Книга: Вопрос жизни. Энергия, эволюция и происхождение сложности
Назад: Глава 2 Что значит жить?
Дальше: Глава 4 Появление первых клеток
Часть II
О жизни
Глава 3
Энергия и начало жизни
Старинные водяные мельницы и современные гидроэлектростанции работают благодаря направленному движению потоков воды. Если перегородить реку плотиной и оставить в ней небольшое отверстие, напор воды будет гораздо мощнее и его хватит, чтобы крутить мельничное колесо. А если позволить потоку течь по широкому руслу, напор уменьшится.
Клетки живых организмов работают по тому же принципу. Метаболический путь можно сравнить с руслом, по которому “течет” углерод. В метаболическом пути последовательность химических реакций катализируется рядом поочередно действующих ферментов. Продукт реакции, катализируемой первым ферментом, является субстратом для второго, и т. д. Ферменты служат как бы берегами, в которых течет углерод, и направляют его ход. Органическая молекула входит в метаболический путь, подвергается ряду химических превращений и выходит уже в виде другой молекулы. Последовательность реакций метаболического пути надежно воспроизводится раз за разом, и исходные вещества и продукты однозначно соответствуют друг другу. В клетке одновременно функционирует множество метаболических путей. Это похоже на сеть взаимосвязанных водяных мельниц, где вода всегда с максимальным напором течет по пересекающимся каналам. Благодаря оптимальному распределению потока для роста клеток требуется гораздо меньше углерода и энергии, чем если бы поток не был ограничен. Вместо того чтобы беспорядочно участвовать в реакциях и на каждом шагу терять энергию, молекулы проходят определенный путь превращений – благодаря работе ферментов. Клеткам не нужна река, бегущая к морю: для работы их “мельниц” достаточно маленьких струек. С точки зрения энергетики, мощь ферментов заключается не столько в том, что они ускоряют реакции, сколько в том, что они делают это специфично, максимизируя выход.
Что было на заре возникновения жизни, прежде чем появились ферменты? Поток был гораздо сильнее. Чтобы расти – увеличивать количество органических молекул – и размножаться, требовалось гораздо больше энергии и углерода, чем сейчас. Клетки современных организмов минимизируют свои энергетические запросы, но мы знаем, что им до сих пор приходится использовать колоссальные объемы АТФ – имеющей повсеместное хождение энергетической “валюты”. Даже самые простые клетки, которые получают энергию, осуществляя реакцию водорода с углекислым газом, производят в процессе дыхания в 40 раз больше отходов, нежели полезной биомассы. То есть чтобы получился 1 г новой биомассы, должно образоваться минимум 40 г мусора. Жизнь – это побочный эффект главной реакции, при которой происходит выделение энергии. Так дела обстоят и после 4 млрд лет эволюции. И если даже современные клетки производят биомассы в 40 раз меньше, чем отходов, представьте, как могло обстоять дело у примитивных клеток, когда не было никаких ферментов. Фермент увеличивает скорость реакции в миллионы раз. Уберите его – и выход биомассы уменьшится в миллионы раз. Вполне возможно, что первым клеткам приходилось производить 40 т отходов (буквально грузовик), чтобы создать 1 г живой материи. Если продолжить сравнение потоков углерода и энергии с потоками воды, то первые клетки были вынуждены устраивать настоящее цунами, чтобы колеса их “мельниц” вертелись.
Вопрос о масштабах энергетических затрат имеет смысл при обсуждении всех аспектов происхождения жизни, однако он редко рассматривается достаточно подробно. Учение о происхождении жизни получило экспериментальное развитие в 1953 году, когда был поставлен знаменитый эксперимент Миллера – Юри. В том же году была опубликована статья Уотсона и Крика о структуре ДНК. Влияние двух этих работ на научный мир оказалось огромным. Область исследований происхождения жизни будто накрыло тенью от крыльев двух громадных летучих мышей. В некотором отношении тень подействовала благотворно, но были и печальные последствия. Великолепная сама по себе работа Миллера – Юри утвердила концепцию “первичного бульона”, занимавшую умы двух поколений ученых. А стараниями Уотсона и Крика в центре науки о живом оказалась генетическая информация и ДНК. Им, несомненно, принадлежат ключевые роли в происхождении жизни, но такое увлечение репликацией и происхождением естественного отбора отвлекло ученых от других важных вещей, например от изучения роли энергии.
В 50-х годах в лаборатории нобелевского лауреата Гарольда Юри работал талантливый аспирант Стэнли Миллер. В 1953 году в рамках эксперимента он пропускал электрические разряды через колбу, содержащую пары воды и смесь восстановленных (обогащенных электронами) газов. Электрические разряды имитировали молнии, а состав газовой смеси был приближен к атмосфере Юпитера. В то время считалось, что атмосфера Юпитера похожа на атмосферу древней Земли: богата водородом, метаном и аммиаком. Результаты эксперимента оказались поразительными: Миллеру удалось синтезировать несколько аминокислот – молекул, которые являются структурными единицами белков и выполняют в клетках важнейшие функции. Проблема происхождения жизни показалась неожиданно простой. В начале 50-х годов это взволновало всех гораздо сильнее, чем двойная спираль Уотсона и Крика, сначала не вызвавшая интереса. Миллер, напротив, стал знаменитостью. Его портрет в 1953 году даже появился на обложке журнала “Тайм”. Его работа оказалась весьма плодотворной. Она по-прежнему не теряет своей значимости, так как это первая экспериментальная проверка гипотезы, относящейся к происхождению жизни: будто разряды молний, проходя через атмосферу восстановленных газов, способны порождать “строительные фрагменты”, входящие в состав клеток. До появления первых организмов эти предшественники могли накапливаться в океанах, постепенно превращая их в “первичный бульон”: обогащенный раствор, содержащий органические молекулы.
Хотя сначала открытие Уотсона и Крика мало кого впечатлило, постепенно заклинание “дезоксирибонуклеиновая кислота” стало действовать. Для многих понятие “жизнь” сводится к информации, записанной в ДНК. Согласно такому представлению, происхождение жизни – это происхождение информации, без которой, разумеется, невозможна эволюция под действием естественного отбора. Происхождение информации, в свою очередь, сводится к вопросу о происхождении репликации, то есть о том, как появились первые репликаторы – молекулы, способные создавать копии самих себя. ДНК слишком сложно устроена, поэтому не годится в первичные репликаторы, но РНК (рибонуклеиновая кислота) – ее более простой и реакционноспособный предшественник – вполне подходит на эту роль. РНК – главное звено на пути от ДНК к белкам, она служит катализатором белкового синтеза, а также матрицей, на которой строится белок. Поскольку РНК способна функционировать и как матрица (подобно ДНК), и как катализатор (подобно белкам), можно предположить, что в первичном РНК-мире она служила предшественником белков и ДНК. Но откуда взялись все нуклеотиды: звенья, которые, соединяясь в цепочки, формируют РНК? Из “первичного бульона” – откуда же еще! Конечно, формирование РНК не обязательно происходило в “первичном бульоне”, но это наиболее простое предположение, позволяющее избежать сложных вопросов, связанных с термодинамикой и геохимией, которые встают при обсуждении других вариантов. Что ж, закроем глаза на слишком сложные вопросы, а с остальными разберутся доблестные молекулярные биологи. Если в последние 60 лет что-нибудь и объединяло ученых, работающих над проблемой происхождения жизни, то это концепция “первичного бульона”. В соответствии с ней, “первичный бульон” дал начало РНК-миру. Простые репликаторы постепенно эволюционировали, усложнялись, начали кодировать метаболические ферменты и, наконец, породили мир ДНК, белков и клеток, с которым мы имеем дело сейчас. Согласно этому воззрению, жизнь представляет собой информацию, которая со временем становится все сложнее.
Слабое место этой теории – проблема энергии. Конечно, энергия представлена и в рамках концепции “первичного бульона” – в виде этих самых вспышек молний. Я как-то вычислил, что для поддержания маленькой примитивной биосферы, по размеру соответствующей той, которая была до возникновения фотосинтеза, лишь за счет молний, то потребовалось бы четыре разряда в секунду на квадратный километр поверхности океана. И это если принимать эффективность роста такой, какой обладают современные организмы. Более подходящий источник энергии – ультрафиолетовое (УФ) излучение. Под его влиянием в атмосфере, содержащей метан и азот, могут образовываться реакционноспособные вещества, например цианид и цианамид – возможные предшественники биологических молекул. УФ-излучение постоянно воздействует на Землю и другие планеты. На древней Земле его интенсивность была выше, потому что защитный озоновый слой еще не образовался, а электромагнитный спектр молодого Солнца был более жестким. Химику-органику Джону Сазерленду даже удалось синтезировать активированные нуклеотиды, создав “правдоподобные первичные условия” (нуклеотиды образовались в ходе реакций с участием цианида и УФ-излучения). Но есть серьезные затруднения. На Земле нет организмов, использующих цианид в качестве источника углерода. И не известно ни одной формы жизни, которая пользовалась бы УФ-излучением как источником энергии. Напротив, оно смертельно опасно. Ультрафиолет губителен даже для современных сложных форм жизни. Хоть он и способен индуцировать образование новых органических молекул, с большей вероятностью он разрушит существующие. УФ скорее простерилизовал бы океаны, чем наполнил их жизнью. Сомневаюсь, что он может служить основным источником энергии на Земле или где-либо еще.
