Книга: Достающее звено. Книга 1. Обезьяны и все-все-все
Назад: Глава 41 Примитивнейшие приматы
Дальше: Заря в преддверии заката: первые человекообразные
Глава 42
Обезьяны
Обезьяны, “сухоносые”, или высшие приматы (Haplorrhini), подразделяются на три большие современные группы: долгопятовых, широконосых и узконосых. Ископаемые обезьяны более разнообразны. Обезьяны в целом отличаются от полуобезьян более развитым мозгом с редуцированными обонятельными луковицами и, соответственно, плохим обонянием, зато хорошим зрением, ориентированными вперед глазницами, более-менее отделенными от височной ямки костной стенкой, свободной верхней губой и богатой мимикой.
Момент расхождения полуобезьян и обезьян определить очень трудно, потому что примитивные представители тех и других чрезвычайно похожи друг на друга. Примером может служить носмипс Nosmips aenigmaticus из Египта, живший 37 млн лет назад и известный лишь по дюжине зубов: по морфологии он годился бы на роль предка в основании адаписовых, прочих полуобезьян и обезьян. Однако датировка носмипса слишком поздняя для того, чтобы считать его универсальным пращуром. Не исключено, что носмипс представляет особую группу приматов.
Долгопятообразные Tarsiiformes
Внижнем эоцене из какой-то группы полуобезьян выделилась линия современных обезьян, но почти сразу она разделилась на долгопятообразных и антропоидов. Поэтому долгопятообразные имеют едва ли не большинство признаков полуобезьян. По этой же причине раньше их включали именно в полуобезьян, потом стали выделять в самостоятельную промежуточную группу, а ныне рассматривают как специализированных и во многом примитивных, но все же обезьян.
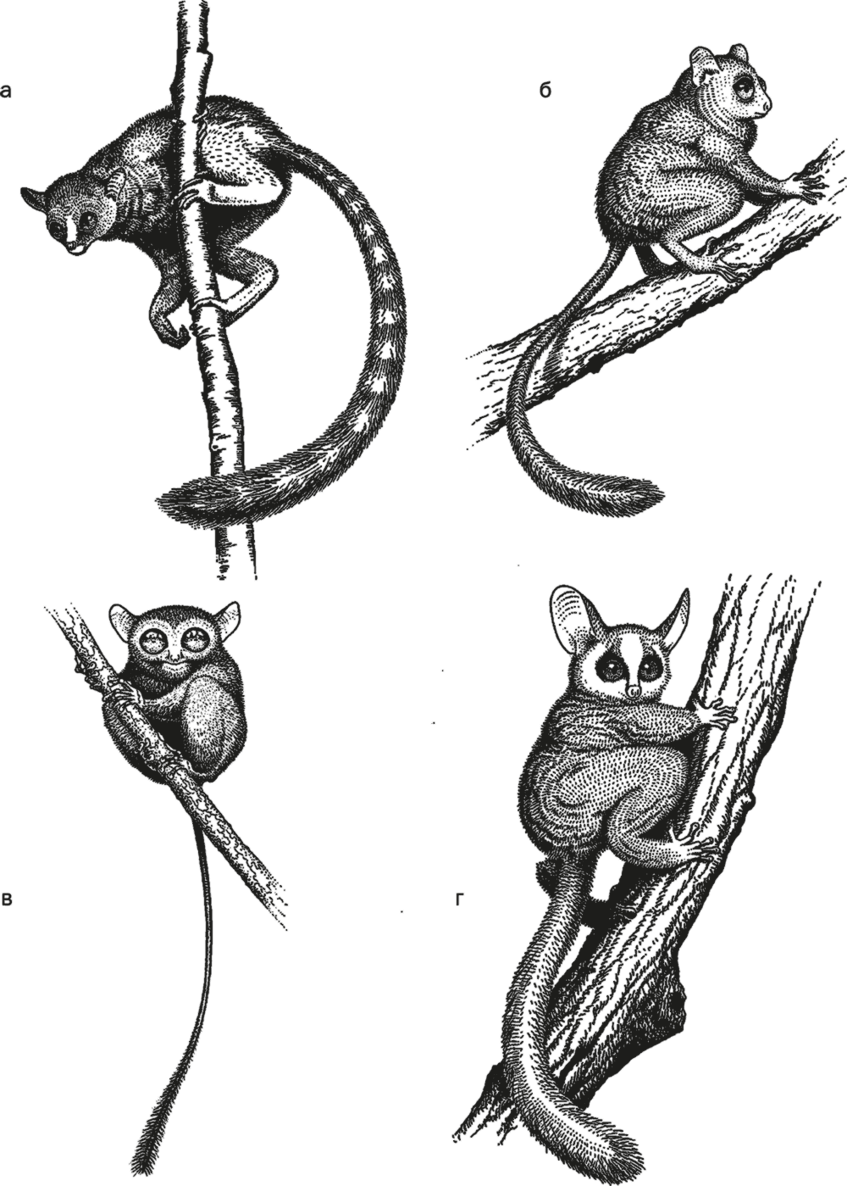
Рис. 30. Archicebus achilles (а), Necrolemur (б), долгопят (в) и галаго (г).
Омомисовые (Omomyiformes) объединяют великое множество эоценовых долгопятообразных. Они были, наряду с адаписовыми, основными участниками второй радиации приматов в раннем эоцене. Не исключено, что их заря зарделась еще в среднем палеоцене Азии, так как в эту эпоху в Китае жил декоредон Decoredon anhuiensis. Однако это существо известно крайне плохо, неспроста изначально декоредон был определен вообще как кондиляртр.
Несравненно лучше сохранился архицебус Archicebus achilles – самая древняя настоящая обезьяна, лучше всех прочих подходящая на роль общего предка долгопятообразных и антропоидов.
Великий Предок – древнейший, наицелейший, примитивнейший и просто красавец
А у Икарушки бедного,
Всеми братками забытого,
Только бледные ножки торчат
Из холодной зеленой водицы…
Д. В. ШагинНаходки целых скелетов древних животных – большая редкость. А если эти животные к тому же жили на деревьях и не любили воду (а скелеты сохраняются преимущественно в водных отложениях), то вероятность обнаружения их останков крайне мала. Потому древнейшие приматы известны плохо и почти исключительно по зубам. Потому сенсацией стала находка почти целого скелета одного из древнейших приматов – архицебуса Archicebus achilles.В очередном номере журнала Nature китайско-американо-французская группа исследователей описала не только новый род и вид, но целое семейство древнейших приматов – архицебид Archicebidae (Ni et al., 2013). Новейшая находка уникальна по всем возможным показателям.Во-первых, датировка: скелет сохранился в отложениях, образовавшихся в самом начале эоцена, 54,8–55,8 млн лет назад. Такого древнего и одновременно целого скелета обезьян еще никогда не находили. Были, правда, скелеты полуобезьян, но эти два главных подразделения отряда приматов – совсем не одно и то же. Как всегда, огромный интерес вызывает момент расхождения главных эволюционных ветвей, зарождение новой группы. Омомисовые известны со среднего палеоцена, но в подавляющем большинстве случаев от них сохранились лишь зубы размером с песчинку. Доселе примитивнейшим представителем группы была тейярдина азиатская Teilhardina asiatica, описанная в 2004 году из отложений границы палеоцена и эоцена Китая, а древнейшими – декоредон Decoredon anhuiensis из середины палеоцена (с его статусом есть большие проблемы, далеко не все палеонтологи признают его за примата), анеморисис Anemorhysis tenuiculus и тетониус Tetonius homunculus из конца палеоцена, еще две тейярдины Teilhardina brandti и T. magnoliana, баатромомис Baataromomys ulaanus и два вида вастаномисов Vastanomys gracilis и V. major, синхронные новоописанному архицебусу. Показательно, кстати, что всплеск разнообразия приходится на границу между эпохами палеоцена и эоцена, ведь эта граница потому и определима, что отражает существенные изменения климата. Однако все эти зверюшки известны по столь скудным останкам, что ученым приходилось напрягать все усилия, чтобы разобраться в их родстве и хоть как-то представить их облик.Тут мы переходим ко второй уникальной особенности архицебуса – его сохранности. Скелет этого зверька сохранился не то чтобы целиком, но настолько, как палеонтологи могли только мечтать. Особенно радуют идеально сохранившиеся ножки и трогательный хвостик. С головой дела обстоят хуже, время не пощадило хрупкий череп, смяв его в не очень внятное пятно. Все же определимых и сравнимых признаков на скелете архицебуса на порядок больше, чем на любой другой находке древнейших обезьян (правда, сравнивать приходится опять же зубы, ибо от других приматов сохранились в основном они).Благодаря этим сохранившимся признакам возможно оценить третью уникальность архицебуса – его крайнюю архаичность. По сочетанию признаков он оказывается максимально приближенным к ожидаемому первопредковому для всех обезьян состоянию: с одной стороны, он лишен явных черт полуобезьян, с другой – отсутствуют специализации. Например, у архицебуса нет сращения большой и малой берцовых костей, нет у него и крайнего удлинения пяточной кости. Эти особенности древних омомисовых и современных долгопятов всегда огорчали исследователей, ведь омомисовые в принципе подходят на роль предков прочих обезьян, но подобные специализации ставят на идее предковости крест. Доселе оставалось предполагать, что более поздние обезьяны произошли от ранних неспециализированных форм омомисовых, но внешний облик таких предков оставался в значительной мере гипотетическим. По одним зубам о строении конечностей много не скажешь. У архицебуса же ноги – какие надо ноги, из таких могли возникнуть как скакательные лапки долгопятообразных, так и хватательные – антропоидов.Конечно, архицебус не мог не преподнести и сюрпризов. Например, в стопе предплюсна имеет умеренную длину, как у антропоидов, плюсна – большую, как у тупай и антропоидов (но в отличие от лемуров, адаписовых и долгопятов), а фаланги – тоже большую, как у долгопятов. Фаланги архицебуса сильно выпрямлены – специфическая особенность; самым длинным пальцем стопы был четвертый – это сближает архицебуса с адаписовыми и долгопятами.На примере стопы отлично видна очередная уникальная особенность архицебуса – сочетание признаков разных групп обезьян. В нем есть основа от долгопятообразных, но немного от широконосых, немного от узконосых, а что-то – и от эосимид, которые сами сочетают черты узко– и широконосых. Много примитивного и мало специализированного – идеальное сочетание для идеального Великого Предка. Ранее известные приматы обычно без особых проблем могли быть классифицированы либо как долгопятообразные, либо как антропоиды. Если бы от архицебуса были найдены только зубы, его тоже без малейших сомнений причислили бы к примитивным долгопятообразным и его имя затерялось бы среди бесчисленных “-омисов”. Но теперь у нас слишком много информации, чтобы так упрощать картину. Становится очевидным, что ранние обезьяны уже имели, хоть бы в зародыше, массу особенностей, одни из которых развились и усилились в одних группах, а другие – в других. В очередной раз приятно видеть, что понятия примитивности и продвинутости крайне относительны.Как же, наконец, выглядел архицебус и какой вел образ жизни?Это было небольшое – с крупную мышь, 20–30 г – животное, с округлой головой, не слишком большими глазами, маленькой суженной мордочкой, крохотным носом и острыми зубами. Очень длинный хвост служил противовесом при прыжках. Скакать он мог благодаря очень длинным задним ногам, а сравнительно короткими передними лапками цеплялся за ветки и хватал насекомых. Впрочем, и способ передвижения, и тип питания не были специализированы. Архицебус прыгал совсем не так специфически, как современные галаго и долгопяты, а мог и бегать на четырех ногах по ветвям. Преимущественно насекомоядная диета также не исключает того, что архицебус мог есть и другую пищу. Судя по размерам глаз, он вел дневной образ жизни.Самый целый из самых древних, наиболее примитивный и одновременно генерализованный неспециализированный, воплотивший в себе идеал Великого Общего Предка – о чем еще можно мечтать?! Приятно, когда в одной находке сочетается столько достоинств! Порадуемся же преодолению еще одной ступени познания!
На самом деле, не только архицебус может быть предком высших обезьян, просто он очень уж хорошо сохранился. Из Индии известен вастаномис Vastanomys (54–55 млн лет назад), который обладал едва ли не самым примитивным среди всех омомисовых строением и, судя по зубам, был не насекомо-, а фруктоядным. Но от вастаномисов сохранились одни зубы, так что мы не знаем о строении их ног или черепа.
Таким образом, омомисовые явились предками долгопятов и высших обезьян – такая версия называется тарзиоидной гипотезой. Слава эволюции, что в самом начале группы омомисовых нашелся не слишком специализированный примат, давший начало более продвинутым обезьянам. Судя по всему, омомисовые специализировались чрезвычайно быстро, не успей зародиться линия антропоидов в начале эоцена, они могли бы и вовсе не появиться. Или же эволюция воспользовалась бы материалом из других групп – адаписовых или, возможно, даже плезиадапиформов.
Основные группы омомисовых – омомиды Omomyidae и микрохэриды Microchoeridae – жили в Северной Америке, Европе, Северной Африке, Южной и Восточной Азии. Те и другие имели очень маленькую – укороченную и суженную спереди – мордочку и округлый череп с огромными направленными вперед глазами. В строении зубов многих видов сохраняется такой примитивный признак, как четыре премоляра. Ноги омомисовых были длинными, со сросшимися малой и большой берцовой костями и резко увеличенной пяткой, образующей цевку. Очевидно, это были ночные, насекомоядные и активно прыгающие животные. Правда, во всех этих чертах омомисовые не превзошли долгопятов, но стремились к идеалу как могли; в силу не слишком далеко зашедшей специализации омомисовые могли внешне напоминать скорее галаго, чем долгопятов.
Микрохэриды были очень разнообразными в размерах животными: от 45 г у Pseudoloris parvulus до 1775 г у Microchoerus erinaceus. Объединяются они благодаря специфическому строению зубов: в частности, первые верхние и нижние резцы несколько увеличены и выдвинуты перед вторыми резцами. Не очень ясно, зачем им нужна была эта специализация.
Омомиды чрезвычайно многочисленны. Большинство их весило от 100 до 200 г, но были и рекордсмены: Teilhardina magnoliana была размером с мышь – 28 г, а Anaptomorphus westi, Gazinius amplus и Stockia powayensis с крысу – 465–475 г, Hemiacodon gracilis тянул на целый килограмм, а представители родов Rooneyia, Ourayia и Macrotarsius – от килограмма до 2520 г! Часть омомид питались преимущественно насекомыми (например, многие в подсемействе Omomyinae и представители трибы Washakiini), однако, в отличие от современных долгопятов, очень многие были фруктоядными (например, почти все представители подсемейства Anaptomorphinae), а некоторые даже листоядными (например, Hemiacodon gracilis).
Кстати, о размерах…
Размеры ископаемых приматов определяются разными способами. Конечно, идеальный случай, когда в нашем распоряжении есть целый скелет. Но это бывает крайне редко. Чаще приходится довольствоваться отдельными зубами. Тогда в дело идет регрессионный анализ: из соотношений размеров зубов (например, площади первого верхнего или длины второго нижнего моляра) и веса тела у современных приматов высчитывается вес ископаемых животных. Дело Ж. Кювье живет и побеждает! Понятно, что погрешность таких вычислений весьма велика – у разных авторов и полученные разными методами цифры могут отличаться в разы, – да и индивидуальную изменчивость никто не отменял, так что публикуемые величины не должны восприниматься слишком буквально, а служат лишь примерным ориентиром.Можно обратить внимание, что почти никогда речь не идет о линейных размерах – длине тела. Дело в том, что приматы – гибкие зверюшки, они удивительно пластичны, даже у человека рост при нескольких измерениях легко может различаться на несколько сантиметров. Так что палеоприматологи предпочитают пользоваться определениями веса.
Altanius orlovi – единственная полуобезьяна из Монголии; он определяет северную известную границу распространения этих животных в Азии. Меж тем фауны Северной Америки и Евразии весьма схожи. Либо обезьяны все же могли преодолевать Берингию, просто мы пока не нашли их в Сибири, либо они периодически форсировали расширявшиеся моря между Северной Америкой и Европой и между Европой и Азией, ибо Европа была островом между двух континентов. По этим же географическим причинам североамериканские омомисовые (как, кстати, и адаписовые) намного разнообразнее европейских: Северная Америка больше по размеру и богаче на условия, вот и животные были отличающимися, тогда как в Европе имелось меньше вариаций экологических ниш, отчего и приматы были схожими (Gilbert, 2005).
Омомисовые в Европе и Северной Америке дожили до границы эоцена и олигоцена (около 34 млн лет назад или чуть позже), после чего вымерли по тем же причинам, что свели в могилу адаписовых и многих прочих животных, процветавших в тропических лесах эоцена.
