Книга: Вопрос жизни. Энергия, эволюция и происхождение сложности
Назад: Глава 3 Энергия и начало жизни
Дальше: Часть III О сложности
Глава 4
Появление первых клеток
“Я так думаю”, – в 1837 году записал Дарвин в дневнике. Эти слова нацарапаны рядом со схематическим рисунком, изображающим дерево жизни. Прошел всего год с момента, когда Дарвин вернулся из плавания на “Бигле”. Спустя 22 года этот (несколько более художественно выполненный) рисунок стал единственной иллюстрацией в “Происхождении видов”. Концепция дерева жизни оказала такое влияние на гипотезы Дарвина и даже на современную эволюционную биологию, что попытки заявить, что она неверна, кажутся кощунством. В 2009 году, спустя 150 лет после публикации “Происхождения видов”, именно такое заявление было крупно напечатано на обложке журнала “Нью сайентист”. Несмотря на провокационный заголовок (призванный привлечь читателей), статья была гораздо сдержанней и содержала вполне определенную мысль. В некотором смысле концепция дерева жизни действительно неверна. Но отсюда не следует, что важнейшая из гипотез Дарвина – эволюция путем естественного отбора – также неверна. Из этого следует лишь, что представления Дарвина о наследственности были неполны. И это не новость. Хорошо известно, что Дарвин ничего не знал о ДНК, генах и законах Менделя, не говоря уже о переносе генов между бактериями, так что механизмы наследственности он представлял себе довольно смутно. Но ничто из этого не дискредитирует дарвиновскую теорию естественного отбора, так что заявление на обложке справедливо лишь в очень узком смысле.
Но фраза с обложки журнала поднимает очень важный вопрос. Дерево жизни подразумевает “вертикальный” способ наследования: родители путем полового размножения передают потомкам копии своих генов. Передача генов происходит в основном в рамках видов, с редкими случаями межвидового скрещивания. Репродуктивно изолированные популяции медленно дивергируют, так как возможности скрещивания между ними снижаются, и в конце концов превращаются в новые виды. Именно так на дереве жизни возникают разветвления. Но у бактерий все гораздо запутаннее. У них нет настоящего полового размножения, как у эукариот, поэтому они не образуют обособленных видов. Определение того, что такое “вид” у бактерий, всегда представляло собой проблему. Но наибольшие затруднения вызывает горизонтальный перенос генов (ГПГ): бактерии способны не только завещать свой геном дочерним клеткам, но и могут раздать его в виде милостыни всем желающим. Но опять-таки, ничто из этого никоим образом не опровергает теорию естественного отбора. Основа осталась той же: наследственность и изменчивость, вот только изменчивость реализуется куда большим числом способов, чем считалось.
Распространенность ГПГ у бактерий ставит перед нами вопрос: что мы вообще можем о них знать? По своей фундаментальности этот вопрос сравним со знаменитым физическим принципом неопределенности. Ведь почти все деревья построены по результатам анализа лишь одного гена. Пионер молекулярной филогенетики Карл Везе после тщательного отбора остановился на рибосомальной РНК (рРНК) малой субъединицы рибосомы. Везе утверждал (и у него были основания), что этот ген универсален у всех организмов и очень редко передается горизонтальным переносом, поэтому филогенетический анализ этого гена покажет “единое истинное дерево жизни” клеток (рис. 15). В некотором смысле это действительно верно: когда клетка делится, дочерние клетки всегда наследуют гены рРНК предковой клетки. Но что произойдет, если через много поколений остальные гены из-за ГПГ окажутся замененными на другие? Со сложными многоклеточными организмами такое случается крайне редко. Мы можем секвенировать рРНК орла, и ее последовательность укажет, что орел – это птица, а отсюда можно сделать вывод, что у орла есть клюв, перья, когти, крылья, что он откладывает яйца, и т. д. Это так, потому что вертикальное наследование генов обеспечивает устойчивую корреляцию между рибосомальным “генотипом” и общим фенотипом организма – гены, кодирующие признаки птицы, наследуются все вместе на протяжении многих поколений. И, хотя они могут в процессе изменяться, изменения редко приводит к непредсказуемым последствиям.

Рис. 15. Знаменитое, но неверное трехдоменное дерево жизни.
Эту схему составил Карл Везе в 1990 году. Дерево построено на основе анализа одного высококонсервативного гена (гена рибосомальной РНК малой субъединицы рибосомы). Местоположение “корня” выявлено анализом расхождений между парами паралогичных генов, общих для всех клеток (которые дуплицировались еще у Последнего всеобщего предка), и оно показывает, что и археи, и эукариоты гораздо ближе друг к другу, чем к бактериям. Но это верно лишь для ключевых информационных генов. Большая доля остальных генов эукариот родственна генам бактерий, а не архей. Так что приведенная схема вводит нас в заблуждение. Ее можно рассматривать исключительно как филогенетическое дерево одного гена, но не как дерево жизни.
Представим теперь, что было бы, если у птиц был бы распространен ГПГ. Последовательность рРНК такого организма указывала бы на то, что перед нами птица. Но взглянув на такую “птицу”, мы увидели бы, что у нее нет крыльев, зато есть хобот, шесть лап, глаза на коленках, шерсть, а еще она мечет икру, как лягушка, и воет, как гиена. Это, конечно, абсурд, но именно с этим мы сталкиваемся, изучая бактерии. Чудовищные химеры ежесекундно пролетают у нас перед глазами, и мы не вопим от ужаса лишь потому, что те, как правило, очень малы и морфологически невзрачны. Тем не менее бактерии (если рассматривать их геномы) всегда химерны, а иные из них в генетическом отношении столь же чудовищны, как наша “птица”. У кого действительно есть причины вопить от ужаса при виде бактерий, так это у ученых-филогенетиков. На основании анализа рРНК мы не можем установить, как выглядит клетка и какой она была в прошлом.
Для чего тогда нужен филогенетический анализ по одному гену, если он ничего не может рассказать нам о происхождении клетки? В некоторых случаях он полезен – это зависит от выбранных временных масштабов и темпов ГПГ. Когда темпы горизонтального переноса низкие (у растений, животных, большинства протистов и некоторых бактерий), то на небольшом временном промежутке мы можем наблюдать устойчивую корреляцию между рибосомальным генотипом и фенотипом. Если же скорость ГПГ высока, корреляция быстро исчезает. Патогенные штаммы кишечной палочки (E. coli) отличаются от безвредных не последовательностями рРНК, а наличием генов, определяющих патогенность. Геномы разных штаммов E. coli могут различаться на 30 % (в 10 раз больше различий, чем между геномами человека и шимпанзе, но при этом мы продолжаем считать все кишечные палочки одним видом). Правда, если нас интересуют патогенные свойства, то филогения рРНК – последнее, что нам пригодится. За долгое время корреляция между рибосомальным генотипом и фенотипом исчезает даже при очень низких темпах ГПГ. Отсюда следует, что почти невозможно установить, какой образ жизни 3 млрд лет назад вела бактерия: за такой срок ГПГ мог заменить почти все ее гены.
Поэтому все величие древа жизни оказывается обманчивым. У нас была надежда, что удастся воссоздать истинную филогению всех клеток, установить, как одни виды возникали из других, проследить за ними до самого истока и воссоздать облик предка всего живого. Если бы мы действительно могли это сделать, мы смогли бы узнать о Последнем всеобщем предке (Last Universal Common Ancestor, LUCA) все, в том числе получить данные о составе его мембраны, среде обитания и потребляемых для роста веществах. К сожалению, мы не можем получить такие точные сведения. Билл Мартин провел очень остроумный тест, который назвал “удивительное дерево с исчезающими ветвями”. Он выделил 48 генов, консервативных для всех живых организмов, и построил филогенетические деревья для каждого гена, чтобы установить родство между 50 бактериями и 50 археями (рис. 16). Вблизи концов ветвей все 48 генов показывали одни и те же родственные взаимосвязи внутри сотни видов бактерий и архей. То же самое наблюдалось и вблизи корня: почти все гены “согласились” с тем, что корень этого дерева жизни находится где-то между бактериями и археями (фактически – с тем, что Последний всеобщий предок был прародителем бактерий и архей). Но при построении глубоких ветвей внутри бактерий или внутри архей оказалось, что все 48 генов показывают разное. Все 48 генов дают разные деревья! Этот может быть проблемой технического характера (размытие сигнала с увеличением расстояния), либо результатом ГПГ – принцип вертикального наследования генов нарушается, когда отдельные гены начинают случайным образом меняться местами. Мы не знаем, какой вариант верен, и пока нет возможности это узнать.
В сущности это означает, что мы не можем определить, какие группы бактерий или архей наиболее древние. Согласно одному дереву, наиболее древняя группа архей – метаногены. Согласно другому, это не так, и реконструкция облика древнейших клеток оказывается практически невозможной. Но даже если удалось бы доказать, что метаногены – действительно древнейшие археи, все равно не было бы уверенности, что они всегда вырабатывали метан, как современные метаногены. В соединении генов в одну последовательность для увеличения силы сигнала тоже мало проку, поскольку каждый ген может иметь свою историю, и такой анализ даст недостоверные результаты.