Ученые, считающие, что УФ-излучение сыграло роль в возникновении жизни, и не утверждают, что оно могло служить основным источником энергии. Однако, по их мнению, ультрафиолет мог способствовать образованию небольших устойчивых молекул, например цианида, которые могли со временем накапливаться. С химической точки зрения, цианид действительно хороший исходный материал для синтеза других органических соединений. Для нас он ядовит, поскольку блокирует клеточное дыхание, но это скорее причудливая особенность земной жизни, а не следствие какого-либо более фундаментального принципа. Главная проблема в том, что достижение достаточной концентрации цианида было крайне маловероятно. Это наводит на мысль о несостоятельности теории “первичного бульона”. Океаны слишком велики. При любой правдоподобной скорости образования цианида его концентрация в океане при 25 °C составила бы около двух миллионных долей грамма на литр, а этого совершенно недостаточно для того, чтобы запустить биохимические процессы. Единственный выход из этого тупика – каким-либо образом сконцентрировать морскую воду. Целое поколение ученых пыталось понять, как это могло произойти. Повысить концентрацию органических веществ в растворе можно путем вымораживания или выпаривания – но это очень жесткие методы, трудносовместимые с поддержанием клетками необходимого гомеостаза. Один сторонник цианидной теории с пеной у рта доказывал, что цианид сконцентрировался (в форме феррицианида) при испарении всего океана в результате метеоритной бомбардировки 4 млрд лет назад. Мне кажется, это просто отчаянная попытка оправдать идею, которая не работает. Такие условия среды слишком неустойчивы. Чтобы приблизиться к возникновению жизни, необходима цепь глобальных изменений внешних условий, но живые клетки при этом представляют собой стабильные структуры (их элементы непрерывно заменяются, но общее строение остается неизменным).
Гераклит говорил, что в одну реку нельзя войти дважды. Но он не имел в виду то, что река перед второй попыткой могла испариться или замерзнуть (или утечь в космическое пространство). Так же, как река течет по постоянному (по человеческим меркам) руслу, так и жизнь непрерывно обновляется, в то время как ее форма остается неизменной. Живые клетки остаются клетками, даже когда все их части заменяются в ходе непрерывного круговорота веществ. Может ли быть иначе? Я сомневаюсь. В отсутствие информации, задающей структуру – как, логично предположить, было на заре возникновения жизни, до появления способных реплицироваться молекул, – структура могла возникнуть и без этого. Но поддержание ее требовало непрерывного поступления энергии. Поток энергии провоцирует самоорганизацию материи.
Вы, вероятно, знакомы с диссипативными структурами Ильи Пригожина, например конвекционными токами, возникающими в кипящем чайнике, или крутящейся воронкой, которую образует вода над отверстием слива. Никакой информации для возникновения таких структур не требуется. Конвекционные ячейки появляются в результате неравномерного нагрева жидкости, а воронка образуется согласно закону сохранения момента импульса. Диссипативные структуры возникают за счет притока энергии и материи. Ураганы, тайфуны, водовороты – все это примеры диссипативных структур. Множество их возникает в океане и атмосфере (из-за разницы в потоках солнечной энергии на экваторе и на полюсах).
Мощные океанические течения и ветра не обусловлены никакой информацией. Они стабильны и непрерывны, как и поток энергии, которая их питает. Большое красное пятно на Юпитере представляет собой громадный шторм, антициклон, в несколько раз превышающий размер Земли. Он существует минимум несколько столетий. Все диссипативные структуры требуют непрерывного поступления энергии – как конвекционные ячейки существуют до тех пор, пока электрический ток заставляет воду кипеть и испаряться. В общем, они представляют собой зримые проявления неравновесных процессов, в которых поток энергии поддерживает структуру до тех пор, пока не будет достигнуто равновесие (в случае звезд это миллиарды лет) и структура не коллапсирует. Главная идея: за счет потока энергии могут появляться устойчивые физические структуры предсказуемого строения. К информации это не имеет никакого отношения, но мы увидим, что такие механизмы способны породить условия, благоприятные для возникновения биологической информации, репликации и отбора.
Согласно современным представлениям, основанным на химическом анализе древнейших горных пород и кристаллов циркона, атмосфера древней Земли была относительно нейтральной и состояла в основном из двуокиси углерода, азота и паров воды, подобно вулканическим газам.
Все живые организмы существуют за счет далеких от равновесия условий окружающей среды: мы сами также диссипативные структуры. Непрерывные дыхательные процессы снабжают клетки свободной энергией, необходимой для того, чтобы фиксировать углерод, расти, производить активные интермедиаты, соединять “строительные блоки” в длинноцепочечные полимеры (углеводы, РНК, ДНК, белки и так далее) и поддерживать свое низкоэнтропийное состояние, повышая энтропию окружающей среды. В отсутствие генов или любой другой информации определенные клеточные структуры, например мембраны и полипептиды, будут самопроизвольно формироваться до тех пор, пока будет продолжаться приток реакционноспособных предшественников – активированных аминокислот, нуклеотидов, жирных кислот, пока будет хватать энергии для формирования этих “строительных блоков”. Клеточные структуры существуют лишь в непрерывном потоке энергии и материи. Отдельные части могут заменяться, но общая структура стабильна и будет существовать, пока не иссякнет поток. Непрерывный приток энергии и материи – как раз то, чего не хватает в “первичном бульоне”. В “бульоне” нет ничего, что могло бы вызвать формирование диссипативных структур, которые мы называем клетками. Ничего, что могло бы заставить клетки расти и делиться: в “первичном бульоне” не было ферментов, которые могли бы запускать метаболизм и управлять его процессами. Образование клеток в таких условиях – задача, прямо скажем, не из легких. Существует ли такая среда, которая способна спровоцировать возникновение первых примитивных клеток? Безусловно, она должна была существовать. Но перед тем как мы приступим к ее поискам, обсудим, что требуется для возникновения клетки.
Как изготовить клетку
Что требуется для изготовления живой клетки? Всем клеткам на Земле присущи шесть базовых свойств. Я не хочу, чтобы книга стала похожей на учебник, но все-таки запишем их:
1) Непрерывное поступление активных форм углерода для синтеза новой органики;2) Приток свободной энергии, необходимой для биохимических процессов – образования новых белков, ДНК и т. д.;3) Катализаторы, чтобы ускорять и направлять эти метаболические реакции;4) Выделение отходов, чтобы процессы в клетке соответствовали второму началу термодинамики и можно было обеспечить прохождение химических реакций в нужном направлении;5) Компартментализация, чтобы отделить внутреннюю среду от внешней, необходимы ячеистые структуры – оболочки клеток;6) Наследственный материал – РНК, ДНК или их эквивалент, – который в деталях бы определял строение и функции биомолекул.
Все прочее (из стандартного набора свойств живых организмов, который школьники заучивают наизусть: движение, чувствительность и т. д.) – например, для бактерий лишь приятные излишества.
Не нужно много размышлять, чтобы прийти к пониманию, что эти шесть свойств глубоко взаимосвязаны и все были необходимы с самого начала. Непрерывное поступление органического углерода, очевидно, критически важно для роста, репликации… в общем, для всего. Даже на примитивном уровне происходила репликация молекул рибонуклеиновой кислоты. РНК – это цепь нуклеотидов. А нуклеотид – органическая молекула, которую нужно где-то взять. Среди исследователей, занимающихся вопросом происхождения жизни, не утихает спор, что возникло прежде: метаболизм или репликация. Но этот спор бессмыслен. Репликация – это удвоение, и расход “строительных блоков” увеличивается в геометрической прогрессии. Если запас “строительных блоков” не восполнять в том же объеме, репликация скоро прекратится.