Долгопятовые (Tarsiiformes) современности обычно объединяются в один род Tarsius, хотя предложено разделить его на три, сохранив прежнее название лишь за сулавесскими видами, тогда как суматранско-калимантанские выделяются в род Cephalopachus, а филиппинские – Carlito (Groves et Shekelle, 2010). Впрочем, разница между ними очень невелика, даром что разошлись островные популяции как минимум еще в миоцене.
В строении долгопятов совмещаются признаки полуобезьян и настоящих обезьян. Это животные размером с крысу, с очень большими глазами и специализированными для прыгания ногами. Они активны ночью и питаются насекомыми. Во многих чертах строения проявляется резкая специализация: глаза столь велики, что не могут вращаться, отчего подвижность шеи резко увеличена, благодаря огромному позвоночному отверстию атланта (первого шейного позвонка) и хитрому строению межпозвоночных сочленений шея может поворачиваться задом наперед; мордочка крайне маленькая; кончики пальцев расширены в виде присосок, большая и малая берцовые кости срастаются, пятка резко удлинена в виде цевки для прыгания по ветвям, а второй и третий пальцы стопы снабжены когтями. Живут долгопяты семейными группами, общаясь друг с другом своеобразным щебетом, похожим на птичий.
О долгопятах подробно говорилось выше, тут помянем лишь их происхождение.
45 млн лет назад в Китае жили как минимум три вида долгопятов или чрезвычайно похожих на них существ, правда, известны они только по таранной и пяточным костям. Один из них весил всего 30 г – вдвое меньше самых маленьких долгопятов: карликовых Tarsius pumilus. Из того же времени Китая описан Xanthorhysis tabrumi, но от него найдены лишь зубы, так что совпадает ли он с одним из трех “таранно-пяточных” видов – непонятно.
Некий долгопятовый зверек жил в Таиланде около 34 млн лет назад, но известен он лишь по одному зубу. В это же время в Китае скакал Oligotarsius rarus.
Миоценовый Hesperotarsius sindhensis обитал 16–18 млн лет назад в Пакистане. Своим существованием он указывает, что древние долгопяты жили далеко за пределами своего нынешнего ареала.
К современному роду относятся три ископаемых вида, причем древнейший – Tarsius eocaenus – жил аж в эоцене, более 37 млн лет назад, в Китае. Почти современно выглядят Tarsius thailandica (18 млн лет назад) и Tarsius sirindhornae (13,1–13,3 млн лет назад) из Таиланда.
Выжить в Индонезии и на Филиппинах долгопятам помогла крайняя специализация, а позволил не слишком сильно менявшийся климат.
Возникновение высших приматов
Время и место возникновения, а также родство первых человекоподобных обезьян-антропоидов – одна из наиболее дискуссионных тем в антропологии. Исследователей тут подстерегает множество трудноразрешимых сложностей: вечный недостаток материалов, крайняя ограниченность доступных для оценки морфологических систем, различие взглядов на таксономическую ценность тех или иных признаков. Поэтому, с одной стороны, каждая новая находка дает много нового, а с другой – интерпретации множатся и найти правду среди них сложно.
Первые человекоподобные-антропоиды известны почти исключительно по обломкам нижних челюстей, зубам и костям стоп. Никто еще не видел целого черепа и тем более скелета такой обезьяны. А ведь выглядеть она должна любопытно: это же переходное звено от полуобезьян к обезьянам. Понятно, что у такой зверюшки – как у любого приличного “достающего звена” – признаки предков и потомков перемежаются в полупереплавленном и частично мозаичном состоянии, что лишь добавляет головной боли систематикам, ибо трудно классифицировать цвета в непрерывном спектре, к тому же ветвящемся.
Начальные этапы эволюции высших приматов-антропоидов известны по четырем группам: эосимидам Eosimiidae, афротарзиидам Afrotarsiidae, амфипитецидам Amphipithecidae и парапитецидам Parapithecidae. Вместе они представляют третью радиацию приматов на границе эоцена и олигоцена. Есть два географических центра, где концентрируются находки: в Китае и Бирме с одной стороны и Египте с Ливией – с другой. Конечно, это не значит, что древнейшие антропоиды жили только там или что именно там от них возникли последующие группы. Просто именно в этих местах были подходящие условия для сохранения ископаемых останков, и именно тут их нашли палеонтологи, которым приходится действовать, как пьянице из анекдота, ищущему ключи под фонарем, потому что там светлее.
Кстати, о границе эоцена и олигоцена…
Эоцен – геологическая эпоха, длинная и теплая, в которую вершились великие дела. Это было важное время. После катастроф конца мелового периода и палеоценовых заморозков Земля вновь покрылась тропическими лесами. Старое уступало место новому, полуобезьяны становились обезьянами, а среди последних появлялись особенные линии, часть из которых оказалась предками современных высших приматов. Многое известно об эоценовых приматах, но чем больше мы узнаем о них, тем мудренее получается картина. Важнейшие события происходили на самом закате эоцена, на границе олигоцена – следующей, более сухой и холодной эпохи. Почти с самого начала эоцена климат постепенно становился все более холодным, а рубеж эпох ознаменовался резким похолоданием-оледенением: в начале олигоцена оледенела Антарктида. Это погубило тропические леса Северного полушария, а вместе с ними – скакавших по ним приматов. Впрочем, во всем есть плюсы: антарктические ледники взяли на себя гигантские массы воды, уровень океана понизился аж на 55 м, так что многие проливы и даже моря исчезли, а между материками возникли мосты суши, например, Северо-Восточная Европа соединилась с Северной Азией, Южная – с Южной, а Африка стала гораздо ближе как к той, так и к другой. В это же время Индия, наконец, доплыла до Азии и воткнулась в нее. Правда, возникшие просторы оказались заняты в основном степями, так что приматы расселяться по ним до поры до времени не могли. Олигоцен – время раздолья примитивных носорогов, лошадей и верблюдов (например, всем известны индрикотерии: даже люди, никогда не интересовавшиеся палеонтологией, видели их в фильме “Звездные войны. Империя наносит ответный удар” в виде четвероногих машин, шагающих по ледяной планете Хот, – отличная, хотя и доведенная до крайности аллегория на реалии олигоцена). Неспроста и ластоногие, ставшие в своем роде символом стужи, появились именно в эту эпоху. Впрочем, не стоит и преувеличивать: даже в самые ужасно холодные этапы олигоцена климат был не морознее современного, а то и существенно теплее.Климатические и экологические перемены привели к исчезновению многих архаичных групп приматов и появлению более прогрессивных – обезьян. Одна из главных интриг – место появления обезьян. Как обычно бывает в палеоантропологии, за звание Великой Прародины борются два региона: Африка и Азия. Полуобезьяны известны из обеих, обезьяны – тоже, но где жили промежуточные звенья – вопрос не до конца решенный. С одной стороны, в Бирме и Китае были найдены многочисленные останки обезьян, относимых к семейству Eosimiidae, живших в среднем, а то и раннем эоцене, в основном около 45 млн лет назад. После их сменили Amphipithecidae, во многом схожие, но потреблявшие в пищу меньше насекомых и больше плодов. Намного позднее в Азии возникают Pliopithecidae. С другой стороны, в Северной Африке обнаружена масса видов, образующих семейства Afrotarsiidae, Parapithecidae, Propliopithecidae и Proteopithecidae. В среднем они моложе эосимид, но древнее многих амфипитецид, хотя по ряду признаков выглядят и прогрессивнее последних. Африканские ранние обезьяны вроде бы хорошо подходят на роль предков всех последующих обезьян, но вопрос – появились ли они изначально в Африке из местных полуобезьян, или их предки пришли все же из Азии. Конечно, неразберихи добавляет крайняя фрагментарность большинства находок и зачастую – неопределенность датировок. В изобилии зубов и обломков челюстей разобраться крайне сложно, а потому любые новые находки принимаются антропологами со смешанными чувствами: с пополнением данных картина вроде бы должна проясняться, но часто запутывается еще пуще.
Древнейшие антропоиды были преимущественно древесными дневными растительноядными животными размером от долгопята до мартышки. Все они характеризуются смесью архаичных и прогрессивных признаков, поэтому систематика их весьма запутана.
Эосимиды (Eosimiidae): Anthrasimias, Bahinia, Eosimias, Phenacopithecus и Phileosimias – крайне примитивные антропоиды из эоцена (55–33 млн лет назад) Индии, Пакистана, Бирмы и Китая. Они сохраняли массу признаков полуобезьян, но по некоторым существенным чертам строения зубов могут быть признаны древнейшими настоящими обезьянами. Самый древний из них – Anthrasimias gujaratensis из Западной Индии, живший 54–55 млн лет назад, – был и самым мелким: он весил всего 75 г. Судя по зубам, он был фруктоядным или даже смолоядным. Более поздние представители семейства были покрупнее – 100–400 г. Самый крупный вид – Bahinia pondaungensis – весил примерно полкило, в самом крайнем варианте – меньше полутора килограммов. От эосимид сохранились в основном зубы и обломки челюстей, так что судить о внешности этих животных трудно. Известны лишь пара обломков черепов и горсть костей стопы эосимид.

Рис. 31. Eosimias (а), Amphipithecus (б) и Apidium (в).
Судя по обломку черепа бахинии Bahinia pondaungensis, ее глазницы, с одной стороны, были сравнительно небольшие, с другой – ориентированы наискосок, как у лемуров и адаписовых, а не вперед, как у обезьян (Rosenberger et Hogg, 2007). Бахиния имела крупную мордочку и высокое подглазничное пространство. Сходство ее черепа с адаписовыми по многим признакам более сильное, чем с омомисовыми. Это может быть следствием конвергенции, сохранения эосимидами примитивных черт в отличие от специализированных омомисовых, или же говорить о том, что эосимиды на самом деле полуобезьяны, а не обезьяны, с коими у них конвергентно схожи лишь зубы.
Верхняя челюсть фенакопитека Phenacopithecus krishtalkai по размерам такая же, как у долгопята, но его подглазничное пространство пропорционально такое же, как у антропоидов, самые мелкие из коих в полтора раза крупнее (Beard et Wang, 2004). Это свидетельствует о небольших размерах глазниц и, соответственно, дневном образе жизни фенакопитека.
Пяточные и таранные кости эосимид из Китая и Бирмы замечательны сочетанием признаков полуобезьян и обезьян (Gebo et al., 2000, 2001, 2002). Отличия между этими группами достаточно существенны, так как полуобезьяны предпочитают передвигаться по вертикальным веткам и очень прочно обхватывают их пальцами, а обезьяны используют гораздо более разнообразный репертуар движений, любят бегать по горизонтальным ветвям и не так сильно цепляются за них. Эосимиды уникальны, так как у них мы воочию видим переход от одного варианта к другому.
Уголок занудства
Полуобезьяны и обезьяны хорошо отличимы по форме костей стопы.На таранной кости латеральная лодыжковая поверхность (суставная фасетка для малой берцовой кости на боковой стороне кости) у полуобезьян наклонная, а у обезьян вертикальная. Также борозда на задней стороне блока, служащая для сухожилия длинного сгибателя большого пальца стопы у полуобезьян расположена латерально, а у обезьян следует плантарно вдоль задней стороны латеральной лодыжковой поверхности. Наконец, задний скат блока таранной у полуобезьян большой, а у обезьян маленький. На таранных костях Eosimias эти три особенности обезьяньи и четко отличаются от полуобезьяньих (Gebo et al., 2000). Впрочем, вопреки кажущейся логике, “обезьяний” вариант более примитивен, нежели “полуобезьяний”, так как именно обезьяны сохранили исходно приматное строение стопы, а полуобезьяны эволюционировали быстрее. Среди приматов эосимиды, что понятно, больше похожи на омомисовых, чем на высших обезьян: их таранная кость умеренно высокая и сравнительно узкая, имеет умеренный угол наклона шейки, мелкий блок и маленький задний скат блока. Обезьяньей и одновременно примитивной чертой выглядит укороченная медиальная лодыжковая поверхность (фасетка для соединения с внутренней лодыжкой большой берцовой кости).На пяточной кости полуобезьян задняя таранная суставная поверхность вытянутая и узкая, с хорошо оформленным плантарным краем, а обезьян – короткая и широкая, с размытым плантарным краем; эосимиды похожи на обезьян. Кубовидная суставная поверхность пяточной у полуобезьян и омомисовых широкая, плоская, веерообразная с центрально расположенной осью вращения, у обезьян круглая, с несуставной вырезкой в медиоплантарной части и смещенной медиально осью вращения; у Eosimias плоская, но округлая с вырезкой и медиально смещенной осью.На основании первой плюсневой кости у полуобезьян имеется большой бугор для прикрепления мощной длинной малоберцовой мышцы, а у обезьян его нет (у омомисовых есть, но не толстый, а узкий и высокий). На первой плюсневой кости из Китая, предположительно принадлежащей эосимиду, указанный бугор был (Gebo et al., 2008).
Примечательно, что в среднеэоценовых экосистемах, где одновременно встречались адаписовые, омомисовые и эосимиды, первые были наиболее крупными, вторые – более-менее средними, а последние – самыми маленькими приматами. Такое распределение напрочь отличается от нынешнего, где долгопяты самые маленькие, полуобезьяны средние, а обезьяны – большие. Как часто бывает, современные стереотипы не срабатывают в приложении к древности. К тому же лишний раз подтверждается истина, что все великое вырастает из малого.
Кстати, о кузнечиках и капусте…
Размеры обезьян связаны с их питанием: мелкие приматы должны поддерживать свой активный метаболизм, для этого им нужно много легкоусвояемых белков, которые легче всего получить, питаясь насекомыми. Крупные лучше сохраняют тепло за счет массы тела, так что им позволительно переходить на менее питательные, зато шире распространенные и легче добываемые ресурсы, самым дешевым из которых являются листья. И те и другие могут есть фрукты, ведь их более-менее легко добыть, но они при этом достаточно питательные. С другой стороны, только насекомыми питаться слишком большой примат тоже не может, потому что будет тратить слишком много усилий на охоту, но получать от этого несопоставимо мало выгоды.Приматологи, изучая диеты современных приматов, выявили несколько надежных закономерностей. В частности, существует так называемая “граница Кея” – вес в 500 г, – разделяющая тех, кто должен и может получать белки, постоянно питаясь только насекомыми, от тех, кто уже никак не способен на такое, зато может позволить себе жевать одни листья (Kay et Covert, 1984). Насекомоядные приматы, регулярно подкрепляющие свои силы фруктами, могут быть значительно крупнее “границы Кея” – до 1,4 кг. Те же, которые едят только листья и фрукты, не бывают легче килограмма (Kirk et Simons, 2001; Ramdarshan et al., 2010). Впрочем, ни одна из этих границ не позволяет оценить степень фруктоядности, так как в качестве десерта фрукты готовы уплетать приматы любого калибра.Практически все эосимиды оказываются легче “границы Кея”, так что с большой вероятностью белки они получали преимущественно из насекомых. Это не так странно, учитывая их вероятное происхождение от омомисовых приматов, многие из которых были именно насекомоядными. Однако строение зубов указывает, что все же основой диеты эосимид были фрукты. Такая диета при увеличении размеров тела легко могла превратиться в ту, что типична для современных антропоидов.
Как уже упоминалось, филогенетические связи эоценовых и олигоценовых “недообезьян” остаются весьма туманными, тут еще неизбежны открытия и откровения. Очередным “достающим звеном” явился верхний моляр древностью 39 млн лет, найденный недавно в Центральном Тунисе. По зубу был описан новый род и вид амамрия Amamria tunisiensis (Marivaux et al., 2014). Казалось бы, всего один зуб, но он очень уж красиво связывает азиатских эосимид и североафриканских проплиопитековых приматов. Признаки амамрии отличаются от типичных эосимиевых как раз в направлении проплиопитековых, а от проплиопитековых – в направлении эосимиевых. Идеальное сочетание для “достающего звена”! Причем среди проплиопитековых сходство обнаруживается у самых древних и примитивных представителей – протеопитецид и олигопитецин, живших как раз чуть позже амамрии, – опять же красивее некуда. Отличия же амамрии от афротарзиид свидетельствуют, что последние мигрировали из Азии в Африку независимо.
