Кстати, о светофорах и доброте…
Иерархия бывает разной – совсем жесткой или же с умом, зависимой от ситуации. Бродячие собаки имеют вожака, но замечено, что при переходе через улицу руководство переходит к другой псине – особо интеллектуальной, умеющей наиболее адекватно отслеживать светофоры и поведение людей. У людей эта способность переключать доминирование в определенных ситуациях развита еще лучше. Классическая иллюстрация – фраза водителя Копытина в сцене погони в фильме “Место встречи изменить нельзя”: “В кабинете у себя командуй, Глеб Егорыч. А тут я…” Люди делают это легко и непринужденно и обычно даже не замечают этого. Например, во время лекции доминант – лектор, в его аудитории могут сидеть самые разные люди – врачи, милиционеры, слесари или чиновники, но все они молча слушают и послушно записывают сказанное. На дороге доминант – инспектор ГАИ, тут он получает власть над врачами, лекторами и слесарями, иногда (а по-хорошему всегда) даже над чиновниками. В школе доминант – учитель, в больнице – врач. Когда там протекает труба, безоговорочным доминантом становится сантехник, временно он получает власть отменить урок или перенести операцию. Более того, когда такое переключение нарушается, общество перестает нормально функционировать. Если в больнице или школе доминантом становится чиновник, охранник или уборщица, то начинается бардак. Сосредоточение власти в одних руках позволяет решать определенные задачи в некоторых ситуациях, но не должно быть излишним. Благодаря разделению ролей и смене доминирования человечество совершило все свои великие достижения.Удивительные изменения социальной структуры произошли в одной из групп павианов, живущей в густо населенной людьми местности. Обезьяны не могут удержаться и грабят плантации; владельцам полей это не нравится, и они берутся за оружие. Конечно, первыми под раздачу попадают самые агрессивные и самые наглые самцы павианов. И вот в результате систематического уничтожения наиболее злобных доминантов резко изменилось соотношение полов – самок оказалось намного больше, чем самцов, обезьяны стали несравненно добрее, уровень агрессии заметно снизился. В этой группе появилось даже такое невиданное среди павианов явление, как груминг – чистка шерсти – самцов самцами. Так вот он – путь к всеобщему счастью, доброте и взаимопониманию!
Особый интерес представляет систематика павианов – проклятье любителей однозначных схем. В зависимости от подхода можно выделить от одного до пяти или даже семи современных видов. Дело в том, что крайние географически разнесенные друг от друга варианты достаточно отчетливо отличаются на глаз. Но в промежутках есть смешанные популяции, в которых идет гибридизация, так что границы между видами оказываются весьма туманными. Получаются цепочки не то видов, не то подвидов, ломающие школьные представления о систематике.

Рис. 38. Черепа проконсула (а) и афропитека (б); реконструкция проконсула.
Уголок занудства
Картина расселения и гибридизации павианов всегда привлекала исследователей (например: Newman et al., 2004).Гвинейские павианы P. papio живут на крайнем западе Западной Африки. Их ареал без малейшего перерыва продолжается в ареал анубиса. Они самые мелкие – вдвое меньше южноафриканских чакм. Гвинейские павианы имеют красновато-бурый цвет и маленькую гриву.Павиан анубис, или оливковый, P. anubis расселен широкой полосой по Сахелю от южных рубежей Сахары до тропических лесов Центральной Африки и от Западной Африки до Восточной, где его ареал соприкасается с ареалами гамадрила (с которым анубис гибридизуется в Эфиопии) и желтым павианом. Анубис имеет массивное сложение, серовато-буровато-зеленоватый цвет и густую, но короткую гриву.Гамадрил, плащеносный или священный павиан, P. hamadryas населяет Аравию и Африканский Рог, похож на P. anubis, с коим регулярно скрещивается, и P. papio. Этот вид отличается также мощной конституцией, серым цветом и богатой гривой.Желтый павиан, или бабуин, P. cynocephalus освоил Африку южнее и восточнее от зоны лесов, на севере соприкасаясь с ареалом анубиса, а на юге – чакмы. Хотя его зона распространения не пересекается с ареалом гвинейского павиана, они прекрасно скрещиваются между собой в зоопарке. Бабуины желтовато-бурые, с вытянутым телом, длинными ногами и хвостом, хотя не столь удлинены и крупны, как чакмы; у бабуинов есть бакенбарды, но нет гривы. Из желтых павианов предлагается выделить в качестве самостоятельного вида бабуина Кинда P. kindae, отличающегося вдвое меньшими размерами и укороченной мордой.Медвежий павиан, или чакма, P. ursinus занял Южную Африку. Северные группы, пограничные с бабуинами, предлагается выделить в отдельный вид сероногого павиана P. griseipes. Чакма – самый крупный вид, с очень вытянутыми пропорциями, серого или темно-бурого окраса, совсем без бакенбард и гривы.
Гелады Theropithecus gelada – эфиопские павианы – внешне очень похожи на обыкновенных павианов, но в реальности отстоят от них больше, чем мангабеи, и выделяются в отдельную подтрибу. Современные гелады населяют исключительно горы Эфиопии, где бывает довольно холодно, почему имеют густую шерсть в виде огромной гривы, переходящей в богатую мантию на плечах и спине. Постоянное карабканье по крутым скалистым горам развило в геладах невероятную цепкость, а по деревьям они никогда не лазают за неимением деревьев. Кстати, образ их жизни навел некоторых исследователей первой половины XX века – например, А. П. Быстрова – на мысль о происхождении прямохождения в горной местности.
Очень много времени гелады проводят сидя на заду, подняв тело вертикально. Это приводит к неожиданной проблеме: ведь у павианов самки демонстрируют готовность к размножению набухающей половой кожей, расположенной вокруг половых органов. А у сидящих самок гелад ее просто не видно. Задача решилась развитием голого участка ярко-красной кожи в виде ошейника, переходящего в треугольник на груди. У самок она набухает кровью и сильно увеличивается во время готовности к размножению, так что у сидящей обезьяны ее видно издалека. У самцов такое же красное пятно имеет также сигнальную функцию, но подтекст меняется – им они сигнализируют о своей доминантности, ведь группы гелад представляют собой гаремы. Впрочем, отдельные гаремы могут собираться в стада вплоть до 670 особей.
По экологической необходимости гелады – почти исключительно травоядные существа, в зависимости от сезона собирающие вершки или корешки, но предпочитающие семена. По этой причине у них отлично развиты противопоставление большого пальца кисти и точечный захват; только человек превосходит их в этом искусстве.
Нельзя не заметить, что в плиоцене и плейстоцене гораздо более многочисленные и широко распространенные гелады были основными конкурентами австралопитеков и первых людей, причем обезьяны имели, казалось бы, очевидное преимущество в размерах и силе. Однако на большей части Африки гелады в итоге вымерли, сохранившись лишь в самых труднодоступных и малонаселенных районах. Очевидно, гоминиды одержали сокрушительную победу в напряженной войне, длившейся пару миллионов лет. Что стало залогом успеха наших предков? Хотелось бы верить, что способность быстро соображать и приспосабливаться к новым условиям, использовать больший спектр ресурсов и воспитывать детей. Но не исключено, победила слабость австралопитеков, тогда как мощь гелад сработала против них, но на наших предков: от 3,3 до 2 млн лет разнообразие и размеры гелад росли, они успешно вытесняли австралопитеков из экологической ниши бродячих собирателей семян, двуногим бедолагам было некуда податься от клыкастых и гривастых супротивников. С горя и безысходности они подались в падальщики и мелкие хищники – уж лучше соревноваться с десятикилограммовым шакалом, чем со стокилограммовой обезьяной. Но поев мяса и закалившись в боях с гиенами и львами, гоминиды набрались ума-разума, взяли в руки копья и вернулись к былым обидчикам уже не как бедные родственники, а с гастрономическим интересом. Тут-то и кончились светлые деньки гелад, да и сами гелады…
Тонкотелые отличаются от мартышковых в узком смысле исключительной древесностью, что привело к грацилизации тела и изменению формы кисти и стопы, а также узкой специализацией к питанию только листьями, что отразилось как на строении зубов (моляры с очень высокими бугорками), ротовой полости (защечные мешки развиты слабо или даже отсутствуют), так и пищеварительного тракта (желудок состоит из трех или даже четырех отделов, в одном из коих живут симбиотические бактерии, а в нижнем выделяется соляная кислота; кроме того, бактерии сидят в увеличенной слепой кишке и специальном отрезке толстой кишки – проксимальной ободочной). Малопитательность листьев и трудности их усвоения приводят к тому, что до трети веса обезьяны может приходиться на недопереваренные листья в ее желудке. Впрочем, как любые приматы, при случае большинство тонкотелых едят и фрукты, и насекомых, и других мелких животных.
Тонкотелые делятся на африканских колобусов и азиатских лангуров. Приятно, что морфологическая логика идеально совпадает с географической.
Африканские колобусы (они же гверецы или толстотелы) менее разнообразны, чем мартышки или азиатские тонкотелые, но не менее красивы. Они делятся на черно-белых Colobus (Colobus), черно-красных C. (Piliocolobus) и зеленых, или оливковых, C. (Procolobus); иногда всем им придается ранг самостоятельных родов. Особенно эффектны черно-белые гверецы, имеющие при общем черном фоне длинную белую шерсть в виде мантии на плечах и огромной кисти на хвосте; виды отличаются пропорциями черного и белого; впрочем, черная гвереца сатана C. satanas абсолютно угольная. Детеныши чисто белые. К сожалению, красота требует жертв: на гверец усиленно охотятся из-за шкур, идущих на статусные наряды и украшения. Древесность гверец почти абсолютная, поэтому их пальцы руки работают как единый крючок, а большой палец редуцируется до маленького бугорка.
Некоторые колобусы чрезвычайно специализированы к питанию листьями даже на фоне других тонкотелых; например, занзибарские красные колобусы Кирка Colobus (Piliocolobus) pennantii kirkii (или Piliocolobus kirkii) вообще неспособны усваивать сахар из спелых фруктов, зато имеют четырехкамерный желудок. Они любят есть уголь, видимо для нейтрализации токсинов, содержащихся в листьях.
Уголок занудства
Многокамерный желудок тонкотелых имеет в предельном варианте четыре камеры (Chivers, 1994; Kay et Davies, 1994).Первая – saccus gastricus – главная часть желудка, в которую впадает пищевод. Ее слизистая содержит желудочные железы, выделяющие слизь с ферментами, и лимфатические узлы.Вторая – pre-saccus gastricus – преджелудок, расположен сбоку параллельно главной части и частично сегментирован. Это дополнительный ферментативный отдел с различными типами слизистой. Он имеется у представителей Procolobus, Piliocolobus, Rhinopithecus, Pygathrix и Nasalis, но отсутствует у Colobus, Semnopithecus, Trachypithecus и Presbytis.Третья – tubus gastricus – трубка наподобие толстой кишки, подразделенная на вздутые сегменты-гаустры, содержащие симбиотических бактерий. В ее нижней части железы продуцируют соляную кислоту и разные ферменты.Четвертая – pars pylorica – пилорическая или привратниковая часть; ее железы выделяют только слизь.
Азиатские тонкотелые невероятно разнообразны. Юго-Восточная Азия и Индонезия – чрезвычайно географически расчлененная территория, на материке много гор с долинами, а острова – вообще лучшая эволюционная лаборатория. Варианты тонкотелых возникали, судя по всему, очень быстро, в течение последних нескольких миллионов лет, а на островах наверняка – последних тысячелетий. Далеко не все формы четко отличимы друг от друга. Поэтому до сих пор среди приматологов нет согласия относительно числа не только видов, но и родов тонкотелых.
Самые “усредненные” азиатские тонкотелые – лангуры хануман Semnopithecus, живущие на Индостане и Шри-Ланке. Лангуры удивительно экологически пластичны: они способны жить в засушливых открытых пространствах и тропических лесах, на побережье и в Гималаях на высоте в 4000 метров над уровнем моря. Это самые обычные животные в Индии, благо лангуры священны и их никто не трогает. Зная свою безнаказанность, эти стройные серые обезьяны наглеют до крайности – не один турист пострадал от их бесцеремонности. Именно лангуры, скорее всего самый обычный их вид S. entellus, послужили прототипом бандерлогов Р. Киплинга.
Намного более красочны зондские лангуры Presbytis: разных цветов, с ирокезами и бакенбардами, эти бойкие обезьяны очень оживляют тропические леса островов Индонезии. Каштановый лангур Presbytis rubicunda с Калимантана имеет приятный красноватый цвет, тогда как лангуры Хосе P. hosei и Томаса P. thomasi – серые, со стоячей на макушке шерстью, точно как у чертей с шишом на средневековых русских иконах (какое раздолье тут открывается для графоманских измышлений на тему индийско-русских связей в незапамятные времена!). Родственные хохлатые лангуры Trachypithecus тоже имеют высокий острый хохол на голове, отчего несколько напоминают хохлатых мангабеев, тем более что и телосложение у них в целом схоже. Видов и подвидов этих обезьян величайшее множество, их систематику впору выделять в особый олимпийский вид спорта. Особенно красив золотоголовый лангур T. poliocephalus. Очковые T. obscurus и серебристые лангуры T. cristatus сами серенькие, зато детеныши у них ярко-желтые.
Особняком стоит немейский тонкотел Pygathrix nemaeus из Индокитая. Это едва ли не самая разноцветная обезьяна из всех: белый, оранжевый, желтый, черный с бурыми подпалинами, серый цвета составляют неповторимый наряд этого примата, причем три подвида еще отличаются друг от друга.
Ринопитеки Rhinopithecus в отечественной литературе часто называются “носатыми обезьянами”, но это неверный – ошибочно укороченный – перевод с английского. В действительности они snub-nosed – коротконосые, так как у них действительно нос укорочен до полного отсутствия, снаружи видны только ноздри. Рокселланов золотой ринопитек Rh. roxellana из Тибета имеет незабываемый золотой цвет густой шерсти и голубую мордочку.
