не двух, но трех и более вариантов данного конкретного «локуса», любые два из которых относятся друг к другу как «доминантные/рецессивные», когда одновременно встречаются в соответствующих локусах двух гомологичных хромосом.
Эксперименты с мутациями, индуцированными рентгеновским излучением, создают впечатление, что каждый «переход», скажем, от нормальной особи к некому мутанту или наоборот, обладает своим «рентгеновским коэффициентом», выражающим процент потомков, которые мутировали данным образом при воздействии на родителей стандартной дозы рентгеновского излучения до зачатия.
Первый закон. Мутация – единичное событие
Более того, законы, управляющие частотой индуцированных мутаций, просты и наглядны. Здесь я буду придерживаться сообщения Н.В Тимофеева-Ресовского, опубликованного в «Биологических обзорах», т. IX за 1934 год. В значительной степени оно касается блистательной работы самого автора. Первый закон таков:
(1) Рост частоты мутаций точно пропорционален дозе облучения, поэтому можно говорить [как это сделал я] о коэффициенте роста.
Мы настолько привыкли к простой пропорциональности, что можем недооценить далеко идущие следствия этого нехитрого закона. Чтобы их понять, давайте вспомним, что, например, цена товара отнюдь не всегда пропорциональна его количеству. При обычных обстоятельствах продавец настолько удивится, если вы купите у него шесть апельсинов, что, когда вы решите взять целую дюжину, может отдать ее дешевле двойной цены за шесть штук. Во времена дефицита происходит обратное. В настоящем случае мы делаем вывод, что первая половинная доза радиации, которая, скажем, вызывает мутации у одного из тысячи потомков, не затрагивает всех остальных – не делает их более склонными или устойчивыми к мутации. Иначе повторная половинная доза не вызвала бы снова лишь одну мутацию на тысячу. Таким образом, мутация не является кумулятивным эффектом, к которому приводят последовательные небольшие дозы радиации, усиливающие друг друга. Она должна представлять собой единичное событие, происходящее в одной хромосоме во время облучения. Что это за событие?
Второй закон. Локализация события
На данный вопрос отвечает второй закон, а именно:
(2) Если варьировать характеристики (длину волны) излучения в широких пределах, от мягких рентгеновских лучей до весьма жестких гамма-лучей, коэффициент останется неизменным, при условии, что вы будете использовать одну и ту же дозу в так называемых рентгенах. То есть если определите дозу, измерив общее количество ионов, производимых на единицу объема в соответствующем стандартном веществе в том же месте и в то же время, когда подвергнутся облучению родители.
В качестве стандартного вещества используют воздух – не только ради удобства, но и потому, что органические ткани построены из элементов с таким же атомным весом. Нижний порог величины ионизации или сопряженных процессов (возбуждений) в тканях получают, умножив уровень ионизации воздуха на отношение плотностей. Таким образом, очевидно – и более серьезное исследование подтверждает это, – что единичное событие, вызывающее мутацию, представляет собой ионизацию (или сходный процесс), происходящую в некоем «критическом» объеме половой клетки. Каков этот критический объем? Его можно оценить на основании частоты мутаций посредством следующего рассуждения: если доза в 50 000 ионов на 1 см3 приводит к вероятности возникновения мутации 1:1000 в любой конкретной гамете (что оказалась в области облучения), «цель», в которую следует «попасть» ионизации для появления этой мутации, занимает лишь одну пятидесятимиллионную долю сантиметра кубического. Эти числа не соответствуют действительности и приведены исключительно ради примера. Реальную оценку дает М. Дельбрюк в статье Дельбрюка, Н. В. Тимофеева-Ресовского и К. Г. Циммера, которая также является основным источником теории, изложенной в следующих двух главах. Он приводит значение десять средних атомных расстояний в кубе, где содержится всего лишь около 103 – тысяча – атомов. Простейшая интерпретация данного результата заключается в том, что существует высокая вероятность вызвать мутацию, если ионизация (или возбуждение) имеет место «не далее чем в десяти атомах» от данного конкретного места хромосомы. Обсудим это подробнее.
В отчете Тимофеева-Ресовского есть практический намек, который я обязан упомянуть, пусть он и не имеет отношения к нашему текущему исследованию. В реальной жизни человек часто подвергается рентгеновскому излучению. Сопряженные с этим прямые опасности – такие как ожоги, лучевой рак, бесплодие – хорошо известны, для защиты от них используют свинцовые экраны, фартуки и т. п., особенно медсестры и врачи, вынужденные регулярно иметь дело с излучением. Суть в том, что, несмотря на успешную защиту от непосредственных угроз, также существует непрямая опасность возникновения вредоносных мутаций в половых клетках – подобных тем, что мы обсуждали, рассматривая неблагоприятный исход близкородственного скрещивания. Выражусь резко, пусть и немного наивно: опасность брака между двоюродными братьями и сестрами может возрасти в том случае, если их бабушка долгое время работала медсестрой и занималась рентгеном. Не следует принимать это близко к сердцу. Однако обществу нужно тревожиться о любой возможности постепенно поразить человеческую расу нежелательными латентными мутациями.