Последние эосимиды дожили в Пакистане до начала олигоцена – Phileosimias kamali и Ph. brahuiorum, они известны всего по двум зубам. Но в это время мир занимали уже новые приматы…
Афротарзииды (Afrotarsiidae) – странная группа приматов с границы эоцена и олигоцена (39–29,5 млн лет назад) Египта, Ливии и Бирмы. В последнее время все настойчивее слышатся голоса палеонтологов, призывающих обратить пристальное внимание на эту группу. Первым был описан афротарзиус Afrotarsius chatrathi, живший примерно 30 млн лет назад в Египте. Как понятно из названия, сначала он был определен как африканский долгопят, благо и размеры у него были более-менее подходящие – от 188 до 357 г. Не так давно в местонахождении Дур-ат-Тала в Центральной Ливии были найдены зубы более древнего (35–39 млн лет назад) представителя того же рода, названного Afrotarsius libycus.
Кстати, о Ливии…
Тут же – в Дур-ат-Тала – обнаружены зубы ископаемого потто Karanisia arenula, древнейшего олигопитекового Talahpithecus parvus и парапитекового Biretia piveteaui (Jaeger et al., 2010). Эта находка представляет огромный интерес, поскольку заполняет географический и хронологический разрыв в представлениях об эволюции сразу нескольких групп древних приматов. Картина пополнилась еще несколькими кусочками мозаики, причем в тех частях, которые доныне оставались темными.В частности, изменились представления о времени возникновения антропоидов. Ранее на основании существовавших палеонтологических данных предполагалось, что человекоподобные появились в Африке в эоцене. Однако новые находки показывают, что к середине эоцена в Африке уже существовал ряд очень разнообразных групп антропоидов. Это может означать, что либо эволюция этих приматов в Африке началась задолго до среднего эоцена (поэтому они успели разделиться на множество таксонов), либо что они возникли и дивергировали за пределами Африки (возможно, в Азии) и затем уже в течение среднего эоцена произошло заселение Африки различными их группами.Таким образом, Ливия может стать источником новых уникальных данных о ранней эволюции приматов, причем самых разных их ветвей. Ранее из Ливии была известна примитивная мартышка Prohylobates simonsi из нижнего миоцена. Осталось найти там еще австралопитеков, и Ливия вырвется в страны-лидеры по ископаемым приматам.
Сходство афротарзиусов с эосимидами позволило классифицировать их как антропоидов. К египетскому виду предположительно относилась большая берцовая кость, на которой есть следы прирастания малой берцовой. Эта черта была сильным аргументом принадлежности афротарзиуса к долгопятовым, однако сопоставление изолированных зубов и костей всегда спорно. Есть даже большие сомнения – действительно ли этой костью обладал примат, а не какое-то другое животное? Строение же зубов Afrotarsius скорее антропоидное, чем долгопятовое.
Афразия вылазила до Африки из Азии
Совсем недавно международная группа ученых описала четыре новых зуба, на сей раз найденных в Бирме (Chaimanee et al., 2012). Казалось бы, четыре зуба – не столь уж велика куча, но уж больно удивительна география находок! Дело в том, что в эоцене, 37 млн лет назад, Северная Африка и Юго-Восточная Азия были разделены широким морем Тетис. На месте нынешних Ближнего и Среднего Востока плескались волны тропического моря, лишь местами разбивавшиеся о берега островов. Азия была напрочь отрезана от Африки, однако же приматы этих континентов удивительно схожи! Бирманскую обезьяну исследователи назвали афразией Afrasia djijidae. Тонкости строения зубов и сравнение их по примитивности – прогрессивности позволяют предположить, что первые подобные приматы возникли в Азии, но именно во время существования афразии переселились в Африку, дав там род Afrotarsius. Более того, авторы исследования утверждают, что многочисленные более поздние североафриканские примитивные антропоиды могли возникнуть тоже от азиатских переселенцев, но от других, нежели Afrasia, например от амфипитецид. Таким образом, море было вполне преодолимо для обезьян, хотя бы и изредка. Конечно, нельзя было в связи с этим забыть вечную проблему возникновения широконосых обезьян Южной Америки, у которых вроде как есть годные предки, но в Африке. Годами ведется спор – как они переправились с материка на материк. Преодоление Атлантического океана в некотором роде сопоставимо с преодолением Тетиса, хотя современные люди обычно удивляются первому событию, но ничего не знают о втором.Показательно, что не только обезьяны форсировали Тетис в поисках африканской благодати. Примерно в то же время аналогичный путь проделали гистрикоморфные грызуны, антракотерии и некоторые другие звери. Либо палеокарты врут, либо мы сильно недооцениваем живучесть животных и их способность к трансокеанским вояжам.В любом случае с новыми находками систематика приматов стала сложнее (обогатившись не только новым родом-видом, но и целым инфраотрядом Eosimiiformes, причем эосимиды оказались не слишком родственными амфипитецидам, хотя раньше предполагалась их тесная связь), география их распространения усложнилась, а поводов для размышлений добавилось. Таким образом, можно закончить великой банальностью: с новыми находками знаний наших прибывает, но и вопросов прибавляется стократ. И это хорошо!
Амфипитециды (Amphipithecidae): Amphipithecus, Bugtipithecus, Ganlea, Krabia, Myanmarpithecus, Pondaungia и Siamopithecus – приматы, также промежуточные между полуобезьянами и высшими обезьянами. Они известны с границы эоцена и олигоцена (35–31 млн лет назад) Бирмы, Таиланда и Пакистана. Большая часть известных признаков позволяет отнести их к человекоподобным обезьянам, хотя сохранялись и примитивные черты, например три премоляра и несросшиеся половинки нижней челюсти. Размер этих животных был примерно с мартышку: от менее чем полкилограмма до 10 или даже более килограммов – заметно крупнее, чем у более древних антропоидов, да и вообще всех эоценовых приматов. Это были гиганты своего времени.
Внешность этих зверей остается загадкой. К амфипитеку в некоторый момент были отнесены два обломка лобных костей, но пересмотр показал, что они с большой вероятностью вообще принадлежали не то что не приматам, но даже не млекопитающим, они вообще непонятно чьи – даже класс этих животных остается неопределенным (Beard et al., 2005).
Кости прочего скелета тоже под вопросом. В слоях с пондаунгией и амфипитеком найден даже фрагментарный скелет без черепа, но как понять – кому он принадлежал, если виды описывались по зубам? По размерам и хроно-географической логике он мог принадлежать пондаунгии. Кости конечностей обладают смесью признаков нотарктид, адапид, лори, цебусовых широконосых, частично оригинальны, а в целом больше всего схожи с костями адаписовых (Ciochon et al., 2001). Либо скелет амфипитецид был гораздо примитивнее, чем их зубная система, либо верно подозрение, что кости принадлежали на самом деле не амфипитеку, а какому-то крупному сиваладапиду (Beard et al., 2007; Marivaux et al., 2008), тем более что зубы двух видов последних тут найдены. К тому же пяточная кость этого скелета не сходится с изолированной таранной из аналогичных слоев. Таранная тоже сочетает признаки полуобезьян и обезьян: с одной стороны, по совокупности измерений она гораздо ближе к таранным костям адаписовых и лемуров, а с другой – по части описательных показателей похожа на кости широконосых обезьян (Gunnell et Ciochon, 2008). Пока кости скелета амфипитецид не будут найдены вместе с черепом, сомнения о принадлежности этих останков продолжат грызть душу палеоприматологов.
Распределение признаков по видам запутано: таиландский сиамопитек Siamopithecus eocaenus – не самый древний, но наиболее примитивный и самый крупный представитель, тогда как пакистанский Bugtipithecus inexpectans – самый поздний, но один из самых примитивных и самый мелкий амфипитецид. Его размеры оказываются чуть ниже “границы Кея”, что предполагает насекомоядность. Более острые и гладкие бугорки зубов указывают на то же. Для большинства амфипитецид была характерна диета из орехов, семян, фруктов и листьев: она устанавливается по строению зубов, а также по значительному развитию нижних челюстей в высоту, очевидно, нагрузки на них были большими (Ramdarshan et al., 2010).
Клыкастая ганлея – гроза орехов
Центральная Бирма богата на находки амфипитецид. Она уже подарила миру собственно самого амфипитека Amphipithecus mogaungensis, самого древнего представителя семейства – мьянмарпитека Myanmarpithecus yarshensis, а также первого из описанных и самого изученного – пондаунгию Pondaungia cotteri. Но открытия продолжаются. В 2009 году был описан новый род и вид – ганлея Ganlea megacanina (Beard et al., 2009). Это была некрупная – 2,3 кг – по амфипитековым меркам обезьяна. Самая выдающаяся (в буквальном смысле) черта ганлеи – огромные нижние клыки. Но не стоит представлять себе оскаленную обезьяну с капающей с клыков кровью. Клыки ганлеи были сильно расширены и уплощены спереди назад, а верхушки их стерты. Больше всего такое строение напоминает резцы грызунов. Кстати, у очень похожего мьянмарпитека и чуть более отличающейся пондаунгии клыки сношены схожим образом. У пондаунгии эмаль моляров толстая и морщинистая, с низкими гребнями, что свидетельствует о питании слабоволокнистой едой. У амфипитека и пондаунгии к тому же верхние резцы широкие и уплощенные, с усиленными корнями, явно приспособленные к раскусыванию какой-то не самой мягкой пищи. Вероятно, эта группа животных была адаптирована к питанию орехами и твердыми семенами. Из современных приматов на них экологически больше всего похожи саки – широконосые обезьяны. Замечательно, что и по многим другим особенностям строения зубов амфипитековые оказываются ближе к южноамериканским широконосым обезьянам (а также североафриканским проплиопитековым), чем к жившим рядом эосимидам. Означает ли такое сходство истинное родство или лишь конвергенцию? Это пока неясно, тем более что география миграций получается уж очень широкой: азиатские амфипитециды должны были переселиться в Северную Африку, а их потомки – совершить оттуда заплыв до Южной Америки. Зато понятно, что ганлея со своими специализациями вряд ли могла быть предком узконосых обезьян.
Krabia minuta – не краб на минуту, а обезьяна на века
Krabia minuta – так названа новая обезьяна, жившая примерно 34 млн лет назад, на границе эоцена и олигоцена, на территории нынешнего Таиланда (Chaimanee et al., 2013). Ее окаменевшие зубы были откопаны трудолюбивыми тайскими руками в отложениях формации Краби, а малые размеры зубов послужили основанием псевдохронометрического видового наименования. Но, конечно, не в названии интерес находки.Крабия – замечательный примат. Большая часть ее особенностей позволяет включить крабию в семейство амфипитецид. Однако ряд признаков зубов ставит ее особняком. Думается, не стоит утомлять ни в чем не повинного Читателя крайней бунодонтностью, сильным развитием лингвального цингулюма и отсутствием буккального, отсутствием гипоконусов и периконусов, редуцированными тригонами, слабыми парастилями и метастилями. Даже прожженные палеоприматологи не знают, что значило в жизни древней обезьяны наличие или отсутствие того или иного бугорка или бороздки. Для ряда признаков смысл более-менее очевиден (например, острые бугорки типичны для насекомоядных, а тупые – для растительноядных приматов), но у многих особенностей глубокого смысла может и не быть вообще. Однако это не мешает палеонтологам тасовать признаки, жонглировать ими в многомерных анализах, оценивать их таксономическую значимость и строить на их основании развесистые филогенетические схемы.Крабия жила в сообществе других приматов – как минимум пяти видов. Кроме нее, по деревьям тех же лесов древнего Таиланда скакали адапис Muangthanhinius siami, сиваладапид Wailekia orientale, амфипитецид Siamopithecus eocaenus, некий мелкий амфипитецид и некий долгопятоподобный зверек, остающийся пока безымянным. Среди этого зоопарка крабия существенно выделялась своими зубами. Главное – сам факт ее сильного отличия. Он свидетельствует о далеко зашедших процессах дифференциации и, стало быть, – длительности эволюции амфипитецид в Азии. Также крабия имеет некоторые черты, связующие ее с более древними азиатскими эосимидами; это важно, ибо доныне между амфипитековыми и эосимидами промежуточные формы были неизвестны. Одновременно особенности крабии сближают ее с африканскими ранними обезьянами и даже современными широконосыми, живущими вообще в Южной Америке. Понятно, что, будучи на несколько миллионов лет моложе африканских проплиопитековых и парапитековых, крабия не может быть их прямым предком, но коли у нее есть признаки древних азиатских эосимид, сама она относится к амфипитековым и одновременно похожа на африканских проплиопитековых, то в сумме это значит, что проплиопитековые возникли от амфипитековых, которые, в свою очередь, произошли от эосимид. Азия побеждает!Конечно, есть но.Во-первых, чехарда датировок. Крабия, как уже сказано, не может быть прямым предком африканских обезьян, поскольку моложе их. Она только демонстрирует связь разных групп через сохранение древних признаков, усложненных позднейшими специализациями.Отсюда вытекает “во-вторых”: некоторые из этих специализаций сами по себе исключают крабию из числа Великих Предков. По некоторым чертам лучше подходит на роль связующего Азию и Африку звена пакистанский амфипитецид бугтипитек Bugtipithecus inexpectans, но он жил вообще в нижнем олигоцене – еще позже крабии. Таким образом, амфипитековый предок африканских обезьян должен был жить в начале или, в крайнем случае, в середине эоцена. Таковой еще никем не найден.В-третьих, сходства крабии с африканскими древнейшими и современными обезьянами могут быть вообще конвергентными. В таком случае нет смысла выводить африканских обезьян из азиатских, у них могут быть собственные автохтонные предки. Такую точку зрения развивают палеоприматологи, изучающие североафриканские находки. Обилие и разнообразие африканских, европейских и североамериканских палеоценовых и раннеэоценовых полуобезьян позволяет сделать широкий выбор возможных предков.Наконец, в-четвертых, искомые Великие Предки вообще не обязаны были жить именно там, где мы находим больше зубов. То, что костеносные отложения сохранились и найдены в Бирме, Китае, Таиланде, Пакистане, Египте и Ливии, не говорит о том, что только там жили древние приматы. Великой Прародиной вполне могла быть, например, Индия, которая к этому времени как раз доплыла до Азии и воткнулась в нее. Проникшие туда приматы запросто могли в новых условиях и при незанятости экологической ниши совершить эволюционный рывок. А ведь еще есть Центральная Африка – terra incognita эоценовой палеоприматологии, есть Западная Африка, Восточная, Южная, наконец.Разнообразие древних приматов было несравнимо большим, чем нам известно. Раскопки продолжаются, новые зубы (а то – чем тафономия не шутит – и целые скелеты!) расскажут новые истории.Сколь много обезьянок древних готовит двадцать первый век….
Последние амфипитециды дожили до начала олигоцена в Пакистане, хотя известны там лишь по восьми зубам, описанным как бугтипитек Bugtipithecus inexpectans. Впрочем, этот вид уж очень специфичен, по части признаков он похож на Plesiopithecus teras, который сам по себе загадочен, по части занимает промежуточное положение между эосимидами и амфипитецидами, хотя моложе тех и других (Coster et al., 2013). Не исключено, что бугтипитек представляет самостоятельную экзотическую линию антропоидизации омомисовых обезьян.
Наступал олигоцен. Эта засушливая эпоха ставила свои задачи, ответы на которые находили уже новые приматы…
Парапитековые (Parapithecoidea): Abuqatrania, Apidium, Biretia, Lokonepithecus, Parapithecus, Qatrania, Serapia, Moeripithecus и Arsinoea – полностью вымершая группа из позднего эоцена и раннего олигоцена (39–30 млн лет назад) Северной Африки, известна почти исключительно из Файюмского оазиса в Египте (только самая ранняя биретия Biretia piveteaui происходит из Ливии, а самый поздний локонепитек Lokonepithecus manai – из Кении). Наряду с эосимидами, афротарзиидами и амфипитецидами они были участниками третьей радиации приматов и с большой вероятностью явились предками последующих обезьян-антропоидов. Как и рассмотренные выше родственники, парапитековые имели смесь примитивных и продвинутых признаков. Причем распределение этих признаков довольно любопытное: ранние парапитековые не имеют ярких специализаций зубов и челюстей, а поздние их приобретают и становятся похожи на более древних эоценовых амфипитецид Азии. Такое хронологически нелогичное распределение, скорее всего, свидетельствует о конвергенции азиатских и североафриканских групп.
В отличие от азиатских примитивных антропоидов, парапитековые известны не только по зубам, но и полноценным черепам. В нашем распоряжении есть реконструированное лицо апидиума Apidium phiomense и целый череп более крупного парапитека Parapithecus grangeri. Внешне они напоминали современных мармозеток. Одной из самых интересных черт является очень маленький размер мозга: 11,4 см³ у парапитека. Это, конечно, больше, чем даже у более крупных адаписовых, но примерно столько же, как у аналогичных по размеру потто и лори, заметно меньше, чем у современных широконосых, и намного меньше, чем у узконосых. Относительная масса мозга получается совершенно “полуобезьянья”. Обращают на себя внимание крупные обонятельные луковицы: они, конечно, на минимальном пределе развития у полуобезьян, зато намного больше, чем у обезьян. Третий примитивный признак – размер зрительного канала, через который проходит зрительный нерв. У современных человекоподобных обезьян нерв и канал заметно толще, нежели у полуобезьян, а у парапитека оказывается ровно средним. Выходит, что мозг первых антропоидов развивался отстающими темпами в сравнении с зубами (Bush, 2004; Bush et al., 2004).