Прямая противоположность ринопитекам – калимантанские обезьяны носачи Nasalis larvatus. Они запоминаются с первого взгляда: у самцов огромный, свисающий вниз нос, придающий им вид пожилого серьезного клоуна на пенсии; у самок он остренький и курносый, делающий их легкомысленными кикиморками. Любопытно, что близкородственный одноцветный носач N. concolor с Ментавайских островов имеет вполне стандартный нос (зато он уникален среди тонкотелых очень коротким хвостом – всего около 15 см). Носачи, кроме своего носа, замечательны размерами: при среднем весе самцов 20,4 кг они являются самыми тяжелыми сугубо древесными приматами, кроме орангутанов.
Заря в преддверии заката: первые человекообразные
Наш путь через дебри эволюции приматов подошел к последнему кустистому ответвлению, от коего доныне сохранилась лишь одна зеленеющая ветка и две откровенно засыхающие. Прочитав про зелень, кое-кто наверняка подумал о себе любимом, но увы – вряд ли среди читателей сей книги есть гиббоны, а в виду имелись именно они. Засыхающие же сучья – это орангутаны и африканские гоминоиды: гориллы, шимпанзе и люди. В прошлом гоминоиды цвели куда как пышнее.
Человекообразные Hominoidea (они же Anthropoidea или Anthropomorpha) отличаются от мартышкообразных в среднем более крупными размерами, большим мозгом, отсутствием защечных мешков и наружного хвоста, а также деталями строения зубов.
Гоминоиды появились в позднем олигоцене и пережили свой расцвет в миоцене. Известно огромное множество ископаемых видов, из которых подавляющее большинство являются тупиковыми формами. Разобраться в них не так-то легко, ведь самый ходовой элемент тут – отдельные зубы или даже их обломки, а у родственных видов они могут быть весьма схожи. Тем не менее ясно, что самыми древними и архаичными являются проконсулиды Proconsulidae и дендропитециды Dendropithecidae. Тех и других из-за их примитивности иногда возводят до ранга собственных надсемейств Proconsuloidea и Dendropithecoidea – шаг, оправданный скорее филогенетически, чем морфологически. Проконсулиды и дендропитециды – почти полностью африканские приматы (единственное исключение – одна находка в Саудовской Аравии), так что возникновение человекообразных в Африке практически несомненно.
Центры происхождения и происхождение мыслей о центрах происхождения мысли
Долгие десятилетия в антропологической литературе бурно обсуждался вопрос о центрах происхождения человекообразных обезьян и человека, причем эти две проблемы обычно никак не разделялись. Вплоть до конца XX века в учебниках обязательным был раздел о “географической прародине человека”.С XIX века рассматривалась вероятность появления человекообразных обезьян и человека в Южной Европе, так как именно тут впервые были найдены останки дриопитеков. Эти воззрения в нетронутом виде дожили до современности в заповедных отечественных школьных учебниках, как испорченный граммофон, воспроизводящий заевшую пластинку времен зари науки.В первой половине XX века многие антропологи считали прямыми предками человека рамапитеков и сивапитеков, многочисленные останки коих сохранились в миоценовых отложениях Пакистана и Индии. Основным аргументом было строение их зубной системы, в частности сравнительно небольшие клыки. К тому же до первой трети XX века находки ископаемых человекообразных обезьян из Африки не были известны, да и долго после эти находки были слишком фрагментарными и немногочисленными. Из Азии же происходили и черепа питекантропов и синантропов – древнейших прямоходящих, известных до открытия австралопитеков (которые к тому же долгое время после первоописания рассматривались многими антропологами как тупиковая ветвь эволюции). В итоге так называемая “гипотеза азиатской прародины” считалась одной из ведущих до 1960-х годов. В настоящее время, с открытием практически непрерывной линии эволюции гоминид в Африке и неоткрытием такой линии в Азии, “азиатская концепция” вспоминается только как элемент истории антропологии.Позже находок в Азии были сделаны богатейшие сборы в Африке. Сейчас описана целая плеяда видов высших африканских приматов, живших с конца олигоцена до современности. Их морфология преемственна от более древних африканских “ранних узконосых” к австралопитекам и современным человекообразным обезьянам. Посему из трех возможных центров происхождения исходного предка человека Африка, бесспорно, побеждает.
Проконсулиды Proconsulidae включают самых древних из известных человекообразных. Внутреннее подразделение проконсулид остается предметом бурных споров, но кажется удобным разделить их на подсемейства проконсулин Proconsulinae, ньянзапитецин Nyanzapithecinae и афропитецин Afropithecinae. Самый примитивный из всех – камойяпитек Kamoyapithecus hamiltoni, живший в конце олигоцена, в интервале от 27,5±0,3 до 24,2±0,3 млн лет назад в Кении. Многими своими чертами камойяпитек схож с сааданиусом, так что его можно считать еще одним достающим звеном – между примитивнейшими узконосыми и человекообразными. Однако удобно включать его уже в проконсулин.
Прочие проконсулины Proconsulinae: Proconsul, Kalepithecus, Limnopithecus, Nacholapithecus – бегали по восточноафриканским лесам 20–15 млн лет назад. Они известны по огромному количеству остатков, в том числе почти целым черепам и скелетам. Больше всех разрекламированы собственно проконсулы, в частности Proconsul heseloni, так как именно к этому виду принадлежит известнейший череп и скелет самки KNM-RU 7290, чей кривоватый анфас чаще, впрочем, фигурирует в популярных книгах под обозначением P. africanus. Большинство проконсулов были размером меньше шимпанзе: P. heseloni – самки 9–11 и самцы 20 кг, P. nyanzae – 35–38 и 26–28 кг, но огромные P. africanus достигали аж 63–87 кг.
Хотя часто проконсулов воспроизводят похожими на шимпанзе, они отличались массой примитивных черт. Проконсулы имели очень слабое надбровье, сравнительно небольшие размеры морды и клыков, узкое носовое отверстие, мелкое небо; мозг у самки KNM-RU 7290 достигал 130,3 см³ (то есть как у крупных самцов сиамангов и вдвое с лишком меньше, нежели у самых захудалых шимпанзе), а его форма больше похожа на мозг широконосых обезьян. Строение зубов проконсулов замечательно генерализованное, то есть годящееся в предки кому угодно: на молярах эмаль сравнительно толстая, но не слишком, цингулюм – утолщение эмали в виде пояска у шейки зуба – бывает развит, а бывает и ослаблен. Кстати, к толщине эмали зубов и цингулюму учеными проявляется повышенный интерес, эти признаки используются в числе основных диагностирующих при систематике древних гоминоидов.
Тело и конечности первых человекообразных совсем не были похожи на шимпанзиные. Бегали все проконсулиды – и проконсулины в частности – примитивно: на четвереньках, с опорой на ладонь; при этом длина рук и ног была примерно одинаковой. Морфологических приспособлений к вертикальному лазанию и подвешиванию на ветвях они либо вовсе не имели, либо только начинали их развивать. Благодаря находкам нескольких скелетов мы знаем, что Proconsul heseloni (17–18,5 млн лет назад) был совсем-совсем четвероногий и горизонтальный, а более поздний (15–16 млн лет назад) нахолапитек Nacholapithecus kerioi из Кении уже начал приобретать способности к вертикальному лазанию и, соответственно, вертикальному положению тела и головы (в частности, у него были чуть длиннее руки) – это были первые робкие подвижки в сторону нашего нынешнего состояния. Оба вида уже были бесхвостыми, так что сия черта, многими воспринимающаяся как сугубо человеческая, как минимум всемеро старше нашего рода.
Уголок занудства
Лопатка Proconsul heseloni больше похожа на таковую колобусов и прочих не подвешивающихся на руках приматов. Грудная клетка сравнительная длинная и узкая. Длинные кости массивные. Шиловидный отросток локтевой очень большой, образующий сустав как с трехгранной, так и с гороховидной костью; кисть длинная и узкая, но ее фаланги сравнительно короткие и слабо изогнутые. В запястье сохраняется самостоятельная центральная кость; таковая имеется у широконосых и мартышкообразных обезьян, тогда как у молодых гиббонов и орангутанов она тоже свободна, но не двигается отдельно от ладьевидной, а у старых может срастаться с последней; у горилл, шимпанзе и человека центральная кость срастается с ладьевидной еще в эмбриональном состоянии.Подвздошная кость проконсула узкая, по форме и ориентации больше похожая на кость мартышкообразных; на седалищной нет седалищной бугристости, так что сидел и спал проконсул в позах, типичных для широконосых, а не узконосых обезьян. Стопа больше всего напоминает стопу древесных, но нечеловекообразных приматов. Проконсул имел особую кость прехаллюкс, соединявшуюся с основанием первой плюсневой; такая же встречается у некоторых широконосых, египтопитеков и гиббонов, но отсутствует у мартышкообразных и высших человекообразных (Lewis, 1972).
Добро пожаловать, экембо! или Новый образ проконсула?
Систематика первых обезьян – хитрая наука на грани искусства. В ход идут как объективные параметры вроде размеров зубов, так и показатели формы, определяемые на глаз. Только пересмотрев сотни зубов, можно научиться различать хитрые одонтологические признаки вроде развития дистолингвального цингулюма, степени изгиба дистального края коронки и формы буккального профиля. Проблема в том, что, во-первых, разные антропологи смотрят на разные зубы, а во-вторых, придают признакам неодинаковый смысл. И конечно, никуда не девается индивидуальная, половая и возрастная изменчивость.Посему не странно, что среди человекообразных палеоприматологи выделяют сильно разное число не только видов и родов, но даже семейств! Особенно лихорадит систематику древнейших представителей – проконсуловых. Хорошо жилось систематикам конца XIX – начала XX века. Сначала всех первобытных человекообразных до кучи валили в единый род Dryopithecus, потом начали выносить азиатских представителей в свои роды (каковых навыделяли больше десятка, из которых валидными оказались… примерно два), наконец, дошли руки и до африканских зверей. Их стали называть проконсулами – Proconsul.Когда критическая масса находок выросла и стали очевидны существенные отличия разных “проконсулов”, опять пошла закономерная волна дробительства: названия родов африканских антропоидов посыпались как из рога изобилия, а свежеиспеченные таксоны стали расползаться по своим подсемействам и семействам.Однако некое ядро оставалось неприкосновенным: несколько видов считались “образцовыми проконсулами”, на которых никто не покушался. Но можно ли устоять перед соблазном выделить еще один род? Никак нельзя! И потому группа палеоприматологов сделала это – описала род Ekembo (McNulty et al., 2015).В роде Proconsul остались виды P. africanus, P. major и P. meswae из кенийских местонахождений Тиндерет, а также P. major из Уганды (этот последний – весьма спорный, другими исследователями он описывался как Ugandapithecus major, Ugandapithecus gitongai и Morotopithecus bishopi; равным образом P. meswae часто рассматривается как Ugandapithecus meswae). Те же останки, что были найдены на берегу озера Виктория в кенийских местонахождениях Кисингири, выделены ныне в новый род Ekembo. В нем оказались виды E. heseloni и E. nyanzae.Уже говорилось, что к виду heseloni принадлежит известнейший череп KNM-RU 7290. Так что же – переписываем учебники? Ставим под картинками новые подписи?Но. Стоит ли спешить? Насколько такая система лучше прежней – покажут новые исследования. Хотя отличия Proconsul и Ekembo показаны достаточно надежно – новоявленный род заметно мельче и грацильнее, – остается неочевидным: достойны ли они родового статуса? Существование разных видов не было секретом и раньше, виды в новом исследовании не подвергаются сомнению. А надежного критерия рода покамест никто из систематиков не предложил. Так что экзотическое “экембо” должно еще подтвердить право на существование. Желательно – практикой: если парадигма двух родов позволит лучше классифицировать новые находки и позволит уточнить родословную первых человекообразных – добро пожаловать, экембо!
Примерно синхронен камойяпитеку руквапитек Rukwapithecus fleaglei, обитавший 25,2 млн лет назад в Танзании. Он близок ньянзапитецинам Nyanzapithecinae: Mabokopithecus, Nyanzapithecus, Rangwapithecus, Turkanapithecus, чьи зубы находят в кенийских и танзанийских отложениях с датировками от 19 до 13 млн лет назад. От прочих проконсулид ньянзапитецины отличаются широкими и короткими резцами, а также другими частностями строения зубов. Лучше прочих изучен турканапитек Turkanapithecus kalakolensis – от него сохранился почти целый череп и значительная часть скелета. Турканапитек был отдаленно похож на шимпанзе, но имел намного меньшие размеры – около 10 кг, гораздо более короткую морду со сравнительно короткими клыками, довольно слабое надбровье даже у самцов, очень широкое межглазничное пространство, сильно расширенные скуловые дуги при суженной мозговой коробке, мозг всего 84,3 см³ – как у макаки, заметно меньше, чем у гиббона, и почти в пять раз меньше, чем у шимпанзе. Ньянзапитецины были вообще не слишком крупными: ньянзапитеки Nyanzapithecus имели размер макаки – 8–11 кг, а рангвапитек Rangwapithecus gordoni весил примерно 15 кг. Короткомордый ньянзапитек назывался в числе возможных предков ореопитека (Harrison, 1986), а листоядный рангвапитек – орангутана, но трудно судить об этом уверенно из-за их фрагментарности.
Третье подсемейство проконсулид – афропитецины Afropithecinae. Его представители: Afropithecus, Equatorius и Morotopithecus – обитали в ранне– и среднемиоценовых лесах Восточной Африки 21,5–14 млн лет назад, а гелиопитек Heliopithecus leakeyi жил в Саудовской Аравии 17–18 млн лет назад. Таким образом, гелиопитек оказывается первым внеафриканским гоминоидом, с него можно вести отсчет выходов из Африки, закончившийся в конце концов расселением сапиенсов. Особо интересно, что гелиопитек примитивнее африканских родственников: этот факт мог определить пути эволюции всех последующих европейских и азиатских гоминоидов.