Но то, что все 48 выбранных Биллом Мартином универсальных генов помещают самую древнюю развилку дерева жизни между бактериями и археями, внушает некоторую надежду. Если мы сможем выяснить, какие свойства являются для бактерий и архей общими, а какие различаются и, по всей видимости, приобретены позднее, мы можем составить “фоторобот” Последнего всеобщего предка. Но и здесь мы сталкиваемся с затруднением: гены, общие для архей и бактерий, могли возникнуть лишь в одной из групп, а в другую попасть путем ГПГ: известно, что вполне возможен горизонтальный перенос между организмами всех трех доменов. Если ГПГ произойдет на ранних этапах эволюции (на пустых участках “удивительного дерева с исчезающими ветвями”), то будет казаться, что эти гены унаследованы от общего предка (вертикальный перенос), хотя на самом деле это не так. И чем полезнее ген, тем активнее он должен был распространяться на ранних этапах эволюции. Для того, чтобы отбросить влияние горизонтального переноса, мы вынуждены снова переключиться на гены, универсальность которых не вызывает сомнений и которые являются общими для представителей всех групп бактерий и архей. Это должно хоть отчасти уменьшить вероятность того, что эти гены распространялись путем горизонтального переноса на ранних этапах эволюции. Но беда в том, что известно менее сотни таких универсальных генов, то есть чрезвычайно мало, и воссозданный по ним портрет Последнего всеобщего предка выходит очень странным.

Рис. 16. Удивительное дерево с исчезающими ветвями.
На рисунке результат сравнения филогенетических схем, построенных для 50 бактерий и 50 архей по 48 универсальным и консервативным генам. Последовательности 48 генов были слиты в одну для увеличения статистической мощности (это обычный метод в филогенетике). По последовательности “супергена” построено дерево, демонстрирующее родственные связи внутри 100 видов. Построили деревья по каждому гену и каждое сравнили с “супергенным”. Для каждой ветви градиентом серого указано число деревьев, построенных по отдельным генам, которые совпадают с деревом, построенным по слитой последовательности. У основания дерева почти все 48 генов выдают то же дерево, что и слитая последовательность, указывая, что бактерии и археи действительно разошлись очень давно. На кончиках ветвей большинство деревьев, построенных по отдельным генам, также совпадают с деревом, построенным по слитой последовательности. Но мы не видим более глубоких ветвей внутри обеих групп: все построенные по отдельным генам деревья имеют иной порядок ветвления, нежели дерево, построенное по слитой последовательности. Это может быть и следствием горизонтального переноса генов, который перетасовал все точки ветвления, и размытием статистического сигнала за невообразимо долгие четыре миллиарда лет.
Мы уже сталкивались с этим (гл. 2). Cудя по всему, у Последнего всеобщего предка уже имелись белки и ДНК, а значит, в то время уже существовал универсальный генетический код, с ДНК считывались РНК-транскрипты, которые, в свою очередь, транслировались в белки на рибосомах – универсальных для всех клеток молекулярных фабриках, строящих белки. Механизмы, необходимые для прочтения ДНК и синтеза белков, задействуют множество молекул белков и РНК, общих для бактерий и архей. Структуры и последовательности этих белков и РНК не несут следов горизонтального переноса между группами – похоже, их механизмы транскрипции и трансляции разошлись на ранних этапах эволюции и затем развивались независимо. Движемся дальше. И бактерии, и археи используют энергию протонного градиента на мембране для синтеза АТФ. Фермент АТФ-синтаза – еще одна изумительная молекулярная машина, не уступающая по замысловатости рибосоме и, по-видимому, не менее древняя. Как и рибосомы, АТФ-синтазы универсальны и консервативны у всех живых организмов. Между бактериальными и архейными АТФ-синтазами есть незначительные структурные различия: по-видимому, эти АТФ-синтазы отделились от предковой АТФ-синтазы Последнего всеобщего предка, а их гены почти не переносились между доменами. Так что у Последнего всеобщего предка, судя по всему, наряду с рибосомами, РНК и ДНК, имелась и АТФ-синтаза. Кроме этого, у архей и бактерий есть несколько общих базовых биохимических путей, например пути синтеза аминокислот и некоторые реакции цикла Кребса, и они, по всей вероятности, присутствовали и у Последнего всеобщего предка. В общем, это почти все.
Что касается различий, то их чрезвычайно много. У архей и бактерий различается большинство ферментов, используемых в репликации ДНК. Хотя, казалось бы, что может иметь более фундаментальный характер для клетки? Разве что мембрана – но и мембраны у архей и бактерий различаются! То же самое можно сказать о клеточной стенке. У архей и бактерий принципиально различное строение обоих барьеров, отделяющих клетки от внешней среды. Почти не представляется возможным установить, чем вместо всего этого располагал Последний всеобщий предок. Различия можно перечислять и далее, но здесь мы остановимся. Из шести фундаментальных для клеток процессов (поток углерода, поток энергии, катализ, репликация ДНК, компартментализация и экскреция) у архей и бактерий сходны лишь первые три, да и то не во всем.
На этот счет существует несколько объяснений. Первое: все признаки у Последнего всеобщего предка могли быть продублированы, а впоследствии бактерии потеряли одну из копий, а археи – другую. Хотя это звучит довольно глупо, его нельзя необоснованно отвергать. Так, известно, что смесь бактериальных и архейных липидов способна образовывать устойчивые мембраны, и поэтому, возможно, у Последнего всеобщего предка имелись липиды обоих типов, а его потомки специализировались, утратив либо один, либо другой тип. Заметим, что этот сценарий может быть справедлив лишь для небольшого числа признаков, иначе возникает проблема, известная как геном Эдема. Если Последний всеобщий предок обладал всеми признаками, а его потомки специализировались, потеряв некоторые из них, то, следовательно, у Последнего всеобщего предка был чрезвычайно большой геном: гораздо больше, чем у современных прокариот. Я думаю, что предполагать, что сложное появилось прежде простого и что на каждую проблему было сразу два решения – все равно, что ставить телегу впереди лошади. К тому же совершенно непонятно, почему потомки Последнего всеобщего предка потеряли по половине всех его признаков. Мне кажется это неправдоподобным.
Вторая гипотеза гласит, что Последний всеобщий предок представлял собой совершенно обычную бактерию с мембраной, клеточной стенкой и репликацией ДНК бактериального типа. Позднее одна из групп его потомков – первые археи – заменила эти предковые структуры другими при адаптации к жизни в экстремальных условиях, например в гидротермальных источниках. Это общепринятое объяснение, которое тем не менее малоубедительно. Почему репликация ДНК у бактерий и архей различается, а процессы транскрипции и трансляции схожи? Если мембраны и клеточные стенки архей суть результат адаптации к жизни в горячих источниках, то почему живущие в аналогичных условиях экстремофильные бактерии сохранили свои мембраны и клеточные стенки и не поменяли их на мембраны и клеточные стенки архейного типа или что-либо иное? Бактерии и археи сосуществуют в одних экологических нишах, но при этом, несмотря на ГПГ, сохраняют фундаментальные различия. Кажется неправдоподобным, что все эти глубокие различия возникли в результате адаптации к жизни в конкретной нише и что все без исключения археи с тех пор зафиксировали эти признаки, хотя в других экологических нишах они совершенно не нужны.
Нам остается принять лишь один вариант. В природе Последнего всеобщего предка нет ничего парадоксального: он действительно использовал энергию ионного градиента для синтеза АТФ, у него была АТФ-синтаза, но при этом не было ни мембран современного типа, ни больших дыхательных комплексов, которые в современных клетках перекачивают протоны. Он действительно обладал ДНК, универсальным генетическим кодом, транскрипцией, трансляцией и рибосомами, но современных механизмов репликации ДНК у него еще не было. Невозможно вообразить, как такая странная клетка могла существовать в открытом океане, но все становится на свои места, если мы поместим ее в щелочной гидротермальный источник. Мы можем понять, как существовал Последний всеобщий предок, изучив, как теперь живут в этих источниках бактерии и археи – те из них, которые используют путь Вуда – Льюнгдаля: вероятно, первичный путь фиксации углерода, чрезвычайно напоминающий геохимические реакции геотермальных источников.
Трудная дорога к Последнему всеобщему предку
В живом мире существует всего шесть путей фиксации углерода – превращения неорганических молекул CO2 в органические. Пять путей довольно сложны, и для их осуществления требуется много энергии – например энергии Солнца (в случае фотосинтеза). Есть еще одна причина, в силу которой Последний всеобщий предок не мог пользоваться фотосинтезом. Это цикл Кальвина – биохимический путь, в ходе которого CO2 преобразуется в сахара. Он встречается лишь у фотосинтетических бактерий (и растений, которые заполучили эти бактерии в качестве хлоропластов). Это означает, что цикл Кальвина вряд ли мог быть у Всеобщего предка. Если последний мог осуществлять фотосинтез, то эта способность должна была исчезнуть у всех архей – а терять такое полезное приспособление, конечно, глупо. Гораздо вероятнее, что цикл Кальвина появился позднее, одновременно с фотосинтезом, и лишь у бактерий. То же самое можно сказать об остальных путях фиксации углерода, за исключением одного. Единственный способ фиксации углерода, который встречается и у бактерий, и у архей – восстановительный путь ацетил-кофермента А (путь Вуда – Льюнгдаля).