Уголок занудства
Сравнительно с более поздними антропоидами, апидиум и парапитек имели более узкую мордочку, их глазницы были еще в значительной степени развернуты вбок, а не смотрели строго вперед. Строение слуховой области, столь важной для систематики, у парапитека в наибольшей степени похоже на мелких современных широконосых и несколько примитивнее (что закономерно), чем у более позднего и сравнительно продвинутого Aegyptopithecus: барабанная пластинка окружает слуховой проход, но сохраняются уплощенные слуховые капсулы, образованные каменистой костью. Неожиданно продвинутой чертой является полное закрытие задней стенки глазницы у парапитека, так что лобная кость контачит с большим крылом клиновидной: широконосые обезьяны имеют в этом месте щель, а подобное парапитековому строение типично для узконосых. У апидиума, впрочем, эта область была схожей с таковой у цебусовых широконосых.Половинки нижней челюсти парапитековых срастаются (впрочем, у стоящей несколько особняком арсиноэи Arsinoea kallimos симфиз нижней челюсти несросшийся). Большинство парапитековых обладало тремя верхними и тремя нижними премолярами, но Parapithecus fraasi – только двумя. Parapithecus grangeri обладал странной и уникальной для приматов специализацией: у него были очень маленькие верхние резцы, а постоянных нижних не было вовсе, так что клыки оказывались самыми передними зубами и сходились между собой. При этом молочные нижние резцы у него наличествовали, а родственный и синхронный вид Parapithecus fraasi имел и постоянные.
Парапитеки были мелкими обезьянками: от 126–242 г для Qatrania wingi до 559–1029 г для Serapia eocaena и несколько больше килограмма (по самой смелой оценке – до 3 кг) для Parapithecus grangeri, то есть крупнее долгопята, но меньше карликовой мартышки. От апидиума сохранилось довольно много останков, что позволило реконструировать целый скелет, от парапитека тоже есть кости конечностей. Их анатомия ясно свидетельствует о древесном образе жизни, четвероногой ходьбе, беге и прыгании по ветвям без подвешиваний на руках и вертикального лазания. Больше всего скелет парапитековых похож на скелет омомисовых и маленьких широконосых обезьян, но отличается от полуобезьян и узконосых. В частности, большая берцовая кость имела прочное соединительнотканное соединение с малой берцовой, подобное тому, что наблюдается у мышиных лемуров и многих широконосых. Внутренняя и наружная лодыжки были крупными, ограничивающими движения голеностопного сустава передне-задним направлением, что способствует силе и точности прыжков. При малых размерах тела такое строение вполне закономерно.
В отличие от предшественников и живших в то же время полуобезьян, чьи гастрономические пристрастия были весьма разнообразны, все парапитековые были фруктоядами, хотя некоторые разнообразили свое меню древесными смолами, а Parapithecus grangeri с большой вероятностью перешел на листоядность. Показательно, что 34 млн лет назад в Египте исчезли крупные фрукто– и листоядные полуобезьяны, эти ниши окончательно заняли антропоиды и сообщество африканских приматов приняло более-менее современный облик (Kirk et Simons, 2001). Впрочем, парапитековые на то и “пара-”, что еще не были совсем “-питеками”. Для того чтобы стать окончательными и безоговорочными обезьянами, им надо было продолжать развитие.
Великолепная миграция: широконосые обезьяны
Широконосые обезьяны (Platyrrhini) – обезьяны Центральной и Южной Америк. По сравнению с узконосыми, широконосые сохранили ряд примитивных, почти полуобезьяньих признаков (например, слуховую капсулу, не полностью замкнутую глазницу, три премоляра), но компенсировали их редкостными специализированными чертами вроде хватательного хвоста и исчезновения третьих моляров у многих видов. Впрочем, для того чтобы распознать широконосую обезьяну, необязательно заглядывать ей в рот и считать зубы, можно узнать ее в лицо: носовая перегородка у нее будет толстая, а ноздри широко разнесены одна от другой и расставлены в стороны. Узконосые же обезьяны называются так не за габариты всего носа, а только за тонкую носовую перегородку; у какой-нибудь гориллы сам нос может быть сколь угодно широченный, но ноздри смотрят в одну сторону и расположены рядышком.
Возникновение широконосых весьма загадочно. Проблема в том, что Южная Америка большую часть своей истории была гигантским островом, полностью отделенным морями от прочих материков. Иногда она соединялась либо с Антарктидой (которая периодически бывала вполне теплым местом), а через нее – с Австралией, либо с Северной Америкой, а через нее – с остальным миром. Однако как-то так сложилось, что предки приматов в Южную Америку в палеоцене и первой половине эоцена не попали. А вот в середине или к концу эоцена – около 40 млн лет назад – они тут появляются, вдруг и без объявления войны. А ведь именно в это время Южная Америка находилась в очередном длительном одиночном заплыве. Ближайшей землей была Северная Америка, но самыми поздними известными оттуда приматами являются омомиды Rooneyia viejaensis и Macrotarsius montanus, имеющие в самом оптимистичном варианте датировку порядка 34 млн лет назад. Однако дело ведь не только в хронологии и географии: позднейшие североамериканские омомиды были весьма специализированными животными, из которых вывести широконосых довольно затруднительно. Конечно, всегда остается вероятность, что мы чего-то не знаем и переходные от омомисовых к широконосым пережили “темные века” на каком-нибудь острове в районе нынешней Панамы. Но тем наука и отличается, что оперирует фактами, а не догадками.
А факты таковы, что в Северной Африке в эоцене и олигоцене известно множество приматов, о части которых уже говорилось выше, в признаках которых там и сям проглядывают явственные черты широконосых. То есть получается замечательная ситуация: достаточно очевидные предки есть в Африке, а потомки – в Южной Америке. Вопрос: как они преодолели Атлантический океан? Потенциально это можно сделать несколькими способами. Во-первых, можно двинуться через Европу и цепочку островов в Северную Америку, а оттуда через Панамский пролив в Южную Америку. Во-вторых, из Африки можно пойти через Азию и Берингов пролив опять же в Северную Америку, а оттуда в Южную. Оба эти варианта неправдоподобны, поскольку предполагают слишком далекие миграции по областям, где по факту раннеолигоценовых приматов мы не находим; а Берингия к тому же совсем неподходящее для приматов место. В-третьих, из Африки теоретически можно добраться до Антарктиды, которая периодически соединялась с Южной Америкой. Этот путь не такой сумасшедший, как может показаться на первый взгляд, поскольку, как уже говорилось, климат в Антарктиде иногда был вполне тропический, а сама она соединялась с Южной Америкой. Главной проблемой были не морозы и айсберги, а морские просторы от Африки до Антарктиды. Мыс Доброй Надежды неспроста называется также Мысом Бурь; хотя в эоцене ситуация с ураганами могла быть несхожей с нынешней, все же вряд ли было какое-то течение, идущее тут с севера на юг. К тому же ископаемые сумчатые из Антарктиды известны, а плацентарные – нет.
Наконец, четвертой возможностью является вояж через Атлантику. На первый взгляд он выглядит тоже крайне сомнительным, но стоит учесть несколько важных моментов. Для начала ширина Атлантического океана была заметно меньшей, чем сейчас, ведь Южная Америка все последующее время двигалась на запад. Многим может показаться, что от Северной-то Америки до Южной всяко ближе, чем от Черного континента, но в реальности Северная миллионы лет ползла на юг, ее нынешнее положение сильно отличается от эоценового, так что расстояние от Северной Америки до Южной тогда было едва ли не большим, чем от последней до Африки. Северное и Южное пассатные течения несут воды от Западной Африки до Бразилии; неспроста в парусную эру капитаны по пути в Индийский океан предпочитали вести свои корабли от Европы сначала до Бразилии, а потом сворачивали обратно – уже к Южной Африке и мысу Доброй Надежды. Понятно, что у обезьян нет лодок, но большие реки Западной Африки – прежде всего Сенегал, Нигер и Конго – выносят в море огромное количество упавших деревьев, иногда сплетенных ветвями и корнями в довольно большие плавучие плоты. На этих естественных понтонах случайно оказываются самые разные животные, в первую очередь, понятно, древесные, в том числе обезьяны. Ясно, что долго без пресной воды они вряд ли могут протянуть, но, повторимся, океан был ýже, течение могло быть быстрее, так как на месте Центральной Америки зиял огромный пролив между Атлантическим и Тихим океанами шириной с хорошее море, да и дожди никто не отменял. К тому же никто и не говорит, что такой путь был налаженной торной дорожкой. Судя по морфологии и генетике современных широконосых обезьян, они происходят от одного предка. Не исключено, что плавание через Атлантику обезьяны совершили всего единожды. Теоретически это вообще могла быть одна беременная самка.
Достоверности сей гипотезе добавляет тот факт, что такой же путь, похоже, совершили предки кавиоморфных грызунов и плохо летающих птиц гоацинов – ныне эндемиков Южной Америки, а также амфисбен и гекконов, подходящие предки коих также обнаруживаются в Африке. Особенно интересно, что с большой вероятностью некоторые из этих групп появлялись в Южной Америке неоднократно. Споры об их происхождении не утихают многие годы, но позиция “африканистов” весьма сильна (Симпсон, 1983).
Как бы там ни было, обезьяны достигли своего Эдема. Они попали в замечательное место, где росли огромные деревья, из коры которых сочилась вкусная смола, на которых висели ароматные плоды и по которым ползали аппетитные насекомые. При этом не было ни одного достойного конкурента (южноамериканские грызуны появились примерно тогда же, но и они были почти все наземными, а немногочисленные древесные были медленными и глупыми; об опоссумах и речи нет), разве что птицы; в ветвях не прятался ни один хищник, специализированный на охоте за приматами и вообще за древесными животными, разве что птицы; да и вообще никто не мог сравниться с приматами в красоте и сообразительности, разве что птицы.
В итоге широконосые обезьяны оказались лучшим образцом кустообразной эволюции, когда в условиях незанятости идеальной экологической ниши на огромной территории из минимума предков за короткое время возникает широкий спектр линий, ближайшие из коих весьма похожи, но крайние варианты которых уже заметно отличаются друг от друга. Интересно, что многие древнейшие широконосые уже очень похожи на современных представителей, что свидетельствует о большой скорости первичного видообразования. Такое положение крайне осложняет систематику широконосых обезьян, поскольку очень трудно провести границы таксонов среди множества схожих родов и видов. В опубликованных таксономических схемах перебраны, кажется, все возможные сочетания семейств и подсемейств и все варианты отнесения подсемейств к тем или иным семействам, которых выделяют от двух до десяти; при этом обычно почти каждый род получает статус самостоятельного подсемейства. Даже молекулярная систематика в такой ситуации не то чтобы пасует, но не имеет преимущества перед классическими морфологическими методами установления родства.
Примерно 30 млн лет независимой от обезьян Старого Света эволюции дали ряд любопытных линий конвергенции, а также своеобразные группы, аналогов которым в Африке и Азии нет.
Древнейшие приматы Южной Америки долгое время были известны из Бразилии, но новейшие открытия сделаны в Перу, в отложениях, возраст которых с наибольшей вероятностью верхнеэоценовый, а то и финально-среднеэоценовый, возможно, древнее 40 млн лет и на десяток миллионов лет старше бразильских. В местонахождении Санта-Роза найдены четыре зуба приматов, из которых, правда, хорошо сохранился только один, а три прочих обломаны, так что исследователи не стали давать им формальные названия, чтобы не затруднять работу в будущем, когда появятся более полные находки.
Самый же сохранный зуб получил наименование перупитека Perupithecus ucayaliensis (Bond et al., 2015). Этот примат оказался очень примитивным – настолько, что его даже нельзя уверенно отнести к широконосым обезьянам. Он явственно архаичнее всех древних и тем паче современных широконосых, зато больше всего похож на зубы талапитека Talahpithecus parvus, который жил в Ливии тоже в конце эоцена (датировка его несколько спорна – 35–36 или 38–39 млн лет назад). Талапитек сам имеет не вполне понятное систематическое положение, так как описан всего по трем зубам, из коих два сломаны. Первоначально он был определен как олигопитековый примат, а олигопитеки – примитивнейшие обезьяны, в целом годящиеся в предки как широконосым, так и узконосым. Есть, правда, одна проблема: у олигопитековых на челюстях сохранилось только по два премоляра, а у широконосых их три. Сколько было премоляров у талапитека и перупитека – неизвестно, но в египетском Файюме есть находки похожих на талапитека обезьян с тремя премолярами. Таким образом, североафриканские корни южноамериканских широконосых вырисовываются с полной определенностью. Самое главное – нигде в мире больше нет более подходящих кандидатов на роль предков широконосых. Перупитек схож с североафриканскими древними приматами, примитивнее всех широконосых и хронологически вписывается аккурат между ними – что еще надо для полного счастья?!
Неожиданно новое открытие дало поддержку, хотя бы и призрачную, для североамериканской версии заселения Южной Америки. До сих пор считалось, что миграция совершилась около 26–27 млн лет назад, тогда как в Северной Америке позднейший примат жил 34 млн лет назад. С открытием же перупитека этот разрыв исчез! Впрочем, как уже сказано, этот аргумент очень слаб, ибо морфология важнее чисто хронологических соображений. В Северной Америке жили только полуобезьяны и омомисовые – довольно специализированные родственники современных долгопятов. Из них крайне проблематично вывести южноамериканских широконосых, а конкретно перупитек совсем на них не похож.
Так что теперь мы можем быть почти на 100 % уверенными (насколько в науке бывает 100 %-ная уверенность), что широконосые возникли из североафриканских антропоидов.
Открытие в Санта-Розе замечательно еще по двум причинам. Во-первых, судя по форме и размерам, три неназванных зуба могут представлять еще два вида, а то и рода приматов. Таким образом, уже в конце эоцена в Америке жили три вида обезьян! Из дополнительных зубов нижний моляр выглядит еще примитивнее, чем зуб перупитека, по признакам он занимает промежуточное положение между амфипитековыми и парапитековыми с одной стороны и широконосыми – с другой. Во-вторых, внимательный глаз бывалого географа может узреть, что Перуанская Амазония находится на прямо противоположном от атлантического побережья конце континента. То есть чтобы приплывшие из Африки приматы оказались у подножия Анд, им надо было пересечь весь материк. Думается, на это ушло какое-то время, очевидно, оно не было потрачено впустую, и по пути успели сформироваться несколько видов. Есть также вероятность, что приматы заселяли Южную Америку неоднократно (несколько миграций предполагалось также для южноамериканских грызунов, амфисбен и гекконов), этим можно объяснить довольно резкое отличие приматов Санта-Розы от более поздних широконосых и отсутствие находок в промежуточные этапы. Впрочем, для таких допущений нет особых оснований: за 10 млн лет признаки как раз должны были существенно трансформироваться, а находок в интервале от 40 до 26 млн нет, так как нет соответствующих местонахождений.
На что были похожи первые обезьяны Южной Америки – понять по зубам трудно. Можно лишь констатировать, что перупитеки были мелкими, вроде современных каллимико или тамаринов, причем неназванные виды были еще на треть меньше. Судя по строению зубов и их размерам, их обладатели были насекомо– и фруктоядными.
Кстати, о судьбе…
Местонахождения ископаемых приматов, к сожалению, весьма редки. Обезьяны – животные древесные, а отложения почти всегда формируются в воде. Чтобы кости примата попали в палеонтологическую летопись, должно свершиться необычайное стечение обстоятельств: зверь должен или умереть около воды, или быть унесенным потоком, или упасть в воду и утонуть; тело не должны растащить и разгрызть хищники и падальщики, кости должны быстро покрыться слоем осадков, а вплоть до современности отложения не должны разрушиться в пыль; наконец, исследователи должны эти слои и кости найти, собрать, распознать и описать. Понятно, что каждый шаг на этом тернистом пути случайностей весьма маловероятен. Обезьяны очень редко падали в воду, крокодилы и пираньи не зевали, превратности геологической судьбы миллионы лет рушили напластования, палеонтологов мало, а планета велика, да и не везде реально производить раскопки по миллиону причин. Нам невероятно повезло, что есть вообще хоть какие-то местонахождения, содержащие останки приматов, хотя бы отдельные зубы, и уж тем более фантастично, что существуют сокровищницы, даже богатые обезьянами.Так что коли не судьба, то уж не судьба, а уж коли повезло – так надо радоваться!