Моротопитек Morotopithecus bishopi – самый древний из афропитецин и один из самых древних гоминоидов, по части признаков он даже примитивнее гиббонов. По строению верхней части бедренной кости он уникален, а если на кого и похож, то на мартышкообразных. Зато моротопитек имел довольно крупные размеры – 40–50 кг – и общий план строения крупной человекообразной обезьяны. В отличие от многих родственников, он достаточно часто лазал в вертикальном положении, цепляясь за ветви преимущественно руками, и даже мог неспешно брахиировать. С некоторой вероятностью он является предком более поздних человекообразных обезьян.
Кстати, о вертикалях и горизонталях…
Вертикальное лазание стало одним из важнейших условий появления прямохождения. Утяжелившиеся человекообразные в некоторый момент перестали много прыгать и перешли к хождению по горизонтальным ветвям. Но ходить на четвереньках по ветвям крупному животному тоже не очень удобно. К тому же замедлившееся и передвигающееся по довольно толстым сучьям животное рискует попасться на обед хищнику. Доселе эта проблема решалась подвижностью, но как раз ее-то гоминоиды и растеряли. Из этого есть оригинальный выход – стать крупнее и сильнее, чтобы уже никакой леопард не посмел напасть (есть другой способ – стать мелким, чтобы хищнику было энергетически невыгодно гоняться за лилипутом, но это путь в интеллектуальный тупик, почти невозможный для высоко социальных человекообразных). Однако тяжелому зверю еще труднее ходить по веткам. Так развивается вертикальное лазание. Но и оно спасает не полностью, так что тенденция спускания на землю будет в такой эволюционной линии просто неизбежной. Что мы и видим в палеонтологической линии приматов. На земле, конечно, опасностей добавляется, но еще частично древесный зверь, который должен уметь подтягивать свое мощное тело на ветвях, имеет такую мускулатуру, что может дать затрещину любому хищнику. Гориллы тому примером.Из-за вертикального лазания тело приобретает новое положение, перпендикулярное тому, что было привычным от ланцетника доселе. Органы, до сих пор мирно подвешенные на арке позвоночника, начинают давить друг друга: сверху без конца стучит беспокойное сердце, ходят ходуном легкие, диафрагма давит на печень и желудок, а кишечнику и мочевому пузырю достается больше всего. А уж что делается с маткой… Ужас просто! Конечно, внутренние органы у приматов с вертикальным положением тела не смещаются слишком уж радикально, печень с сердцем не меняются местами, но определенные изменения, безусловно, происходят. Например, сердце теперь должно качать кровь вверх, а не вперед, особенно сложно вернуть венозную кровь обратно.Таким образом, положение тела при вертикальном лазании стало важнейшим залогом, частично поводом и даже гарантией развития прямохождения. Кстати, не в этом ли причина сохранения четвероногости у павианов, тоже вышедших в саванну? У них не было крупных вертикально лазавших предков, так что и появление в павианьей линии прямохождения было бы чудом. Некоторым суррогатом является типичная для павианов поза отдыха – сидя, с выпрямленной спиной, – но из статичной позы активного хождения на двух ногах не создашь.
Часть похожих находок из Кении и Уганды описана как угандапитеки Ugandapithecus – U. meswae, U. legetetensis, U. major и U. gitongai, в последовательном ряду которых размеры все время увеличивались (Pickford et al., 2009). Ugandapithecus major раньше назывался Proconsul major, но резко отличается от проконсулов строением скелета – не только размерами 60–90 кг, но и более существенными чертами. Однако самостоятельный статус угандапитеков признается не всеми исследователями.
Лучше всех из афропитецин изучен афропитек Afropithecus turkanensis, от которого сохранился почти целый череп и много более фрагментарных частей. Уникальность афропитека в резко сниженном половом диморфизме по размерам клыков, притом что сами клыки очень большие и очень толстые. Обычно у приматов, да и вообще любых млекопитающих, особенно нехищных, крупные размеры клыков однозначно указывают на мужской пол их обладателя (достаточно посмотреть на мандрилов). Если же самцы с самками отличаются не очень резко, то и клыки у них небольшие (чему примером может служить человек). Афропитек же являет собой разительное исключение: самцы и самки этого вида надежнее различались по строению премоляров, но не клыков. Судя по всему, такая особенность была следствием питания какими-то твердыми семенами или орехами, которые афропитеки грызли как раз клыками (Rossie et MacLatchy, 2013). Из-за этого же афропитеки имели характерную внешность: сильно выступающую вперед морду, широченные резцы, огромные височные мышцы и, соответственно, сагиттальный гребень на своде черепа, уменьшенные гайморовы пазухи, высокую нижнюю челюсть. Кстати, в меньшей степени то же было характерно для нахолапитека, а среди современных обезьян – для мохнатых саки Chiropotes и уакари Cacajao (нелишне вспомнить и ганлею), так что тут мы имеем очередной пример конвергенции.
Экваториус Equatorius africanus, живший 15,4–15,6 млн лет назад в Кении, судя по строению костей, вел полуназемный образ жизни (Patel et al., 2009). Конечно, он не достиг даже степени наземности павианов, но это был первый заход гоминоидов на сушу – маленький шаг для экваториусов, но огромный скачок для гоминоидов! Не пройдет и 10 млн лет, как потомки проконсулид полностью освоят африканскую саванну.
Уголок занудства
Чтобы Читатель проникся не только антропологическими байками, но нюхнул и настоящей науки, полезно привести без перевода на человеческий язык список признаков наземности и древесности экваториуса, позаимствованный из статьи, посвященной этому вопросу (Patel et al., 2009).Признаки, свидетельствующие о полуназемности: большой бугорок выступает выше головки плечевой кости; на дистальной части плечевой медиальный надмыщелок в состоянии ретрофлексии; проксимальная часть локтевой с выдающимся дорзально локтевым отростком, блоковидная вырезка глубокая, венечный отросток хорошо развит; головки метакарпалий с хорошо развитыми поперечными дорзальными гребнями и пальмарно широкой суставной поверхностью; фаланги кисти короткие, массивные и прямые; проксимальные фаланги кисти укорочены относительно метакарпалий сравнительно с большинством наземных мартышкообразных.Древесные или примитивные Proconsul-подобные черты: диафиз плечевой в состоянии ретрофлексии, выражен отдельный латеральный надмыщелковый гребень; лучевая кость прямая и грацильная; головка лучевой циркулярная; метакарпалии короткие, с дорзально суженными проксимальными суставными поверхностями; проксимальные фаланги кисти относительно метакарпалий длиннее, чем у постоянно наземных мартышкообразных типа Theropithecus и Papio.Для восприятия этой и прочей подобной тарабарщины надо бы для начала знать анатомические плоскости и направления. Сагиттальная плоскость проходит спереди назад и сверху вниз и делит тело на правую и левую стороны, парасагиттальная параллельна ей, но идет не строго посередине; фронтальная плоскость проходит сверху вниз и поперек, отделяет перед от зада; трансверзальная, она же поперечная плоскость делит тело поперек, для человека она же горизонтальная и будет проходить параллельно земле. Медиально – близко к середине, к центру тяжести (позвоночник в теле); мезиально – близкий термин, используется либо как синоним, либо как обозначение середины на некой протяженности (локоть на руке); латерально – сбоку (ухо на голове); дорзально – на спинной стороне (лопатка на туловище; в том числе на тыльной стороне кисти); вентрально – на брюшной стороне (грудина на туловище); краниально – в направлении переднего конца тела и головы; каудально – в направлении заднего конца тела и хвоста; проксимально – ближе к началу, телу и центру тяжести (плечо на руке, локоть на предплечье, запястье на кисти); дистально – ближе к концу, дальше от тела и центра тяжести (пальцы на руке, запястье относительно предплечья, локоть относительно плеча); волярно или пальмарно – на ладонной стороне кисти; плантарно – со стороны стопы; ульнарно – на предплечье или кисти со стороны локтевой кости и мизинца; радиально – на предплечье или кисти со стороны лучевой кости и большого пальца; лябиально или буккально – на зубе или челюсти со стороны губ или щеки; лингвально – на зубе или челюсти со стороны языка.
Otavipithecus namibiensis уникален своим географическим положением, ибо его челюсть найдена в Намибии, в местности, ныне явно не приспособленной для человекообразных обезьян. С некоторой вероятностью отавипитек родственен афропитецинам, но при сравнительно поздней датировке – 13 млн лет назад – он крайне примитивен и по части признаков сближается вообще с плиопитековыми и дендропитецидами. Возможно, Южная Африка была своего рода заповедником архаичных форм.
До сих пор я подчеркивал примитивные и негоминоидные черты проконсулид, так что Читатель вправе спросить: “Так почему же, собственно, они считаются первыми человекообразными?” Конечно, несколько больше признаков проконсулид сближают их именно с антропоидами. Например, сроки прорезывания постоянного первого нижнего моляра у афропитека были существенно большими, нежели у мартышкообразных, зато в точности как у шимпанзе (Kelley et Smith, 2003). Значит, детство было удлинено, а это – один из многочисленных шагов к разуму.
Дендропитециды (Dendropithecidae): Dendropithecus, Micropithecus и Simiolus – жили в начале и середине миоцена (20–14,5 млн лет назад) в Кении и Уганде. Одна из особенностей этих странных обезьян – мощные, недлинные, но толстые конические клыки у самцов. От микропитека Micropithecus clarki сохранились части черепа, так что мы знаем, что эти животные имели большие глазницы, широкое межглазничное пространство, сравнительно маленькую морду и грацильную нижнюю челюсть. Поэтому они, равно как и дендропитеки Dendropithecus macinnesi, назывались в качестве возможных предков гиббонов. Географически это выглядит странно, ибо дендропитециды неизвестны в Азии, а гиббоны – в Африке. К тому же дендропитециды бегали по веткам на четвереньках и не имели каких-либо приспособлений к брахиации.
От скелетов дендропитецид сохранилось не так мало костей, особенно от симиолуса Simiolus enjiessi, благодаря чему нам известно, что они обладали вытянутыми грацильными конечностями примитивного строения, напоминающими скорее вариант широконосых, нежели мартышкообразных или человекообразных обезьян (Rose et al., 1992). Таким образом, множество очень примитивных черт дендропитецид позволяют считать их параллельной остальным человекообразным линией эволюции, сохранявшей массу пережитков едва ли не эоценовых времен.
Некоторые древние человекообразные имеют своеобразную морфологию, позволяющую выделить их в отдельные группы. Например, несколько спорным статусом обладают грифопитециды Griphopithecidae – первые в основном неафриканские гоминоиды. Их номинативное подсемейство Griphopithecinae представлено единственным родом грифопитек Griphopithecus. Этот или очень похожий примат жил 17 млн лет назад на юге Германии (Böhme et al., 2011; Heizmann et Begun, 2001). От него сохранился лишь один поломанный зуб, но это первый европейский гоминоид. Другие виды грифопитеков: G. alpani, G. darwini и G. suessi – обитали около 14–15 млн лет назад в Турции, Австрии и Швейцарии. Это были довольно крупные древесные четвероногие поедатели фруктов, твердых плодов и орехов, жившие во влажных субтропических лесах. О них известно крайне мало, но грифопитеки были, вероятно, общими предками всех следующих неафриканских гоминоидов.
В целом похожие кениапитецины Kenyapithecinae жили 13,4–14 млн лет назад в Кении (Kenyapithecus wickeri) и Турции (K. kizili). Они тоже могли есть какие-то твердые семена, хотя точно это утверждать трудно.
Человекообразные в рыжих тонах:
Азия и Европа
Расселившиеся за пределы Африки гоминоиды дали новый всплеск разнообразия. Местами оно приобретало довольно экзотичные формы, как, например, на острове Гаргано в Средиземном море, где жил ореопитек Oreopithecus bambolii, выделяемый в самостоятельное семейство ореопитецид Oreopithecidae. Эта небольшая обезьяна отличалась массой специфических черт, в числе коих резко уменьшенные резцы, сравнительно маленькие клыки у самцов – свидетельство пониженной агрессивности и слабой иерархичности, необычайное сходство верхних и нижних моляров, центроконид – особый бугорок на молярах. Специфику скелета, полупрямохождение и умелые ручки ореопитека мы уже достаточно подробно разобрали выше, тут же повторимся лишь, что наиболее вероятные предки сего таинственного зверя обнаруживаются в Африке, где ньянзапитек и мабокопитек демонстрируют с ним замечательное сходство. Какими извилистыми путями кенийских обезьян занесло на сказочный северный остров – загадка зоогеографии.
Другая тайна – происхождение гиббонов.
Долой гипотезу гигантского гиббона!
Даешь гипотезу карликового гиббона!
Плиобатес из Испании – новый предок?
