Книга: Что такое жизнь?
Назад: Глава 1 Классический физический подход к предмету
Дальше: Глава 3 Мутации
Глава 2
Механизм наследственности
Оно [бытие] извечно; и законы Хранят, тверды и благосклонны Залоги дивных перемен.И. В. Гёте. Завет
Ожидания классического физика не тривиальны, но неверны
Итак, мы пришли к выводу, что организм и испытываемые им биологически значимые процессы должны обладать в высшей степени «многоатомной» структурой и быть защищены от случайных «одноатомных событий». Это существенно для того, говорит нам «наивный физик», чтобы организм мог подчиняться достаточно точным физическим законам, на которых основана его удивительно регулярная и упорядоченная работа. Как эти выводы, достигнутые, биологически выражаясь, a priori, то есть с чисто физической точки зрения, согласуются с реальными биологическими фактами?
На первый взгляд кажется, будто данные выводы тривиальны. Лет тридцать назад какой-нибудь биолог вполне мог сказать, что хотя для популярного лектора уместно подчеркнуть значимость статистической физики, в частности для организма, сама идея банальна. Ведь не только тело взрослого представителя любого высшего вида, но и каждая составляющая его клетка содержит «космическое» число разнообразных атомов. И каждый наблюдаемый нами физиологический процесс внутри клетки или в ходе ее взаимодействия с окружающей средой, судя по всему – по крайней мере, так казалось тридцать лет назад, – затрагивает столь огромное число отдельных атомов и атомарных процессов, что соответствующим законам физики и физической химии ничто не грозит, несмотря на очень строгие требования, накладываемые статистической физикой на «большие числа». Эти требования я только что проиллюстрировал на примере правила √n.
Теперь мы знаем, что это мнение ошибочно. Как мы вскоре увидим, небольшие группы атомов – слишком небольшие, чтобы подчиняться точным статистическим законам, – играют важнейшую роль в упорядоченных и закономерных процессах в живом организме. Они управляют наблюдаемыми крупномасштабными особенностями, которые организм приобретает в ходе развития, определяют важные характеристики его работы – и во всем этом проявляются четкие и строгие биологические законы.
Следует начать с краткого описания ситуации, сложившейся в биологии, особенно в генетике. Иными словами, нужно суммировать нынешнее положение дел в предмете, которым я не владею. Тут ничего не поделаешь, и я прошу прощения, в первую очередь – у биологов, за дилетантский характер моей сводки. Кроме того, прошу дозволения изложить основные идеи в определенном смысле категорично. Нельзя ждать от бедного физика-теоретика компетентного обзора экспериментальных доказательств, которые включают, с одной стороны, множество длинных и красиво переплетающихся серий искусных экспериментов по скрещиванию, а с другой – прямые наблюдения за живой клеткой, выполненные с использованием изощренной современной микроскопии.
Генетический код (хромосомы)
Позвольте мне использовать термин «структура» организма в том смысле, в каком биолог использует «четырехмерную структуру», имея в виду не только строение и работу этого организма во взрослом возрасте или на какой-то другой конкретной стадии, но его онтогенетическое развитие в целом, от оплодотворенной яйцеклетки до зрелости, когда он приступает к размножению. Известно, что всю эту четырехмерную структуру определяет строение одной-единственной клетки – оплодотворенной яйцеклетки. Более того, мы знаем, что структура, в сущности, определяется строением небольшой части этой клетки – ее ядра. В типичном «состоянии покоя» ядро обычно выглядит как сеть хроматина, распределенная по клетке. Однако в ходе жизненно важного процесса клеточного деления (митоз и мейоз) оно выглядит как набор частиц, обычно в форме фибрилл или палочек, называемых хромосомами. Их число составляет 8 или 12 – или, у человека, 48. Однако мне следовало записать эти показательные числа как 2 (×) 4, 2 (×) 6… 2 (×) 24… и говорить о двух наборах, в привычном биологу смысле. Хотя отдельные хромосомы иногда отчетливо видны и различимы по форме и размеру, два их набора почти идентичны. Один набор приходит от матери (яйцеклетка), другой – от отца (оплодотворяющий сперматозоид). Именно эти хромосомы – или, возможно, лишь осевые скелетные фибриллы того, что мы видим под микроскопом как хромосомы, – содержат неким образом закодированную структуру будущего развития особи и ее функционирования в зрелом состоянии. Каждый полный набор хромосом содержит полный код, таким образом, две копии кода присутствуют в оплодотворенной яйцеклетке, которая является самой ранней стадией развития будущего организма.