Рис. 32. Branisella boliviana.
Собственно сами широконосые достоверно известны из верхнего олигоцена и нижнего миоцена. Древнейшей обезьяной Южной Америки, от которой сохранилось много останков, была браниселла Branisella boliviana, найденная в Боливии и имеющая датировку 26–27 млн лет назад. В 1991 году из тех же слоев был описан Szalatavus attricuspis, но, видимо, в реальности это та же браниселла. Показательно, что из всего бестиария ископаемых широконосых браниселла едва ли не единственная, не имеющая очевидной близости к какому-либо современному подсемейству, зато подходящая на роль общего предка. Всего на миллион лет моложе аргентинский Dolichocebus gaimanensis, но он уже может быть определен как капуцин – дифференциация началась.
Кстати, о древности…
Датировка южноамериканских фаун – непростое дело. До появления и широкого использования радиометрических методов датировки мнения о возрасте тех или иных слоев выдвигались достаточно произвольно. В Старом Свете обычно есть возможность прокоррелировать слои из разных местонахождений, сравнить виды и прикинуть как минимум последовательность и относительный возраст отложений. Так и были созданы сами понятия эр, периодов и эпох, и описаны они были именно на основе европейских фаунистических комплексов. Однако южноамериканские животные столь сильно отличаются от европейских и даже североамериканских, что геохронологические шкалы, построенные для Южной Америки, оказываются “вещью в себе”. С последовательностью еще как-то разобрались, а вот с абсолютным возрастом долго не могли. Конечно, это привело к перекосам, причем преимущественно в сторону удревнения по сравнению с реальностью. В немалой степени этому способствовала архаичность южноамериканских зверей.Например, слои “сантакрусия” в аргентинской Патагонии сначала считались верхнеэоценовыми, потом финально-олигоценовыми, потом нижнемиоценовыми и, наконец, даже верхнемиоценовыми, то есть разброс был в пределах половины всего кайнозоя! Абсолютные датировки расставили все на свои места: сантакрусий соответствует все же нижнему миоцену. А ведь именно в этих отложениях были найдены останки гомункулюса Homunculus patagonicus: одно дело, если он синхронен адаписовым и омомисовым приматам, и совсем другое – проконсулам или сахелянтропам, есть же разница! Сейчас мы знаем, что гомункулюс жил в одно время с проконсулами.
В отличие от более древних приматов, широконосые известны по весьма полным находкам, в том числе немалому числу целых черепов и нескольким скелетам. Поэтому мы точно знаем, что ископаемые широконосые принципиально не отличались от современных. Все они, без единого исключения, были древесными животными. Почти все были мелкими, как и нынешние их потомки, среди которых самые тяжелые – ревуны – редко превышают 12 кг. Только позднеплейстоценовые бразильские протопитек Protopithecus brasiliensis, картеллес Cartelles coimbrafilhoi и кайпора Caipora bambuiorum весили порядка или более 20 кг – в полтора раза тяжелее самых откормленных современных широконосых (раньше их реконструировали даже до 25–35 кг, но такие оценки наверняка завышены: Halenar, 2011).
Уголок занудства
Как уже говорилось, даже первые представители широконосых весьма похожи на современные формы и, очевидно, были их прямыми предками. Например, колумбийская Stirtonia и бразильский Cartelles сильно напоминают современных ревунов, колумбийский Miocallicebus – обезьян-прыгунов, аргентинский Tremacebus – ночных обезьян мирикин, бразильские Caipora, Solimoea и Protopithecus – паукообразных обезьян, аргентинский Dolichocebus, чилийский Chilecebus, колумбийская Laventiana, бразильский Acrecebus и панамский Panamacebus transitus – капуцинов, перуанская Canaanimico, аргентинские Mazzonicebus, Soriacebus, Propithecia, Carlocebus и Homunculus, колумбийские Mohanamico, Nuciruptor и Cebupithecia – уакари, аргентинская Killikaike и колумбийская Neosaimiri – беличьих обезьян саймири, колумбийская Cebupithecia и аргентинский Carlocebus – саки, колумбийские Lagonimico, Micodon и Patasola – игрунок.Возможно, уже в середине миоцена появляются современные роды, по крайней мере, в Бразилии из этого времени описан Cebus macrognathus.
Когда Северная Америка соединилась с Южной, по Панамскому перешейку на юг хлынули орды новых животных: оленей, верблюдов, саблезубых тигров, медведей. На север миграции тоже были, но гораздо скромнее. Среди гигантских ленивцев, броненосцев и опоссумов были и приматы. Будучи тропическими животными, слишком далеко они, конечно, не продвинулись – их остановили степи и пустыни Северной Мексики, но Центральную Америку заселили плотно. Первым поспешил перебраться на новые земли Panamacebus transitus – около 20 млн лет назад, даже еще до полного соединения материков, когда между ними оставался небольшой пролив.
Еще интереснее, что обезьяны умудрились попасть на все крупные Антильские острова. Как эти древесные животные доплыли до них – остается загадкой, впрочем меньшей, чем тайна проникновения приматов в саму Южную Америку. Течения исправно выносят из Ориноко и других рек немалое число деревьев, а Гвианское течение влечет их на север – как раз вдоль Малых Антильских островов к Большим. Первым большим портом был Гаити, откуда известны плейстоценовый Insulacebus toussaintiana, родственный ночным обезьянам, а также прыгун Antillothrix bernensis, чьи субфоссильные останки датированы вообще голоценом (позднейшая дата – 3850 лет назад). Западнее лежит Куба. В миоцене тут жила родственница прыгунов паралуатта Paralouatta marianae, в плейстоцене ее сменила P. varonai. Интересно, что древнейшая паралуатта имеет очень крупные по широконосым меркам размеры, это как бы пример островного гигантизма. “Как бы” – потому что реальный размер был, конечно, не слишком значительным, примерно как у современных крупных ревунов. До совсем недавнего времени на Кубе жили буроголовая коата Ateles fusciceps, сейчас обитающая в Панаме, Колумбии и Эквадоре, A. anthropomorphus и, вероятно, еще один вид паукообразных обезьян.
Дальше всего забрался ксенотрикс Xenothrix mcgregori – он известен из плейстоцена Ямайки. Обращает на себя внимание отсутствие третьих моляров – как верхних, так и нижних, что вроде бы роднит ксенотрикса с игрунками. Однако, скорее всего, сие сходство сугубо конвергентно, так как подавляющая часть признаков достаточно специфична (благодаря чему обычно выделяется самостоятельное подсемейство Xenotrichinae, иногда даже признаваемое семейством) или сближает ксенотрикса с прыгунами.
Родство многих островных видов остается под вопросом. Подробные исследования показали, что Antillothrix, Paralouatta и Xenothrix весьма родственны между собой и ближе всего к современным прыгунам Callicebus (Horovitz et MacPhee, 1999). Это может значить, что до островов добрались далеко не все широконосые; в отсутствие конкуренции прыгуны дали свой всплеск видообразования (выражаясь умно – адаптивной радиации) с появлением конвергентных форм: Antillothrix разными чертами был несколько похож на саймири, капуцинов и игрунок, крупная Paralouatta – на ревунов, Xenothrix – на уакари. Впрочем, как уже было помянуто, до Кубы добрались и паукообразные обезьяны.
Любопытно, что теоретически обезьяны могли пропутешествовать и дальше: морские течения несут воды от Кубы до Флориды, где для приматов условия хоть и неидеальные, но терпимые. Да и расстояние там явно не больше, чем от Африки до Бразилии. Почему не осуществилась такая миграция – неизвестно. Скорее всего, просто по случайности, ведь не каждый день обезьяны совершают трансокеанские круизы на бревнах. Может, сказалось отсутствие очень крупных рек на Кубе или кипарисовые болота Флориды оказались все же слишком экзотическим местом для приматов.
Разнообразие широконосых обезьян очень велико, выделяется множество родов и видов, но систематика их очень запутана, единства среди антропологов по этому вопросу нет. Чаще выделяют семейства цебусовых и игрунковых.
Цебусовые (Cebidae) покрупнее, их размер колеблется от масштаба крысы до средней собаки. Это практически полностью древесные животные, на землю они почти не спускаются. В связи с этим хвост у многих цебусовых – паукообразных, ревуновых и капуциновых – приобрел хватательную функцию и действует как пятая конечность. Обезьяны могут брать им предметы, а не самые тяжелые паукообразные даже подвешиваются на нем без помощи рук и ног. На нижней стороне кончика хвоста нет шерсти, зато имеются папиллярные узоры, подобные кожным узорам на ладонях, стопах и пальцах. Рисунки линий индивидуальны, потенциально обезьян можно определять по отпечаткам хвоста; впрочем, зверюшки эти законопослушны, злостных правонарушений не совершают, так что надобности в “каудоскопии” нет.
Едят цебусовые преимущественно фрукты и листья, но и любую животную пищу, какую могут найти. Широконосые – дневные животные, кроме нескольких видов дурукули Aotus (они же мирикини или ночные обезьяны, иногда выделяемые в особое подсемейство Aotinae), являющихся единственными ночными обезьянами, а также родственных им прыгунов-тити Callicebus (Callicebinae), ведущих сумеречный образ жизни. Большие глаза дурукули делают их похожими на лемуров или сов, откуда происходит еще одно их название – совиные обезьяны. Прыгуны крайне разнообразны внешне, некоторые очень красивы. Они любят сидеть рядышком на ветке, трогательно переплетясь хвостиками. Между прочим, воротничковый прыгун, или белорукий тити-вдовушка, Callicebus torquatus имеет самый маленький среди приматов кариотип из 16 хромосом в диплоидном варианте.
Среди подсемейства цебусовых (Cebinae) собственно цебусы или капуцины Cebus имеют самый большой относительный размер мозга и сложное поведение. Как в неволе, так и в природе они используют простейшие орудия труда, например раскалывают орехи или раковины моллюсков о камни. Капуцины больше, чем другие широконосые, потребляют животную пищу. Сообразительность и живой нрав этих зверюшек обеспечили их успех у дрессировщиков и, в частности, на поприще киноиндустрии: именно капуцин играет обезьянку-привидение в фильме “Пираты Карибского моря”.
Ревуны Alouatta (Alouattinae) уникальны своим голосом: с помощью огромного горлового мешка-резонатора они могут кричать на несколько километров. Крики не беспорядочны: обязательно есть обезьяна-запевала, которая задает речовку могучему хору. Вся анатомия головы ревунов подчинена вокальной мании, особенно у взрослых самцов: нижняя челюсть с широченными восходящими ветвями, подъязычная кость имеет вид черпака, горло огромных размеров, а борода еще больше увеличивает визуальный эффект. Во время крика ревуны характерно, очень сосредоточенно и весьма брутально вытягивают губы трубочкой, чтоб, не дай бог, не потерять еще пару децибел. Кроме того, у ревунов указательный палец кисти работает вместе с большим, но отставляется от прочих – уникальный для приматов вариант.
Паукообразные обезьяны (Atelinae): коаты Ateles, шерстистые обезьяны Brachyteles и Lagothrix – имеют крайне вытянутые руки и ноги и длинный цепкий хвост. Пальцы на руках образуют своеобразный крючок, а большой палец редуцируется до маленького бугорка или вообще полностью, – это замечательный пример конвергенции с колобусами и гиббонами. Передвигаются паукообразные обезьяны брахиацией, то есть на руках, часто с минимальным участием ног. Конечно, так залихватски, как у гиббонов, у паукообразных скакать не получается, зато помощь цепкого хвоста придает передвижению особое своеобразие. За счет стройности тела относительный вес мозга паукообразных обезьян – один из наибольших среди мало-мальски крупных животных.
Саковые, или чертовы, обезьяны (Pitheciinae) – одни из самых эффектных приматов Южной Америки. Мохнатые саки Chiropotes satanas, Ch. chiropotes и родственные виды имеют окладистую бороду и раздвоенную шапочку на голове, сильно выступающую вперед: такое впечатление, что на них надели странный парик не по размеру. У Ch. albinasus к этой красоте добавляется ярко-розовое лицо. Белолицый саки Pithecia pithecia – вид с самым сильным половым диморфизмом среди приматов: самцы угольно-черные с широким плоским белым или оранжевым лицом, а самки серые с пестринками. Детеныши окрашены как самки, и только с возрастом самцы приобретают свое великолепие. У родственного вида P. monachus разница не так сильна, зато на голове самцов есть шапочка волос в стиле “под горшок”. Существуют и другие, тоже оригинальные виды.
Лысые уакари Cacajao calvus действительно лысые, при этом голова у них воспаленно-красного цвета. На неподготовленных людей они производят впечатление плешивых больных, но нет – так и должно быть, просто понятия о красоте бывают разные. Остальное же тело покрыто очень длинной и красивой шерстью, причем у подвида C. calvus calvus она белоснежно-белая, а C. calvus rubicundus – рыжая. Уакари иначе называются короткохвостыми саки, ведь хвост у них действительно короткий – каких-нибудь 15 см при почти полуметровом размере самого зверя. Редукция связана, как часто бывает у приматов, с крупными размерами и медленным передвижением, причем обычно по нижним ветвям деревьев. C. melanocephalus, наоборот, имеет густую шерсть на голове с начесом вперед: именно родственные виды должны максимально отличаться для избегания межвидового скрещивания.
Саймири, или беличьи обезьяны, Saimiri (Saimiriinae) – самые маленькие цебусовые, примерно четверть метра без хвоста, который в полтора-два раза длиннее тела. Это в своем роде апофеоз примата: мультяшные обезьяны максимально похожи именно на этих жизнерадостных существ с круглыми выразительными глазами, цепкими пальчиками, непрестанно что-то ищущих, исследующих, ковыряющих, бегающих огромными стаями, чирикающих и корчащих друг другу мордочки. Пять видов, из которых наиболее известен Saimiri sciureus, заселяют практически всю Южную Америку и часть Центральной.
Второе семейство широконосых – игрунковые (Callithrichidae), его представители известны также как мармозетки и тамарины. Они отличаются от цебусовых очень мелкими размерами – вплоть до величины мыши: карликовая игрунка Cebuella pygmaea может иметь длину тела всего 10 см (то есть длину пальца), плюс 20 см хвоста, и весить 100 г, хотя обычно они несколько крупнее. Прочие игрунки раза в полтора-два больше, но, конечно, тоже неспроста называются игрунками. Мелкие размеры нынешних игрунок – древнее приобретение. Уже среднемиоценовые Patasola magdalena и Micodon kiotensis имели уменьшенные габариты. Однако синхронный Lagonimico conclucatus весил 1,2 кг – многовато для любой современной игрунки; он же сохранял третьи верхние и нижние моляры, хотя и уменьшенные (Kay, 1994).
Мармозетки имеют только два моляра в каждой половинке челюсти, поскольку размеры черепа крайне малы и третьи просто не помещаются. Ручки настолько крошечные, что игрунки не могут обхватывать ветки пальчиками, поэтому большие пальцы не противопоставляются остальным, а ногти на руках и ногах вторично приобрели вид когтей; только на большом пальце стопы сохраняется плоский ноготь. По стволам эти крошечные зверьки передвигаются наподобие белок, в том числе вниз головой.
Игрунковые обезьяны представляют уникальную форму сообщества, в котором размножается только одна доминирующая самка, а все остальные члены группы, включая самцов, ухаживают за детенышами. Таким образом, мы имеем тут некий аналог социальной структуры общественных насекомых или голых землекопов. Конечно, у игрунок остальные самки не теряют способности к размножению и не превращаются в специализированных рабочих особей, но когда-то и муравьи с чего-то начинали. В отличие от остальных приматов, у мармозеток чаще всего рождается двойня (бывает и один, и три дитятки, но гораздо реже). Не очень понятно, в чем причина этого; есть мнение, что мелкой самке проще родить двух маленьких детенышей, чем одного крупного, хотя логика тут сомнительная, ведь проще-то родить одного мелкого. У многих видов большую часть заботы о малютках берет на себя отец: он сажает их на спину, носит с собой, чистит и оберегает, холит и лелеет. Только для кормления молоком папаша отдает ненаглядных отпрысков маме, но потом ревниво забирает обратно.
Игрунковые делятся на собственно игрунок родов Callithrix (иногда из него выделяются еще Callibella и Mico) и Cebuella, а также тамаринов родов Leontopithecus и Saguinus. Разнообразие тех и других крайне велико: усы и хохлы, гривы и кисточки на ушах, пятна и полосы – все идет в ход для отличия от родственников.