Но и это утверждение не совсем верно. Путь Вуда – Льюнгдаля у бактерий и архей несколько отличается, и к этим различиям мы обратимся ниже. А сейчас рассмотрим, почему этот путь имеет хорошие шансы оказаться предковым, хотя данные филогенетики противоречивы и не могут подтвердить (впрочем, и опровергнуть) его раннее происхождение. Археи, живущие за счет пути Вуда – Льюнгдаля, – это метаногены, а бактерии – ацетогены. Одни деревья присваивают древнее происхождение метаногенам, другие – ацетогенам, а третьи указывают, что обе группы возникли довольно поздно, и их простота отражает скорее специализацию и оптимизацию, нежели близость к предковому состоянию. Сосредоточившись лишь на филогенетике, мы рискуем остаться ни с чем. Впрочем, никто нас и не заставляет ориентироваться только на нее.
Путь Вуда – Льюнгдаля начинается с H2 и CO2 – веществ, которые в изобилии имеются в щелочных гидротермальных источниках. Реакция CO2 и H2 с образованием органики экзергонична, то есть происходит с выделением энергии. Теоретически она должна быть самопроизвольной, но на практике существует энергетический барьер, из-за которого эта реакция протекает невероятно медленно. Метаногены используют протонные градиенты, чтобы преодолеть этот барьер. (Я постараюсь доказать, что этот прием был характерен для общего предка.) Как бы то ни было, метаногены и ацетогены растут лишь за счет реакции H2 и CO2: это их единственный источник углерода и энергии. Этим путь Вуда – Льюнгдаля выгодно отличается от остальных пяти способов фиксации углерода. Как заметил геохимик Эверетт Шок, “это бесплатный завтрак, за который тебе еще и платят”. Может, завтрак этот довольно скуден, зато в гидротермальных источниках его подают целый день.
И это еще не все плюсы. В отличие от других, путь Вуда – Льюнгдаля короткий и прямой. Он позволяет за небольшое число стадий перейти от простых неорганических молекул к веществу, в котором сосредоточен метаболизм клетки: маленькой и очень активной молекуле ацетил-кофермента А. Это универсальный “крючок”, который удерживает молекулы, чтобы они могли быть обработаны ферментом. Сам “крючок” не столь важен – важно то, что он держит: в данном случае это ацетильная группа. В слове “ацетил” тот же корень, что в слове “ацетат”. Ацетаты – соли уксусной кислоты, простой двухуглеродной молекулы, центральной в биохимии всех клеток. Будучи присоединенной к коферменту А, ацетильная группа находится в активированном состоянии (“активированный ацетат” – активированная уксусная кислота), что позволяет ей легко вступать в реакции с другими органическими молекулами и так участвовать в процессах биосинтеза.
Таким образом, в рамках пути Вуда – Льюнгдаля из CO2 и H2 за небольшое число шагов активированные молекулы синтезируются до тех пор, пока энергия не выделится в количестве, достаточном для того, чтобы обеспечить не только образование нуклеотидов и других молекул, но и их объединение в длинные цепи: ДНК, РНК, белки и т. д. В состав ферментов, катализирующих первые несколько стадий, входят неорганические кластеры, содержащие железо, никель и серу, которые участвуют в переносе электронов на CO2, в результате чего образуются активные ацетильные группы. Эти неорганические кластеры, по сути, являются минералами, более или менее идентичными по структуре железосерным минералам, которые осаждаются в гидротермальных источниках (рис. 11). Геохимия щелочных источников настолько сходна с биохимией метаногенов и ацетогенов, что язык не поворачивается назвать ее “аналогичной”. Аналогия подразумевает лишь поверхностное сходство, а здесь сходство настолько глубоко, что речь идет скорее о гомологии (когда одна форма дает начало другой). В ходе плавного превращения неорганики в органику из геохимии выросла биохимия. По словам Дэвида Гарнера, “неорганические вещества породили органические”.
Но, наверное, главное преимущество ацетил-кофермента А заключается в том, что он – перекресток путей углеродного и энергетического метаболизма. На важную роль ацетил-кофермента А в происхождении жизни указал в начале 90-х годов Кристиан де Дюв, правда, рассуждая о “первичном бульоне”, а не о щелочных источниках. Ацетил-кофермент А не только обеспечивает органический синтез, но и способен реагировать непосредственно с фосфатом с образованием ацетилфосфата. Хотя сейчас ацетилфосфат не так важен в энергетическом плане, как АТФ, он до сих пор широко используется живыми организмами и может выполнять почти все те же функции, что и АТФ. Как замечено выше, АТФ не просто высвобождает энергию: он осуществляет реакции дегидратации, в которых одна молекула воды отщепляется от двух аминокислот или других “строительных блоков”, скрепляя их в цепь. Проблема дегидратации аминокислот в растворе та же самая, что и в случае выжимания белья под водой, но именно этим и занимается АТФ. В лаборатории мы показали, что ацетилфосфат может выполнять ту же функцию, потому что его химические свойства в основе такие же. Это означает, что древний метаболизм углерода и энергии мог осуществляться за счет другого простого тиоэфира – ацетил-кофермента А.
Но так ли он прост? Двухуглеродная ацетильная группа, может, и простая, но кофермент А – сложная молекула, которая, несомненно, является продуктом естественного отбора, а значит, появилась довольно поздно. Впрочем, существуют простые “абиотические” эквиваленты ацетил-кофермента А. Высокая активность ацетил-кофермента А обусловлена тиоэфирной связью (между атомом серы и атомом углерода, который, в свою очередь, связан с кислородом):
R – S – CO – CH3
Здесь R – это радикал (остаток) молекулы, в данном случае кофермент А. CH3 – метильная группа. Но в качестве R не обязательно выступает кофермент А. Вместо него может быть какая-нибудь простая группа, например метильная (CH3). Тогда получится небольшая молекула – метилтиоацетат:
CH3–S – CO – CH3
Это активный тиоэфир, аналогичный по химическим свойствам ацетил-коферменту А, но достаточно простой, чтобы его можно было получить в результате реакции H2 и CO2 в гидротермальных источниках – и действительно, он был получен Клаудией Хубер и Гюнтером Вехтерсхойзером, в качестве исходных веществ взявших лишь CO и CH3SH. Более того, метилтиоацетат, как и ацетил-кофермент А, должен быть способен непосредственно реагировать с фосфатом, образуя ацетилфосфат. Таким образом, этот активный эфир через образование ацетилфосфата теоретически мог обеспечивать синтез новых органических молекул, а также их полимеризацию с образованием сложных цепей, например белков и РНК. Эту гипотезу мы сейчас проверяем при помощи настольного реактора (нам действительно удалось получить ацетилфосфат, хотя и в низкой концентрации).
Внутри микропор щелочных гидротермальных источников примитивный вариант пути Вуда – Льюнгдаля, в принципе, может дать все, что нужно для возникновения примитивных клеток. Я предположил бы три стадии. На первой стадии протонные градиенты на тонких неорганических перегородках, содержащих каталитические железосерные минералы, обусловили образование небольших органических молекул (рис. 14). Органические вещества концентрировались внутри более прохладных пор посредством термофореза и, в свою очередь, улучшали каталитические свойства минералов (гл. 3). Так зародилась биохимия: непрерывное образование и концентрирование активированных предшественников вынуждало молекулы реагировать друг с другом и формировать простые полимеры.
Вторая стадия: формирование простейших органических протоклеток внутри пор источника как естественный исход физического взаимодействия органических веществ. Это были простые клеткоподобные структуры, образующиеся в результате самоорганизации материи, пока не обладающие ни каким-либо генетическим базисом, ни реальной сложностью. Я думаю, что эти простые клетки осуществляли органический синтез за счет протонного градиента, но теперь на собственных органических мембранах (так, липидные бислои образуются самопроизвольно из жирных кислот), а не на неорганических стенках источника. Для этого не нужны белки. Протонный градиент может способствовать образованию метилтиоацетата и ацетилфосфата и обеспечивать углеродный и энергетический метаболизм. На этой стадии возникает ключевое отличие: отныне органическая материя формируется внутри самой клетки и этим процессом движут естественные протонные градиенты на органических мембранах. Мне нравится слово “движут”: думаю, оно подходит по смыслу лучше всех остальных. Оно позволяет подчеркнуть, что это не пассивные химические процессы, а вынужденные – инициированные непрерывным потоком углерода, энергии, протонов. Этим реакциям нужно произойти: это единственный способ разрешить нестабильное состояние, в котором пребывают восстановленные, насыщенные водородом щелочные жидкости, попадающие в окисленные, кислые, богатые ионами металлов океанские воды. Единственный путь достигнуть желанного термодинамического равновесия.
Третья стадия – возникновение генетического кода и настоящей наследственности. Это позволило протоклеткам более или менее точно воспроизводить себя. Наиболее ранние формы отбора, основанные на соотношении скоростей синтеза и распада, открыли путь настоящему естественному отбору, в ходе которого популяции протоклеток с генами и белками начали бороться за существование в порах источников. В конце концов в результате действия стандартных механизмов эволюции в ранних клетках появились белки со сложным строением, в том числе рибосомы и АТФ-синтаза (эти белки сохранились у всех организмов). Мне представляется, что Последний всеобщий предок, прародитель и бактерий, и архей, обитал в микропорах щелочных гидротермальных источников. Это значит, все три стадии, от абиотического начала и до Последнего всеобщего предка, протекали внутри пор источников. Все они обеспечивались протонными градиентами на неорганических стенках или органических мембранах. А появление сложных белков, например АТФ-синтазы – это последний шаг на каменистом пути к Последнему всеобщему предку.