Называя структуру хромосомных фибрилл кодом, мы имеем в виду, что придуманный Лапласом всеведущий ум, мгновенно постигающий все причинно-следственные связи, может по ней узнать, вырастет ли из яйцеклетки при подходящих условиях черный петух или рябая курица, муха или кукуруза, рододендрон, жук, мышь или женщина. На вид все яйцеклетки удивительно схожи, а даже если и различаются – как в случае сравнительно крупных яиц птиц и рептилий, – эти различия затрагивают не основные структуры, а питательный материал, добавленный по очевидным причинам.
Разумеется, термин «код» является узким. Хромосомные структуры также участвуют в реализации развития, которое кодируют. Они объединяют в себе свод законов и исполнительную власть – или, если использовать другое сравнение, план архитектора и строительное мастерство.
Рост тела путем клеточного деления (митоз)
Как хромосомы ведут себя во время онтогенеза?
Рост организма осуществляется последовательными клеточными делениями. Такое клеточное деление называют митозом. В жизни клетки это не настолько частое событие, учитывая множество клеток, из которых состоит наше тело. Вначале рост осуществляется быстро. Яйцеклетка делится на две «дочерние клетки», они на следующих стадиях дают 4, затем 8, 16, 32, 64 и т. д. клеток. Частота делений неодинакова в различных частях растущего организма, и это нарушает регулярность числа клеток. Но на основании быстрого роста посредством простых вычислений можно установить, что в среднем 50–60 последовательных делений достаточно, чтобы получить число клеток, обнаруживаемое во взрослом человеке, или в десять раз больше, если учесть замену клеток в течение жизни. Таким образом, мое тело лишь на 50 или 60 поколений отстоит от яйцеклетки, которой я был.
Во время митоза каждая хромосома дуплицируется
Как ведут себя хромосомы в ходе митоза? Они дуплицируются – оба набора, обе копии кода. Данный процесс подробно изучен под микроскопом и очень важен, но слишком сложен, чтобы подробно описывать его здесь. Суть заключается в том, что каждая из двух «дочерних клеток» получает в наследство два полных набора хромосом, полностью идентичных набору родительской клетки. Поэтому все клетки тела имеют одинаковый хромосомный набор. Биолог простит мне удивительный случай мозаичности, который я опустил в своем кратком описании.
Хотя мы до сих пор плохо понимаем данный процесс, судя по всему, для работы организма важно, что каждая клетка, даже самая малозначимая, обладает полной (двойной) копией кода. Раньше в газетах писали, что во время своей африканской кампании генерал Монтгомери позаботился о том, чтобы каждый солдат его армии был подробно осведомлен о его планах. Если это соответствует действительности (а это вполне может ей соответствовать, учитывая сообразительность и надежность его отрядов), это отличная аналогия для нашего случая, в котором соответствующее утверждение в буквальном смысле верно. Самым удивительным фактом является двойственность хромосомного набора, поддерживаемая посредством митотических делений. Данную выдающуюся особенность генетического механизма особенно хорошо подчеркивает единственное исключение из правила, какое мы сейчас обсудим.
Редукционное деление (мейоз) и оплодотворение (сингамия)
Вскоре после начала развития особи резервируется группа клеток, которая на более поздней стадии даст начало так называемым гаметам – сперматозоидам или яйцеклеткам, – необходимым для воспроизведения организма во взрослом возрасте. «Резервирование» означает, что пока они не выполняют никаких других функций и претерпевают значительно меньшее число митотических делений. Именно посредством исключительного, или редукционного, деления (мейоза) в зрелом возрасте из этих запасенных клеток образуются гаметы, обычно незадолго до того, как должна осуществиться сингамия. При мейозе двойной набор хромосом родительской клетки просто разделяется на два одинарных, по одному для каждой из двух дочерних клеток, или гамет. Иными словами, во время мейоза не происходит митотического удвоения числа хромосом, оно остается постоянным, и потому каждая гамета получает только половину – то есть одну полную копию кода, а не две (например, у человека только 24, а не 2 (х) 24 = 48).