Можно предположить, что первые репликаторы состояли не из органики, а были глинистыми минералами и т. п. (Эту идею выдвинул Грэм Кернс-Смит.) Но это не очень удачное решение: минералы недостаточно пластичны для того, чтобы кодировать нечто хоть немного приближающееся к РНК по уровню сложности, хотя они и способны быть хорошими катализаторами. Но если минералы не могут выступать репликаторами, нужно найти наиболее короткий и быстрый путь от неорганических молекул до органических, способных выполнять функцию репликаторов, например РНК. Если согласиться с тем, что нуклеотиды были синтезированы из цианамида, бессмысленно рассматривать неизвестные интермедиаты. Лучше сразу перейти к сути и принять, что некоторые среды на древней Земле были способны предоставлять необходимые для репликации органические “строительные блоки” – активированные нуклеотиды. Даже если цианамид не годится на роль отправной точки, тенденция к образованию поразительно сходного набора органических веществ в разных условиях (под действием электрических разрядов в восстановительной атмосфере, в ходе химических процессов на астероидах, в автоклавах под высоким давлением) наводит на мысль, что определенные молекулы (в число которых, возможно, входят нуклеотиды) наиболее предпочтительны с точки зрения термодинамики. В первом приближении, образование органических репликаторов требует непрерывного поступления в среду органического углерода. Поэтому мы не рассматриваем среды с очень низкой температурой: хотя органика при замерзании растворов может концентрироваться между кристаллами льда, нет механизма, который бы работал при низких температурах и мог восполнить запас нужных для продолжения процесса “строительных блоков”.
Также необходима энергия. Для того чтобы “строительные блоки” (аминокислоты или нуклеотиды) объединились и сформировали длинноцепочечный полимер (белок или РНК), нужно их активировать. Это, в свою очередь, требует источника энергии, например АТФ или похожего. Возможно, даже очень похожего. В водном мире, каким была Земля 4 млрд лет назад, источник энергии должен был обеспечивать полимеризацию длинноцепочечных молекул. В ходе полимеризации при образовании каждой связи отщепляется одна молекула воды (реакция дегидратации). В водных растворах с молекулами, подвергающимися дегидратации, возникает та же проблема, как при попытке выжать белье под струей воды. Некоторые выдающиеся исследователи были столь озадачены этим затруднением, что поддержали идею возникновения жизни на Марсе, где вода присутствует в гораздо меньшем объеме. Возникнув на Марсе, жизнь добралась сюда на метеоритах – и, выходит, все мы в некотором роде марсиане. Но, конечно, жизнь на Земле прекрасно существует в воде. Любой клетке трюк дегидратации удается тысячи раз в секунду. И люди это умеют: мы совмещаем реакцию дегидратации с расщеплением АТФ – при каждом расщеплении АТФ из среды забирается одна молекула воды. В совокупности реакции дегидратации и “регидратации” (гидролиза) приводят к переносу воды – и к высвобождению энергии, заключенной в связях АТФ. Это упрощает дело: все, что нужно – это непрерывное поступление АТФ или более простого его аналога, например ацетилфосфата. Мы займемся вопросом, откуда они могли взяться, в следующей главе. Пока отметим, что для репликации в воде нужно непрерывное обильное поступление в среду и углерода, и чего-либо похожего на АТФ.
Итак, репликация, углерод и энергия: три из шести факторов. А что насчет отделения от внешней среды? Это также связано с концентрированием. Биологические мембраны состоят из липидов, а те, в свою очередь, – из жирных кислот или изопреноидов (которые присоединяются к глицериновой головке). Когда концентрация жирных кислот превышает пороговое значение, они самопроизвольно формируют везикулы, похожие на клетки и способные расти и делиться, если их “подкармливать” новыми жирными кислотами. Чтобы обеспечить образование жирных кислот, требуется непрерывный приток органического углерода и энергии. А чтобы жирные кислоты (или нуклеотиды) накапливались быстрее, чем они рассеиваются, нужно обеспечить локальное повышение их концентрации с помощью каких-либо физических процессов или за счет природной компартментализации, чтобы сделать возможным формирование более крупных структур. В этих условиях формирование везикул вовсе не является таинством: это наиболее физически стабильное состояние, при котором общая энтропия повышается. (См. предыдущую главу.)
Если активированные “строительные блоки” поступают непрерывно, то простые везикулы будут расти и делиться самопроизвольно, в соответствии с оптимальным соотношением площади поверхности и объема. Представьте шарообразную везикулу – простую “клетку” с разнообразными органическими молекулами внутри. Везикула растет, встраивая в мембрану новые липиды и забирая в свою полость органические вещества. Теперь представим, что она вдвое увеличилась в размерах: вдвое увеличилась поверхность мембраны и вдвое возрос объем органических веществ внутри. Что при этом происходит? Если площадь поверхности возрастает вдвое, объем увеличивается более чем вдвое: площадь поверхности возрастает как квадрат радиуса, а объем – как его куб. Если скорость накопления содержимого клетки не будет выше, чем скорость увеличения площади поверхности, везикула превратится в нечто вроде гантели, а это уже на полпути к формированию двух новых везикул. Иными словами, арифметический рост приносит нестабильность, которая с большей вероятностью приведет к делению и удвоению, а не к дальнейшему увеличению везикулы. Вопрос лишь в том, сколько времени пройдет до того, как растущая везикула разделится на меньшие пузырьки. Так, непрерывный приток органических углеродных предшественников приводит не только к образованию примитивных клеток, но и к зачаточной форме клеточного деления. Кстати, этим путем – почкованием – делятся L-формы бактерий, у которых нет клеточной стенки.
Из-за проблемы соотношения площади поверхности и объема размер клетки ограничен. Это связано просто с поступлением реагентов и выведением отходов. Ницше сказал, что “брюхо служит причиной того, что человеку не так-то легко возомнить себя Богом”. На самом деле экскреция – это термодинамическая необходимость, от которой не смогли бы уклониться и божественные создания. Чтобы любая реакция могла протекать в прямом направлении, ее конечные продукты должны удаляться. Это не более загадочно, чем поведение людей на железнодорожной станции. Если уезжающие не смогут садиться в поезд с той же скоростью, с какой прибывают новые пассажиры, образуется толпа. В случае клеток скорость образования белков зависит от скорости поступления активированных предшественников (активированных аминокислот) и удаления отходов (метана, воды, CO2, этанола – всего, что может образоваться в реакции с выделением энергии). Если отходы не будут физически удаляться из клетки, это помешает протеканию прямой реакции.
Это еще один фундаментальный недостаток концепции “первичного бульона”, в котором плавают и реагенты, и отходы. Этот “бульон” не способен дать никакого импульса, никакой движущей силы, инициирующей новые химические процессы. И чем больше становится клетка, тем больше она похожа на бульон. Из-за того, что объем клетки возрастает быстрее, чем площадь ее поверхности, относительная скорость поступления углерода и удаления отходов через внешнюю мембрану по мере увеличения объема клетки должна падать. Клетка размером даже с футбольный мяч не сможет функционировать: это будет просто “бульон”. (Конечно, страусиное яйцо по размеру почти с мяч, однако желточный мешок – это просто запас пищи. Зародыш гораздо меньше.) На заре возникновения жизни природные скорости поступления углерода и удаления отходов должны были обусловливать небольшой объем клеток. Похоже, была необходима также некая разновидность физической канализации.
Это подводит нас к вопросу о необходимости катализаторов. Сейчас живые организмы пользуются белковыми катализаторами – ферментами, но РНК также может иметь некоторую каталитическую активность. Проблема в том, что РНК – уже довольно сложный полимер. Она состоит из многочисленных “строительных блоков” – нуклеотидов, и каждый нужно сначала синтезировать, активировать, а после соединить их в длинную цепь. До этого момента РНК едва ли способна выступать катализатором. Какой бы процесс ни дал начало РНК, сначала он должен был инициировать образование других, более простых органических молекул – главным образом аминокислот и жирных кислот. Получается, древний “РНК-мир” в любом случае был загрязнен многими другими органическими молекулами. Та идея, что РНК самостоятельно обеспечила возникновение метаболизма, абсурдна, даже если она сыграла ключевую роль в происхождении репликации и белкового синтеза. Так что послужило катализатором зарождающейся биохимии? Возможный ответ: это неорганические комплексы, например сульфиды металлов (особенно железа, никеля и молибдена). Они и по сей день служат кофакторами нескольких древних, сохранившихся у всех организмов консервативных белков. Хотя мы склонны приписывать каталитические свойства именно белкам, нередко они лишь оптимизируют работу кофактора, который определяет природу реакции. Оторванные от белка, кофакторы не слишком эффективны и не очень специфичны как катализаторы, но это лучше, чем ничего. Степень их эффективности зависит, опять же, от выхода реакции. Первые неорганические катализаторы лишь начали направлять потоки углерода и энергии на образование органики, но благодаря им необходимость в “цунами” отпала и стало достаточно спокойной реки.
Все эти простые органические вещества (особенно аминокислоты и нуклеотиды) проявляют некоторую каталитическую активность по отношению и к самим себе. В присутствии ацетилфосфата аминокислоты могут даже объединяться друг с другом, формируя пептиды – недлинные цепочки аминокислот. Устойчивость пептидов отчасти зависит от их взаимодействия с другими молекулами. Гидрофобные аминокислоты или пептиды, объединяясь с жирными кислотами, существуют дольше. Заряженные пептиды, которые связываются с неорганическими кластерами, например железосерными, могут быть более стабильными. Естественные связи между пептидами и минеральными кластерами могут повышать каталитическую активность минералов, а затем эти связи будут подвергаться отбору в зависимости от того, насколько связь стабильна. Представьте себе минеральный катализатор, осуществляющий органический синтез. Некоторые продукты реакции связываются с минеральным катализатором, что повышает их устойчивость и улучшает каталитические способности минерала (по крайней мере меняет их специфичность). Теоретически такая система может обеспечить появление более богатой и сложной органической химии.