Известно великое множество ископаемых обезьян. Но сотни уже известных видов – явно капля в море реально существовавших разновидностей. Чем больше мы знаем, тем лучше мы понимаем, чего мы не знаем. И при этом известно, в каких слоях и каких местах лучше всего искать то, чего больше всего хотелось бы найти. Поэтому палеоприматологи рыщут по карьерам и оврагам всего мира в поисках чего-то новенького. И поэтому закономерно открытие не только новых видов, но и более крупных систематических единиц.А посему великое дело – описание плиобатеса Pliobates cataloniae, выделенного в самостоятельное семейство Pliobatidae (Alba et al., 2015). Его останки найдены в Северо-Западной Испании и имеют возраст 11,6 млн лет. Сохранность уникальна: палеонтологам редко достаются скелеты, включая череп, конечности и даже кости запястья и стопы.Череп плиобатеса выглядит очень странно: мозговая коробка относительно большая, хотя и приплюснутая, мордочка крохотная, глазницы огромные и совершенно круглые. При этом мозгов было не так уж богато – 69–75 см³, как не у самой крупной мартышки. Даже у гораздо более древних проконсулов мозга было в два с лишним раза больше, да и современные гиббоны обгоняют испанскую обезьяну в полтора раза. Особое внимание привлекают махонькие клыки, едва выступающие за уровень соседних зубов, отчего череп плиобатеса смахивает на остов вампира-недокормыша. Питался плиобатес в основном фруктами.Руки плиобатеса очень длинные и вытянутые. Судя по строению запястья, в основном он бегал по горизонтальным ветвям, хотя уже мог подвешиваться к веткам снизу. Однако он не умел лихо брахиировать подобно нынешним гиббонам, в этом смысле плиобатеса нельзя назвать слишком специализированным – хорошая черта для потенциального Великого Предка.Открытие плиобатеса проливает свет на эволюцию всех человекообразных. Дело в том, что мелкие обезьяны вообще редко и плохо сохраняются в ископаемой летописи. Из-за этого создается впечатление, что первые человекообразные – проконсулиды – были крупными, а лишь потом часть из них измельчала. Но так ли это? Размер тела плиобатеса был очень маленький – всего 4–5 кг, то есть с хорошую кошку.Отдельные черты плиобатеса выглядят очень примитивно и сближают его с загадочными дендропитеками, другие же весьма похожи на гиббоньи. Вместе с тем по совокупности признаков плиобатес лишь едва прогрессивнее проконсулид и примитивнее всех прочих человекообразных. Получается, что такими – мелкими древесными древолазами-недобрахиаторами – должны были быть общие предки гиббонов и крупных человекообразных, да и нас тоже. Стало быть, плиобатес – очередное достающее звено, Великий Предок?Авторы первоописания именно на этой пафосной ноте и заканчивают свое исследование: возможно, единый прародитель всех человекообразных был более гиббоноподобным, чем орангутано-шимпанзе-гориллоподобным.Что ж, байка про “питекантропа – гигантского гиббона” давно отправлена в разряд забавных выкрутасов истории палеоантропологии. Пришла пора “концепции карликового гиббона”?Впрочем, есть парочка зловредных но.Самое обидное, датировка плиобатеса не такая уж запредельная. В это же время в Африке некие обезьяны уже начали выглядывать из джунглей в саванну. Задолго до испанского зверя по Африке и Азии уже скакали весьма разнообразные человекообразные, и почти никто из них не был специфически схож с гиббонами.В частности, длинные кости рук плиобатеса выглядят все же чересчур вытянутыми. Конечно, они еще не были явно брахиаторными, но находились явно на пути к этому состоянию. Странно, если бы сравнительно короткие руки проконсулов сначала удлинились до состояния плиобатеса, а потом снова укоротились в большинстве линий, кроме гиббоньей. Некогда бурно обсуждавшаяся “брахиаторная гипотеза”, согласно которой предки человека были брахиаторами, в чистом виде давно признана несостоятельной, хотя, конечно, никто не отрицает, что элементы брахиации в локомоторном репертуаре древнейших человекообразных присутствовали.Как уже говорилось выше, и с мозгами у плиобатеса не фонтан. Неужели со времен проконсулов они сначала уменьшились, чтобы потом снова увеличиться?Кроме того, как часто бывает, подводит география. Конечно, не слишком вероятно, что предки гиббонов возникли на крайнем западе Европы, а потом расселились до Тихого океана и сохранились лишь в Юго-Восточной Азии и Индонезии. Вряд ли плиобатес был прямым предком гиббонов. Но из всех наличных ископаемых он ближе всего к ним.Сочетание неподходящей датировки и локализации с мозаикой генерализованно-специализированных черт может говорить о том, что плиобатес, скорее всего, являлся реликтовым потомком примитивных человекообразных, сохранившимся на краю Земли, тогда как в Африке и Азии эволюция уже дала гораздо более продвинутых обезьян. Кто-то очень похожий на плиобатеса, но еще более примитивный, живший раньше на полтора десятка миллионов лет и в Африке, действительно мог быть Великим Предком. Тогда проконсулиды – далеко не самые примитивные человекообразные, тогда многое сходится, но возникают и новые вопросы.Дело за малым: найти аналог плиобатеса, но в Африке и с датировкой 25–30 млн лет.
Подходящий кандидат на звание предка гиббона – юаньмоупитек Yuanmoupithecus xiaoyuan из Южного Китая (7,1–9 млн лет назад), однако до крайности плохо известный – всего лишь по нескольким зубам.
Гиббоновые Hylobatidae населяют с плиоцена поныне Юго-Восточную Азию и Индонезию. Резкая специализация, очевидно, ставит их особняком среди прочих человекообразных. Прямые предки гиббоновых точно не известны, на их роль выдвигались разные ископаемые приматы – Propliopithecus из олигоцена Египта, Pliopithecus и Crouzelia из миоцена Европы, Laccopithecus и Yuanmoupithecus из миоцена Китая, Dionysopithecus из миоцена Китая, Пакистана и Таиланда, Krishnapithecus из миоцена Индии, Dendropithecus, Limnopithecus и Micropithecus из миоцена Восточной Африки. Низведение корней гиббоновых аж к нижнему олигоцену создало мнение о более раннем их отделении от прочих гоминоидов, чем даже проконсулид, а резкие отличия от человека – о примитивности в сравнении с крупными человекообразными. Этому немало способствовали устоявшиеся стереотипные представления о человеке как венце эволюции, шимпанзе – как почти-человеке, дриопитеках – как почти-шимпанзе, а проконсулах – как почти-дриопитеках. В реальности же проконсулиды заметно примитивнее, специализации гиббоновых не примитивны, а, напротив, очень и очень продвинуты, а сама филогенетическая линия может быть далеко не столь древней. Гиббоны запросто могли появиться в самом конце миоцена или даже в плиоцене; древнейший достоверный обнаружен Эженом Дюбуа в Триниле вместе с питекантропом. Известные ископаемые гиббоны принадлежат уже к современным родам и даже современным видам, так что почти не отличаются от них. Таким образом, фактически гиббоны являются высшим и самым продвинутым вариантом эволюции человекообразных.
Гиббоновые – животные не очень большие, весом от 5 до 12 кг; самые отъетые сиаманги могут достигать 23 кг. Они имеют грацильный череп с ослабленным рельефом, маленькую морду с небольшими зубами, но длинными тонкими клыками, относительно тела крупный, а на самом деле маленький головной мозг – около 100 г, примерно как у среднеразмерных мартышкообразных. Руки гиббонов чрезвычайно удлинены и намного больше длины ног и тела. Такие пропорции обусловлены замечательным способом локомоции – брахиацией, то есть передвижением только с помощью рук. Гиббон раскачивается на ветке на одной руке, перебрасывается на другую ветку другой, а ноги в этом никак не участвуют. Специализация к брахиации у гиббонов так велика, что они не могут передвигаться по земле на четырех ногах и при необходимости спуститься на землю ходят на двух, поднимая руки вверх. Пальцы кисти образуют крючок, удобный для цепляния за ветви. Большой палец на руке замечательно длинный – вдвое больше, чем у человека, но практически не противопоставляющийся остальным, тем более что из-за крайней удлиненности пясти кончик большого пальца достает только до основания остальных. Из-за этого гиббонам крайне трудно что-либо брать рукой, они очень неуклюже сгребают предметы всей кистью. Она у гиббонов рассчитана на растяжение, а не сжатие: в частности, в запястье есть особая кость os Daubentonii, расположенная в толстом мениске, который укрепляет соединение шиловидного отростка локтевой кости с гороховидной костью.
Кстати, о скалолазах…
Измерения профессиональных скалолазов показали, что у них сравнительно с обычными людьми весьма короткие ноги. Длинные ноги много весят, а это очень мешает при карабканье по вертикальным стенам. Так что у эволюции всегда есть возможности для неожиданных поворотов: возникни необходимость, из человека запросто может получиться брахиирующее чудо с мощными длинными руками и коротенькими, но тоже цепкими ножками.
Специализированной чертой гиббонов являются маленькие седалищные мозоли, иногда подчеркнутые длинной шерстью выделяющегося цвета; на седалищных костях для их крепления имеются большие плоские поверхности. Скорее всего, седалищные мозоли гиббонов возникли конвергентно с мозолями мартышкообразных, а не являются общим древним наследием, за это говорят особенности их строения, прикрепления и развития (McCrossin et Benefit, 1992). Тут же можно упомянуть, что седалищные подушки шимпанзе устроены совсем иначе (состоят в основном из жира, а не утолщенной кожи). С изобретением стульев человек тоже вступил на скользкую дорожку, ведущую к развитию седалищных мозолей; коли у родственников они появлялись, то куда и нам деваться.
Гиббоны многочисленны по меркам современных человекообразных – специалисты выделяют от пяти до шестнадцати видов, объединяемых иногда в один род Hylobates, а иногда разделяемых на Hylobates, Bunopithecus, Nomascus и Symphalangus, которым не менее часто придается подродовой статус. Впрочем, принадлежность к разным родам или подродам не мешает гиббонам давать гибридов. Все это свидетельствует о значительном сходстве разных гиббонов и наверняка об их совсем недавнем эволюционном происхождении. Самые известные – белорукий гиббон H. lar, белощекий H. concolor, серебристый H. moloch и белобровый H. hoolock.
Белорукие H. lar и чернорукие гиббоны H. agilis имеют несколько контрастных вариантов окраски – черный, красно-коричневый и белый, а у китайского белощекого H. leucogenys, белощекого H. concolor и белобрового H. hoolock самцы черные, а самки белые или золотистые.
Самый большой гиббон – сиаманг Hylobates (Symphalangus) syndactylus или Symphalangus syndactylus – отличается срастанием указательного и среднего пальцев на стопе (часто ошибочно пишут, что у них сращены пальцы на кисти); за такое строение сиамангов называют еще симфалангами. Вторая особенность сиамангов – огромный горловой мешок у самцов и самок, позволяющий им петь громче всех прочих человекообразных, кроме разве что тирольцев и оперных примадонн. Также сиаманги – самые длиннорукие приматы из всех: их руки в полтора раза длиннее ног.
Живут гиббоны семейными группами – папа, мама и детеныш. Эти обезьяны уникальны своими вокальными способностями – они поют по утрам на вершинах деревьев всей семьей, чтобы соседи знали, чья тут территория.
Кстати, о птичках…
Гиббоны, как известно, поют. Но этим занимаются не только они: во-первых, иногда поют и люди (хотя гораздо реже гиббонов, которые делают это ежедневно), во-вторых, многие птицы. При этом часть птиц поет вполне творчески, им надо долго этому учиться, у них бывают диалекты и индивидуальные уровни мастерства, бездари и таланты (это не касается некоторых халтурщиков типа петухов и кукушек, чьи вокализации на сто процентов врожденны и ни от чего не зависят). Люди еще и говорят и, что характерно, тоже этому долго учатся. Удивительно, но у птиц, поющих с обучением, генетика и нейрофизиология пения оказались необычайно похожи на генетику и нейрофизиологию речи у людей (Pfenning et al., 2014). За то и другое отвечают очень похожие комплексы генов и близкие по происхождению и функциям участки мозга. Ясно, что способности эти развились совершенно независимо, так что тут мы имеем замечательный пример генетической конвергенции. Более того, разные поющие птицы приобрели свои замечательные способности независимо друг от друга, ведь единого предка-певца никогда не существовало.Было бы чрезвычайно интересно сравнить в том же аспекте гиббона и человека: думается, наши дарования тоже имеют независимое происхождение. Судя по всему, некоторые сложные признаки можно реализовать не столь уж большим количеством генетических путей: сколь ни богата природа на выдумки, все же и она имеет ограничения.
Следующие гоминоиды отличаются друг от друга уже совсем незначительно, а потому обычно объединяются в рамках одного семейства. Тут появляются варианты, как это семейство называть. Традиционно крупных человекообразных разделяли на понгид и гоминид. Отличия человека от орангутанов, горилл и шимпанзе вроде бы слишком очевидны, чтобы сваливать их в кучу. Однако филогения позволяет классифицировать их иначе. Ведь по пути к человеку раньше всех отделилась линия орангутанов, намного позже среди африканских человекообразных – линия горилл, затем шимпанзиная разошлась от человеческой, а уж в самом конце появились два вида шимпанзе и разные виды людей, из которых все, кроме одного, исчезли. Таким образом, филогенетически получается, что человек ближе к шимпанзе, чем шимпанзе к горилле, а к горилле ближе, чем горилла к орангутану.
Уголок занудства
Кладистика – способ построения систематики на основе исключительно порядка ветвления филогенетического древа. Все потомки одного предка считаются принадлежащими к одной таксономической группе. В кладистике используются свои термины, придающие ей дополнительную внешнюю беспристрастность, политкорректность и наукоподобие. Плезиоморфный признак – присущий предкам и общий для всех потомков, по-простому – примитивный; симплезиоморфия – родство по такому признаку – считается незначимым и не учитывается при построении схемы. Апоморфный признак характеризует только какую-то конкретную группу живых существ, по-человечески такой называется специализированным или продвинутым; синапоморфия – родство по такому признаку – признается единственно значимым для установления близости групп. Гомоплазия – сходство признаков в разных группах, возникшее независимо, – то же самое, что результат конвергенции.В кладистике фактически игнорируются идеи об ароморфозах и идиоадаптациях, уровнях и значениях эволюционных преобразований. Это сделано с целью объективизировать систему, избавить ее от оценочных суждений. Построение филогенетических деревьев по возможности автоматизировано и формализовано. Это удобно, особенно для не слишком опытных морфологов, к тому же ограниченных во времени грантами и сроками сдачи статей в печать. Однако выбор анализируемых признаков по-прежнему остается за человеком, а применяемый при их обсчете кластерный анализ дает не столь однозначные результаты, как хотелось бы, так что мнение исследователя исключить все же не удается. Посему кладистический подход в систематике, с одной стороны, завоевал почти абсолютное господство в современной биологии, с другой – постоянно критикуется.