Игрунки питаются древесным соком; для его получения животные надгрызают стволы острыми резцами. Погрызы затягиваются тягучей смолой, которую игрунки тоже едят. В итоге все дерево покрывается овальными наростами; когда на нем не остается живого места, игрунки находят себе новое дерево. Только белоухие игрунки Callithrix aurita не умеют надгрызать кору из-за слишком маленькой мордочки, но все равно основу питания составляет смола и древесный сок, они пользуются наработками других видов. До половины рациона игрунок составляют насекомые, главным образом большие кузнечики и личинки, которых обезьянки постоянно ищут в трещинах коры своими тонкими пальчиками. Не пропускают они фрукты, птичьи яйца и даже птенцов, ящериц и лягушек.
Внешне особенно красива обыкновенная игрунка Callithrix jacchus с пучками белого пуха на ушах и пестрой серо-желтой спинкой; несколько других видов в целом похожи на нее, но, например, у белолицей игрунки C. geoffroyi кисточки ушей серые, а у черноухой C. penicillata – черные и к тому же висячие.
В отличие от игрунок, у тамаринов клыки намного больше, а резцы значительно меньше, а потому они неспособны прогрызать кору деревьев. Тем не менее они тоже любят древесные соки, смолы и нектар, хотя чаще, чем игрунки, ловят насекомых и прочих животных, а также едят фрукты и цветы. Среди тамаринов особенно выделяется императорский Saguinus imperator – с огромными висячими усами а-ля Вильгельм II, только у обезьянки усы висят вниз, а у кайзера торчали вверх. По-королевски выглядит и эдипов тамарин S. oedipus – с белоснежным плюмажем на голове.
Очень впечатляют пышной гривой львиные тамарины, из которых Leontopithecus rosalia выделяется золотым цветом. Другие могут быть контрастны: например, L. caissara имеет золотую шерсть с черными лицом, руками и хвостом, а L. chrysomelas – черную шерсть с золотыми лицом, руками и хвостом.
Гельдиева мармозетка, или каллимико, Callimico goeldii (Callimiconinae) в целом похожа на других представителей семейства, однако имеет три моляра, то есть может представлять потомков исходной для игрунок группы. Она равномерно окрашена в черный цвет.
Амазония велика, а потому раз в несколько лет исследователи находят в сельве новые виды широконосых обезьян. К сожалению, леса безжалостно вырубаются, и многие виды оказываются на грани вымирания. Часть из них, как золотистых тамаринов, удается спасать в зоопарках, но часть может пропасть, не будучи даже открыта. Есть, конечно, и радостные моменты: в 2013 году в Бразилии была вновь найдена мармозетка Марка Callithrix marcai, с 2008 года считавшаяся уже вымершей.
Бывают и “закрытия”. Как-то по чучелу из одного музея был описан особый вид игрунки замечательного пепельного цвета. Однако потом выяснилось, что чучело принадлежало ранее известной мармозетке, шкурка которой просто покрылась густым слоем пыли за долгие годы хранения.
Широконосые обезьяны известны своей склонностью образовывать группы из представителей разных видов, рядом с которыми могут кормиться и не приматы. Такое сообщество оказывается лучше защищено от хищников: например, еноты-носухи лучше нюхают, попугаи – слышат, а обезьяны – видят и соображают. Какой-нибудь онцилле или оцелоту нелегко обмануть сразу всех этих животных. Так что толерантность заложена в глубине души широконосых обезьян; если будущее человечество возникнет из капуцинов, оно наверняка будет терпимее нынешнего.
Новые приматы Старого Света: примитивнейшие узконосые обезьяны
Узконосые обезьяны (Catarrhini) – обезьяны Старого Света: Африки, Азии и Европы. Они появились уже в верхнем эоцене, а на границе с олигоценом стали достаточно разнообразными. Как всегда бывает, переходные группы трудно однозначно классифицировать, так как они зависают между предками и потомками. В данном случае палеоприматологи нашли элегантный выход: примитивнейшие узконосые, которые не могут быть однозначно определены как мартышковые или человекообразные, выделяются в парвотряд “ранних узконосых” Eocatarrhini, тогда как более продвинутые формы, имеющие непосредственное отношение к современным, обозначаются как “истинные узконосые” Eucatarrhini. Маленькая буковка в названии, но огромный скачок в морфологии!
“Ранние узконосые” (Eocatarrhini) сами по себе подразделяются на две группы: более древние и примитивные проплиопитековые (Propliopithecoidea) известны исключительно с границы эоцена и олигоцена Северной Африки (почти исключительно из Египта), а поздние и относительно продвинутые плиопитековые (Pliopithecoidea) – из миоцена Уганды, Европы и Азии.
Проплиопитековые (Propliopithecoidea) имели еще довольно много архаичных черт строения, поэтому их систематический статус неустойчив: разные палеоприматологи предлагали объединять их с парапитековыми или, напротив, включать в состав человекообразных. Самые ранние формы – самые загадочные, что, в общем-то, закономерно.
Олигопитековые (Oligopithecinae), названные по роду Oligopithecus из нижнего олигоцена (31–33 млн лет назад) оазиса Файюм в Египте (предположительно туда же включаются катопитек Catopithecus browni из более старых отложений того же Файюма и еще более древний талапитек Talahpithecus parvus из верхнего эоцена Ливии), разными исследователями определялись как самостоятельное семейство Oligopithecidae надсемейства проплиопитековых (Propliopithecoidea), как подсемейство семейств проплиопитецид (Propliopithecidae) или плиопитецид (Pliopithecidae; в варианте с выделением подсемейств Propliopithecinae, Pliopithecinae и Oligopithecinae), как синоним проплиопитецид или парапитецин (Parapithecinae), включались в понгид (Pongidae) и даже в надсемейство адаписовых (Adapoidea), а также в качестве семейства в инфраотряд Paracatarrhini гипотряда Anthropoidea. Можно надеяться, что новые находки позволят точнее определиться в этом сложном вопросе. В то же время и в том же месте, что олигопитеки, жил странный примат – протеопитек Proteopithecus sylviae, выделенный в собственное семейство Proteopithecidae; впрочем, по общему эволюционному уровню, размерам и образу жизни он не очень отличался от своих соседей.
Очень здорово, что приматы Файюма были массовыми животными, особенно многочисленны были катопитеки, на втором месте по частоте – протеопитеки. От них, да и других приматов сохранились не только зубы, но и черепа и части скелетов. Благодаря такому богатству нам известны многочисленные подробности строения и образа жизни египетских позднеэоценовых и раннеолигоценовых обезьян; ни о каких других синхронных приматах у нас нет столь обширных сведений.
Олигопитековые и протеопитек еще имели некоторые примитивные черты: например, барабанная пластинка у них не полностью окружала слуховой канал. Зато был и прогресс: скажем, в челюстях осталась всего пара премоляров – черта, не очень надежно, но наглядно разграничивающая узконосых и всех прочих приматов. Олигопитековые имели вес от трети до полутора килограммов, вели дневной образ жизни, питались преимущественно фруктами, дополняя их листьями и иногда насекомыми; протеопитек мог вдобавок к тому лакомиться смолами и нектаром.
Рис. 33. Череп Aegyptopithecus zeuxis и реконструкция Propliopithecus.
Любопытно, что половой диморфизм по размерам клыков у катопитека и особенно протеопитека был выражен сильнее, чем у схожих по размеру тела современных обезьян (Simons et al., 1999). Крупные размеры клыков у самцов и мелкие у самок свидетельствуют о существенной межсамцовой конкуренции. Группы катопитеков и протеопитеков состояли либо из нескольких самцов со строгой иерархией и множества самок, либо вообще из одного самца с гаремом самок. Такая структура нетипична для полуобезьян, зато характерна для многих узконосых.
Пяточная кость протеопитека замечательна чудной смесью признаков, одни из которых почти идентичны чертам омомисовых, другие – видимо, конвергентно – мартышковых, а кои – так и вообще адаписовых (Gladman et al., 2013). Очевидно, на этом эволюционном уровне строение конечностей отставало от челюстей и зубов.
Позднейшим олигопитековым был некий неопределенный примат, живший в начале олигоцена в Египте. Он отличался карликовыми размерами – существенно меньшими, чем у Catopithecus и Oligopithecus (Seiffert et Simons, 2013). Трудно сказать, что довело его до жизни такой, а в конце концов и до вымирания.
Кстати, о Файюме…
Файюмский оазис широко известен среди искусствоведов файюмскими портретами, сделанными на крышках саркофагов (кстати, это отличный источник антропологической информации). Но палеонтологи любят Файюм совсем не за это: в нескольких карьерах они нашли и продолжают раскапывать сотни или даже тысячи окаменелостей эоценовых и олигоценовых животных. В Файюме найдены одни из древнейших слонов и слоновых прыгунчиков, даманов и сирен, антракотериев и китов, странных арсиноитериев и еще более странных птолемайид, летучих мышей и грызунов. Нынешняя пустыня 41–29 млн лет назад была цветущим садом, в болотах которого жизнь била ключом. Из Файюма известны самые разные приматы, в том числе предковые для нескольких крупных современных групп.
Проплиопитековые в узком смысле (Propliopithecinae) не слишком разнообразны – Aegyptopithecus zeuxis и четыре вида рода Propliopithecus – и известны только из Файюма в узком хронологическом интервале 29,5–31,5 млн лет назад, но чрезвычайно важны филогенетически. Один клык найден в Анголе.
Сложность разграничения парапитековых и “ранних узконосых” отлично видна на примере оманской находки в местонахождении Таках. Тут был обнаружен обломок нижней челюсти с пятью зубами. Первоначально он был описан как мэрипитек Moeripithecus markgrafi – вид, ранее известный из египетского Файюма (Thomas et al., 1991). Мэрипитек похож на проплиопитека, почему включался в проплиопитековых и даже иногда обозначался как Propliopithecus markgrafi. Зубы из Файюма, описанные как Propliopithecus ankeli, некоторыми палеоприматологами тоже определялись как принадлежавшие Moeripithecus markgrafi. Однако другие исследователи склонны обращать большее внимание на примитивные черты мэрипитека и включать его в парапитековых. В то же время оманская челюсть больше сближается с видом Propliopithecus ankeli, который признается самостоятельным и остается в проплиопитековых. И такая путаница сопровождает исследование почти каждой находки примитивных обезьян с границы эоцена и олигоцена.
Другой пример – алжирипитек Algeripithecus minutus: он описывался как парапитековая или плиопитековая обезьяна, однако с большей вероятностью является адаписовой полуобезьяной (Tabuce et al., 2009).
Из проплиопитековых лучше всего изучен египтопитек Aegyptopithecus zeuxis из олигоцена Египта (29,5–30,2 млн лет назад), от которого сохранились несколько целых черепов и фрагментарные скелеты. На первый взгляд египтопитек напоминал современных макак, благо он практически полностью соответствует им и размерами (2,5–6 кг), и пропорциями. Однако приглядевшись, в строении египтопитека нетрудно заметить массу примитивных черт: задняя стенка глазницы закрыта, да не вполне, барабанная пластинка окружает слуховой канал, но тоже не полностью, – тот и другой признаки развиты примерно как у широконосых обезьян. Совсем подкачал мозг: он у египтопитека вдвое меньше, нежели даже у карликовой мартышки (Radinsky, 1973; Simons et al., 2007). И относительный размер, и пропорции мозга египтопитека вообще оказываются как у мадагаскарских лемуров, хотя участки мозга, отвечающие за зрение, у египтопитека чуть больше, а за обоняние – чуть меньше; впрочем, на фоне современных обезьян эти “чуть” не считаются. Между прочим, это говорит о том, что увеличение мозга у широконосых и узконосых обезьян шло независимыми путями, коли уж общие предки по этому показателю находились на уровне развития полуобезьян. А это, в свою очередь, позволяет надеяться на появление в будущем новых разумных видов.
Судя по маленьким глазницам, египтопитек был дневным, а их разворот вперед говорит о хороших способностях к лазанию по деревьям. О том же свидетельствует строение конечностей: эти животные бегали по ветвям примерно как современные ревуны. В строении конечностей вообще много сходства именно с широконосыми обезьянами: руки и ноги были одинаковой длины, относительно толстые и короткие, неспособные полностью выпрямляться, зато очень цепкие. Далеко прыгать египтопитеки вряд ли умели, но хвост не утеряли, хотя вряд ли он был очень длинным.
Строение зубов свидетельствует, что египтопитеки и проплиопитеки питались почти исключительно фруктами, восполняя недостаток белков листьями. Таким образом, насекомоядное прошлое наконец-то было окончательно изжито.
Обилие черепов позволило оценить половой диморфизм и даже возрастные изменения египтопитеков: самки были существенно – вдвое – меньше и имели меньшие клыки и сагиттальные гребни на черепе. У проплиопитеков разница полов была далеко не такая значительная. С возрастом, как положено, росла морда и увеличивался сагиттальный гребень (Simons, 1987).
Плиопитековые (Pliopithecoidea), вероятно, были потомками проплиопитековых. Эта странная группа процветала от начала до конца миоцена (20–7 млн лет назад) в Китае, Таиланде, Пакистане и Европе, а одна спорная находка сделана в Уганде. Систематика плиопитековых остается предметом бурных дискуссий. С одной стороны, они объединялись с проплиопитековыми, с другой – с гиббонами. В одном из новейших вариантов плиопитековые делятся на дионисопитецид (Dionysopithecidae) и плиопитецид (Pliopithecidae); последние же – на более древнее подсемейство плиопитецин (Pliopithecinae) и более молодое крузелин (Crouzeliinae).
Кстати, о миоцене…
Граница олигоцена и миоцена ознаменовалась существенным потеплением и таянием льдов Антарктиды. Тепло, правда, не вернулось к эоценовому уровню, но все же существенно превосходило современные показатели. Карта мира в целом и Евразии в частности приобрела более-менее современный вид, только юг Европы еще существенно отличался: как бы Средиземное море тянулось аж до середины нынешних Украины и Казахстана, а посреди этих вод возвышались острова, которых не найти на современной карте, в числе коих знаменит Гаргано.Примерно с середины миоцена опять стало холодать, причем ударными темпами, так что с климатической точки зрения границу между эпохами логично было бы проводить именно тут – порядка 12–14 млн лет назад.
Дионисопитециды (Dionysopithecidae) представлены всего тремя нижне-среднемиоценовыми (12–20 млн лет назад) видами из Таиланда, Китая и, видимо, Пакистана – Dionysopithecus orientalis, D. shuangouensis и Platodontopithecus jianghuaiensis, но известны фрагментарные и плохо изученные находки, которые могут представлять иные роды и виды. Сложность, как обычно, в том, что найдены почти исключительно изолированные зубы. Отдельными чертами эти животные уже настолько напоминали человекообразных обезьян, что некоторые палеонтологи объединяют их с проконсулами, хотя в целом они примитивнее плиопитецид. Дионисопитек числится в сонме вероятных предков гиббонов, благо и размером он был похож – около 15 кг. Вместе с тем пяточная кость платодонтопитека довольно примитивна и похожа на кости плиопитеков и проконсулид, а если выбирать между мартышковыми и человекообразными, то на мартышковых. Фаланги платодонтопитека очень вытянуты, по этому показателю их превосходят только гиббоны (Harrison et Yumin, 1999). Дионисопитециды были вероятнейшими предками плиопитецин, которые, таким образом, имели прародину в Азии.
Не исключено, правда, что плиопитековые возникли не в Азии, а в Африке. За это говорит существование ломорупитека Lomorupithecus harrisoni в Уганде 15–20 млн лет назад и возможного плиопитекового в Кении в местонахождении Форт Тернан, правда, уже в конце миоцена 12,5–14 млн лет назад (его зуб был отнесен к Limnopithecus – роду, раньше причислявшемуся к плиопитековым, но ныне считающемуся проконсулидом, то есть человекообразным). Ломорупитек известен по разбитому черепу, в целом похожему на плиопитековых Европы. Его древность – едва ли не старше дионисопитецид – и географическое расположение говорят, что корни плиопитековых могут быть глубже, чем нам известно.
Плиопитецины (Pliopithecinae) сыграли большую роль в истории палеонтологии, поскольку их останки впервые были найдены и описаны еще в середине XIX века. Благодаря этому они вошли во все учебники антропологии, хотя, строго говоря, наверняка были тупиковой группой приматов, имеющей к человеку лишь косвенное отношение. Они известны из Европы и Китая, где существовали не слишком долго – от 17 до 11,6 млн лет назад. Достоверно известен лишь один род Pliopithecus, в котором выделяются шесть-семь похожих видов. Некоторые роды от схемы к схеме кочуют между плиопитецинами и крузелинами.