Как построить клетку с нуля? Нужен непрерывный поток активного углерода и доступной химической энергии, который проходит через примитивный катализатор, преобразующий небольшую долю потока в новые органические вещества. Поток должен быть ограничен таким образом, чтобы органические вещества (в том числе жирные кислоты, аминокислоты и нуклеотиды) достигали высоких концентраций, но ничто не препятствовало выводу отходов. Сосредоточение потока может быть достигнуто путем естественного ограничения или компартментализации, что напоминает управление потоками воды на мельнице: это повышает силу потока в отсутствие ферментов и, соответственно, уменьшает потребность в углероде и энергии. Лишь если скорость образования новой органики превышает скорость, с которой она теряется, уходя во внешнюю среду, станет возможно концентрирование веществ, а затем самопроизвольное формирование структур, таких как клеткоподобные везикулы, РНК и белки.
Ясно, что это лишь намеки на будущую клетку: необходимое, но не достаточное. А сейчас сосредоточимся на очень важном моменте. Без интенсивного притока углерода и энергии, пропущенного через неорганические катализаторы, возникновение и развитие клеток невозможно. Я готов поспорить, что такая закономерность действует во всей Вселенной: учитывая потребность в углероде (см. предыдущую главу), термодинамика диктует необходимость непрерывного потока углерода и энергии через природные катализаторы. Строго говоря, это требование заставляет исключить почти все среды, которые в разное время предлагались на роль места зарождения жизни: теплые водоемы (к сожалению, Дарвин в их отношении был неправ), “первичный бульон”, пористые вулканические породы, побережья, другие планеты (в рамках идеи панспермии) – можете сами продолжить список. Но это требование не исключает гидротермальные источники. Напротив, они отлично ему соответствуют. Гидротермальные источники – как раз та разновидность диссипативных структур, которую мы ищем: работающие на непрерывном потоке и далекие от равновесия электрохимические реакторы.
Гидротермальные источники как потоковые реакторы
Большой призматический источник в Йеллоустонском национальном парке своей устрашающей оранжево-желто-зеленой окраской напоминает мне Око Саурона. Все дело в фотосинтетических пигментах бактерий, которые в качестве донора электронов пользуются выделяющимися из вулканических источников водородом и сероводородом. Хотя йеллоустонские бактерии-фотосинтетики почти ничего не могут рассказать о происхождении жизни, они дают представление о первобытной мощи вулканических источников. Если лишить растительности скалы вокруг источников, пейзаж стал бы напоминать место, где 4 млрд лет назад зародилась жизнь.
Правда, 4 млрд лет назад Земля была покрыта водой. Возможно, горячие источники имелись и на каких-нибудь крошечных вулканических островах, но в основном горячие источники находились глубоко под водой. Открытие подводных гидротермальных источников в 70-х годах XX века вызвало ажиотаж: не потому, что об их существовании никто не догадывался (их легко обнаружить по шлейфу теплой воды), а потому, что никто не представлял себе мощь “черных курильщиков” и не знал, что они выступают средоточием жизни на океанском дне. Оно представляет собой почти лишенную жизни пустыню. “Черные курильщики”, похожие на обвалившиеся дымоходы, непрерывно выбрасывающие клубы черного дыма, дали приют удивительным животным: гигантским червям, утратившим рот и анус, двустворчатым моллюскам с большую тарелку, безглазым креветкам. По плотности жизни “черные курильщики” сопоставимы с тропическими лесами. Открытие “черных курильщиков” привлекло биологов и океанографов, но особенно – ученых, занимавшихся проблемой происхождения жизни. Микробиолог Джон Баросс сразу оценил важность открытия. Он в большей степени, нежели остальные исследователи, сосредоточил внимание на необыкновенной силе и мощности неравновесных процессов, происходящих в гидротермальных источниках, скрытых от солнечного света километрами океанской воды.
Впрочем, “черные курильщики” не отрезаны от Солнца. Обитающие там животные существуют за счет симбиоза с бактериями, которые окисляют сероводород, выделяющийся из “курильщиков”. Именно сероводород является главной причиной неравновесного состояния в “черных курильщиках”: сероводород – восстановленное соединение, поэтому он реагирует с кислородом с выделением энергии. Вспомните механизм дыхания, описанный в предыдущей главе. Бактерии используют сероводород как донор электронов, кислород как акцептор электронов, а выделяющуюся энергию тратят на синтез АТФ. Но ведь кислород – побочный продукт оксигенного фотосинтеза, и до его появления на Земле кислорода быть не могло. Так что бурное развитие жизни вокруг “черных курильщиков” полностью, хотя и не непосредственно, зависит от солнечного света. Четыре миллиарда лет назад подводные гидротермальные источники выглядели совсем иначе.
Что останется, если убрать кислород? “Черные курильщики” возникают при прямом контакте морской воды с магмой в спрединговых зонах срединно-океанических хребтов или в других вулканически активных местах. Через океанское дно вода просачивается в вулканические камеры, которые залегают неглубоко под поверхностью дна. В них вода мгновенно нагревается до нескольких сотен градусов, насыщается растворенными сульфидами металлов и становится сильнокислой. Когда перегретая вода под давлением прорывается через дно, она резко охлаждается и растворенные в ней сульфиды железа, например пирит (“золото дураков”), сразу же кристаллизуются. Взвесь сульфидов железа образует тот самый черный “дым”. Четыре миллиарда лет назад “черные курильщики” были устроены почти так же – кроме того, что они были совершенно непригодными для жизни. Ведь жизнь целиком зависит от химического потенциала. Он обеспечивается наличием кислорода, а его тогда не было. Сероводород очень тяжело заставить реагировать с CO2 с образованием органики, особенно при высоких температурах. Но один революционер от науки, немецкий химик, патентный поверенный Гюнтер Вехтерсхойзер решил доказать, что на самом деле все не так, и весьма в этом преуспел. С конца 80-х годов Вехтерсхойзер опубликовал ряд новаторских статей, в которых очень подробно описал путь восстановления CO2 до органических молекул на поверхности сульфидов железа. Этот процесс он назвал пиритным пуллингом. Вехтерсхойзер говорил о “железосерном мире”, в котором железосерные минералы (FeS) катализируют образование органических молекул. По своей структуре такие минералы представляют собой кристаллы, собранные из повторяющихся ячеек из ионов двухвалентного железа (Fe2+) и сульфид-ионов (S2–). Крошечные неорганические кластеры из ионов Fe2+ и S2–, известные как железосерные кластеры, до сих пор выполняют ключевые функции во многих ферментах, в том числе участвующих в дыхании. Структура железосерных кластеров идентична кристаллической структуре минералов сульфида железа (FeS), например макинавита и грейгита (рис. 8, 11). Такие минералы могли катализировать реакции на заре возникновения жизни. Но, несмотря на то, что железосерные минералы – прекрасные катализаторы, Вехтерсхойзер экспериментально доказал, что гипотеза пиритного пуллинга в своем первоначальном виде не работает. Только взяв более реакционноспособный монооксид углерода (CO), Вехтерсхойзеру удалось синтезировать органические молекулы. И тот факт, что неизвестны организмы, способные расти за счет пиритного пуллинга, подтверждает, что дело не в неудачных экспериментах: идея и в самом деле плоха.