Наиболее крайний вариант систематики предложили молекулярные биологи (Goodman et al., 1998). Согласно им, семейство Hominidae состоит из одного подсемейства Homininae (неясно, зачем его тогда вообще выделять, но так сделано в первоисточнике), в котором выделяются три трибы. Первая триба – Oreopithecini. Вторая – Hylobatini – с единственной подтрибой Hylobatina включает два монотипичных рода: Symphalangus syndactylus и Hylobates lar. Вторая триба – Hominini – включает две подтрибы. Первая – Pongina с сивапитеком Sivapithecus и орангутаном Pongo. Вторая – Hominina с четырьмя родами: Samburupithecus, Graecopithecus, Gorilla и Homo. Род Homo подразделяется на два подрода: шимпанзе H. (Pan) (с видами H. (P.) troglodytes и H. (P.) paniscus) и H. (Homo) sapiens, сюда же вклиниваются австралопитеки (Goodman et al., 1998). В принципе, если попытаться привести систематику водорослей, жуков и приматов к общему знаменателю, такая схема вполне оправдана. Жаль только, в ней напрочь растворяются почти все ископаемые формы – им просто не хватает места; на практике такой схемой невозможно пользоваться. Посему она не нашла доброго отклика в сердцах антропологов, а лишь осталась навеки в истории науки как величайшая попытка низвести человека с высот исключительности.
Существует огромное количество других вариантов классификации крупных человекообразных, в подавляющем их числе признается лишь одно семейство гоминид Hominidae, но мы используем более консервативную версию, дабы подчеркнуть различия и специализации разных групп. О неисключительности человека и так было довольно сказано выше, пора обратить взор на специфику.
Понгиды Pongidae (или Anthropomorphidae) – крупные человекообразные обезьяны, населявшие со среднего миоцена экваториальные и тропические леса Африки, Европы и Азии. Четыре вида сохранились по наши дни, но ископаемых форм гораздо больше, и они гораздо разнообразнее.
Азиатская часть филогенетического куста представлена рамапитецинами Ramapithecinae, гигантопитецинами Giganthopithecinae, дриопитецинами Dryopithecinae и понгинами Ponginae.
Рамапитецины Ramapithecinae – Sivapithecus и Ankarapithecus – жили от 12,5 до 8 млн лет назад. Подавляющая часть останков найдена в Сиваликских холмах на стыке Пакистана, Индии и Непала. В первой половине XX века была описана целая куча родов и видов сиваликских приматов, в ход шли почти все, даже самые фрагментарные находки. Сейчас признается от трех до пяти видов сивапитеков Sivapithecus: самый древний и некрупный S. indicus (10,5–12,5 млн лет назад, самцы 30–45 кг, самки 20–25 кг); крупнейший S. parvada (10,1 млн лет назад, самцы 60–75 кг, самки 30–45 кг), их потомок S. sivalensis (8,5–9,5 млн лет назад, промежуточного между предыдущими размера), а также его возможные синонимы S. punjabicus и S. simonsi. Разница между всеми ними не такая уж большая, что наглядно видно по диагностике самого целого черепа, определявшегося и как S. indicus, и как S. punjabicus, и S. sivalensis.
Кстати, о чехарде…
Как известно, ученые, занимающиеся классификацией, делятся на “дробителей” и “объединителей”. Так было всегда и пребудет вовек. Первые склонны давать новое название каждому древнему огрызку, вторые готовы слить воедино и слона с тенреком. Посему систематика ископаемых гоминоидов чрезвычайно запутанна. Исторически так сложилось, что первыми стали известны европейские дриопитеки, затем азиатские сивапитеки, а после – африканские проконсулы. Чаще всего одни и те же находки обозначались разными авторами под несколькими названиями, которые потом могли оказаться разнесенными даже в разные семейства. Потому одни и те же слова встречаются в самых разных сочетаниях: Proconsul africanus назывался Dryopithecus africanus, Sivapithecus africanus и Kenyapithecus africanus; Equatorius africanus имеет частичные синонимы Kenyapithecus africanus, Griphopithecus africanus, Proconsul nyanzae и Sivapithecus africanus; Nyanzapithecus vancouveringorum в литературе может быть помянут как Dryopithecus (Rangwapithecus) vancouveringi, Proconsul vancouveringi или Rangwapithecus vancouveringi; Ugandapithecus major – как Proconsul major, Dryopithecus (Proconsul) major или Morotopithecus bishopi. Род Sivapithecus – это то же, что Ramapithecus, Bramapithecus, Palaeopithecus, Anthropopithecus, Sugrivapithecus и Dryopithecus (Sivapithecus), тогда как Dryopithecus – Adaetontherium, Anthropodus, Neopithecus, Hispanopithecus и Hungaropithecus. Очень не повезло роду кениапитеков, ведь под названием Kenyapithecus africanus кроются останки и Proconsul africanus, и Equatorius africanus, и Mabokopithecus clarki, и Kenyapithecus wickeri. Разобраться в этом балагане крайне трудно даже узким специалистам, особенно учитывая крайнюю фрагментарность большинства находок.
От сивапитеков сохранилось огромное количество челюстей и зубов, а также часть черепа. Череп замечательно похож на орангутаний: с высокой вогнутой мордой, резко выступающими челюстями, вытянутыми вертикально овальными глазницами, высоко поднятыми над носовым отверстием, резко суженным межглазничным пространством. Нижняя челюсть весьма тяжелая. Верхние центральные резцы очень широкие, а боковые маленькие, клыки умеренно крупные, моляры с толстой эмалью и без цингулюма.

Рис. 39. Черепа Sivapithecus (а), Ankarapithecus (б) и Dryopithecus (в).
Найдены и кости посткраниального скелета сивапитеков, хотя сравнительно немногочисленные. Они замечательно отличаются от соответствующих костей современных человекообразных. Например, верхняя часть плечевой больше напоминает вариант павианов, а нижняя – африканских человекообразных; нижняя часть большой берцовой формой суставной поверхности похожа на таковую у мартышкообразных, но показателем сечения и внутренней лодыжкой – человекообразных, прежде всего шимпанзе; крючковидная кость не имеет крючка. Кости запястья приспособлены к костяшкохождению – с опорой на согнутые пальцы (Begun et Kivell, 2011); этот способ, видимо, возникал среди человекообразных независимо как минимум трижды: в линиях, ведущих к орангутанам, гориллам и шимпанзе. Однако строение фаланг указывает, что сивапитек ходил с опорой на ладонь (Rose, 1986). Не очень ясно, что значат все эти противоречия. Видимо, сивапитеки были очень своеобразными животными: на лицо орангутанами, руками и ногами – частично павианами, частично шимпанзе. Понятно только, что прямого аналога им сейчас нет.
Таз сивапитека
Люди встали на две ноги примерно 7–6 млн лет назад. Часто по умолчанию считается, что предки были в принципе похожи на современных человекообразных обезьян – орангутанов, горилл и шимпанзе, а то и гиббонов. Дескать, скакали по веткам этакие “протошимпанзе”, потом слезли с дерева и стали ходить на двух ногах. А кто не слез, тот так и прыгает до сих пор по лесам. Обезьяны сохранили примитивные черты, мы изменились.Но так ли это?На самом деле, много ли мы знаем о способе передвижения древних приматов? Как это вообще узнать? Ведь чаще всего от них находят зубы и обломки челюстей, кости скелета сохраняются редко и плохо. Если в Африке еще найдены несколько скелетов проконсулов, то в Азии с этим совсем беда. До сих пор бесчисленные сивапитеки и их родичи известны почти исключительно по зубам, мы более-менее представляем, что они ели, но как они добирались до плодов – пока оставалось загадкой.Но все тайное когда-нибудь становится явным. В 1990 году в Пакистане, на плато Потвар, была найдена тазовая кость. Через двадцать лет – в 2010 году – она была определена как кость примата. А в 2015 году вышла статья с ее описанием (Morgan et al., 2015). Судя по месту обнаружения и датировке – 12,3 млн лет назад, кость принадлежала Sivapithecus indicus, потому что ранее тут нашли верхнюю челюсть и три зуба именно этого вида. Размеры кости позволили предположить, что это была самка весом около 20–25 кг – как раз подходящий размер, ранее предполагавшийся на основании размеров зубов.Исследователи сравнили таз сивапитека с костями множества прочих обезьян. Выводы были довольно неожиданными.Судя по совокупности признаков таза, сивапитеки были неторопливыми четвероногими приматами, предпочитавшими осторожно и даже вдумчиво ходить по ветвям, как более-менее горизонтальным, так и по вертикальным и расположенным наискосок – в любых направлениях, способными далеко отводить конечности в сторону и даже подвешиваться на трех или четырех лапках, главное – без экстремальных прыжков и легкомысленных подвешиваний и раскачиваний на руках. Впрочем, строение рук говорит все же о некоторых способностях к подвешиванию на ветках; очевидно, разные части скелета эволюционировали с разной скоростью.Таз сивапитека был ориентирован вдоль позвоночника, как у мартышкообразных. У современных человекообразных крылья подвздошной кости все же несколько расходятся в стороны; человек же являет крайний вариант в этом ряду. “Поясничный индекс” – соотношение ширины поясницы к ее передне-заднему диаметру – у человекообразных обезьян больше 120, то есть “талия” очень широкая. У широконосых обезьян Южной Америки это же соотношение примерно равно 100, то есть они круглые в сечении. Мартышкообразные имеют индекс меньше сотни, как и сивапитек, у которого указатель равен 90–100.Таз сивапитека очень массивен. Учитывая не слишком большие размеры тела, такое усиление говорит о значительных нагрузках, передающихся от опоры, и – в совокупности с другими параметрами (например, усилением подвздошной бугристости) – о четвероногом передвижении.Ширина седалищного бугра у приматов здорово коррелирует с наличием или отсутствием седалищных мозолей. Сивапитек, что приятно, попадает все же в группу без мозолей, то есть ближайших человеческих родственников.Вертлужная впадина сивапитека очень глубокая, что означает усиление тазобедренного сустава, чем сивапитек напоминает мартышковых. Однако по площади полулунной поверхности (суставной поверхности вертлужной впадины) он ближе к паукообразным широконосым и крупным человекообразным обезьянам. От другого вида – Sivapithecus sivalensis – известна бедренная кость, ее головка круглая, почти полностью погруженная в вертлужную впадину. Сочетание получается необычное: с одной стороны, сустав очень прочный, с другой – весьма подвижный, способный к вращательным движениям. Обычно это вещи не то чтобы взаимоисключающие, но, скажем осторожно, плохо сочетающиеся. Вероятно, это связано с четвероногим хождением по ветвям в сочетании с отсутствием хвоста. Хорошо мартышкам бегать по ветвям – они балансируют длинным хвостом. А попробуйте удержаться на суку, будучи довольно крупным зверем и без никакого уравновешивателя. Придется активнее двигать поясницей, а это дает повышенные нагрузки на суставы, что и вызывает тот странный комплекс, который мы видим у сивапитека.Кстати, можно вспомнить, что и плечевые кости сивапитеков больше схожи с мартышковыми, а не орангутаньими. То же можно сказать и о других известных посткраниальных костях сивапитеков.Кстати, по всем основным параметрам новообретенный таз мало отличается от таза проконсула Proconsul nyanzae. По многим частным параметрам и общему комплексу проконсул оказывается закономерно более мартышкообразным, чем сивапитек. Впрочем, у сивапитека больше черт вообще широконосых обезьян. Сивапитек отличался от проконсула одновременно большей подвижностью суставов ноги и большей же их прочностью.Но челюсти с зубами никто не отменял. А по ним сивапитеки – ближайшие родственники орангутанов. Исследователи честно признаются, что пока не знают, как объяснить такое расхождение (тут самое место появиться креационистам и радостно закричать: “АГА!!!” – но мы не дадим им этого шанса). Есть две гипотезы. Согласно первой, сходство сивапитеков и орангутанов по зубам является конвергентным, случайным, возникшим в силу близкой диеты. Конвергенция – обычнейшее явление в живом мире. Среди приматов тому тоже немало примеров. Можно, скажем, вспомнить челюсти колобусов и ореопитеков или плезиадаписов и руконожек. Ясно, это не отменяет факта, что сивапитеки – человекообразные приматы, но они могут представлять особую тупиковую линию развития.Второй вариант: сивапитеки действительно родственны орангутанам, но отличались способом передвижения. В строении конечностей конвергенция распространена не реже, если не чаще, чем в зубах. Скажем, кисти паукообразных, колобусов и гиббонов близки по целому ряду показателей; если задача – зацепиться за ветку и быстро ее отпустить, то решение будет одинаковым, тем более что исходник-то общий, все же так или иначе родственники. Теоретически эволюция конечностей может совершаться очень быстро, так что не исключено, что вплоть до 10 млн лет назад или даже дольше способ передвижения человекообразных был совсем не таким, как сейчас. В этом случае нынешнее состояние орангутанов, горилл и шимпанзе – столь же новомодное эволюционное изобретение, как и прямохождение человека.Интуитивно антропологи часто склонны отдавать первенство важности черепу, но нет ли в этом “культа черепа”? Поэтому исследователи таза сивапитека корректно не дают преимущества ни одной из двух версий. Будет больше костей – будет больше знаний.
Ближайший и синхронный родственник гималайских сивапитеков – анкарапитек Ankarapithecus meteai – найден в Центральной Турции. От него сохранилась часть лица, в целом аналогичная сивапитечьей и орангутаньей, хотя не такая плоская. Нижняя челюсть тоже похожа на таковую сивапитеков – высокая и очень массивная.
Гигантопитеки Giganthopithecinae были поздними потомками азиатских сивапитеков. Возможное достающее звено между ними символизируют два моляра из Сивалика в Пакистане с древностью 9–11,6 млн лет назад, описанные как Chinjipithecus atavus, но выглядящие как уменьшенные копии зубов гигантопитека (Koenigswald, 1981).
Надежно выделяются два вида гигантопитеков: более древний и мелкий североиндостанский Gigantopithecus giganteus (он же G. bilaspurensis и Indopithecus giganteus; 10,1–6 млн лет назад) и молодой крупный южнокитайско-вьетнамско-яванский G. blacki (известный также как Giganthropus; 1,96–0,38 млн лет назад). Гигантопитеки – феноменальные животные – известны из большого числа местонахождений и в огромном интервале времени. Однако, как уже упоминалось, от них сохранились сотни зубов и ни одного черепа, лишь пять нижних челюстей позволяют хоть как-то представить внешний вид этих колоссальных зверей.