В этой книге мы не рассматриваем детали древней биохимии – откуда взялся генетический код и прочие столь же сложные проблемы. Это по-настоящему важные вопросы, и ими занимаются некоторые выдающиеся исследователи. Мы до сих пор не знаем точных ответов на эти вопросы, но все идеи на эту тему предполагают обильное поступление активированных предшественников. Вот, например, красивая догадка о происхождении генетического кода, предложенная Шелли Копли, Эриком Смитом и Гарольдом Моровицем: каталитические динуклеотиды (два соединенных вместе нуклеотида) могут инициировать образование аминокислот из более простых предшественников, например пирувата. Эта остроумная схема показывает, как генетический код мог возникнуть из детерминистической химии. Для тех, кому это интересно, см. главу о происхождении ДНК в моей книге “Лестница жизни”. Но все эти гипотезы исходят из наличия постоянного снабжения нуклеотидами, пируватом и прочими предшественниками. Вопрос таков: какие движущие силы вызвали возникновение жизни на Земле? Главное, что мне хотелось бы подчеркнуть – отсутствие принципиальной сложности насчет того, откуда могли взяться углерод, энергия и катализаторы, обеспечившие образование сложных биологических молекул и дальнейшее развитие вплоть до появления генов, белков и Последнего всеобщего предка.
Рассмотренный сценарий с участием щелочных источников хорошо сочетается с биохимией метаногенов – архей, которые живут за счет H2 и CO2 и пути Вуда – Льюнгдаля. Эти клетки (вероятно, древние) генерируют на мембране протонные градиенты (мы рассмотрим, как именно), производя то же самое, что щелочные гидротермальные источники предоставляют даром. Протонный градиент обеспечивает прохождение пути Вуда – Льюнгдаля, и в этом участвует встроенный в мембрану железосерный белок – энергопреобразующая гидрогеназа (Ech). Он перебрасывает протоны через мембрану на другой железосерный белок – ферредоксин. Тот, в свою очередь, восстанавливает CO2. Выше я предположил, что в щелочных источниках естественные протонные градиенты на тонких стенках из сульфида железа сделали возможным восстановление CO2 за счет изменения восстановительных потенциалов H2 и CO2. Полагаю, что в энергопреобразующей гидрогеназе происходят те же самые процессы, только в нанометровом масштабе. Ферменты способны поддерживать определенные физические условия (в том числе концентрацию протонов) внутри своих полостей шириной в несколько ангстрем, и энергопреобразующая гидрогеназа могла обладать такими свойствами. Если так, то может существовать непрерывный переход от древнего состояния, в котором полипептиды стабилизируются, связываясь с железосерными минералами, встроенными в мембраны протоклеток, – и современным состоянием, в котором генетически закодированная энергопреобразующая гидрогеназа обеспечивает энергией метаболизм углерода у современных метаногенов.
Как бы то ни было, в мире генов и белков энергопреобразующая гидрогеназа использует протонный градиент, созданный за счет энергии, выделившейся в ходе синтеза метана, чтобы осуществить восстановление CO2. А метаногены пользуются протонным градиентом для непосредственного синтеза АТФ с помощью АТФ-синтазы. Таким образом, и углеродный, и энергетический метаболизм осуществляются за счет протонных градиентов, точно таких же, как в источниках (а источники предоставляли их бесплатно). Возможно, именно так был устроен углеродный и энергетический метаболизм у первых протоклеток, обитавших в щелочных источниках. Это звучит достаточно правдоподобно, но на самом деле, если полагаться на естественные градиенты, возникают некоторые проблемы, очень серьезные и увлекательные. Мы с Биллом Мартином поняли, что может быть лишь одно возможное решение – и оно сулит многое в понимании причин фундаментальных различий бактерий и архей.
Проблема проницаемости мембраны
Внутренние мембраны наших собственных митохондрий почти непроницаемы для протонов. Это необходимое свойство. Какой смысл перекачивать протоны через мембрану, если они тут же устремляются обратно, будто бы сквозь бесчисленные дырочки? С тем же успехом можно носить воду в решете. В митохондриях находится электрическая цепь, в которой мембрана выступает диэлектриком. Протоны перекачиваются через мембрану, и большинство их возвращается, проходя сквозь белки, которые функционируют как турбины. В случае АТФ-синтазы синтез АТФ происходит за счет прохождения протонов сквозь крошечный вращающийся мотор. Но заметьте: система основана на активном перекачивании. Если заблокировать помпы, все процессы в клетке остановятся – как если стихнет ветер и на мельнице зерно перестанет перемалываться в муку. Это и происходит при отравлении цианидом: он блокирует протонную помпу на конце дыхательной цепи митохондрий. Если работа помп в составе дыхательной цепи затруднена, протоны могут продолжить втекать через АТФ-синтазу несколько секунд до того, как концентрация протонов не станет равной по обе стороны мембраны и ток прекратится. Трудно дать смерти столь же четкое определение, как жизни, но безвозвратная потеря мембранного потенциала приближается к таковому.
Как энергия естественного градиента протонов могла осуществлять синтез АТФ? Это сопряжено с “цианидной проблемой”. Представьте себе протоклетку внутри поры источника, которая получает энергию за счет протонного градиента. Одна сторона клетки постоянно омывается океанской водой, другая – потоками щелочных гидротермальных жидкостей (рис. 17). Четыре миллиарда лет назад океаны, вероятно, были слабокислыми (pH = 5–7), а гидротермальные жидкости – такими же, как сейчас (pH = 9–11). Получается, перепады pH были довольно резкими, в 3–5 единиц. Иначе говоря, разница концентраций протонов внутри и снаружи источника могла различаться в 1–100 тыс. раз. Представим, что концентрация протонов внутри клетки совпадает с таковой для гидротермальных жидкостей. Это значит, что концентрация протонов внутри и снаружи клетки различается и протоны будут стремиться внутрь по градиенту концентрации. Впрочем, за несколько секунд поток иссякает и не возникнет снова, пока протоны не будут удалены из клетки. Этому есть две причины. Во-первых, концентрация по обе стороны мембраны быстро уравнивается. Во-вторых, важную роль играет электрический заряд. Протоны (H+) заряжены положительно, но в морской воде их заряд компенсируется отрицательно заряженными атомами, например ионами хлора (Cl–). Проблема в том, что протоны проходят через мембрану гораздо быстрее, чем ионы хлора, и приток отрицательно заряженных частиц не компенсирует приток положительно заряженных. Таким образом, внутренняя сторона мембраны относительно внешней среды становится заряженной положительно, и это препятствует дальнейшему притоку H+. Итак, без насоса, который позволил бы выкачивать протоны из клетки наружу, естественные протонные градиенты бесполезны. Они быстро приходят в равновесие, а равновесие эквивалентно смерти.

Рис. 17. Клетка, живущая за счет природного протонного градиента.
В центре изображена клетка с проницаемой для протонов мембраной. Клетка втиснута в расщелину в неорганическом барьере, который внутри микропористого источника разделяет две фазы жидкостей. Верхняя фаза – это слабокислая океанская вода с pH = 5–7 (в рамках этой модели ее pH принят за 7), которая просачивается внутрь глубокой поры. В нижнюю фазу входят щелочные гидротермальные жидкости, которые просачиваются из одной поры в другую через соединительные каналы (их pH примерно 10). Ламинарное течение предполагает отсутствие турбулентных завихрений и смешивания, характерных для жидкостей, протекающих сквозь небольшие ограниченные пространства. Протоны (H+) могут проходить прямо сквозь липидную мембрану или через встроенные в нее белки (на рисунке обозначены треугольной фигурой) по градиенту концентрации протонов, меняющемуся от кислой воды океана до щелочных гидротермальных потоков. Гидроксид-ионы (OH–) текут в противоположном направлении, из гидротермальных жидкостей в океанскую воду, но исключительно через мембрану. Интенсивность потока протонов зависит от проницаемости мембраны для H+, нейтрализации OH–(с образованием H2O), количества мембранных белков, размера клетки и заряда на мембране, накопленного в результате перемещения ионов из одной фазы в другую.