Плиопитеки из всех ископаемых приматов в наибольшей степени похожи на гиббонов, причем как строением черепа, так и зубов и конечностей. Неспроста самый примитивный европейский вид Pliopithecus piveteaui был даже описан под названием Hylobates antiquus. Впрочем, это не значит, что плиопитеки больше всего похожи на гиббонов из всех современных приматов; при таком ракурсе сравнения плиопитеки сближаются скорее с широконосыми или мартышкообразными. Плиопитеки были довольно резвыми древолазами, но точно не брахиаторами.
Особенно здорово изучен Pliopithecus vindobonensis (15–15,5 млн лет назад), так как в Словакии были найдены многочисленные его останки, а Г. Запфе описал их в монументальном труде (Zapfe, 1960).
Уголок занудства
Плиопитек имел укороченную и суженную морду, сравнительно грацильную мозговую коробку округлой формы с более-менее вертикальным лбом, умеренно развитые височные линии, либо не сливавшиеся, либо образующие низкий сагиттальный гребень. Глазницы довольно крупные, ориентированные почти вперед, но все же несколько вбок, их край утолщен почти в виде кольца. Наглядное отличие от мартышкообразных – широкое межглазничное пространство. Скуловая область очень низкая. Угол нижней челюсти выступает назад. Особенности строения зубов выдают фруктоядность. Примитивной чертой является неполное срастание барабанной пластинки – для миоцена очень примитивная черта. В сравнении с гиббоном плиопитек имел практически той же длины ноги, но куда как более короткие и толстые руки. Если у гиббона руки заметно длиннее ног, то у плиопитека – наоборот; пропорциями он больше напоминал ревунов или павианов. Кисти и стопы плиопитека были удлинены, а фаланги сужены и изогнуты, хотя и не в такой степени, как у гиббона. Тело, судя по всему, было тонкое и длинное. Строение крестца наводит на мысли о сохранении хвоста.
Крузелины (Crouzeliinae): Plesiopliopithecus, Crouzelia, Barberapithecus, Anapithecus, Egarapithecus и Laccopithecus – более поздняя и продвинутая группа плиопитековых. Они известны почти исключительно из Европы (14,5–9 млн лет назад), только Laccopithecus robustus жил в Южном Китае (6–8 млн лет назад). Крузелия Crouzelia и лаккопитек Laccopithecus – очередные предполагавшиеся предки гиббонов.

Рис. 34. Череп Pliopithecus vindobonensis.
Больше останков сохранилось от анапитека Anapithecus hernyaki, особенно в венгерском местонахождении Рудабанья (9,5 млн лет назад). Это обезьяна весила 15 кг. Замечательно, что при малых различиях в размерах тела самцы и самки существенно отличались размерами клыков. Это говорит о своеобразной социальной структуре анапитеков. В черепе анапитеков вполне объемно повторяется план строения черепа более древнего плиопитека: крупные размеры, округлая мозговая коробка, сравнительно вертикальный лоб, чуть развернутые вбок глазницы с утолщенным краем, широкое межглазничное пространство, короткая морда с низкой скуловой областью. Судя по зубам, анапитеки, как и прочие плиопитековые, были фруктоядными (DeMiguel et al., 2013), хотя раньше предполагалась их листоядность. Есть и специфика: верхнечелюстные пазухи анапитеков уменьшены, хотя и не исчезли, как у мартышкообразных. Кости остального скелета тоже похожи на плиопитечий вариант, хотя фаланги изогнуты сильнее.
Кстати, о бедрах…
Сложность изучения фрагментарных останков нагляднейше видна в истории с бедром из Эппельсгейма (Köhler et al., 2002). В Германии еще в 1820 г. была найдена бедренная кость довольно большого размера. Первоначально она была описана как принадлежащая дриопитеку, хотя получила сразу три названия – Paidopithex rhenanus, Pliohylobates eppelsheimensis и Pliosymphalangus. После было показано, что она заметно отличается от варианта, типичного для человекообразных обезьян, зато весьма напоминает бедренную кость Pliopithecus vindobonensis. Однако европейские плиопитеки весили около 10 кг, а эппельсгеймский примат – 22. По размерам к нему приближается Pliopithecus zhanxiangi, но он найден в Китае. В 1935 г. в том же местонахождении был найден клык самца примата, описанный как Semnopithecus eppelsheimensis, который впоследствии определялся как Pliopithecus, Rhenopithecus или Dryopithecus. В итоге плиопитековая его природа была признана наиболее вероятной, но размеры зуба говорят о том, что его обладатель был вдвое меньше, чем владелец бедра, так что отнесение их к одному виду опять же совершенно необязательно. Далее, в рядом расположенном местонахождении Сальмендинген нашлись зубы, определенные как Anapithecus hernyaki – то есть опять же, плиопитековый примат. А анапитек весил 15 кг – уже ближе к размерам эппельсгеймской обезьяны. Однако в 1990-х гг. в плиопитековой природе этих зубов появились основательные сомнения, и исследователи стали склоняться к дриопитековой версии. Наконец, очередное подробное морфологическое исследование показало, что бедро наверняка не принадлежало дриопитеку, им мог обладать либо неизвестный доселе очень крупный плиопитековый примат, либо вообще совсем своеобразная узконосая обезьяна из “третьей группы” (первые две – плиопитековые и человекообразные).
Плиопитековые являют собой своеобразный аналог широконосых обезьян в Старом Свете, евразийский эксперимент: с наибольшей вероятностью они возникли в Азии, но дали всплеск разнообразия в Европе, где до этого крупных растительноядных приматов не было. Олигоценовые холода выкосили европейских жителей лесов, к середине этой эпохи тут исчезли и адаписовые, и омомисовые. Как и в случае с Южной Америкой, свободные экологические ниши просто-таки томились – когда же их займут новые жильцы, и те не заставили себя ждать. Как только в миоцене новые теплые леса заколосились от Испании до Таиланда, в рекордные сроки кустообразная эволюция заполнила европейские пущи крикливыми пожирателями плодов и листьев. Именно по этой причине быстрого видообразования из малого числа предков так трудно классифицировать плиопитековых – тут параллель с широконосыми тоже совершенно очевидна.
Правда, в это же время из Африки и с Ближнего Востока подтянулись человекообразные и мартышкообразные, составившие плиопитековым мощную конкуренцию. В конце же миоцена и вовсе стало грустно: леса сохли и заменялись саваннами. В Африке это привело к появлению австралопитеков, а в Европе – к глобальному вымиранию лесных приматов. Как-то не вставалось им на две ноги.
Последние крузелины – они же последние плиопитековые и самые специализированные среди прочих – исчезли 9 млн лет назад в Европе (Egarapithecus narcisoi) и 6–8 – в Китае (Laccopithecus robustus).
Минутка фантазии
Давным-давно никто не считает плиопитековых предками человека. Но это не отменяет того, что все задатки для появления разумности у этих обезьян были. Они обладали относительно большим мозгом и маленькими челюстями, не слишком сильным половым диморфизмом, не имели резких специализаций в строении конечностей. В каком-то смысле они находились даже в выигрышных условиях по сравнению с предками австралопитеков Африки, ведь условия Европы были более суровыми, а стало быть – располагающими к прогрессивной эволюции. В северных широтах раньше стали исчезать тропические леса, появляться саванны и степи. Тут бы плиопитекам и слезть с деревьев, заселить просторы и устроить марш на восток, юг и куда еще захочется… Что остановило их эволюцию – не вполне очевидно. С одной стороны, вымирание плиопитековых совпадает с концом миоцена и резким похолоданием, с другой – южнокитайские виды, жившие далеко на юге, потенциально могли развиваться дальше. Надо думать, основную роль сыграла конкуренция с мартышкообразными и человекообразными: “истинные узконосые” оказались быстрее, бодрее и нахальнее. Показательно, что у анапитеков постоянные зубы прорезывались заметно раньше, чем у мартышкообразных сопоставимого размера (Nargolwalla et al., 2005). Предполагалось, что раннее взросление было связано с усиленным прессом со стороны хищников или листоядностью, но новейшие исследования говорят скорее о фруктоядности анапитеков. Возможно, ускоренное развитие было способом конкуренции с “истинными узконосыми”. Как бы то ни было, укороченное детство означает меньшие возможности для обучения и в итоге интеллектуальное отставание от конкурентов.
“Истинные узконосые”
“Истинные узконосые” Eucatarrhini захватили Старый Свет. Они отличаются от прочих обезьян повышенной интеллектуальностью, исключительно дневным образом жизни (кроме сторожей и ночных диспетчеров), почти исключительной растительноядностью (хотя все не прочь при случае поесть мяса, но специализаций к этому никаких нет) и специфическими чертами типа закрытости задней стенки глазницы, отсутствия слуховых капсул и полного прирастания барабанной пластинки.
Среди узконосых выделяют две главные группы: мартышкообразных и человекообразных. Судя по всему, они разделились очень рано и достаточно быстро. Долгое время момент их расхождения был покрыт мраком неизвестности. Существование общего предка мартышкообразных и человекообразных давно предсказывалось палеоприматологами. По всем выкладкам он должен был жить в Африке или на Ближнем Востоке в олигоцене, между 30 и 25 млн лет назад: около древнейшей даты мы знаем проплиопитековых, а около молодой – уже дифференцированых мартышкообразных и человекообразных. Но предок оставался неуловим.

Рис. 35. Череп Saadanius hijazensis.
Наконец, в 2010 году и это звено стало достающим: на западе Саудовской Аравии был найден отличнейший череп (Zalmout et al., 2010). Обезьяна была названа сааданиусом Saadanius hijazensis. Его датировка – 28–29 млн лет назад, то есть идеально вписывается в требуемый интервал. Столь же красиво-промежуточна и его морфология – настолько предковая, что самого сааданиуса невозможно отнести ни к мартышкообразным, ни к человекообразным. Великому Предку – великие почести: специально для него было выделено особое надсемейство сааданиоид (Saadanioidea) с единственным семейством сааданиид (Saadaniidae). Правда, при жизни его величие не было должным образом оценено: на черепе есть следы клыков какого-то хищника. Зато теперь мы можем воздать ему должное. Сааданиус был довольно крупным зверем – 15–20 кг, хотя самки наверняка были меньше. Судя по зубам, он был фруктоядным. К сожалению, пока не найдены кости его рук и ног, так что мы можем лишь догадываться, как он передвигался, но, думается, в этом сааданиус вряд ли сильно отличался от проплиопитековых и макак.
Уголок занудства
Сааданиус обладал своеобразным набором признаков: у него была сильно вытянутая морда, большие клыки, височные линии, сходящиеся в сагиттальный гребень, не было лобных пазух (они отсутствуют у проплиопитековых, плиопитековых и мартышкообразных, зато типичны для большинства человекообразных), но имелись верхнечелюстные синусы (их также нет у мартышкообразных, но они есть у проплиопитековых, плиопитековых и человекообразных), зато барабанная пластинка полностью срасталась с другими частями височной кости и целиком окружала слуховой проход (в отличие от проплиопитековых и “ранних узконосых”, но как у “истинных узконосых”), моляры были большими и широкими. Это самая генерализованная узконосая обезьяна, какую только можно представить.
Древнейшая мартышкообразная обезьяна и древнейшая человекообразная обезьяна, или Oldest monkey, oldest ape
В качестве древнейших мартышкообразных и человекообразных назывались разные животные. Лучшим кандидатом на роль древнейшей мартышки до недавнего времени считался викториапитек Victoriapithecus macinnesi, скакавший 15–20 млн лет назад по территории нынешних Кении и Уганды. Прохилобатес Prohylobates tandyi из Египта и норопитек Noropithecus bulukensis из Кении жили около 17 млн лет назад. Древнейшим же человекообразным признавался камойяпитек Kamoyapithecus hamiltoni, обитавший 23,9–27,8 млн лет назад в Кении, хотя он настолько схож с сааданиусом в своих примитивных чертах, что далеко не все исследователи были готовы признать его человекообразным приматом. Более надежные кандидаты – лимнопитек Limnopithecus legetet (17–22 млн лет назад, Кения и Уганда), угандапитек Ugandapithecus meswae (19–21,5 млн лет назад, Кения) и моротопитек Morotopithecus bishopi (20,6 млн лет назад или древнее, Уганда), хотя и их положение на древе приматов неоднозначно. Рангвапитек Rangwapithecus gordoni (19 млн лет назад, Кения) еще надежнее, но моложе. Имеющийся временной разрыв между сааданиусом и его потомками по эволюционным меркам не столь уж большой, но досадный. Естественно, дотошные палеоприматологи не могли смириться с таким положением дел. Благо недра Африки еще далеко не иссякли на свидетельства прошлого.В 2013 году американо-австралийско-танзанийская группа исследователей опубликовала описание двух древнейших обезьян (Stevens et al., 2013). Их останки, как водится, не слишком эффектны внешне (от одной сохранился обломок челюсти с одним моляром, от другой – половина нижней челюсти с четырьмя зубами), зато для специалистов несут ценнейшую информацию. Останки были найдены в Юго-Западной Танзании, около города Мбейя, в долине Руква, в местонахождении Нсунгве. Геологический слой, содержавший челюсти, образовался 25,2 млн лет назад.Левый нижний моляр был описан как нсунгвепитек Nsungwepithecus gunnelli. Его особенности больше всего напоминают характерные для мартышкообразных. Но он древнее самых древних мартышек на 5 млн лет!Четыре зуба из нижней челюсти были названы руквапитеком Rukwapithecus fleaglei. Детали строения выдают принадлежность их обладателя к примитивнейшим представителям человекообразных обезьян. Похожие существа иногда объединяются в подсемейство ньянзапитецин (Nyanzapithecinae) семейства проконсулид (Proconsulidae). Проконсулиды настолько примитивны, что не все ученые считают их настоящими человекообразными, хотя ясно, что они в любом случае предки “настоящих” человекообразных. И вот теперь мы имеем первейшего из первых или даже “нулевых”.Что значат 3 млн лет, отделяющие сааданиуса от нсунгвепитека и руквапитека? Много это или мало? Для сравнения – 3 млн лет назад наши предки были австралопитеками и еще не научились делать каменные орудия. Ясно, что за 3 млн лет могли смениться несколько видов, появиться даже несколько линий. Как часто бывает, в этих неведомых пока трех миллионах таятся очередные “недостающие звенья” между достающими “еще не вполне” и “древнейшими уже”. Великая заслуга и удача палеоприматологов – найти и описать истоки основных групп современных обезьян. Конечно, всегда хочется большего. Когда-то величайшим достижением считалось обнаружение любого древнего примата, теперь же того и гляди раздадутся недовольные голоса: “А почему вам нечего сказать об этих трех миллионах лет? Что это вообще за жалкие зубы?” Не будем слушать критиканов! Порадуемся лучше открытию очередных достающих звеньев. Работы в Африке продолжаются, хочется верить, что скоро мы увидим останки такого существа, про которое трудно будет сказать – ближе он к общему предку сааданиусу, мартышкообразному нсунгвепитеку или человекообразному руквапитеку.Благодаря работе палеоприматологов теперь мы знаем, когда и где разошлись пути мартышек и наш, когда и где были заложены очередные человеческие черты, когда и где был сделан еще один шаг к разуму.
Идеальные обезьяны: мартышкообразные
Итак, линии мартышкообразных и человекообразных разошлись. Их можно отличать по целому ряду признаков. Например, у мартышкообразных есть защечные мешки, седалищные мозоли и хвост, но нет гайморовых пазух в верхней челюсти, у человекообразных гораздо крупнее головной мозг и специализированнее конечности, отличия имеются и в строении зубов. Мартышкообразные в среднем меньше по размеру. Все эти и другие особенности позволяют распознавать ископаемых представителей даже по весьма фрагментарным останкам. К сожалению, они обычно и бывают фрагментарными.
Мартышкообразные (Cercopithecoidea), с одной стороны, многочисленны и внешне очень разнообразны, но с другой – удивительно монолитны филогенетически. Между крайними формами есть почти непрерывные морфологические переходы.
Древнейшие мартышковые появились не менее 25 млн лет назад. Они выделяются в особое семейство викториапитецид Victoriapithecidae: Nsungwepithecus, Victoriapithecus, Noropithecus, Prohylobates, Zaltanpithecus и, возможно, еще парочка родов. Только древнейший Nsungwepithecus жил в олигоцене, остальные – нижне– и среднемиоценовые, дожившие до 12 млн лет назад. Это исключительно африканские животные, их останки найдены в Танзании, Кении, Уганде, Египте и Ливии, так что возникновение мартышкообразных на Черном континенте практически неоспоримо.