Обычно гигантопитеков воспроизводят как неких кинг-конгов немереного роста. В действительности аллометрические закономерности могли выразиться в быстрейшем росте челюстей сравнительно с габаритами тела, так что в реальности гигантопитеки, вероятно, были всего на пятую часть или четверть длиннее, но в полтора-два раза тяжелее горилл (Johnson, 1979).
Нижние челюсти гигантопитеков поражают масштабами – в полтора раза больше, чем у гориллы, у которой морда вообще-то тоже не кукольная. Зубная система уникальна для приматов: моляры, премоляры и даже клыки схожей формы, огромных размеров, с бугристой и складчатой эмалью, чуть ли не как на зубах копытных травоядных. Особенное же сходство моляры гигантопитеков имеют с зубами панд – удивительная конвергенция приматов и медведеенотов. Нижние резцы этих странных обезьян относительно уменьшены, а клыки толстые, но короткие. Судя по всему, гигантопитеки были огромными фрукто– и листоедами, практически гарантированно наземными – под огромным весом деревья просто ломались бы.
Кстати, о натюрмортах…
Бамбук превосходен тем, что его мало не бывает, а растет он очень быстро, это сорняк, который захочешь – не изведешь. Но он же имеет тот огромный минус, что после цветения, которое захватывает сразу все бескрайние заросли одновременно, эта деревянистая трава сохнет и гибнет. Цветение бамбука случается едва ли не раз в сто лет, но это критическое время, когда все зависимые от него животные попадают под лезвие естественного отбора. Трагична не только гибель от голода, но и резкое сокращение генетического разнообразия; возникает так называемое “бутылочное горлышко” – выживают единицы, и в следующее поколение переходит очень малая часть вариантов генов. А суженная изменчивость чревата уязвимостью при последующих вызовах природы. Когда же “бутылочные горлышки” повторяются регулярно (а в случае с бамбукоедами это практически неизбежное проклятие), то все представители вида становятся фактически клонами. Любая недобрая мутация или существенное изменение окружающей среды – колебание климата, эпидемия, новый хищник – запросто может сгубить всю популяцию в одно мгновение. Если гигантопитеки действительно были специализированными пожирателями бамбука, остается удивляться, как они вообще протянули столько времени.Цветение бамбука приводит к бутылочному горлышку – такой вот смертельный эволюционный натюрморт мог свести гигантопитеков в могилу…Впрочем, новейшие исследования показывают, что гигантопитеки не были слишком специализированы и питались самой разнообразной лесной растительностью. Их исчезновение было вызвано, видимо, банальным сокращением тропических лесов во время похолоданий.
Дриопитецины Dryopithecinae: Anoiapithecus, Dryopithecus и Lufengpithecus – своеобразные приматы, жившие от 12,3 до 8,5 млн лет назад в Европе и от 11 до 1,9 млн лет назад или даже позже в Китае.
Специфический и самый древний представитель – анойяпитек Anoiapithecus brevirostris – известен из отложений в Каталонии с возрастом 12,3–11,9 млн лет. Название – “короткомордый” – лучше всего характеризует его главную особенность.
Наиболее известны и лучше всех изучены собственно дриопитеки Dryopithecus. Их многочисленные останки описывались из самых разных европейских стран с середины XIX века. Попервоначалу и сгоряча, за неимением более подходящих кандидатов, дриопитеки были зачислены в предки человека; эта версия до сих пор тлеет в школьных учебниках, хотя уже сто лет назад была отвергнута в науке. Сейчас ясно, что дриопитеки были в некотором роде аналогом европейских орангутанов.
Уголок занудства
За полтора столетия было придумано много альтернативных названий для дриопитеков, но сейчас большинство исследователей выделяют всего от трех до пяти видов. D. fontani – самый древний вид, обитавший во Франции, Австрии и, видимо, Северной Испании 11–12 млн лет назад; D. crusafonti – испанский дриопитек (возможно, синоним D. laietanus), 10,5 млн лет назад; D. brancoi – его многочисленные останки найдены в Венгрии, а один зуб – в Германии, 9,7 млн лет назад; D. laietanus – самый поздний испанский вид, 9,6–9,7 млн лет назад. Иногда испанские D. crusafonti и D. laietanus выделяются в особый род Hispanopithecus. К нему может присоединяться D. brancoi, но он же часто определяется как Rudapithecus hungaricus, так как голотипом (образцом при первоописании) для D. brancoi послужил третий моляр из Германии, который, строго говоря, мог принадлежать кому угодно, а большинство находок Rudapithecus сделано в Венгрии. Венгерские находки стали также основой для описания видов Ataxopithecus serus и Bodvapithecus altipalatus, никем ныне не признающихся за самостоятельные. В Грузии найден кусок верхней челюсти с двумя зубами, названный Udabnopithecus garedziensis, но с наибольшей вероятностью принадлежащий все к тому же виду D. brancoi.Кроме того, в Китае раскопана нижняя челюсть с возрастом 6–8 млн лет назад, получившая название D. wuduensis; она вполне похожа на челюсти дриопитеков, сомнение вызывает в основном географическое положение – больно уж далеко от Европы.
Дриопитеки были животными размером несколько меньше современных карликовых шимпанзе – 15–45 кг, в целом похожими на сивапитеков и орангутанов: с резко покатым лбом и умеренным надбровьем, округлыми глазницами, весьма широким носовым отверстием, не слишком вытянутой мордой с высоким альвеолярным отростком и скошенными скулами (чем заметно отличаются от сивапитеков и анкаропитека), очень мощной нижней челюстью, большими клыками, узкими и длинными центральными верхними резцами. Наглядным отличием от сивапитеков является тонкая эмаль моляров, сходством – отсутствие цингулюма.
Скелет дриопитеков известен достаточно хорошо, особенно благодаря находкам в Рудабанье в Венгрии и Кан-Йобатерес в Испании. Тело их было приспособлено как к четвероногому хождению по ветвям, так и – даже в большей степени – к вертикальному лазанию и подвешиванию на ветвях; такого сочетания типов передвижения нет среди современных человекообразных. Достижением дриопитеков стало удлинение рук – наконец-то эта специфическая черта современных человекообразных впервые проявилась у явных ископаемых родственников. У более древних проконсулид руки были короткие; ореопитеки оказались еще более длиннорукие, но они жили в среднем чуть позже дриопитеков. Пальцы на руках дриопитеков были тоже длинные и сильно изогнутые, явно приспособленные для цепляния за ветви. В целом строение рук больше всего похоже на вариант орангутанов, что, в общем-то, закономерно. Вместе с тем позвоночник больше похож на вариант широконосых и гиббонов, а большая берцовая – макак и гиббонов, чем современных крупных человекообразных обезьян.
Половой диморфизм у дриопитеков был достаточно сильным, что, впрочем, типично почти для всех мало-мальски крупных обезьян.
Кстати, о Рудабанье…
В Венгрии, в замечательном местонахождении Рудабанья, обнаружены останки двух видов приматов – плиопитецид Anapithecus hernyaki и дриопитеков Dryopithecus brancoi. Замечательно, что они были схожих размеров и образа жизни. Возникает закономерный вопрос: как же они решали проблему конкуренции? Исследование тонкостей строения моляров и микростертости эмали показало, что, хотя и анапитеки, и дриопитеки ели в основном сочные фрукты, первые разбавляли их листьями, а вторые – твердыми плодами или орехами (Deane et al., 2013).На основании половозрастного распределения останков в Рудабанье было сделано предположение, что дриопитеки D. brancoi охотились за детенышами анапитеков A. hernyaki: среди первых встречены почти только взрослые особи, а среди последних много детенышей (Kordos et Begun, 2001). Однако более тщательные исследования показали, что, во-первых, разница частот не столь уж очевидна, а самое главное – останки двух видов залегают в несколько разных отложениях, так что они, видимо, предпочитали различные условия жизни (Armour-Chelu et al., 2005). Так что картина охоты “почти-шимпанзе” за “почти-гиббоном”, видимо, останется не воплощенной в красках…
Позднейшие и географически обособленные дриопитецины люфенгпитеки Lufengpithecus найдены в Юго-Западном Китае. Как обычно бывает, исследователи разродились фонтаном имен: L. keiyuanensis, 8,3–10,5 млн лет назад (он же S. xiaolongtanensis, S. yunnanensis и Dianopithecus progressus, не говоря о комбинациях с родовыми названиями Laccopithecus, Dryopithecus, Sivapithecus и Ramapithecus), L. hudienensis, 7,1–8,2 млн лет назад (он с большой вероятностью может быть синонимом L. keiyuanensis и точно синоним Lufengpithecus yuanmouensis, Sinopithecus xiaoheiensis, Homo erectus zhupengensis или H. habilis zhupengensis, а также H. orientalis – просто праздник какой-то!), и L. lufengensis, 6,2–6,9 млн лет назад (который тоже фигурирует в литературе как минимум под 13 названиями!).
Люфенгпитеки известны по четырем черепам, распределенным по-даосски гармонично: два детских и два взрослых, из коих один – самца и один – самки. Правда, все эти черепа не слишком гармонично расплющены в лепешки, но и в этом можно найти свое дао. Реконструированные черепа в чем-то схожи с дриопитеками – верхние центральные резцы узкие и длинные, а в чем-то отличны – глазницы прямоугольные, верхний край носового отверстия расположен выше нижнего края глазницы, клыки у самцов тонкие и очень длинные, эмаль зубов толстая.
Три сказа о люфенгпитеках
Сказ первый: о том, как мудрецы взглянули на лик Люфенгпитека и как Люфенгпитек Китай и Европу сроднил
Хорошей сказке полагается начинаться примерно так: давным-давно, в незапамятные времена (1988 г.), в далеком-предалеком Южном Китае, в цветущей долине Юаньмоу, один трудолюбивый китаец рубил бамбук / копал канаву / искал козу, да и нашел череп. Удивился китаец премного и отнес череп Великим Мудрецам, жившим в Университете. А был то череп детеныша обезьянки. Он был маленький, на левый глаз кривенький, но все ж красивый. И за это назвали его YV0999, что в переводе значит Lufengpithecus hudienensis. Все любовались черепом, и многие мудрецы писали про него ученые статьи. Однако ж нелегко сложилась судьба YV0999: лишь одна черно-белая зернистая фотография его гуляла по бескрайним просторам интернета, и несмотря на то, что часто поминали его, существовал он в полном одиночестве. Никто не хотел сравнивать его с детенышами других обезьян, и полноценного анализа за последующие 24 года (конечно, эпичнее было бы тридцать лет и три года, но, видать, мудрецы поторопились) ни один мудрец так и не сделал. Но всякому сроку приходит свой конец. Два мудреца, жившие по разные стороны Великого Океана, – Джей Келли из пыльной Аризоны и Фенг Гао из дождливой Юньнани – удивительной силой интернета соединили свои невероятные усилия и сняли проклятье одиночества с YV0999 (Kelley et Gao, 2012). Навострили они волшебные Циркули Скользящие, пошли в хранилища заветные и разом измерили 10 черепов детенышей шимпанзе, 9 – горилл и 11 – орангутанов (лучше б, конечно, сорок сороков, но столько не нашлось, а как по сусекам поскребли, по амбарам помели – вот и набралось 30 черепов, это вам не две горсти все ж!) ажно по 17 признакам (лучше б, конечно, по тьме тьмущей и чтоб без счета, но, как говорилось, череп кривенький был, махонький). Составили они потом Матрицу Многостолбовую Малострочную, расчехлили Великую Машину Счетную, вложили в нее циферки полученные, да такой завели в ней Анализ Кластерный да Дискриминантный, что аж стон по земле пошел! И выдала им Машина картинку дивную да цветную притом. Глянули на нее Джей и Фенг и аж заколдобились. Да и было отчего! Ибо выходило из той картинки цветной две вещи: одна явная, а другая – тайная. Явная была такова, что YV0999 вовсе не схож с рыжим зверем-орангутаном индонезийским, что казалось очевидным мудрецам древности, кои в былые времена говаривали (а то и пописывали): “Ай до чего ж похож наш люфенгушка на предка орангутанушки! – вылитые дед да внучок!”, да длинные седые бороды при том поглаживали. А зато выходило из той картинки кластерной, что схож YV0999 с обезьяном-шимпанзе африканским. Зело чудно показалось это Джей и Фенгу, ибо обезьян-шимпанзе живет в далекой Африке, а до Царства Орангутанов от Юаньмоу – рукой подать.Впрочем, у картинки и оборотная сторона была, потаенная, – в средней части ее смешались в кучу (хочется ляпнуть “кони, люди”, но это совсем из другой сказки, про удивительную Вента-Мицену Гишпанскую) гориллы, шимпанзе и орангутаны. Так-то часто бывает, что самое дивное на виду лежит, но глаз мимо ходит да дивноты той не замечает. А сталось то оттого, что Алгоритм все выбранный решает в Анализе Кластерном, а всех тех Алгоритмов – тьма тьмущая да еще один с привесочком.Не только измерили Джей и Фенг черепа, но и описали таинственные дискретные признаки, тайное генетическое значение коих никому доселе неведомо, но всегда всех интригует безмерно. И вышла у них табличка предивная, ибо в ней-то почти все признаки, окромя двух с половиною, сошлись у YV0999 и зверя-орангутана, а не сошлись с обезьянами африканскими, что не в лад пошло с картинкою кластерной.Но не остановились на том Джей и Фенг, провели они еще и фаун сравнение. И обнаружили они, что фауны, в коих находили доселе люфенгпитеков, вовсе не схожи по составу своему с фаунами, в коих сивапитеки да хоратпитеки обнаружены. Хоть и мало о чем говорит сие, но как бы намек тут некий мудрецам приоткрылся.И задумались тогда крепко Джей и Фенг: кому ж родня тогда YV0999? И открылось им, что близок он к стволовой линии гоминид, кои, окромя горилл, и шимпанзей, и человеков в себя включают сродственно. Все ж никак не может стоять Поднебесная без того, чтобы и в ней стволовой Великий Предок не нашелся. А чтоб явнее та истина всем казалася, сравнили Джей и Фенг YV0999 с Rudapithecus из страшной, дикой и вечно холодной Восточной Европы (хоть и найден Rudapithecus в Европе Центральной, однако ж глобусы в Аризоне и Юньнани, видимо, малость перекошенные, али долго на боку лежали-сплющились, али карту еще Клавдий Птолемей рисовал – то нам неведомо). И, конечно, нашли они сходства необычайные, удивления достойные, в зубах заключающиеся. Впрочем, сравнение то было предварительное и целью имело интригу вызвать и зуд мозговой у приматологов.Смелы были Джей и Фенг: задумали они поразить прочих мудрецов смелою догадкою, что шли Люфенгпитеки до Китая не по берегу морскому, как примитивные косматые Pongo делали, а по пути Северному, через горы высокие Тибетские, через степи широкие Ордосские, через пески зыбучие Монгольские да до самых до лесов дремучих Южнокитайских. Долог и труден был путь Великого Предка, да на то он и Великий, чтоб все препоны преодолеть да окольною дорогою диких понгов обогнать. И неважно показалось Джей и Фенгу, что на Северном Пути о гоминоидах и слыхом не слыхивали, и нюхом не нюхивали: на то она и Догадка Ученая, чтоб затем в находках воплотиться и всех своею глубиною и прозорливостью поразить.А после сели Джей и Фенг за свои компьютеры многоклавишные да разноязыкие, да и написали статью о четырех страничках, да и опубликовали ее врагам на зависть, а нам на удивление. Тут и сказке конец, а кто прочел, тот молодец.