Но есть исключение. Если мембрана почти непроницаема для протонов, они не смогут попасть внутрь. Протоны входят в клетку, но не могут выйти. Но если мембрана в высокой степени проницаема, тогда другое дело. Протоны и в этом случае попадают внутрь клетки, но теперь они могут покидать ее, хоть и пассивно, через проницаемую мембрану на другой стороне клетки. В результате проницаемая мембрана представляет собой менее значительное препятствие для потока. Более того, гидроксид-ионы (OH–) из гидротермальных жидкостей проходят через мембрану примерно с той же скоростью, что и протоны. Встречаясь друг с другом, H+ и OH– реагируют с образованием воды (H2O), при этом положительный заряд протонов ликвидируется. При помощи классических электрохимических уравнений можно рассчитать скорости входа и выхода протонов из гипотетической клетки (ее компьютерной модели) как функции от мембранной проницаемости. Именно это и сделал Виктор Сохо, химик, интересующийся масштабными биологическими проблемами и работающий над диссертацией вместе со мной и Эндрю Помянковски. Зная соотношение равновесных концентраций протонов, мы можем рассчитать свободную энергию (ΔG), доступную лишь за счет протонного градиента. Доступная движущая сила зависит от проницаемости мембраны для протонов. Если мембрана слишком проницаема, протоны устремляются внутрь, а затем быстро исчезают, нейтрализованные стремительным потоком ионов OH–. Но мы обнаружили, что и при очень высокой проницаемости мембраны протоны проходят через мембранные белки (например через АТФ-синтазу) быстрее, чем между липидами самой мембраны. Это означает, что за счет потока протонов может происходить синтез АТФ или восстановление углерода энергопреобразующей гидрогеназой. Рассмотрев разницу концентраций и заряд, а также особенности работы АТФ-синтазы и других белков, мы показали, что лишь клетки с высокопроницаемыми мембранами могут использовать протонные градиенты для углеродного и энергетического метаболизма. Примечательно, что такие клетки теоретически могут получить столько же энергии за счет протонного градиента в три единицы pH, сколько современные клетки получают в ходе дыхания.
Они могут получать и гораздо больше. Большую долю времени метаногены, как следует из названия, занимаются тем, что производят метан. В среднем метаногены вырабатывают примерно в 40 раз больше отходов (метана и воды), чем органических веществ. Вся энергия, полученная в ходе синтеза метана, идет на перекачивание протонов (рис. 18). Так-то! Метаногены тратят почти 98 % своего энергетического бюджета на создание протонных градиентов при помощи метаногенеза – и менее 2 % на образование новой органической материи. С естественными протонными градиентами и проницаемыми мембранами такие затраты энергии не требуются. Объем доступной энергии при этом почти такой же, но дополнительный расход уменьшается примерно в 40 раз: весьма существенное преимущество! Только представьте: в 40 раз больше энергии! Даже мои маленькие сыновья не превосходят меня настолько. Выше я отметил, что примитивным клеткам должно было требоваться больше углерода и энергии, чем современным. Отсутствие необходимости перекачивать протоны предоставляет гораздо больше углерода и энергии.
Рассмотрим проницаемую клетку, находящуюся в среде с естественным градиентом протонов. Помните, что мы живем в эпоху генов и белков, которые сами являются продуктом естественного отбора, действовавшего на протоклетки. Наша клетка с проницаемыми мембранами может использовать непрерывный поток протонов, чтобы обеспечивать метаболизм углерода при помощи энергопреобразующей гидрогеназы. Этот белок позволяет клетке осуществлять реакцию H2 и CO2 с образованием ацетил-кофермента А, а следом и всех “строительных блоков” жизни. Клетка также может использовать протонный градиент, чтобы осуществлять синтез АТФ при помощи АТФ-синтазы. И, конечно, она может использовать АТФ, чтобы полимеризовать аминокислоты и нуклеотиды с образованием новых белков, РНК и ДНК и, наконец, собственных копий. Важно, что нашим проницаемым клеткам не надо тратить энергию на перекачивание протонов, а значит, они должны хорошо расти, несмотря на слабую эффективность древних ферментов.
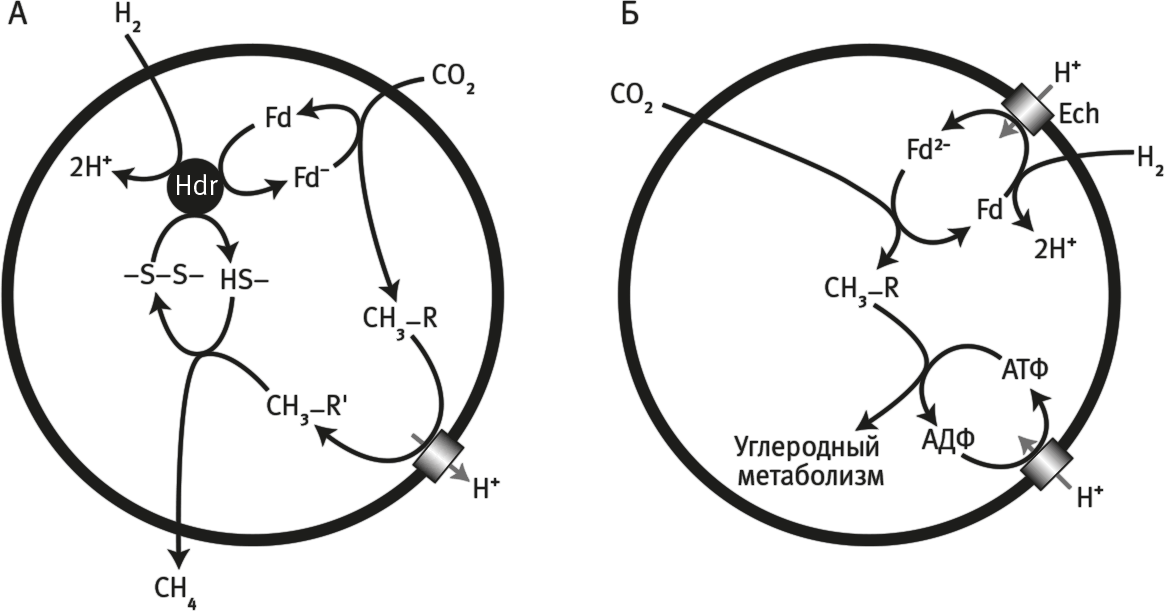
Рис. 18. Получение энергии в ходе образования метана.
А. Энергия, которая выделяется в реакции между H2 и CO2, расходуется на транспорт протонов сквозь клеточную мембрану. Фермент гидрогеназа (Hdr) катализирует одновременное восстановление ферредоксина (Fd) и дисульфидной связи (–S – S–, используя два электрона H2. Ферредоксин, в свою очередь, восстанавливает CO2 до метильной группы (–H3), которая связывается с кофактором (R). Метильная группа затем перемещается на второй кофактор (R'), и в процессе выделяется энергия в количестве, достаточном для перекачивания через мембрану двух H+ (или Na+). На финальном этапе группа CH3− восстанавливается группой HS– до метана (CH4). В итоге некоторая часть энергии, выделившейся из H2 и CO2 в ходе образования метана (CH4), сохраняется на клеточной мембране в виде градиента H+ (или Na+).
Б. Протонный градиент непосредственно используется двумя особыми мембранными белками, которые направляют энергию градиента на углеродный и энергетический метаболизм. Энергопреобразующая гидрогеназа (Ech) прицельно восстанавливает ферредоксин (Fd), который снова отдает электроны CO2. Образуется метильная группа (–H3), которая реагирует с CO. Складывается путь Вуда – Льюнгдаля. Подобным образом через АТФ-синтазу благодаря потоку H+ осуществляется синтез АТФ и, тем самым, энергетический обмен.
Однако такие проницаемые клетки привязаны к месту, и их выживание целиком зависит от гидротермальных потоков. Если поток иссякнет или сместится, они обречены. Хуже того: в этих условиях они не могут эволюционировать. Не имеет смысла улучшать параметры мембраны: напротив, если мембрана станет малопроницаемой, протонный градиент быстро нарушится – при условии, что не будет никакого способа избавиться от протонов внутри клетки. Таким образом, клетки, которые приобрели более “современные” непроницаемые мембраны, в ходе естественного отбора должны элиминироваться. Конечно, лишь до тех пор, пока они не научатся перекачивать протоны, но это не менее проблематично – мы уже знаем, что нет смысла перекачивать протоны через проницаемую мембрану. Наше исследование подтверждает, что перекачивание протонов остается невыгодным, даже если уменьшить проницаемость мембраны на целых три порядка.
Разберем это подробнее. У проницаемой клетки в среде с протонным градиентом есть доступ к большому количеству энергии, которого достаточно для того, чтобы обеспечить ее углеродный и энергетический метаболизм. Если в результате некоего эволюционного фокуса полностью функциональный насос будет помещен в мембрану, это не будет иметь смысла: энергии с насосом будет столько же, сколько без насоса. (Перекачивание протонов сквозь проницаемую мембрану бессмысленно – они возвращаются обратно.) Попробуем уменьшить проницаемость мембраны в десять раз: энергии не становится больше. А в сто раз? То же самое. В тысячу? Бесполезно. Почему? Уменьшая проницаемость мембраны, мы получаем возможность перекачивать протоны эффективнее, но при этом нарушается их природный градиент, и энергию становится неоткуда брать. Перекачивать протоны выгодно лишь тогда, когда в мембране много насосов, а сама мембрана почти непроницаема. И это серьезная проблема: ни насосы, ни современные малопроницаемые мембраны сами по себе не давали никаких преимуществ, а значит, не могли закрепиться. Однако они существуют. Так что мы упускаем из виду?