Клетки с одним набором хромосом называют гаплоидными (от греч. ἁπλοῦς – одиночный). Таким образом, гаметы гаплоидны, а обычные клетки тела диплоидны (от греч. διπλοῦς – двойной). Иногда встречаются особи с тремя, четырьмя… в общем, с множественными наборами хромосом. Их называют триплоидными, тетраплоидными… полиплоидными.
В ходе сингамии гаплоидная мужская гамета (сперматозоид) и гаплоидная женская гамета (яйцеклетка) сливаются в оплодотворенную диплоидную яйцеклетку. Она получает один хромосомный набор от матери и один – от отца.
Гаплоидные особи
Еще один момент требует уточнения. Для нас он не слишком важен, однако представляет значительный интерес, поскольку демонстрирует, что практически полный код «структуры» содержится в каждом одинарном наборе хромосом.
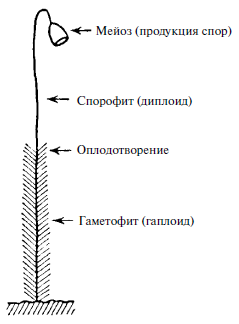
Рис. 5. Чередование поколений
В некоторых случаях непосредственно после мейоза не происходит оплодотворения, и гаплоидная клетка («гамета») претерпевает многочисленные митотические деления, приводящие к формированию полностью гаплоидной особи. Например, самец пчелы – трутень – развивается партеногенетически, то есть из неоплодотворенных, а значит, гаплоидных яйцеклеток матки. У трутня нет отца! Все клетки его тела гаплоидны. Можно назвать его огромным сперматозоидом – и действительно, как известно, это его единственная функция. Однако данная точка зрения нелепа. Такой случай не уникален. Существуют семейства растений, у которых гаплоидная гамета, образующаяся путем мейоза и называемая спорой, падает на землю и, подобно семени, развивается в истинно гаплоидное растение, по размерам сравнимое с диплоидным. На рис. 5 приведено схематическое изображение мха, распространенного в наших лесах. Облиственная нижняя часть представляет собой гаплоидное растение, называемое гаметофитом, потому что на его верхнем конце развиваются половые органы и гаметы, которые путем взаимного оплодотворения дают диплоидное растение – голый стебель со споровой коробочкой наверху. Когда коробочка раскрывается, споры попадают на землю, из них развивается облиственный стебель и т. д. Эту последовательность событий называют чередованием поколений. Можно взглянуть подобным образом на человека и животных. Однако «гаметофит» у них, как правило, представляет собой короткоживущее одноклеточное поколение, сперматозоид или яйцеклетку. Наше тело соответствует спорофиту. Наши «споры» – запасенные клетки, из них посредством мейоза образуется одноклеточное поколение.
Значимость редукционного деления
Важным, действительно судьбоносным событием в процессе размножения особи является не оплодотворение, а мейоз. Один набор хромосом достается от отца, другой – от матери. Ни случай, ни рок не может повлиять на это. Каждый человек обязан половиной своего наследственного материала отцу и половиной – матери. То, что обычно одна из них доминирует, объясняется другими причинами, до которых мы еще дойдем. Разумеется, самый простой пример такого доминирования – пол.
Но если проследить вашу наследственность до бабушки и дедушки, ситуация изменится. Возьмем набор хромосом моего отца, точнее, одну их пару, например № 5. Это верная копия № 5, которую отец получил от своего отца, или № 5, какую отец получил от своей матери. Вопрос был решен с вероятностью 50:50 в ходе мейоза, случившегося в организме моего отца в ноябре 1886 года и произведшего сперматозоид, который несколько дней спустя принял участие в зачатии меня. Аналогичную же историю можно рассказать про хромосомы № 1, 2, 3… 24 из отцовского набора и, mutatis mutandis, из материнского набора. Более того, все 48 случаев независимы друг от друга. Даже если бы я знал, что отцовская хромосома № 5 попала ко мне от моего деда Йозефа Шредингера, хромосома № 7 имеет равные шансы также принадлежать ему – или его жене Марии, в девичестве Богнер.