Сказ второй: о том, как лик Люфенгпитека взглянул на мудрецов и как Люфенгпитек Китай обособил
Велика страна Китай, бесчисленны ее жители, беспредельны рисовые поля, и недра ее бездонны. Редкий шахтер потревожит своей мотыгой лигниты Жоатонга. Миллионы лет лежат под бамбуковой сенью нетронутые богатства – бивни слонов-стегодонов, зубы зайцев-атилепусов, черепа обезьян-люфенгпитеков. Но даже в Поднебесной время неумолимо течет. Вот уж изобретены порох и компас, построена и уже развалилась Великая Стена, основана Вечная Империя и сметена Пламенной Революцией. Доходит черед и до древних костей. Но тяжек труд палеонтолога: прочны камни, трухлявы кости, жарко печет солнце Юньнани, катится пот по спине землекопа. Но трудолюбие китайцев не зря вошло в поговорку. И вот из земли Шуйтангба впервые за 6 млн лет (6,0–6,5) появляется невиданное лицо. Плоско оно, круглы его глазницы, широки его скулы, и радует это китайский взгляд. Счастливы И Супин, Ден Ченлун и Ю Юшан. Скорее чистят они треснувший лоб и крохотную челюсть, с удивлением стряхивают вековую пыль с молочных зубов. Кто перед ними? Неужели и это – предок китайцев? Лунгупо и Лунгудун, Нанкин и Чжоукоудянь, Дали и Цзиньнюшань, Мапа и Жижендун, Малудун и Лунлин – не счесть кандидатов на эту почетную должность.И вот уже созвана международная команда исследователей, трещат клавиши, гудят томографы, горят процессоры: кипит работа. Шесть-семь миллионов лет – важное время. В те времена в Африке ранние австралопитеки, робко озираясь, выходили из джунглей на простор саванн, в Европе доживали свой век последние дриопитеки, на средиземноморских островах гибли в зубах злых кошек беспечные ореопитеки, в Азии могучие гигантопитеки соревновались с пандами в пожирании бамбука, а по ветвям будущей Юньнани скакали неведомые обезьяны.Много придумано названий этим обезьянам, да трудно в них разобраться. Самые древние человекообразные Юньнани найдены около Кайюани и названы Lufengpithecus keiyuanensis (8,3–10,5 млн лет назад), более поздних из Юаньмоу прозвали Lufengpithecus hudienensis (7,1–8,2 млн лет назад), а самых последних из Шихуйба в Люфенге – Lufengpithecus lufengensis (6,2–6,9 млн лет назад). Но любят ученые люди латинский язык, а потому обогатились списки такими сказочными словосочетаниями, как Homo erectus wushanensis и Homo erectus zhupengensis, Sinopithecus xiaolongtanensis и Sinopithecus xiaoheiensis, Sivapithecus yunnanensis и Dianopithecus progressus, а также многими иными, дивными в своем благозвучии. А все потому, что обычно в руки исследователей попадают лишь зубов обломки да челюстей куски, и лишь мудрейший может разведать их истинную суть, да и то с трудом и не сразу. Бывали, конечно, и лучшие дни, найдены были и черепа люфенгпитеков, но не на радость, ибо так были сплющены и раздавлены толщами земляными, что даже китайские мастера доселе расправить лишь один смогли. Был, впрочем, и светлый луч во тьме прошедших эпох: милое лицо детеныша малого найдено было в незапамятном 1988 году, а спустя всего 24 года, в году 2012-м, описано как необычайно особенное, для всех гоминоидов изначальное.И вот ныне второй луч осветил тайны минувшего: второе лицо, тоже дитяти несмышленого и столь же милое. Антропологи китайские соединили усилия свои с антропологами американскими и описали череп сей (Ji et al., 2013). И ведь что дивно: нет в мире находок черепов детенышей человекообразных, а в Юньнани два уж таких. Новоявленный детеныш был отнесен к виду Lufengpithecus lufengensis. Хоть и мал тот детеныш, да пришлось его сравнивать с другими находками, ранее сделанными: с черепом детским первым и реконструированным взрослым. Да с обезьянами иными – орангутанами бородатыми. И вроде как похож люфенгпитек на орангутана, да вроде и не похож. Из чего вывод сделан был закономерный об особливой эволюции эндемичной в Южном Китае обезьян человекообразных, не имеющей ничего общего с путями орангов ли, горилл ли или шимпанзе каких. Ибо в Китае все вообще особенное и не может там быть банальностей заурядных и обычностей обыденных.Велика страна Китай, и недра ее бездонны. Много еще хранят они черепов, зубов и отпечатков. И многое еще узнаем мы о прошлом. А пока шелестит бамбук под дождем, панда листья жует, да рис колосится…
Сказ третий: о том, как безликий Люфенгпитек запутал мудрецов, о первом китайце и последнем люфенгпитеке
В далекие-предалекие времена (1985–1988 гг.) в туманных горах китайской Сычуани, покрытых бамбуковыми лесами и славных плюшевыми пандами, отважные китайские палеонтологи копали пещеру Лунгупо. Глубок был раскоп, многочисленны находки. Чего только не нашли доблестные ловцы знаний: и огромные мослы китайских мастодонтов, и крошечные китайских хомяков, и истлевшие мощи макак с проциноцефалами, и складчатые зубы гигантопитеков с лошадями. Все глубже рыли упорные искатели, и все больше чудес им открывалось. На пятом уровне наградой им был булыжник со сколом, на седьмом они наткнулись на верхний резец, а на восьмом ждала их сенсация – еще один обработанный булыжник да часть нижней челюсти с двумя зубами! Тут-то и разыгралась китайская антропология, да вместе с археологией. Ведь слои те были не простые, а очень древние, аж 1,96 млн лет датированные. Не было доселе видано таких древних зубов в Китае, да и во всей Азии. То есть зубов-то в Китае хватает, даже весьма славна Поднебесная драконьими зубами, которые исстари лекари перетирали на лекарства (страшно подумать, сколько уникальных окаменелостей сгинуло в их ступках!), однако же таких, чтобы как бы человеческими были, да двумя миллионами бы датировались, – это невиданно было и неслыханно. Потому-то и возликовали мудрецы китайские и назвали те находки Homo erectus wushanensis, а для непонятливых пояснили, что этот древнейший китаец был даже больше похож на H. habilis, только Конфуций учил скромными быть, отчего и назвали эректусом (Wanpo et al., 1995). И много после полицентристы торжествующе поминали его как первого человека в Китае, да и в Азии, да уж и в мире, чего уж мелочиться-то.Но не только в Китае мудрецы мудрят. Нашлись смутьяны и среди рыжих мохнатых варваров запада лаовай, кои все с ног на голову поставили: якобы коли H. erectus возник в Азии так уж слишком рано, то не мог он в сложении современного человека участвовать. Появились и такие знатоки, даобицзы “большеносые”, которые усомнились в том, что челюсть человечья (Schwartz et Tattersall, 1996). А потом и вовсе распоясались янгуйцы, “заморские черти”: взяли да написали статью, где по полочкам разложили, что резец, конечно, человечий, но даже слишком уж современный и запросто мог завалиться из вышележащего слоя, камни вообще ничем не похожи на орудия, а челюстью Лунгупо жевал не человек, а обезьяна вроде люфенгпитека (Etler et al., 2001).Сказка о двухмиллионолетнем китайце растворилась в туманах Сычуани, зато появилась новая интрига: выходит, реликтовые дриопитецины дожили до гораздо более поздних времен, чем доселе считалось. А дао-то все едино – что первый человек, что последний люфенгпитек, всяко Поднебесная в центре мира стоит, а находки в ней уникальнее, чем где-либо за ее пределами.
Понгины Ponginae (они же Simiinae) в современности представлены всего одним видом – орангутан Pongo pygmaeus.
Происхождение орангутанов изучено неожиданно хорошо. В качестве отдаленных предков предполагались сивапитеки и люфенгпитеки, но в последнее время были найдены, видимо, совсем прямые пращуры. В Индокитае обнаружены зубы и челюсти трех видов хоратпитеков: Khoratpithecus chiangmuanensis жил в Таиланде 10–13,5 млн лет назад, Kh. ayeyarwadyensis – в Бирме 8,8–10,4 млн лет назад, а Kh. piriyai – опять в Таиланде 7–9 млн лет назад. Ясно, что ареал был наверняка шире, но, как обычно, что и где найдено, то и найдено.
Хоратпитеки отличаются от орангутанов ровно настолько, насколько должны бы отличаться далекие предки. Интересно, что флора в местонахождении древнейшего хоратпитека обнаруживает явные африканские корни; теоретически приматы могли в это время перемещаться от Африки до Индокитая. К сожалению, у нас слишком недостаточно останков африканских гоминоидов этого времени.
Собственно род Pongo достаточно хорошо представлен в палеонтологической летописи. Найдено множество зубов и даже два скелета – целый взрослого и фрагментарный детеныша – во вьетнамской провинции Хоа-Бин (Bacon et Long, 2001). По останкам из разных мест возрастом от среднего плейстоцена до голоцена описаны как минимум шесть подвидов ископаемых орангутанов, в том числе из Вьетнама, Южного Китая, Бирмы и с Явы. Некоторые китайские находки были определены как самостоятельный вид P. fossilis, а вьетнамские – P. hooijeri. Большинство этих вариантов морфологически слабо отличаются от нынешних “лесных людей”, но были более крупными. Думается, уменьшение размеров от плейстоцена до современности явилось прямым следствием преследования людьми и вытеснения орангутанов в наихудшие закоулки джунглей. На Яве орангутаны дожили до вполне исторических времен, вероятно вплоть до XVII века.
Ныне сохранился лишь один вид. Иногда его, правда, делят на калимантанский P. pygmaeus и суматранский P. abelii виды, но разница между ними весьма несущественна: суматранский покрупнее, поярче, чаще ест фрукты, насекомых и прочих животных, больше времени проводит на деревьях и использует орудия труда. В литературе часто упоминается, что два вида просуществовали более чем 1,5 млн лет в изоляции друг от друга, но в реальности Ява с Калимантаном соединялись и намного позже, а расшифровка полного генома позволила оценить время дивергенции в 400 тыс. лет (Locke et al., 2011; впрочем, позже оно было рассчитано опять в 1 млн лет: Prado-Martinez et al., 2013). В некоторой степени разделение орангутанов на два вида обусловлено нуждами их сохранения: чем уникальнее популяция, тем легче убедить людей ее беречь. Любопытно – генетики утверждают, что генетическое разнообразие суматранских орангутанов больше, чем калимантанских, тогда как морфологи среди последних выделяют три подвида, а среди суматранских – ни одного.
Современные орангутаны – рыжие косматые длиннорукие обезьяны. Лицо орангутанов легко узнаваемо уплощенностью и даже вдавленностью, близко посаженными глазами, очень сильно выступающими челюстями. У взрослых самцов имеются незабываемые пупырчатые кожистые выросты вокруг лица и на груди, длинные усы, борода и выражение а-ля “восточный мудрец”. Уши такие маленькие, что их видно только у детенышей и иногда самок, самцы же кажутся совсем безухими. Орангутаны предпочитают жить на деревьях, но из-за большого веса вынуждены передвигаться очень неторопливо и осторожно. Длинные руки приспособлены к брахиации, но не быстрой, как у гиббонов, а медленной. Крупные самцы, вес которых в среднем 45–90 кг, но может достигать 190 кг, часто ходят по земле, опираясь при этом на согнутые фаланги пальцев, однако не совсем так, как это делают гориллы и шимпанзе. Самки заметно меньше – 40–45 кг, они почти все время проводят на деревьях с детенышами. Суматранские орангутаны реже спускаются на землю, так как там их ждут тигры; на Калимантане полосатые хищники тоже водились, но были выбиты людьми не то еще в плейстоцене, не то лет двести назад. Так или иначе, из крупных человекообразных орангутаны – самые древесные. Из-за этого их руки чрезвычайной длины, большой палец кисти крайне укорочен и почти нефункционален, зато пястные кости крайне вытянуты и изогнуты. Нога тоже изменена соответствующе: большой палец стопы почти полностью редуцирован – у 60 % орангутанов концевой фаланги и ногтя вообще нет, особенно в этом преуспевают самки, что наводит на мысль о сцепленном с полом наследовании признака (Tuttle et Rogers, 1966).