Сейчас я расскажу историю, которая служит прекрасным примером того, как действует научная интуиция. Мы с Биллом Мартином размышляли над этим вопросом, и вдруг нас осенило: метаногены используют белок-антипортер. Эти метаногены выкачивают ионы натрия (Na+), а не протоны (H+), но все равно у них есть проблемы, связанные с тем, что протоны накапливаются внутри. Антипортер обменивает Na+ на H+, работая по принципу двустороннего турникета или карусельной двери. На каждый Na+, который входит в клетку, приходится один выходящий H+. Это протонная помпа, работающая за счет градиента натрия. Однако антипортеры неразборчивы: им все равно, в каком направлении работать. Если градиент H+ превышает градиент Na+, антипортер может легко начать работать в обратном направлении: при вхождении в клетку одного H+ один Na+ будет выходить наружу. Ага! Если бы наша клетка с проницаемой мембраной, сидящая в гидротермальном источнике, приобрела антипортер Na+/H+, он функционировал бы как Na+-помпа, работающая за счет протонов! На каждый H+, входящий в клетку через антипортер, один Na+ должен был ее покидать. Теоретически антипортер мог преобразовывать природный протонный градиент в биохимический градиент натрия. Но что именно это нам дает? Я должен подчеркнуть, что это мысленный эксперимент на основе известных свойств белка. Но, согласно нашим расчетам, их роль может быть удивительно важна. Как правило, липидные мембраны примерно на шесть порядков менее проницаемы для Na+, чем для H+. Мембрана, высокопроницаемая для протонов, почти не пропускает ионы натрия. Выкачайте протон – и он тут же вернется. Выкачайте ион натрия из той же самой клетки – и вы еще долго его не увидите. Это значит, что антипортер может работать за счет природного градиента протонов: на каждый H+, входящий в клетку, один Na+ будет выталкиваться. Пока мембрана остается проницаемой для протонов, как и раньше – поток протонов через антипортер не будет утихать, способствуя выходу Na+. Так как мембрана менее проницаема для Na+, выброшенный Na+ скорее останется снаружи. Вернее, он будет заходить обратно в клетку через мембранные белки, а не возвращаться прямо через мембрану, просачиваясь между липидами. Это означает, что энергия потока ионов Na+ может быть использована для выполнения полезной работы.
Конечно, это возможно лишь в том случае, если мембранные белки, обеспечивающие углеродный и энергетический метаболизм – энергопреобразующая гидрогеназа и АТФ-синтаза, – не делают различия между Na+ и H+. Звучит абсурдно, но вполне может быть правдой. Оказывается, АТФ-синтаза некоторых метаногенов может почти одинаково легко использовать H+ и Na+. Даже на сухом языке химии они получили наименование promiscuous, “неразборчивые”. Скорее всего, причина кроется в одинаковом заряде и очень близком радиусе этих двух ионов. Хотя H+ по размеру значительно меньше, чем Na+, протоны редко пребывают в изоляции. Переходя в раствор, они формируют связи с водой с образованием H3O+. Радиус этого иона почти соответствует радиусу Na+. Другие мембранные белки, включая энергопреобразующую гидрогеназу, также неразборчивы в отношении H+ и Na+ – вероятно, по тем же причинам. Самое главное, что перекачивание Na+ становится отнюдь не бесполезным. Если использовать природные протонные градиенты, выкачивание Na+ становится по сути “бесплатным”. При наличии градиента натрия большая часть ионов Na+ входит в клетку сквозь мембранные белки, например энергопреобразующую гидрогеназу и АТФ-синтазу, а не непосредственно сквозь липидный слой мембраны. Теперь мембрана лучше изолирована, а значит, вероятность “короткого замыкания” гораздо ниже. Теперь для обеспечения углеродного и энергетического метаболизма можно использовать большее число ионов, а каждый ион выкачивать с большей выгодой.
У этого простого приобретения несколько неожиданных и своеобразных эффектов. Один очевиден: выкачивание ионов натрия наружу клетки уменьшает их концентрацию внутри клетки. Известно, что многие базовые ферменты, имеющиеся и у бактерий, и у архей (например ферменты, ответственные за транскрипцию и трансляцию), в результате отбора приобрели строение, оптимальное для работы при низкой концентрации Na+ – хотя их эволюция, скорее всего, протекала в океане, где концентрация Na+ была высокой даже 4 млрд лет назад. Раннее возникновение антипортера может объяснять, почему все клетки приспособлены к низкой концентрации натрия, хотя их развитие происходило в среде с высокой его концентрацией.
Пока для нас важнее вот что: антипортер дополняет существующий протонный градиент градиентом Na+. Клетка, как и прежде, получает энергию благодаря природному градиенту H+, и, соответственно, ее мембрана должна оставаться проницаемой для протонов. Но теперь у нее появился еще и градиент натрия, который, согласно нашим расчетам, позволяет получать на 60 % больше энергии, чем прежде, когда использовался лишь протонный градиент. Это дает клеткам большое преимущество. Во-первых, клетка с антипортером получает больше энергии, а значит, способна расти и размножаться быстрее, чем клетки без антипортера. Во-вторых, такие клетки могут существовать и при менее выраженных протонных градиентах. В нашем опыте клетки с проницаемыми мембранами хорошо росли, когда перепад концентраций протонного градиента был равен примерно трем единицам pH, а ведь это и соответствует разнице pH кислой воды океана (pH около 7) и щелочных гидротермальных жидкостей (pH примерно 10). (То есть концентрация протонов в океанской воде на три порядка выше, чем в щелочных потоках источника.) За счет более эффективного использования естественного протонного градиента клетки с антипортером способны выжить, даже если градиент pH менее двух единиц. Наличие антипортера дает им преимущество в конкурентной борьбе с другими клетками, позволяя расселяться по “родному” гидротермальному источнику и захватывать соседние источники, если они объединены в сеть. Но из-за того, что клетки по-прежнему полностью зависимы от природного протонного градиента, они не могут покинуть источник. Для этого требуется еще один шаг.
Хотя клетки с антипортером пока не способны удалиться от источника, в некоторой степени они к этому подготовлены. Антипортер – это “преадаптация”, необходимый первый шаг, который способствует дальнейшему эволюционному развитию. Неожиданное решение! По крайней мере, я удивился. Наличие антипортера впервые порождает условия для активного перекачивания. Я уже упоминал, что нет смысла перекачивать протоны через проницаемую мембрану: они будут возвращаться. Но если у вас есть антипортер, это становится выгодным. Будучи выброшены наружу, некоторые протоны просачиваются внутрь не сквозь мембрану, а через антипортер, заставляя натрий выходить из клетки. Так как мембрана пропускает натрий хуже, большая часть энергии, затраченной на то, чтобы выкачать протоны, сохраняется в виде ионного градиента на мембране. Шанс, что выброшенный протон так и останется снаружи, немного увеличивается. А это означает, что, выкачивая протоны, вы получаете преимущество, пусть и небольшое, тогда как раньше это не приносило вообще никакой пользы. Выкачивание обретает смысл только при наличии антипортера.
И это не все. Когда возникла протонная помпа (насос), стало выгодно изменить параметры мембраны. В условиях природного протонного градиента абсолютно необходимо иметь проницаемую для протонов мембрану. В то же время перекачивать протоны через мембрану абсолютно нет смысла. Антипортер отчасти исправляет положение: благодаря нему из природного протонного градиента можно извлечь больше энергии. Однако антипортер не в полной мере делает клетку независимой от этого градиента. Но все же, имея антипортер, можно выкачивать протоны с пользой, а значит, зависимость от природного градиента снижается. И теперь – лишь теперь! – становится выгодно иметь в меньшей степени проницаемую мембрану. Небольшое снижение проницаемости делает перекачивание протонов чуть выгоднее. Если изменить ее еще немного, то преимущество, соответственно, еще вырастет. И так далее. Впервые мы имеем устойчивую селективную движущую силу, одновременно способствующую эволюции и протонных помп, и современных липидных мембран. Так клетки приобрели возможность оборвать пуповину, связывающую их с природными протонными градиентами: они стали вольны покинуть источники, чтобы найти свое место в огромном, пока еще пустом мире.
В отличие от филогенетики, которая мало что может сказать с высокой степенью достоверности, эти физические ограничения позволяют нам указать точную последовательность эволюционных шагов, начиная с зависимости от природных протонных градиентов и заканчивая, по сути, современными клетками, которые создают собственные протонные градиенты на непроницаемых мембранах (рис. 19). Более того, теперь можно объяснить глубинные различия между бактериями и археями. И те, и другие производят АТФ, используя протонные градиенты на мембранах, хотя мембраны у представителей этих доменов сильно отличаются. Так же дело обстоит и с другими чертами, включая сами мембранные насосы, клеточную стенку и механизм репликации ДНК.
О причинах фундаментальных отличий бактерий и архей
Мы рассмотрели вероятные условия зарождения жизни на Земле с точки зрения энергетики. Затем сузили область условий и остановились на щелочных гидротермальных источниках, в которых постоянные потоки углерода и энергии сочетаются с естественной компартментализацией и присутствием каталитических минералов. Но есть проблема: источник энергии и углерода в этих источниках – CO2 и H2, а их нелегко заставить реагировать друг с другом. Как мы видели, энергетический барьер этой реакции потенциально возможно преодолеть за счет энергии геохимических протонных градиентов на тонких полупроводниковых перегородках в порах гидротермальных источников. За счет образования реакционноспособных тиоэфиров, например метилтиоацетата (функциональный аналог ацетил-кофермента А), протонные градиенты могли стимулировать развитие путей метаболизма углерода и энергии, что привело к накоплению в порах источника органических молекул, а облегчение реакций “дегидратации” способствовало образованию ДНК, РНК, белков и других сложных полимеров. Уклонившись от рассмотрения таких подробностей, как возникновение генетического кода, я ограничился тем, что в этих условиях теоретически возможно образование примитивных клеток с белками, закодированными в генах. В популяции клеток возможен настоящий естественный отбор, и я предположил, что Последний всеобщий предок – прародитель архей и бактерий – мог быть результатом отбора в популяциях таких примитивных клеток, обитающих в порах щелочных гидротермальных источников и зависящих от естественных протонных градиентов. Под воздействием отбора появилось множество замысловатых белков, в том числе рибосомы, энергопреобразующей гидрогеназы и АТФ-синтазы: они универсальны для живых организмов и консервативны.