Кроссинговер. Местоположение признаков
Однако чистая случайность получила еще больше возможностей смешать в потомке наследие дедов и бабок, чем может показаться на основании вышеизложенного. Ранее мы полагали, будто конкретная хромосома целиком досталась нам от деда или от бабки, иными словами, что отдельные хромосомы передаются неделимыми. В действительности это не так, по крайней мере, не всегда. Прежде чем разойтись в ходе редукционного деления, например, происходящего в организме отца, любые две «гомологичные» хромосомы вступают в близкий контакт друг с другом и обмениваются целыми участками, как показано на рис. 6. Благодаря этому процессу, называемому кроссинговер, два признака, расположенные в соответствующих частях данной хромосомы, окажутся разделены у внука, который получит одно из них от деда, а другое от бабки. Процесс кроссинговера снабдил нас важной информацией о расположении признаков на хромосомах. Для полноценного рассмотрения следовало бы очертить концепции, которые будут представлены в следующей главе (такие как гетерозиготность, доминантность и т. д.), однако это выведет нас за рамки данной книги, а потому позвольте сразу перейти к сути.
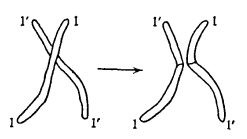
Рис. 6. Кроссинговер. Слева: контакт между двумя гомологичными хромосомами. Справа: гомологичные хромосомы после обмена и расхождения
Если бы не кроссинговер, два признака, за которые отвечает одна и та же хромосома, всегда передавались бы вместе, ни один потомок не смог бы получить один из них, не унаследовав при этом другой. Однако два признака, находящиеся на различных хромосомах, будут разделены с вероятностью 50:50 – или со 100 % вероятностью, если расположены на гомологичных хромосомах одного предка, которые не могут передаться вместе.
Кроссинговер нарушает эти правила. Таким образом, вероятность данного события можно оценить, тщательно отслеживая процентный состав потомства в длительных экспериментах по скрещиванию. Анализируя статистику, следует принять рабочую гипотезу, что «связь» между двумя признаками, расположенными на одной хромосоме, нарушается тем реже, чем ближе они находятся друг к другу. В таком случае ниже вероятность, что точка обмена окажется между ними, в то время как признаки, расположенные на противоположных концах хромосом, разделяются при каждом кроссинговере. То же самое верно для рекомбинации признаков, расположенных на гомологичных хромосомах одного предка. Таким образом, на основании «статистики связи» можно получить некую «карту признаков» каждой хромосомы.
Эти ожидания полностью подтвердились. В проведенных экспериментах, в основном – но не только – на дрозофиле, изученные признаки действительно разделяются на группы, не сцепленные друг с другом, число которых равняется числу отдельных хромосом (четыре у дрозофилы). В каждой группе можно составить линейную карту признаков, количественно соответствующую степени сцепления между любыми двумя признаками в данной группе. Поэтому нет сомнений, что они действительно расположены на хромосоме, причем линейно, что предполагает ее палочковидная форма.
Разумеется, изображенный здесь схематический механизм наследственности выглядит поверхностным и бесцветным, даже слегка наивным. Ведь мы не сказали, что именно называем «признаком». Кажется неправильным и невозможным расчленять на отдельные «признаки» структуру организма, который по существу является единым, «целым». В любом конкретном случае мы утверждаем, что пара предков различалась по явно заданным характеристикам (например, у одного были голубые глаза, а у другого – карие), и потомок в данном смысле похож на одного из них. То, что мы обнаруживаем в хромосоме, и есть причина этого различия. (Технический термин для этого – «локус», или, если имеется в виду лежащая в его основе гипотетическая структура, «ген».) На мой взгляд, различие в признаках само по себе является фундаментальной концепцией, а не признаком, вопреки очевидному лингвистическому и логическому противоречию данного утверждения. Различия в признаках дискретны, как мы поймем из следующей главы, когда будем обсуждать мутации, и я надеюсь, что приведенная здесь сухая схема обретет жизнь и заиграет красками.
Максимальный размер гена
Мы только что ввели термин «ген» – гипотетический материальный носитель определенной наследственной характеристики. Теперь следует подчеркнуть два момента, в высшей степени значимые для нашего исследования. Во-первых, размер – а точнее, максимальный размер – такого носителя; иными словами, до сколь малого объема мы сможем проследить местоположение признака? Во-вторых, постоянство гена, которое следует из устойчивости наследственной структуры.