Уголок занудства
От викториапитеков Victoriapithecus macinnesi сохранились целый череп, многие сотни зубов и остатки скелетов, так что их строение прекрасно изучено. Очевидно, эта обезьяна была обычнейшим видом. План строения черепа викториапитека откровенно мартышковый: выступающая морда, развернутые вперед глазницы, довольно сильное надбровье, узкое межглазничное пространство, длинные и узкие носовые кости, отсутствие лобных и верхнечелюстных синусов. Викториапитеки весили 3–4 кг, крупные самцы могли достигать аж 7–10,5 кг. Конечности были приспособлены к быстрому четвероногому бегу как по ветвям, так и по земле. Показательно, что фаланги викториапитека были короче, чем у мелких древесных мартышек, и даже короче, чем у полуназемных мандрилов, но такие же, как у наземных павианов и мартышек-гусаров. Викториапитек, судя по строению седалищной кости, имел седалищные мозоли и подвижный хвост (McCrossin et Benefit, 1992). Иначе говоря, седалищные мозоли были уже у первых мартышкообразных, еще до их разделения на современные группы.
В целом викториапитек удивительно похож на самых обычных современных мартышкообразных типа мартышек или макак. Получается, этот план строения тела оказался настолько удачным, что не возникало никакой нужды его менять больше десяти следующих миллионов лет. Это универсальное строение, идеально приспособленное сразу к разным типам местообитаний. Небольшими модификациями из него можно получить массу других вариантов – приспособлений к каким-то конкретным условиям.
Изобилие молочных зубов викториапитеков на острове Мабоко в Кении позволяет изучить не только особенности взросления этих обезьян, но и социальную структуру (Benefit, 1994). Клыки самцов встречаются намного чаще, чем клыки самок. Такое распределение говорит об организации общества наподобие макак: молодые самцы покидают группу, в которой родились, по полной программе испытывают невзгоды и опасности одинокой жизни и гораздо чаще гибнут. В группах же оказывается много взрослых самок и совсем мало самцов. Опять же это – удивительно стабильный в течение миллионов лет вариант.

Рис. 36. Череп Victoriapithecus.
Показательно, что едва ли не самая примитивная часть викториапитека – мозги (Gonzales et al., 2015). Особенно впечатляют размеры его обонятельных луковиц, контрастирующие с маленьким общим объемом: по этим показателям викториапитек больше похож на лемуров, египтопитеков и парапитеков. Объем мозга был 35,6 см³ – вдвое больше, чем у самок, и чуть больше, нежели у самых крупных самцов египтопитеков (по старым оценкам – 54 см³ – выходило, что викториапитек почти догонял мартышек среднего размера). На примере викториапитека можно воочию убедиться, что, при всей важности мозга для приматов, он на самом деле зачастую эволюционировал по остаточному принципу – после конечностей и зубов. Сколь часто звучит, что повышенный интеллект для приматов – самое святое! Сколь часто палеонтологическая реальность опровергает пафосную теорию…
Мартышковые (Cercopithecidae) подразделяются на собственно мартышковых Cercopithecinae (мартышки, макаки, мангабеи, мандрилы, павианы и гелады) и тонкотелых обезьян Colobinae (как это ни странно, их синоним – толстотелые; колобусы, носачи и лангуры). Они известны как минимум с позднего миоцена, примерно с 10 млн лет назад, причем уже во множестве видов; менее достоверные останки известны и из среднемиоценовых отложений, когда мартышковые, видимо, и появились. Все древнейшие находки: Colobus, Libypithecus, Microcolobus – сделаны в Африке.
Тонкотелые начали захватывать мир раньше: в верхнем миоцене представители рода мезопитеков Mesopithecus заселили Южную Европу, дошли до Ирана, Афганистана и Индии, мьянмарколобусы Myanmarcolobus yawensis уже жили в Бирме, а лангуры Presbytis – в Индии. В начале плиоцена в европейских лесах – от Франции до Болгарии, Молдавии и Украины – жевали листья родственные долихопитеки Dolichopithecus. В середине плиоцена тонкотелые освоили Азию, причем к этому моменту они оккупировали не только весь нынешний ареал, но распространились и гораздо севернее – мезопитеки до Германии, а парапресбитисы Parapresbytis eohanuman аж до Северной Монголии и Забайкалья. В конце плиоцена долихопитеки Dolichopithecus (Kanagawapithecus) leptopostorbitalis добрались до Японии, в Бирме и Индии появляется современный род хануманов Semnopithecus, а в Китае – носатые обезьяны Rhinopithecus. Своя эволюция продолжалась и в Африке: на протяжении плиоцена и плейстоцена тут обитали Cercopithecoides, Kuseracolobus, Rhinocolobus, Paracolobus и Colobus.
Собственно мартышковые вышли из Африки позднее тонкотелых: в начале плиоцена они прибыли в Европу, а в конце плиоцена жили уже по всей Европе и Азии. Без сомнения, всех превзошли макаки Macaca, которые появились еще в верхнем миоцене Северной Африки и огромное количество видов коих с начала плиоцена разбрелось от Испании, Франции и Германии до Китая, Японии и Индонезии. В их числе по всему югу Европы – в Италии, Испании, Германии, Франции, Венгрии – был распространен бесхвостый макак Macaca sylvanus, ныне обитающий на севере Африки и на Гибралтарской скале. В Европе жили и другие, ныне вымершие виды макак. В прошлом они населяли также Кавказ и даже Монголию (Macaca anderssoni). В нижнем плейстоцене в Китае некоторые виды достигли приличных размеров, неспроста один из них назван мегамакакой Megamacaca lantianensis. В конце плиоцена особенно удачным оказался близкий к макакам род парадолихопитеков Paradolichopithecus: его представители известны от Франции, Румынии и Греции (P. arvernensis) до Таджикистана (P. suschkini) и Китая (P. gansuensis). Тогда же по Индии и Китаю бегали проциноцефалы Procynocephalus. Удивительным образом, ни макаки, ни парадолихопитеки не проникли в тропические части Африки. Судя по всему, Сахара была лучшим барьером, чем даже морские проливы.
В Африке жизнь кипела своим чередом, плиоценовые саванны просто кишели павианами Parapapio, Pliopapio, Gorgopithecus, Dinopithecus, Procercocebus и первыми представителями уже современных родов павианов Papio и мангабеев Cercocebus. Целый ряд видов ископаемых гелад теропитеков Theropithecus на протяжении плиоцена и плейстоцена заселял всю Африку, в конце плиоцена попал в Индию, а в плейстоцене каким-то образом просочился в Испанию (что имеет прямое отношение к появлению тут человека – ведь в теории сюда можно попасть через Гибралтар или же добраться вдоль бережка в обход Средиземного моря). Многие древние павианы были гораздо крупнее современных. Так, гелада Theropithecus oswaldi могла достигать веса в 100 кг, тогда как даже большой самец крупного мандрила весит всего 35 кг. Такие павианы, как Dinopithecus и Gorgopithecus, вполне достойны названия саблезубых обезьян, ибо размеры их клыков были не меньше, чем у пресловутых саблезубых тигров.
Минутка фантазии
Интересно, что в эволюции павианов прослеживается универсальная тенденция к увеличению головного мозга. Хотя темпы такого роста не слишком высоки, он вполне достоверен и происходил тогда же, когда в Африке жили австралопитеки, причем у последних мозг увеличивался едва ли не меньшими темпами. Таким образом, павианы вполне могли бы обогнать двуногих конкурентов и развиться в собственное парачеловечество, если бы… Вероятно, причиной остановки прогресса послужила чрезмерная “бюрократизация” в обществе павианов, их повышенная агрессивность, жесткая иерархичность и приверженность строгому распорядку и субординации.
В конце плиоцена в Африке появляются современные мартышки Cercopithecus, а в середине плейстоцена современные виды зеленых мартышек Chlorocebus aethiops и мартышек-гусаров Erythrocebus patas.
Плейстоценовые оледенения выморозили обезьян в Европе и значительной части Азии, а опустынивание Средней Азии и Ирана истребило местных приматов; африканская и азиатская части ранее единого ареала оказались разделены. В итоге сейчас мартышковые обитают только в Африке, Южной, Восточной и Юго-Восточной Азии. От европейских обезьян ничего не осталось, популяция макак на Гибралтарской скале на самом юге Пиренейского полуострова, скорее всего, не реликтовая, а заселена сюда в исторические времена людьми. В Западной Азии чудом сохранились лишь павианы-гамадрилы на юго-западе Аравийского полуострова; предполагалось, что они могли быть завезены человеком сравнительно недавно, но анализ мтДНК показал, что заселение свершилось более 12 тыс. лет назад, а стало быть, скорее всего, без помощи людей (Kopp et al., 2014).
Несмотря на плейстоценовые потери, мартышковые являются одной из самых экологически пластичных и эволюционно преуспевающих групп приматов.
Мелкие виды мартышковых – мартышки и мангабеи – обычно сугубо древесные формы, средние – макаки – одинаково ловко бегают и по земле, и по ветвям, а крупные – мандрилы, павианы и гелады – наземные. Мартышковые заняли весьма широкий спектр экологических ниш, но все являются дневными, стадными и растительноядными.
Африканские мартышки: обыкновенные Cercopithecus, черно-зеленые Allenopithecus, зеленые Chlorocebus, гусары Erythrocebus и карликовые Miopithecus – возникли совсем недавно – около 2,5 млн лет назад и позже, фактически они современники нашего рода Homo. Однако, в отличие от гоминид, число видов которых скорее уменьшалось со временем, пока не свелось к одному, мартышки бурно наращивали свое разнообразие. Конкретные виды отличаются зачастую экологическими тонкостями: например, мартышка диана Cercopithecus diana держится только в первичном лесу и очень не любит заходить во вторичный, похожая мартышка салонго Cercopithecus dryas процветает во вторичном и даже предпочитает его первичному, тогда как большая белоносая мартышка Cercopithecus nictitans любит держаться в переходной зоне от дождевого леса к саванне. Зеленые мартышки, или верветки, Chlorocebus ведут уже полудревесный-полуназемный образ жизни в довольно засушливых районах Африки. Мартышки гусары Erythrocebus patas приспособились к жизни в открытой саванне, где они бегают подобно мелким антилопам. Большинство мартышек питаются фруктами и в меньшей степени листьями, но голубая мартышка Cercopithecus mitis в большом количестве ест также медленных беспозвоночных.
Ареалы многих видов совпадают почти полностью, ведь зона лесов в Африке ограничена, так что среди мартышек хорошо поработал половой отбор. Большинство видов отличаются почти исключительно формой и цветом пятен и пучков волос на лице, это ясно даже по названиям: желтоносая, совинолицая, голуболицая, белощекая, чернощекая белоносая, коронованная, чубатая, усатая, бородатая, белогубая, красноухая мартышка с пятном на носу, большая белоносая мартышка, малая мартышка с белым пятном на носу. Те же и другие соревнуются в окраске живота или хвоста: белогорлая, краснобрюхая, золотобрюхая, золотая, краснохвостая, рыжехвостая. Калейдоскоп настолько пестр, популяционная и индивидуальная изменчивость столь велики, а переходы такие постепенные, что среди систематиков никогда не утихнут споры даже о числе родов и видов, не говоря уж о подвидах.
Макаки Macaca – самые холодоустойчивые обезьяны (кроме человека); особенно выделяются в этом отношении тибетские M. thibetana и японскиея макаки M. fuscata, каждую зиму сталкивающиеся с морозом и снегом. Эксперименты показывают, что макаки вполне могут жить на круглогодичном вольном выпасе в климате современной Германии. В военном Харькове три сбежавших из разбомбленного зоопарка макаки прожили без малого два года – перезимовали две зимы, – партизаня в здании Госпрома в самом центре города.
Достаточно крупные размеры и всеядность делают макак универсальными обезьянами и позволяют им выживать в самых разных местах – от скалистых утесов до густых лесов, от морских побережий до поселений людей в качестве воришек-нахлебников-мусорщиков. Жизнь шумными компаниями, подвижность и сообразительность – отличная защита от большинства хищников.
Самые банальные макаки – азиатские резусы M. mulatta и крабоеды M. fascicularis, а также североафриканские маготы M. sylvanus.
Резко выделяются устрашающей внешностью некоторые представители группы целебесских макак, или черные павианы, – M. hecki, M. maura, M. nigra, M. nigrescens. У этих угольно-черных обезьян поистине демонический облик: странная вытянутая морда, хохол на макушке и огромные зубы.
Другие виды имеют свой неповторимый облик: индийский львинохвостый макак M. silenus обладает шикарной белой гривой и пушистой кисточкой на длинном хвосте, тогда как у близкородственного свинохвостого M. nemestrina и грива гораздо скромнее, и хвостик крючком.
Африканские мангабеи: обыкновенные Cercocebus, бородатые Lophocebus и горные Rungwecebus – филогенетически наиболее близки павианам, но ведут древесный образ жизни; фактически они занимают ту же экологическую нишу, что макаки в Азии, тут мы видим пример своего рода микроконвергенции на уровне подтриб в пределах близкородственной в целом группы. У мангабеев хвост очень длинный и имеет очень слабую хватательную способность – уникальный случай среди узконосых приматов, хотя, конечно, они бесконечно далеки от возможностей цепкохвостых широконосых. Внешность некоторых видов примечательна панковскими хохлами-ирокезами и залихватскими гусарскими усами или бакенбардами, а то и бровями а-ля Мефистофель. Танзанийский Rungwecebus kipunji уникален тем, что умудрился быть неописанным до 2005 года, проживая в более-менее исследованной зоологами местности.
Рис. 37. Павианы: гвинейский P. papio (а), анубис P. anubis (б), гамадрил P. hamadryas (в), бабуин P. cynocephalus (г), чакма P. ursinus (д).
Мандрилы – Mandrillus и павианы Papio – крупные животные с клыками как у леопарда, очень сильные и агрессивные. Среди них особенно цветасты западноафриканские мандрилы с их ярко раскрашенной мордой и задом. Рекордные самцы мандрила Mandrillus sphinx могут достигать метра в длину и 50 кг веса! Это предел для современных мартышковых обезьян. Впрочем, обычно они в полтора раза меньше, а самки и вовсе весят в среднем 15 кг. Другой рекорд мандрилов – способность собираться в огромные стада, самое внушительное из которых состояло из примерно 1300 особей! Очевидно, что набеги таких орд на плантации местных жителей заканчиваются праздником для мандрилов и катастрофой для поселян.
Кстати, о леопардах и уроках…
В некоторых африканских странах возникает хитрая связь между браконьерством и успеваемостью школьников: когда охотники истребляют хищников, прежде всего леопардов, то плодятся мандрилы и павианы, они разоряют поля, на охрану и защиту коих посылаются дети, которые прогуливают уроки и в итоге плохо учатся. Мораль – берегите природу!
И мандрилы, и павианы питаются большей частью растительной пищей, но регулярно ловят мелких животных типа дукеров или детенышей антилоп. Показательно, что это никоим очевидным образом не отражается на их зубочелюстном аппарате; разрывают же они добычу руками, хотя бы и не голыми, а мохнатыми.
Мандрилы ведут более-менее древесный образ жизни, тогда как павианы уже почти полностью наземны и забираются на деревья только на ночевку или спасаясь от хищников.
Наземные павианы представляют вариант жестко построенного иерархического общества, где каждая особь кому-либо подчинена и кого-либо подчиняет. Иерархия основана на физической силе и жизненном опыте. Все дозволено лишь самым маленьким детенышам и вожакам стада. При переселениях стадо павианов представляет собой образец построения группы на марше. В центре стада движутся самки с детенышами, возглавляет шествие вожак, а по сторонам охраняют самцы. Жесткая иерархическая система держится на альфа-самцах. Интересно, что почти все нововведения и инициативы рождаются в “низах общества” – среди павианов самого низшего ранга, чаще среди самок, которые не так заняты поддержанием своего социального статуса, но которым достаточно плохо живется, чтобы хотелось стремиться к лучшему. Тот же эффект ярко был продемонстрирован в наблюдениях над группой японских макак, которых исследователи подкармливали бататами. Когда одна самка низшего ранга случайно уронила батат в воду, она обнаружила, что чистым его гораздо приятнее есть – песок на зубах не скрипит. Она стала регулярно мыть клубни, от нее научился ее детеныш, а после нововведение распространилось “от низов к верхам”. Последними научились мыть клубни альфа-особи; им и так хорошо, их позиция – лучше, когда ничего не меняется, пусть все навеки застынет в стабильности.
Раньше павианов часто предлагали в качестве модели первобытного человеческого стада. Однако закрепленность и регламентированность взаимоотношений среди членов такого общества не дает возможности развития, закрепления новаций и распространения их по всем социальным слоям. Поэтому маловероятно, что отношения среди древнейших людей были похожи на отношения среди павианов.