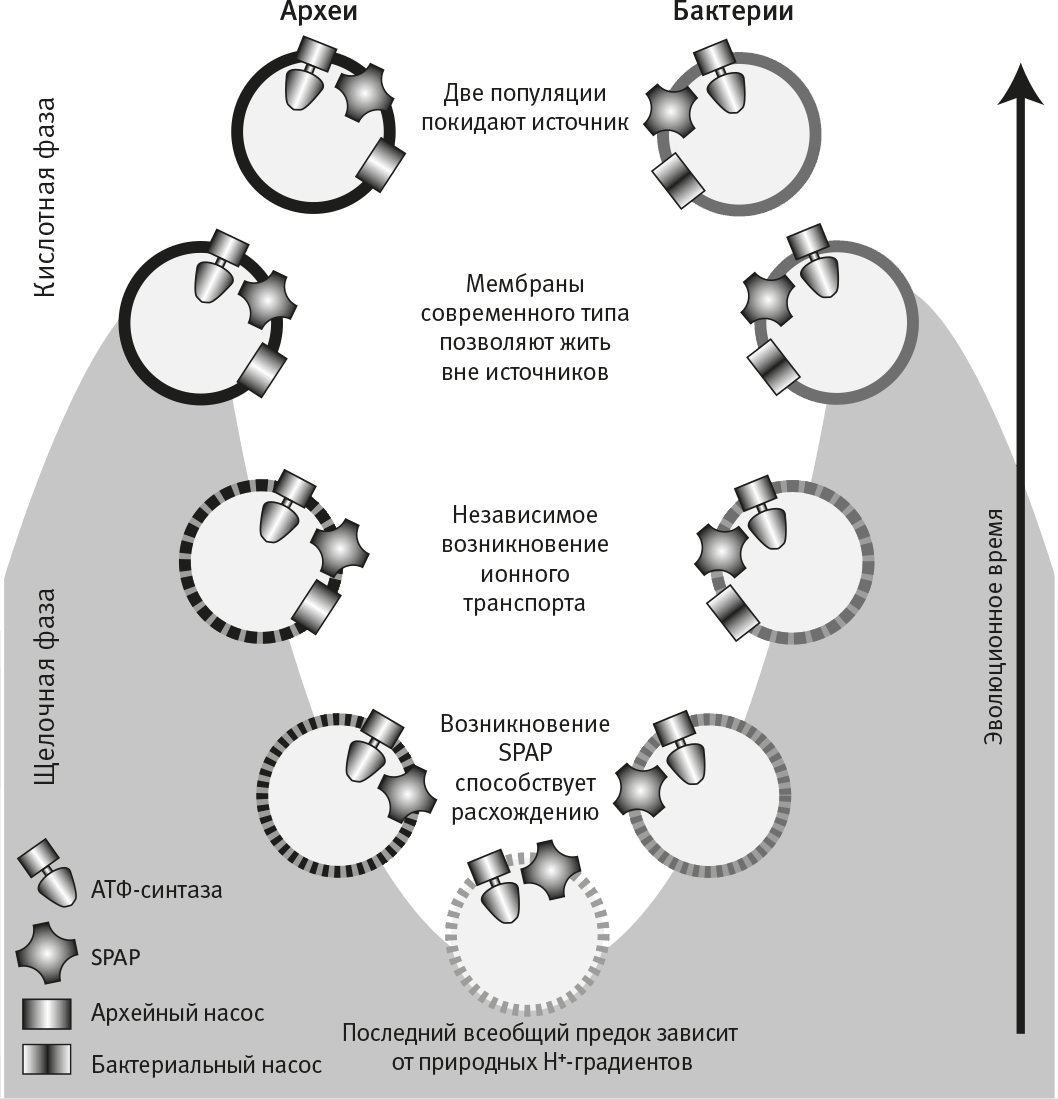
Рис. 19. Происхождение бактерий и архей.
Возможный сценарий расхождения бактерий и архей, который опирается на математическую модель доступности энергии в условиях естественных протонных градиентов. Для простоты на рисунке изображена лишь АТФ-синтаза, но тот же принцип действует в отношении других мембранных белков, таких как энергопреобразующая гидрогеназа (Ech). Естественный градиент протонов в источнике может обеспечивать синтез АТФ до тех пор, пока мембрана остается “протекающей”, так что не имеет смысла делать мембрану непроницаемой, потому что это лишит клетку возможности использовать естественный градиент (внизу). Натрий-протонный антипортер (SPAP) дополняет геохимический протонный градиент биохимическим градиентом натрия, что позволяет клетке выживать при менее выраженных градиентах, а это способствует распространению и дивергенции популяций клеток внутри источника. При наличии SPAP выкачивать H+ становится выгодно. С таким насосом, в свою очередь, становится выгодно делать мембрану менее проницаемой для H+. Когда степень мембранной проницаемости для протонов приближается к современному уровню, клетки становятся независимыми от естественных градиентов и могут покинуть источник. Бактерии и археи изображены покидающими источник независимо друг от друга.
В принципе Последний всеобщий предок для обеспечения энергетического обмена и метаболизма углерода мог обойтись естественными протонными градиентами, АТФ-синтазой и энергопреобразующей гидрогеназой, но для этого ему понадобились бы мембраны с очень высокой проницаемостью. В таких условиях Последний всеобщий предок не смог бы приобрести непроницаемые мембраны современного типа, как у бактерий или архей, поскольку это привело бы к исчезновению естественного протонного градиента. Но вот антипортер, преобразовывающий естественные протонные градиенты в биохимические градиенты натрия, мог оказаться полезен: он увеличивает эффективность использования энергии, а это позволяет клеткам выживать при более низких значениях протонных градиентов. Такие клетки получили возможность распространяться и заселять непригодные для остальных клеток участки гидротермальных источников, что, в свою очередь, способствовало образованию разрозненных популяций. Получив способность выживать в широком диапазоне условий, клетки смогли “заражать” соседние источники и расселяться по всему древнему дну, где в то время было много очагов серпентинизации.
Антипортер, кроме прочего, впервые сделал выгодным активный транспорт ионов. Так мы приходим к различиям пути Вуда – Льюнгдаля для метаногенов и ацетогенов. Судя по этим различиям, активный транспорт ионов независимо возник в двух разных популяциях клеток, отделившихся от общей предковой популяции за счет приобретения антипортера. Напомню, что метаногены и ацетогены принадлежат к двум разным доменам прокариот: археям и бактериям, то есть к двум самым древним ветвям “дерева жизни”. У бактерий и архей очень схожи системы транскрипции и трансляции, рибосомы, синтез белков, но чрезвычайно сильно различаются такие фундаментальные структуры, как клеточные мембраны. Я отметил, что они также различаются деталями пути Вуда – Льюнгдаля, хотя он является для них предковым признаком. Отличие и сходство этого пути могут рассказать очень о многом.
Как и метаногены, ацетогены производят ацетил-кофермент А за счет реакции H2 и CO2 через серию аналогичных последовательных реакций. И метаногены, и ацетогены для того, чтобы обеспечить энергией систему активного транспорта ионов, прибегают к бифуркации (разветвлению) потока электронов. Этот механизм совсем недавно открыт немецким микробиологом Рольфом Тауэром и его коллегами, и это, пожалуй, самый крупный прорыв в биоэнергетике за последние десятилетия. Формально Тауэр уже на пенсии, но результаты его исследований пролили свет на загадку биоэнергетики, над которой ученые десятилетиями ломали головы: как загадочным микробам удается расти, когда это противоречит всем стехиометрическим расчетам. Эволюция, как часто бывает, оказалась хитрее нас. Бифуркацию потока электронов можно сравнить с краткосрочным энергетическим “кредитом”. Как мы отметили, реакция H2 с CO2 суммарно экзергонична (протекает с выделением энергии), но первые ее стадии эндергоничны (требуют затраты энергии). Бифуркация потока электронов – хитроумный механизм, который позволяет направить часть энергии, выделяющейся на поздних, экзергонических стадиях восстановления CO2, на выплату “энергетической задолженности” за протекание ранних, эндергонических стадий. Так как на поздних стадиях выделяется больше энергии, чем требуется для протекания ранних стадий, часть этой энергии может запасаться в форме протонного градиента на мембране (рис. 18). Суммарный результат всех этих процессов – перенос протонов через мембрану за счет энергии, выделившейся в ходе реакции H2 и CO2.