Что касается размера, то существуют две независимые оценки. Одна опирается на генетические доказательства (эксперименты по скрещиванию), а другая – на цитологические свидетельства (прямое микроскопическое наблюдение). Первая по сути своей несложна. После того как мы выявим указанным выше способом на конкретной хромосоме достаточное число различных («широкомасштабных») признаков (например, у мушки-дрозофилы), останется разделить измеренную длину этой хромосомы на число признаков и умножить на поперечное сечение. Разумеется, мы считаем различными только те признаки, которые время от времени разделяет кроссинговер, а потому они не могут определяться одной и той же структурой – микроскопической или молекулярной. С другой стороны, очевидно, что наша оценка может дать лишь максимальный размер, поскольку число признаков, обнаруженных генетическим анализом, постоянно растет.
Другая оценка, пусть и основанная на микроскопическом наблюдении, в действительности является косвенной. Определенные клетки дрозофилы, а именно клетки слюнных желез, по какой-то причине сильно увеличены, как и их хромосомы. На этих фибриллах можно различить плотный рисунок из поперечных темных полос. С. Д. Дарлингтон отметил, что число этих полос (в его случае – 2000) хотя и намного превышает число генов, расположенных на хромосоме, как установлено в экспериментах со скрещиванием, но находится в пределах того же порядка. Он склонен считать эти полосы настоящими генами (или границами между ними). Поделив длину хромосомы, измеренную в клетке нормального размера, на число полос (2000), он получает объем гена, равный кубу с гранью 300 Å. Учитывая приблизительность оценки, можно предположить, что такой же результат был получен и первым способом.
Маленькие числа
Подробное обсуждение связи статистической физики со всеми перечисленными фактами – а точнее, связи этих фактов с использованием статистической физики в живой клетке – последует позднее. Но позвольте сейчас привлечь внимание к тому, что 300 Å – лишь 100–150 атомных расстояний в жидкости или твердой материи, а значит, ген определенно не может содержать больше миллиона или нескольких миллионов атомов. Это число слишком мало (с точки зрения √n), чтобы обеспечить упорядоченное поведение в соответствии со статистической физикой, а следовательно, и физикой вообще. Оно слишком мало, даже если все эти атомы играют одну и ту же роль, как в газе или капле жидкости. Но ген – определенно не гомогенная капля жидкости. Это крупная белковая молекула, в которой каждый атом, каждый остаток, каждое гетероциклическое кольцо выполняет особую функцию, отличную от функций других схожих атомов, остатков или колец. Так считают ведущие генетики, например Холдейн и Дарлингтон, и вскоре нам придется обратиться к генетическим экспериментам, практически доказывающим эту точку зрения.
Постоянство
Теперь второй важный вопрос: какой степени постоянства ожидать от наследуемых признаков, а значит, и материальных структур, которые их переносят?
Ответ можно дать без специальных исследований. Факт, что мы обсуждаем наследуемые признаки, свидетельствует о признании их почти незыблемого постоянства. Не надо забывать, что ребенку передается от родителя не просто та или иная черта – кривой нос, короткие пальцы, предрасположенность к ревматизму, гемофилии, дихромазии и т. п. Подобные признаки мы можем выбрать для изучения законов наследственности. Однако в действительности речь идет о полной (четырехмерной) структуре «фенотипа», видимой и проявляемой природе особи, какая воспроизводится без значимых изменений от поколения к поколению, неизменная на протяжении веков – хотя и не десятков тысячелетий, – всякий раз переносимая материальной структурой ядер двух клеток, которые объединяются, чтобы дать оплодотворенную яйцеклетку. Это чудо – уступающее лишь другому чуду, оно, пусть и тесно связанное с первым, лежит в иной плоскости. Я имею в виду тот факт, что мы, чье существование основано на чудесных взаимодействиях такого рода, все же обладаем способностью в значительной степени постигать их. Думаю, это понимание – первого чуда – может стать почти полным. Однако второе может остаться непостижимым для человека.
Назад: Глава 1 Классический физический подход к предмету
Дальше: Глава 3 Мутации