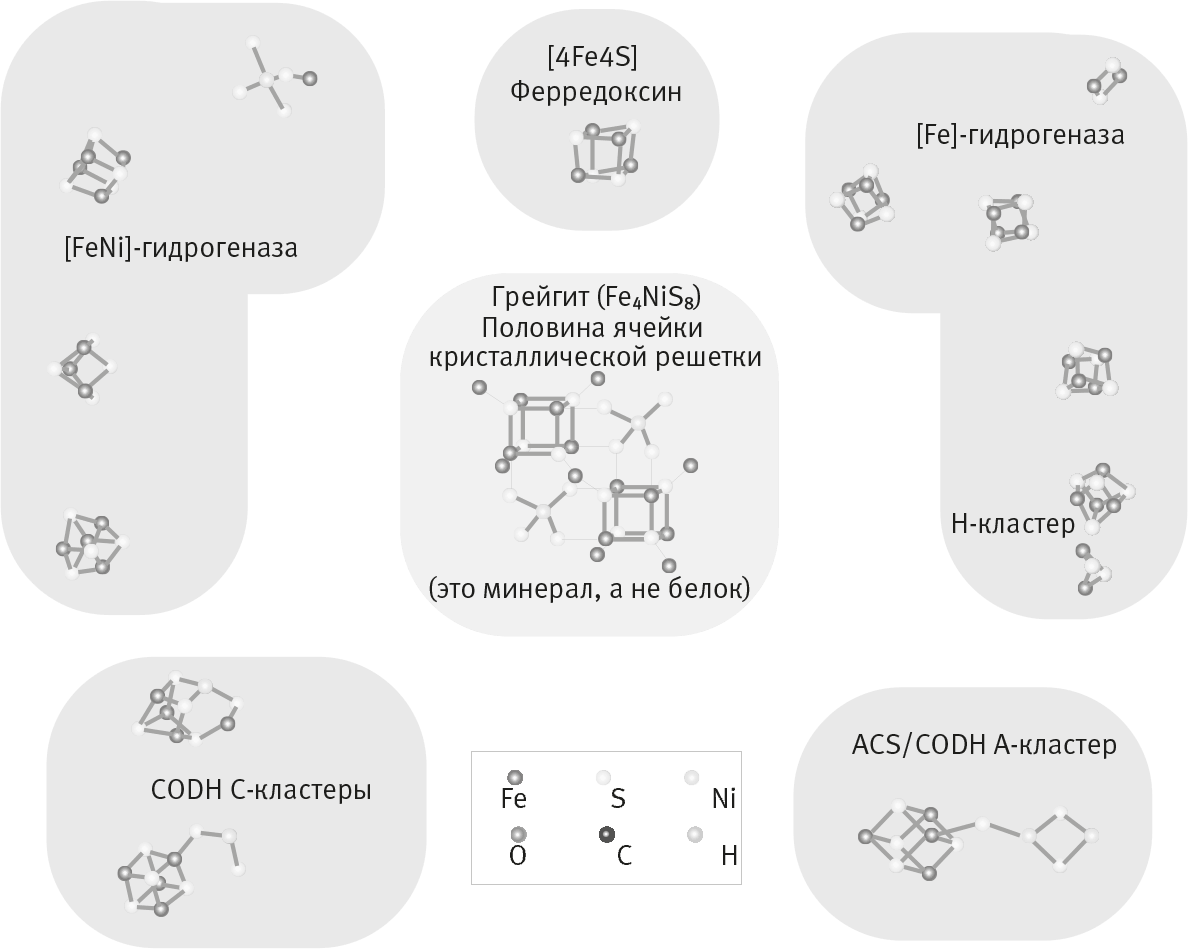
Рис. 11. Железосерные минералы и железосерные кластеры.
Близкое сходство железосерных минералов и железосерных кластеров, входящих в состав современных ферментов (Russell and Martin 2004). В центре показана повторяющаяся кристаллическая структурная единица грейгита (эти единицы в совокупности образуют решетку). Справа и слева – железосерные кластеры в составе белков. Их структура напоминает структуры грейгита и сходных минералов, например макинавита. Закрашенные области отражают форму и размер названных белков. Изображенные белки, как правило, содержат несколько железосерных кластеров (с никелем или без него).
В жерлах “черных курильщиков” содержится и CO, но его концентрация ничтожно мала, так что органическую химию так не построить. (Концентрация CO в “черных курильщиках” в 1–1000 тыс. раз ниже, чем CO2.) Есть и другая проблема. Внутри “черных курильщиков” чрезвычайно горячо: температура воды в жерле достигает 250–400 °C, но на большой глубине она не кипит из-за высокого давления. При такой температуре CO2 – самое стабильное углеродное соединение. Это означает, что синтез органических веществ в этих условиях не может осуществляться: любая синтезированная органика будет быстро разрушена и снова превратится в CO2. Предположение, что органические реакции могут протекать на поверхности минералов, также сомнительно. Если органические молекулы будут оставаться связанными с поверхностью минералов, их образование в конце концов остановится. Если же молекулы будут диссоциировать, их сразу смоет в океан, и синтез органики в прямом смысле вылетит в трубу. “Черные курильщики” еще и очень недолговечны, период их существования исчисляется десятками лет – слишком мало для зарождения жизни. Хотя “черные курильщики” представляют собой неравновесные диссипативные структуры и с их помощью можно разрешить некоторые проблемы гипотезы “первичного бульона”, их нестабильность и жесткие условия препятствуют синтезу хрупких органических соединений, необходимых для развития жизни. Но все же “черные курильщики” сыграли в этом процессе очень важную роль: насытили воды древнего океана важными для катализа ионами металлов, например железа и никеля, выщелоченных из магмы.
Преимуществами этих ионов могли пользоваться подводные источники и другого типа – щелочные гидротермальные источники (рис. 12). На мой взгляд, эти источники позволяют решить все проблемы, которые возникали с происхождением жизни в “черных курильщиках”. Щелочные гидротермальные источники, в отличие от “черных курильщиков”, образуются не из-за вулканической активности (и поэтому выглядят не столь впечатляюще), но на роль потоковых электрохимических реакторов подходят гораздо лучше. То, что щелочные источники отвечают требованиям жизни, показал геохимик Майк Рассел, опубликовав в 1988 году короткую статью в журнале “Нейчур”. В 90-х годах он написал ряд теоретических работ. Впоследствии к нему присоединился Билл Мартин, который обогатил исследования горячих источников своими микробиологическими идеями, и вместе они обнаружили множество неожиданных параллелей между источниками и живыми клетками. Как и Вехтерсхойзер, Рассел и Мартин утверждают, что жизнь началась с реакций между простыми молекулами, например H2 и CO2 – подобно тому, как автотрофные бактерии из простых неорганических предшественников синтезируют все свои органические молекулы. Рассел и Мартин также подчеркивают роль каталитических способностей железосерных минералов на ранних стадиях возникновения жизни. Мысль Рассела, Мартина и Вехтерсхойзера вращается вокруг горячих источников, железосерных минералов и происхождения автотрофности, и может показаться, что они говорят об одном и том же. В действительности их идеи радикально различаются.
Щелочные горячие источники возникают не при взаимодействии воды с расплавленной магмой, а при гораздо более мягких процессах – химических реакциях воды с горными породами. Мантийные горные породы, богатые оливином, реагируют с водой, превращаясь в гидратированный минерал серпентинит (змеевик) – красивый камень с пестрой зеленоватой расцветкой, напоминающей змеиную кожу. Серпентинит часто используют для облицовки (вспомните здание ООН в Нью-Йорке). Химическая реакция, в ходе которой оливин реагирует с водой и превращается в серпентинит, носит зловещее название “серпентинизация” (“озмеение”). Побочные продукты этой реакции и сыграли ключевую роль в возникновении жизни.

Рис. 12. Глубоководные гидротермальные источники.
Сравнение активного щелочного гидротермального источника из Затерянного города (А) с “черным курильщиком” (Б). Длина масштабной линейки в обоих случаях соответствует 1 м. Высота щелочного источника может достигать 60 м (как 20-этажное здание). Белая стрелка вверху указывает на зонд, прикрепленный к вершине источника. Более светлые участки щелочных источников – наиболее активные. В отличие от “дыма” “черных курильщиков”, гидротермальные жидкости щелочных источников не выпадают в осадок (и поэтому не выглядят как дым). Название “Затерянный город” было продиктовано возникающим там ощущением заброшенности и пустоты, однако это ощущение не вполне соответствует действительности.
Оливин богат соединениями двухвалентного железа и магния. При взаимодействии оливина с водой двухвалентное железо окисляется до трехвалентного – до соединений вроде ржавчины. Это экзотермическая (идущая с выделением тепла) реакция. В ее ходе выделяется большое количество газообразного водорода, который растворяется в теплой щелочной воде, содержащей гидроксид магния. Поскольку оливин – распространенный компонент мантии, эта реакция происходит преимущественно на дне океана вблизи спрединговых зон, где обнажаются свежие мантийные породы. Впрочем, мантийные породы в редких случаях взаимодействуют с водой непосредственно – она просачивается на глубину нескольких километров и уже там реагирует с оливином. Теплая, щелочная, насыщенная водородом жидкость поднимается (так как ее плотность ниже, чем у холодной морской воды), остывает, реагирует с растворенными в океане солями, и из нее выпадает осадок, который и формирует торчащие из морского дна столбы.