Загадка кроется в различии путей бифуркации электронов у метаногенов и ацетогенов. Хотя и те, и другие используют весьма сходные железо-никелево-серные белки, многие другие белки и детали механизма бифуркации электронов у них различаются. Как и метаногены, ацетогены запасают энергию, выделяющуюся при реакции H2 с CO2, в форме градиента H+ или Na+ на мембране. В обоих случаях этот градиент расходуется на поддержание углеродного и энергетического метаболизма. И у метаногенов, у ацетогенов есть АТФ-синтаза и энергопреобразующая гидрогеназа, но, в отличие от метаногенов, ацетогены не используют энергопреобразующую гидрогеназу для непосредственного поддержания углеродного метаболизма. Напротив, у некоторых ацетогенов энергопреобразующая гидрогеназа работает в обратном направлении. Ацетогены и метаногены сильно различаются в деталях углеродного метаболизма, и эти детали кажутся настолько фундаментальными, что некоторые специалисты утверждают даже, что сходство метаногенов и ацетогенов есть следствие конвергентной эволюции или горизонтального переноса генов, а не происхождения от общего предка.
Сходство и различия метаногенов и ацетогенов складываются в единую картину, если допустить, что Последний всеобщий предок зависел от природных протонных градиентов. В этом случае ключ к пониманию различий в механизмах создания мембранных градиентов у метаногенов и ацетогенов – это направление естественного потока протонов через энергопреобразующую гидрогеназу. Был ли он направлен внутрь клетки и осуществлял фиксацию углерода – или был обращен наружу через белок, который сейчас функционирует как мембранный насос, выкачивающий из клетки протоны (рис. 20)? Думаю, что поток протонов в предковой популяции протоклеток был направлен вовнутрь, и его энергия расходовалась на фиксацию CO2. Затем две отдельные популяции независимо приобрели механизмы создания протонных градиентов. В популяции, от которой произошли ацетогены, произошло обращение работы энергопреобразующей гидрогеназы: она стала окислять ферредоксин и за счет выделившейся энергии выкачивать протоны из клетки. Но это простое и полезное приобретение порождает проблему: восстановленный ферредоксин, который ранее использовался для фиксации углерода, теперь расходуется на транспорт протонов. Поэтому ацетогенам прошлось изобрести новый способ восстановления углерода, который не зависит от ферредоксина. Их предки нашли хитроумный выход: бифуркация потока электронов, которая позволяет осуществлять непрямое восстановление CO2. Вполне возможно, что происхождение базовой биохимии ацетогенов вытекает из этого простого допущения – инверсии работы энергопреобразующей гидрогеназы, в результате чего ацетогены приобрели протонный насос, а с этим и ряд проблем, которые им пришлось решить.
В другой популяции клеток, давшей начало метаногенам, был найден альтернативный путь создания ионных градиентов. Клетки в этой популяции, как и их предшественники, восстанавливали ферредоксин, используя энергию протонных градиентов, и при помощи восстановленного ферредоксина осуществляли фиксацию углерода. Но этим клеткам пришлось с нуля изобрести ионный насос. Ну, почти с нуля: они переоборудовали для этой цели другой белок. Вероятнее всего, они превратили свой антипортер в прямой насос. Осуществить это несложно, но возникла проблема: как снабдить насос энергией? Метаногены приобрели другой тип бифуркации электронов. Они пользовались для этого теми же белками, что и ацетогены, но сообразно своим потребностям построили совершенно иной путь, позволяющий подключить ионный насос. Различия в углеродном и энергетическом метаболизме у метаногенов и ацетогенов связаны с направлением потока протонов через энергопреобразующую гидрогеназу. Можно выбрать: либо внутрь, либо наружу. Метаногены и ацетогены выбрали разные решения.
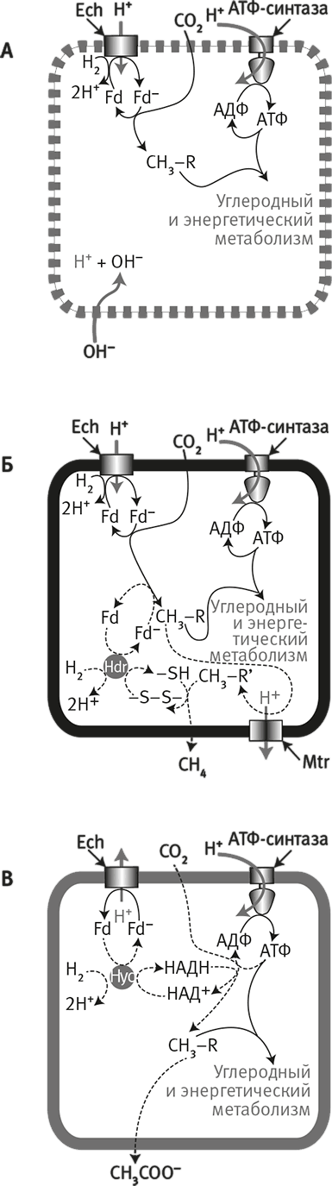
Рис. 20. Модель эволюции активного транспорта ионов.
Гипотеза происхождения активного транспорта ионов у бактерий и архей, основанная на направлении потока протонов через мембранный белок Ech.
А. В предковом состоянии естественный протонный градиент поддерживает углеродный и энергетический метаболизм посредством энергопреобразующей гидрогеназы (Ech) и АТФ-синтазы. Это путь возможен лишь до тех пор, пока мембрана “проницаема” для протонов.
Б. Клетки метаногенов (предположительно, именно такой образ жизни вели предки архей) также используют Ech и АТФ-синтазу для углеродного и энергетического метаболизма. Однако непроницаемые для протонов мембраны уже не позволяют им использовать естественные протонные градиенты. Поэтому им пришлось найти новые биохимические пути (пунктирные линии) и изобрести новый ионный насос (метилтрансферазу, Mtr) для получения собственных H+– или Na+-градиентов. (Обратите внимание: эта схема получается в результате комбинации схем А и Б на рис. 18.)
В. Ацетогены (предположительно древнейшая ветвь бактерий). Ech стала работать в обратном направлении, выкачивая протоны из клетки за счет энергии окисления восстановленного ферредоксина. Поэтому ацетогенам не потребовалось изобретать новый ионный насос, но пришлось найти новый путь фиксации CO2 – с использованием НАДН и АТФ (пунктирные линии). Этот гипотетический сценарий позволяет объяснить все сходства и различия пути Вуда – Льюнгдаля у метаногенов и ацетогенов.
Когда обе группы клеток тем или иным способом приобрели механизмы активного транспорта ионов, им наконец стало выгодным усовершенствование мембран. На ранних этапах развития мембраны современного типа, обогащенные фосфолипидами, не приносили никакой пользы и даже были бы вредны. Но вскоре после того, как клетки приобрели антипортер и ионные насосы, стало выгодным встраивать в мембраны липиды с глицерольными полярными группами. Такие липиды, по-видимому, независимо появились в двух разных доменах, поэтому археи и бактерии в мембранных липидах используют зеркальные стереоизомеры глицерола (гл. 2).
Теперь, когда клетки приобрели активный транспорт ионов и мембраны современного типа, они смогли покинуть гидротермальные источники и выйти в океан. От общего предка, который жил за счет естественных протонных градиентов источников, отделились две ветви: археи и бактерии. Нет ничего удивительного ни в том, что бактерии и археи приобрели разные клеточные стенки, защищающие в незнакомой среде, ни в том, что они сконструировали различные механизмы репликации ДНК. У бактерий в ходе деления клетки ДНК прикрепляется к мембране за особый участок – репликон, что позволяет дочерним клеткам получить по копии родительского генома. Структура задействованных в этом молекулярных комплексов, как и многих других компонентов репликации ДНК, должна хотя бы отчасти зависеть от механизма прикрепления ДНК к мембране. Независимое происхождение клеточных мембран позволяет объяснить различия в репликации ДНК у архей и бактерий. То же самое относится к строению клеточных стенок. Перед постройкой клеточной стенки необходимо транспортировать все предшественники наружу через специальные мембранные поры. То есть синтез клеточной стенки зависит от свойств мембран, а отсюда следует, что у архей и бактерий он должен различаться.
Несмотря на то, что из базовых принципов биоэнергетики не удается вывести необходимость существования фундаментальных различий бактерий и архей, мы можем объяснить, как и по каким причинам они могли возникнуть. Глубокие различия между двумя доменами прокариот не имеют ничего общего с адаптацией к экстремальным условиям, например высокой температуре. Вероятнее всего, эти различия появились в ходе дивергенции предковой популяции клеток, мембраны которых по биоэнергетическим причинам должны быть полупроницаемыми. Хотя дивергенцию архей и бактерий нельзя предсказать исходя из базовых принципов, тот факт, что обе группы используют энергию протонного градиента на мембране, следует из физических законов, которые мы обсуждали в последних двух главах. Щелочные гидротермальные источники – наиболее подходящий кандидат на роль “колыбели жизни” на Земле или где-либо еще во Вселенной. Благодаря гидротермальным источникам клетки стали использовать природные протонные градиенты, в конце концов научившись создавать их самостоятельно. Готов поспорить, что если где-то во Вселенной существуют живые клетки, они также хемиосмотические, а значит, им пришлось столкнуться с теми же проблемами, что и на Земле. Далее мы узнаем, почему из необходимости энергии протонных градиентов следует, что сложная жизнь во Вселенной должна быть редким явлением.
Назад: Глава 3 Энергия и начало жизни
Дальше: Часть III О сложности