В отличие от “черных курильщиков”, щелочные источники не имеют никакого отношения к магме и формируются не над магматическими камерами в центрах океанического спрединга, а в нескольких милях от них. Вода в них не перегретая, а горячая: 60–90 °C. У них нет сквозного канала, из которого поток воды выходит в океан. Вместо этого они пронизаны сетью связанных друг с другом микроскопических пор. И вода в них не кислая, а сильнощелочная. По крайней мере, это те их свойства, которые предсказал Рассел в начале 90-х годов и которые лежали в основе его теории. Голос Рассела, одинокий и страстный, раздавался на конференциях, призывая ученых отвлечься от хтонической мощи “черных курильщиков” и обратить внимание на тихую гавань щелочных источников. Но никто не прислушивался к нему до 2000 года, когда был открыт первый подводный щелочной источник, названный Затерянным городом. Он удивительно соответствует почти всем предсказаниям Рассела, вплоть до местоположения – около 10 миль в сторону от Срединно-Атлантического хребта. Когда все это происходило, я стал задумываться о связи биоэнергетики с происхождением жизни и впервые написал об этом (в книге “Кислород”, опубликованной в 2002 году). Эти идеи сразу привлекли к себе внимание. Для меня в гипотезе Рассела важнее всего то, что она однозначно связывает происхождение жизни с протонным градиентом. Остается вопрос: как именно?
Как важно быть щелочью
Щелочные гидротермальные источники обеспечивают точно те условия, которые требуются для возникновения жизни: мощный приток углерода и энергии, проходящий над неорганическими катализаторами, и специфические ограничения этого потока, которые позволяют добиться высоких концентраций органики. Гидротермальные жидкости насыщены растворенным водородом и, в меньших количествах, другими восстановленными газами, включая метан, аммиак и сероводород (который в щелочной среде превращается в сульфид-ион). Гидротермальные источники Затерянного города и другие известные щелочные источники имеют микропористую структуру: у них нет главного “жерла”. Составляющая их порода похожа на минерализованную губку, где крошечные, от микрометров до миллиметров в диаметре, поры разделены тонкими стенками, вместе образуя огромный лабиринт, сквозь ходы которого просачиваются наружу гидротермальные жидкости. Из-за того, что эти жидкости не перегреваются, контактируя с магмой, их температура способствует не только синтезу органических молекул, но и тому, что жидкости вытекают медленно. Вместо того чтобы неистово извергаться, жидкости спокойно протекают над каталитическими поверхностями. И гидротермальные источники существуют тысячелетиями – как Затерянный город, которому минимум 100 тыс. лет. Рассел указывает, что если перевести в более подходящие для химических процессов единицы времени, получится 1017 микросекунд. Неимоверно долго!
Теплые течения по лабиринту крошечных пор способствуют концентрированию органических молекул (в том числе аминокислот, жирных кислот и нуклеотидов), обеспечивая возрастание концентраций до значений, в миллионы раз превышающих изначальные. Это становится возможным благодаря термофорезу. Принцип примерно такой же, как в стирке: в стиральной машине мелкое белье нередко скапливается внутри большого пододеяльника. Оба этих явления зависят от кинетической энергии. При высоких температурах маленькие молекулы (и белье) кружатся и танцуют, вольные двигаться в любом направлении. А по мере того как гидротермальные жидкости смешиваются друг с другом и остывают, кинетическая энергия органических молекул падает, и они уже вовсе не так свободны (это и происходит с носками, когда их окутывает одеяло). Это означает, что они, скорее всего, уже не покинут место, где оказались. Так органические молекулы скапливаются в областях с более низкой кинетической энергией (рис. 13). Эффективность термофореза отчасти зависит от размера молекул: крупные, вроде нуклеотидов, концентрируются сильнее, чем молекулы меньшего размера. Низкомолекулярные конечные продукты, например метан, легко покидают гидротермальный источник.
Итак, постоянное гидротермальное течение сквозь микропористые стенки источников должно концентрировать органику путем активного динамического процесса, который не изменяет равновесное состояние (как, например, вымораживание или упаривание), а, напротив, сам является равновесным состоянием. Более того, термофорез обеспечивает химические взаимодействия органических молекул, благодаря чему внутри пор источника могут возникать диссипативные структуры.

Рис. 13. Термофорез позволяет добиться очень высоких концентраций органики.
А. Щелочной гидротермальный источник из Затерянного города (в разрезе). Заметна пористая структура стенок: здесь нет центрального жерла, вместо него – лабиринт пор (от микрометров до миллиметров в диаметре).
Б. Органические вещества, например нуклеотиды, теоретически могут достигать концентраций, более чем в 1 тыс. раз превышающих первоначальные, путем термофореза за счет конвекционных токов и тепловой диффузии в порах источника.
Г. Пример экспериментального термофореза, осуществленного в нашем реакторе в Университетском колледже Лондона. Здесь показан флуоресцентный органический краситель (флуоресцеин) в пятитысячекратной концентрации на пористой керамической пене (диаметр 9 см).
Д. Другое флуоресцентное вещество, хинин, концентрируется еще сильнее, минимум в миллион раз.
Возможно, это звучит слишком хорошо, чтобы быть правдой. Вблизи щелочных гидротермальных источников Затерянного города цветет жизнь, пусть и представленная в основном бактериями и археями. В этих источниках в небольших количествах образуется органика: метан и следы других углеводородов. Но новых форм жизни там определенно не возникает, и даже среды, богатой органикой, в результате термофореза не образуется. Конечно, уже живущие там бактерии подчищают ресурсы, но есть и более фундаментальные причины.
“Черные курильщики” 4 млрд лет назад отличались от сегодняшних. И химия щелочных гидротермальных источников в то время была иной. Конечно, в чем-то они очень похожи. Сам процесс серпентинизации не должен был измениться: теплые, насыщенные водородом щелочные потоки, по всей видимости, растекались по морскому дну и тогда. Но химия океана была совсем другой, а это должно было сказаться на минеральном составе щелочных источников. Сейчас “курильщики” Затерянного города сложены в основном из карбонатов (арагонита), а другие похожие источники, открытые позднее (например Стритан в Исландии), состоят из глин. Что творилось в катархейских океанах 4 млрд лет назад, мы не можем точно сказать. Неизвестно, какие именно структуры должны были тогда формироваться, но ясно, что двумя ключевыми факторами, определяющими их тип, являлось отсутствие кислорода и гораздо более высокая, чем сейчас, концентрация CO2 в воздухе и океане. Из-за этих отличий древние щелочные источники в роли потоковых реакторов должны были быть гораздо эффективнее.
В отсутствие кислорода железо переходит в раствор в виде двухвалентного иона. Мы знаем, что в древних океанах было много ионов железа: они выпали в осадок, образовав огромные слоистые железные формации (гл. 1). Большая часть этих растворенных ионов попала в воду из “черных курильщиков”. Также мы знаем, что железо должно было осаждаться, формируя щелочные гидротермальные источники – не потому, что мы это наблюдали, а потому, что это должно быть обусловлено химическими закономерностями (и мы можем воссоздать этот процесс в лабораторных условиях). Железо будет выпадать в осадок в виде гидроксидов и сульфидов, которые образуют каталитические кластеры. Такие кластеры входят в состав ферментов, контролирующих метаболизм углерода и энергии (например в составе белка ферредоксина). Когда кислорода не было, минеральные стенки щелочных источников должны были содержать каталитически активные железные минералы – скорее всего с примесями других активных металлов, например никеля и молибдена (который растворяется в щелочных жидкостях). А это очень похоже на настоящий потоковый реактор: насыщенные водородом жидкости циркулируют в лабиринте микроскопических пор, стенки которых каталитически активны и где удерживаются и концентрируются продукты, а отходы – удаляются.
Но что именно вступает в реакцию? Вот мы и подобрались к сути: роли высоких концентраций CO2. В щелочных гидротермальных источниках в наше время углерода относительно мало, потому что большая часть доступного неорганического углерода осаждается на стенах источника в виде карбоната (арагонита). По всей видимости, в катархее, 4 млрд лет назад, концентрация CO2 была существенно выше (в 100–1000 раз), чем сейчас. Высокие концентрации CO2 не только обеспечивали источники углеродом, но и делали воду океанов кислее, что препятствовало осаждению карбоната кальция. (В наши дни возрастающая концентрация CO2 – и, как следствие, закисление океанов – представляет угрозу для коралловых рифов.) Уровень pH современных океанов – около 8, это слабощелочная среда. В катархее же океаны, скорее всего, были нейтральными или слабокислыми, с pH = 5–7 (геохимические данные не дают точные значения). Благодаря сочетанию высокой концентрации CO2, слабокислых вод океанов, щелочных потоков и тонких стенок, содержащих FeS, стали возможными химические процессы, которые в иных условиях было непросто осуществить.
Химическими процессами управляют термодинамика и кинетика. Законы термодинамики предопределяют, какие из состояний материи более стабильны – иными словами, какие молекулы будут получаться в результате реакции при условии неограниченного времени. Законы кинетики указывают скорость реакций и позволяют понять, какие продукты образуются при реакции за фиксированный срок. Исходя из термодинамических закономерностей, CO2 будет реагировать с водородом (H2) с образованием метана (CH4). Это экзотермическая реакция, она протекает с выделением тепла. За счет этого повышается энтропия окружающей среды – по крайней мере в определенных условиях, благоприятствующих протеканию реакции. Реакция пойдет самопроизвольно, если предоставить ей такие условия (в данном случае это умеренная температура и отсутствие кислорода). Если температура слишком высока, CO2 становится стабильнее метана. А в присутствии кислорода водород будет реагировать преимущественно с ним, образуя воду. Четыре миллиарда лет назад умеренная температура и бескислородная среда щелочных источников должны были способствовать реакции CO2 с H2 с образованием CH4. Даже сейчас, в присутствии кислорода, Затерянный город производит немного метана. Геохимики Ян Аменд и Том Макколлом рассчитали, что в условиях щелочного гидротермального источника образование органических веществ из H2 и CO2 остается термодинамически выгодным до тех пор, пока в среде нет кислорода. Это впечатляет. В таких условиях, при 25–125 °C, формирование тотальной клеточной биомассы (аминокислот, жирных кислот, углеводов, нуклеотидов и т. д.) из H2 и CO2 действительно экзергонично. Это значит, что органическая материя при таких условиях должна самопроизвольно формироваться из H2 и CO2. В результате образования клеток выделяется энергия и повышается общая энтропия.
Но (и это сильное возражение) заставить реагировать H2 и CO2 очень непросто: препятствует кинетический барьер. Хотя термодинамика и утверждает, что они должны самопроизвольно реагировать, некое препятствие не дает этому случиться. H2 и CO2 почти безразличны друг к другу. Чтобы заставить их вступить в реакцию, нужно много энергии: бомба, которая проломит барьер. Тогда H2 и CO2 вступят в реакцию. Вначале будут образовываться частично восстановленные вещества. CO2 может принимать электроны только парами. Приобретая пару электронов, CO2 превращается в формиат (HCOO–). Приняв еще два электрона – в формальдегид (CH2O), а следующие два – в метанол (CH3OH). Наконец, последняя пара дает полностью восстановленный метан (CH4). Конечно, живые организмы состоят не из метана – основой жизни служит частично восстановленный углерод, в грубом приближении представляющий собой нечто среднее между углеродом в формальдегиде и углеродом в метане. Это означает, что на пути возникновения жизни из CO2 и H2 стоят два кинетических барьера. Первый нужно преодолеть, чтобы получить формальдегид или метанол. А второй не должен быть преодолен! Приняв H2 и CO2 в свои теплые объятия, клетка в последнюю очередь желала бы, чтобы реакция пошла до конца и образовался метан. В этом случае ее содержимое разлетелось бы, рассеялось, улетучилось. Похоже, жизнь точно знает, как сделать первый барьер ниже и как поддерживать второй (опуская его лишь при необходимости энергии). Но что случилось в самом начале?
Если бы существовал простой и экономичный способ заставить CO2 реагировать с H2, затрачивая на это не более энергии, чем получается на выходе, мы немедленно им воспользовались бы. Это был бы огромный шаг на пути к решению глобальной энергетической проблемы. Представьте себе: искусственный фотосинтез, с помощью которого можно расщепить воду на H2 и O2! И это уже реальность. Но у водородной энергетики есть свои недостатки. Гораздо лучше заставить реагировать H2 и CO2 из воздуха, получая природный газ – или даже синтетический бензин! Тогда можно сразу сжигать этот газ на электростанциях. При этом выброс CO2 уравновешивался бы его забором. Мы смогли бы остановить повышение концентрации CO2 в атмосфере, избавиться от зависимости от ископаемого топлива. Энергетический кризис нам точно не грозил бы. Пока мы не преуспели в поиске экономичного осуществления этой реакции. Вряд ли можно добиться более высокого выхода. А вот клетки, даже самые простые, делают это постоянно. Так, метаногены получают нужные им для роста углерод и энергию в результате реакции H2 с CO2. Но вот еще более сложный вопрос: как эта реакция могла идти до возникновения живых клеток? Вехтерсхойзер отверг такую возможность. Он решил, что жизнь не могла начаться с реакции CO2 и H2, потому что те просто не стали бы реагировать. Даже высокое давление, как на дне океана, где находятся гидротермальные источники, не заставило бы H2 вступить в реакцию с CO2. Вот почему Вехтерсхойзер отстаивал идею пиритного пуллинга. Но один способ все же есть.
Протонная сила
Окислительно-восстановительные реакции предполагают перенос электронов от донора (в данном случае H2) к акцептору (CO2). Готовность молекулы отдавать свои электроны называют восстановительным потенциалом. Это не слишком правильная аналогия, но она довольно проста для понимания. Если молекула “хочет” избавиться от своих электронов, восстановительный потенциал принимает отрицательное значение. Чем сильнее она хочет это сделать, тем отрицательнее потенциал. Напротив, если атом или молекула жаждут электронов и готовы их отрывать почти отовсюду, то восстановительный потенциал положителен (можно рассматривать это как притягивание отрицательно заряженных электронов). Кислород “хочет” захватывать электроны (окисляя любое вещество, у которого он их забирает), демонстрируя свой очень высокий положительный восстановительный потенциал. Все эти понятия на самом деле относятся к так называемому стандартному водородному электроду, но сейчас не стоит об этом беспокоиться. Важно, что молекула с отрицательным восстановительным потенциалом склонна избавляться от электронов, передавая их любой молекуле с более высоким восстановительным потенциалом, но не куда-либо еще.
В этом и проблема H2 и CO2. При нейтральном pH = 7 восстановительный потенциал H2 составляет –414 мВ. Если H2 отдаст свои два электрона, от него останется два протона: 2H+. Восстановительный потенциал водорода отражает этот динамический баланс – склонность H2 терять электроны, превращаясь в H+, и склонность 2H+ захватывать электроны, образуя H2. Если бы CO2 приобрел эти электроны, он превратился бы в формиат. Но восстановительный потенциал формиата равен –430 мВ. Это означает, что он склонен отдавать электроны на H+ с образованием H2 и CO2: фактически это обратная реакция. Формальдегид и того хуже. Его восстановительный потенциал примерно равен –580 мВ. Он крайне неохотно удерживает собственные электроны и с легкостью отдаст их протонам (с образованием H2). Таким образом, если рассматривать реакцию при pH = 7, Вехтерсхойзер прав: H2 никоим образом не сможет восстановить CO2. Но некоторые бактерии и археи живут именно благодаря этой реакции, а значит, она возможна. Мы подробно рассмотрим, как они это делают, в следующей главе. А сейчас все, что нам нужно знать: что бактерии, живущие на H2 и CO2, могут расти, лишь получая энергию за счет трансмембранного протонного градиента. Это и есть ключ.
Восстановительный потенциал молекулы часто зависит от pH – иначе говоря, от концентрации протонов. Причина достаточно проста. При переносе электрона переносится и отрицательный заряд. Если молекула, которая восстанавливается, может принять еще и протон, продукт получается более стабильным, потому что положительный заряд протона компенсирует отрицательный заряд электрона. Чем больше протонов доступно для компенсации заряда, тем проще осуществить перенос электрона. Веществу становится легче принять пару электронов. Действительно, восстановительный потенциал повышается при закислении среды примерно на 59 мВ с каждой единицей pH. Чем кислее раствор, тем проще передать электроны на CO2 и получить формиат или формальдегид. К сожалению, то же самое относится и к водороду. Чем раствор кислее, тем проще перенести электроны на протоны с образованием водорода H2. И если просто изменить pH, это не возымеет эффекта. Восстановить CO2 при помощи H2 по-прежнему будет невозможно.
А теперь подумайте о протонном трансмембранном градиенте. Концентрация протонов – кислотность – по разные стороны мембраны различается. Точно такое же различие наблюдается в щелочных источниках. Щелочные гидротермальные потоки держат путь через лабиринт микропор. Сквозь них же течет слабокислая океанская вода. В некоторых местах потоки соприкасаются: при этом океанская вода, насыщенная CO2, отделяется от щелочных потоков тонкой неорганической стенкой, содержащей FeS минералы с полупроводниковыми свойствами. Восстановительный потенциал H2 в щелочных условиях становится ниже: водород в такой среде отчаянно желает отринуть свои электроны, а оставшийся после этого H+ может спариться с OH– в щелочных потоках, порождая стабильную воду. При pH = 10 восстановительный потенциал H2 равен –584 мВ: это сильный восстановитель. А при pH = 6 восстановительный потенциал для формиата равен –370 мВ, для формальдегида он равен –520 мВ. Иными словами, при такой разнице водороду довольно легко восстановить CO2 с образованием формальдегида. Единственный вопрос: как именно электроны переносятся с H2 на CO2? Дело в том, что железосерные минералы в тонких неорганических перегородках микропористых источников проводят электроны – пусть не так хорошо, как медная проволока. Теоретически физическое строение щелочных источников могло обеспечить восстановление CO2 водородом, в результате чего могла получиться органика (рис. 14). Фантастика!
Но так ли это? Вот она, красота науки: это простой вопрос, который можно проверить. Не то чтобы это было просто. Некоторое время я уже пытаюсь это сделать в лаборатории вместе с химиком Барри Херши и аспирантами Александрой Уичер и Элоем Кампруби. На деньги фонда “Леверхульм траст” мы построили настольный реактор, чтобы попробовать провести эти реакции. Осаждение в лаборатории тонких полупроводниковых стенок из сульфида железа – дело не слишком простое. Еще одна проблема – формальдегид нестабилен и он “хочет” вернуть свои электроны протонам, а при этом вновь образуются H2 и CO2. Легче это происходит в кислотной среде. Определенный pH и точная концентрация водорода критически важны. И в лаборатории, разумеется, затруднительно воспроизвести размеры настоящих источников (десятки метров в высоту), а тем более обеспечить высокое давление (это позволяет добиться гораздо более высокой концентрации газов, например водорода). Эксперимент прост в том смысле, что вопрос четко сформулирован и проверяем, а ответ поможет многое узнать о происхождении жизни. Нам удалось получить формиат, формальдегид и другие простые органические вещества (включая рибозу и дезоксирибозу).
Пока примем теорию такой, какова она есть, и предположим, что реакция пойдет именно так, как предсказано. Что произойдет? Медленный, но устойчивый синтез органических молекул. (Каких именно молекул, разберем в следующей главе. Пока лишь отметим, что это предположение проверяемо.) После образования органики ее концентрация будет увеличиваться в тысячи раз по сравнению с изначальной за счет термофореза, обеспечивая формирование везикул и, возможно, таких полимеров, как белки. (И это предположение можно проверить в лаборатории.) Первые шаги вдохновляют: флуоресцентный краситель флуоресцеин, по размеру такой же, как нуклеотид, в нашем потоковом реакторе увеличивает свою концентрацию минимум в 5 тыс. раз, а хинин может концентрироваться еще сильнее (рис. 13).
Итак, что означают рассуждения о восстановительных потенциалах? Они одновременно и ограничивают спектр, и выявляют широкий диапазон условий, при которых во Вселенной могла возникнуть жизнь. Да, из-за подобных вещей ученые часто выглядят так, как будто они живут в изолированном мирке, потерявшись среди абстрактных размышлений. Может ли тот факт, что восстановительный потенциал водорода падает вместе с pH, приблизить нас к разрешению проблемы? Да. В условиях щелочных гидротермальных источников H2 будет реагировать с CO2 с образованием органических молекул. Почти в любых иных условиях этого не случится. Выше я успел отвергнуть почти все остальные среды, которые могли обеспечить происхождение жизни. Исходя из законов термодинамики, мы установили, что для того, чтобы построить живую клетку с нуля, требуется непрерывный поток активного углерода и химической энергии, проходящей над примитивными катализаторами в системе, которая ограничивает и направляет движение потока. Лишь гидротермальные источники могут обеспечить необходимые условия, и лишь единственная их разновидность – щелочные гидротермальные источники – обладает абсолютно всеми необходимыми свойствами. Но идея щелочных источников одновременно порождает серьезную проблему – и дает красивое ее решение. Серьезная проблема состоит в том, что хотя эти источники богаты водородом, водород не будет просто так вступать в реакцию с CO2 с образованием органики. А красивое решение – это физическое строение щелочных источников: естественные протонные градиенты на тонких полупроводниковых стенках в теории могут инициировать образование органических веществ. А затем обеспечить их концентрирование. На мой взгляд, все это имеет смысл! Учтите при этом, что жизнь использует (до сих пор) трансмембранные протонные градиенты, чтобы обеспечивать энергией и углеродный, и энергетический метаболизм, и мне хочется стенать вместе с Джоном Арчибальдом Уилером: “О, могло ли быть иначе! Как мы все могли так долго оставаться слепы?!”
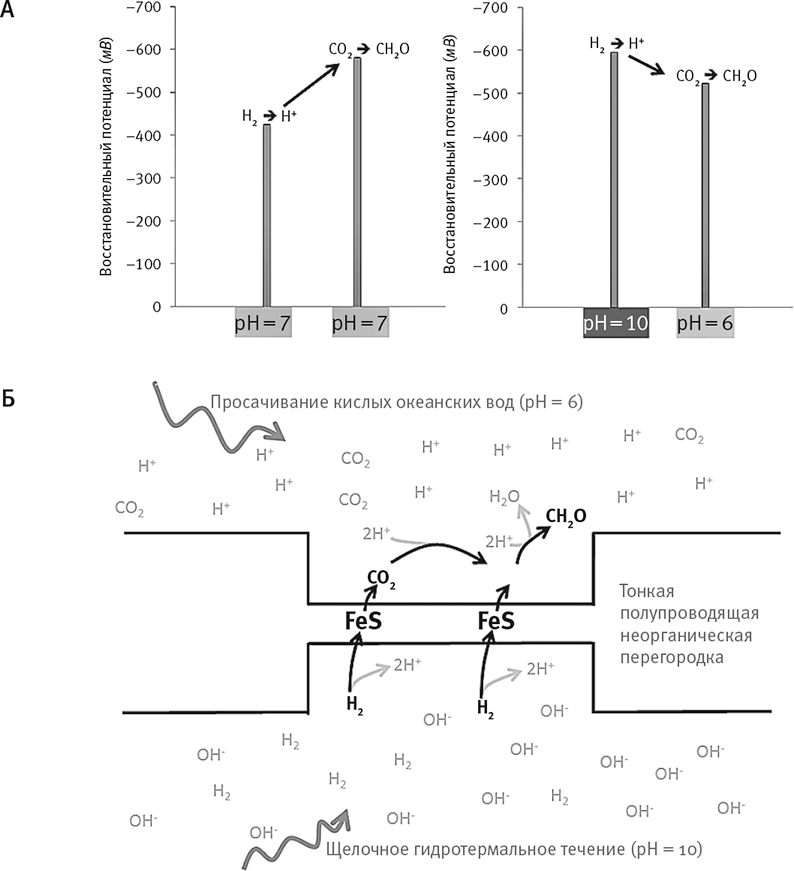
Рис. 14. Как получить органику из H2 и CO2.
А. Зависимость потенциала восстановления от водородного показателя (pH). Чем более отрицателен потенциал восстановления, тем выше вероятность, что вещество отдаст один или более своих электронов. Чем он положительнее, тем вероятней, что вещество примет электроны. Обратите внимание, что значения на шкале y становятся отрицательнее с увеличением высоты. При pH = 7 H2 не способен отдавать электроны на CO2 с образованием формальдегида (CH2O). Реакция будет идти преимущественно в противоположном направлении. Однако если H2 окажется в среде с pH = 10 (как в гидротермальных щелочных источниках), а CO2 – при pH = 6 (как в древнем океане), теоретически возможно восстановление CO2 до CH2O.
Б. В пористом источнике жидкости с pH = 10 и pH = 6 могли находиться в соседних порах и отделяться друг от друга тонкой полупроницаемой стенкой, в составе которой имелся сульфид железа (FeS), ускоряющий реакцию восстановления CO2 до CH2O. FeS выступает в роли катализатора, как и в процессе нашего дыхания перемещая электроны H2 к CO2.
Но – давайте успокоимся. Восстановительный потенциал ограничивает и одновременно выявляет набор условий, при которых должна была возникнуть жизнь. Мы выяснили, что условия, которые в наибольшей мере способствовали бы возникновению жизни, реализуются в щелочных источниках… Что, и это все? Должны быть другие варианты! О’кей, может быть. В бесконечной Вселенной возможно все, но не все вероятно. Возникновение жизни в щелочных источниках вероятно. Такие источники формируются в результате химической реакции между водой и оливином – одним из самых распространенных минералов во Вселенной, главной составляющей космической пыли и аккреционных дисков, из которых формируются планеты, включая Землю. Серпентинизация оливина может происходить даже в космосе – при насыщении водой космической пыли. Согласно некоторым гипотезам, когда образовалась наша планета, вода при возрастании температуры и давления дала начало океанам. Как бы то ни было, оливин и вода – одни из самых широко распространенных веществ во Вселенной. Другой необходимый компонент – газ CO2. Он входит в состав атмосферы большинства планет Солнечной системы и найден в атмосфере экзопланет.
Определенный минерал, вода и газ: вот список необходимого для жизни. Мы найдем все это почти на любой влажной каменистой планете. В соответствии с законами химии и геологии, эти вещества будут формировать теплые щелочные гидротермальные источники с протонными градиентами между тонкостенными каталитическими микропорами. Мы можем на это рассчитывать. Возможно, особенности их химии не всегда будут способствовать жизни. Этот эксперимент идет прямо сейчас на каждой из 40 млрд подобных Земле планет одного только Млечного Пути. Мы живем в чашке Петри вселенского масштаба. Как часто прекрасные условия дают начало жизни, зависит от того, что происходит дальше.
Назад: Глава 2 Что значит жить?
Дальше: Глава 4 Появление первых клеток