Орангутаны заслужили славу очень спокойных и мирных животных, но не так давно выяснилось, что на Суматре самки регулярно охотятся на медленных лори (Hardus et al., 2012), а на Калимантане детеныши и подростки иногда ловят сомиков в подсохших водоемах (Russon et al., 2014; кстати, какой подарок ученых сторонникам “гипотезы водной обезьяны”! Правда, те все равно не читают научных журналов…). Едят они и других животных, когда те подворачиваются под руку, но предпочитают все же дурианы – очень колючие, очень вонючие, но очень вкусные плоды. Орангутаны довольно часто используют орудия труда, хотя и самые простецкие – разного рода палки-ковырялки для добывания термитов, меда из пчелиных гнезд и очистки родственных дурианам плодов Neesia от шерстовидных волокон; широкими листьями они трогательно укрываются от дождя как зонтиками или используют их как прихватки, чтобы брать колючие ветки. Однако образ жизни орангутанов мало способствует развитию трудовых навыков, тут у них маловато перспектив.
Социальная структура современных орангутанов сильно нарушена, так как люди слишком много преследуют их и вытесняют в неудобные местности. Раньше, по-видимому, орангутаны были куда как более общительными существами, жизнь группами и обучение детенышей должны были существенно развивать их интеллект (Schaik, 2002). Однако в настоящее время на Калимантане они живут поодиночке – многому ли научишься в таких условиях? На Суматре их первоначальный образ жизни сохранился чуть лучше: самец контролирует большую территорию, которая включает участки нескольких самок с детенышами, хотя и тут пары образуются только на период размножения. Поэтому и здесь у деток возможности для развития не лучше калимантанских. Лишь в резерватах и зоопарках они могут проявить свои потенциальные дарования. Какая судьба ждет рыжих “лесных людей”?..
Кстати, о тайнах…
Часть ископаемых человекообразных пока не находит себе места в классификации. Kogolepithecus morotoensis из Уганды, живший 17,5 млн лет назад, известен по четырем зубам, Helladopithecus semierectus из Греции (17 млн лет назад) – по обломку бедренной кости, Langsonia liquidness из Вьетнама (475 тыс. лет назад) – по пяти зубам из единственной пещеры Там-Хуен. В отложениях формации Нгорора в Туген-Хиллс в Кении (12,4–12,5 млн лет назад) обнаружены зубы как минимум одной крупной (схожей с проконсулом – но поздновато для этого рода!) и двух мелких человекообразных обезьян, а также “не мартышкообразного узконосого” примата. Обломок нижней челюсти из Нигера с датировкой 5–11 млн лет назад – единственная находка человекообразного примата из Западной Африки. Фрагмент более всего похож на челюсть шимпанзе, но обнаружен далеко за пределами современного ареала шимпанзе и горилл, зато сравнительно недалеко от чадского местонахождения сахелянтропа. Samburupithecus kiptalami из Кении (8,5–9,5 млн лет назад) имел размер современной гориллы; он образует хронологический мостик от хорошо изученных ранне– и среднемиоценовых гоминоидов к современным. Еще один – пока безымянный – вид крупных человекообразных известен по единственному премоляру из Накали в Кении с датировкой 9,8–9,88 млн лет назад; там же найдены останки еще двух видов “мелких не мартышкообразных узконосых”. Исчезновение тропических лесов в Восточной и Южной Африке лишило нас находок; предки шимпанзе и горилл, судя по всему, жили где-то в Центральной Африке, но там их останки пока не обнаружены.
Человекообразные в черных тонах: африканские человекообразные – почти люди и люди
Подсемейство Gorillinae (оно же Anthropopithecinae или Anthropithecinae) практически никем не признается за валидное в современной систематике. Отличия горилл и шимпанзе от людей в масштабе всех приматов слишком незначительны, да к тому же гориллы отделились от шимпанзино-человеческой линии заметно раньше разделения шимпанзе и людей, а потому обычно всех африканских человекообразных объединяют в гоминид Hominidae и гоминин Homininae. Такой подход совершенно логичен филогенетически и узаконен таксономически. Но: если объединять шимпанзе с человеком в одном подсемействе или даже роде, то как подчеркнуть специфику человеческой линии? Будь бы еще только один вид современного человека, можно было бы обойтись безо всякого подчеркивания. Так ведь ископаемых двуногих предков очень много, они достаточно разные и все отличаются от шимпанзе. Можно бы, конечно, обойтись трибами и подтрибами, но зачем мелочиться? Может антрополог раз в жизни спокойно предаться антропоцентризму? Это всяко проще, чем городить странные конструкции типа “крупные африканские нечеловеческие человекообразные обезьяны” и “крупные африканские человеческие человекообразные обезьяны” – в разделении гориллин и гоминин смысл тот же, но воплощение более простое.
Происхождение горилл и шимпанзе покрыто мраком неизвестности. Впрочем, в последние годы в нем появились первые робкие лучики света: из Эфиопии был описан хорорапитек Chororapithecus abyssinicus, обитавший тут в конце миоцена 8 млн лет назад (Suwa et al., 2007), а из Кении – накалипитек Nakalipithecus nakayamai, живший 9,8–9,88 млн лет назад (Kunimatsu et al., 2007).
Хорорапитек Chororapithecus abyssinicus известен из местонахождения Бетиха по девяти зубам размером с горилльи и очень на горилльи похожим. Отличия заключаются почти только в более низких бугорках моляров и толстой эмали. Посему он из всех известных ископаемых приматов больше всех похож на предка гориллы. Впрочем, распределение толщины эмали у хорорапитека довольно специфическое, так что совсем прямым предком он, вероятно, все же не был. Да и географически хорорапитек расположился не слишком удачно для того, чтобы быть пращуром горилл, – Эфиопия не Уганда и не Конго. Палеоэкологические реконструкции указывают на обитание хорорапитека в прибрежных зарослях вокруг рек и озер, не так далеко от открытых саванн – окончание миоцена и обезлесение Африки были не за горами.
Накалипитек Nakalipithecus nakayamai известен по обломку нижней челюсти с тремя зубами и еще одиннадцати изолированным зубам. Не столь богато, как хотелось бы, но, по крайней мере, теперь исследователи точно знают, где лежат сокровища, так что новые открытия нам гарантированы. Накалипитек имел размеры примерно как у самок орангутанов и горилл. Его клыки весьма короткие, хотя и массивные, похожие на заостренные премоляры, а эмаль моляров толстая. Зубы накалипитека здорово похожи на зубы Ouranopithecus macedoniensis, горилл и шимпанзе, так что из всех прочих обезьян он идеально подходит на роль последнего общего предка современных африканских человекообразных и человека. Вот оно – очередное “достающее звено”, Великий Предок, столь долго искомый. И пусть пока в наших руках лишь горсть зубов, зато нам известно, где искать и что искать.
Существование в Туген-Хиллс, Бетихе и Накали нескольких видов мелких и крупных человекообразных в интервале от 12,5 до 9,8 млн лет назад ставит крест на гипотезе вымирания африканских человекообразных в конце миоцена и реконкисте их из Европы. А ведь в некоторый момент такая концепция была довольно хорошо разработана: согласно ей, средне-позднемиоценовые пертурбации климата сгубили гоминоидов Африки, а прямыми предками современных горилл, шимпанзе и человека являются европейские уранопитеки, грекопитеки и пиеролапитеки, вторично вернувшиеся на исходную прародину.
Действительно, картина достаточно странна: Европа времени около 10 млн лет назад оказывается едва ли не богаче человекообразными, чем Африка. Впрочем, такая ситуация – в большей степени итог недостатка подходящих местонахождений в Африке, плохой сохранности окаменелостей и их недостаточной изученности.
Но европейские “африканские” гоминоиды сами по себе интересны. Древнейший из них – пиеролапитек Pierolapithecus catalaunicus – жил в Испании 11,9 млн лет назад. От него сохранился почти целый скелет с черепом, хотя и основательно погрызенный каким-то хищником.
Череп пиеролапитека своеобразен: с низким, широким и чрезвычайно сильно выступающим лицом, умеренным надбровьем, низкими широкими глазницами, широким межглазничным пространством и очень широким носом, почти квадратным глубоким небом, огромными узкими клыками, широкими верхними центральными резцами. В этом комплексе сочетаются как примитивные, так и продвинутые черты: в анфас пиеролапитек похож на гориллу, а вот в профиль – скорее на сааданиуса или афропитека. В целом он вполне подходит на роль предка современных африканских человекообразных, только вот география подводит – Северная Испания все же не граничит с Африкой. Теоретически приматы могли бы перебираться через Гибралтар, но в реальности это трудновато, скорее уж они мигрировали вдоль берегов Средиземного моря через Ближний Восток.
Строение тела пиеролапитека наконец-то похоже на современных крупных человекообразных: грудная клетка широкая и уплощенная спереди назад, поясница укорочена и расширена; исчез сустав между локтевой костью и запястьем; таз имеет промежуточное строение между мартышкообразными и проконсулами с одной стороны и современными человекообразными – с другой, что примерно соответствует варианту гиббонов; коленная чашечка низкая, широкая и толстая (отличается от узкой и тонкой широконосых, мартышкообразных, Proconsul, Nacholapithecus и гиббонов, но аналогична надколеннику современных крупных человекообразных и человека). Впрочем, еще не все признаки пришли в современное состояние. Судя по строению фаланг, пиеролапитек ходил на четвереньках с опорой на ладонь, а не согнутые фаланги. Очевидно, пиеролапитеки совершили переход от четвероногого бегания по ветвям к вертикальному лазанию по деревьям, хотя еще не были способны подвешиваться на руках, как современные человекообразные.
Уголок занудства
Ладоне– и костяшкохождение отражается на разных костях скелета, сильнее всего, очевидно, на пястных и фалангах кисти. На головках пястных костей и основаниях проксимальных фаланг суставные поверхности у ладонеходящих приматов ориентированы дорзально и сглажены для облегчения отгибания пальцев вверх, а у костяшкоходящих – на головках пястных тоже дорзально, но с ограничителем разгибания в виде поперечного костного гребня, а на проксимальных фалангах – проксимально, то есть с торца. У ладонеходящих на пальмарной стороне основания проксимальной фаланги имеются два мощных бугорка для укрепления крупных сесамоидов, ограничивающих канал для сухожилия длинного сгибателя, весьма напрягающегося при чрезмерном разгибании пальца, а у костяшкоходящих этих бугорков нет. Головки проксимальных фаланг у ладонеходящих вытянуты спереди назад, а у костяшкоходящих – дорзо-пальмарно, а сама фаланга из-за этого сильнее изогнута, чтобы следующая – медиальная – оказалась опорой, лежащей дорзальной поверхностью горизонтально на земле.
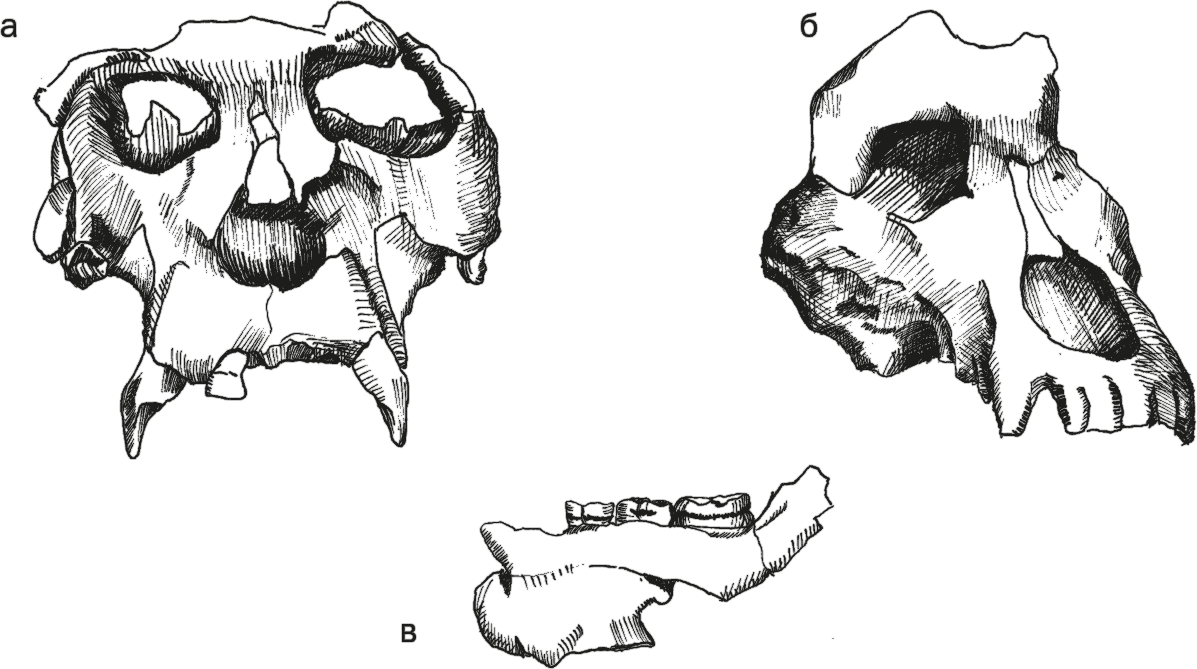
Рис. 40. Черепа Pierolapithecus catalaunicus (а), Ouranopithecus macedoniensis (б) и челюсть Nakalipithecus nakayamai (в).
Пиеролапитеки жили в теплых влажных лесах и питались преимущественно фруктами.
Любопытно, что пиеролапитеки обитали одновременно и в тех же местах, что и дриопитеки. Посему некоторые палеоприматологи не желают пускать их в тесный кружок африканской родни и прямо относят испанских “недогорилл” к Dryopithecus fontani или, более осторожно, к трибе Dryopithecini. Однако отличия пиеролапитеков от дриопитеков все же достаточно заметны, а прогрессивные черты и сходство с современными человекообразными несомненны.
На противоположном конце Европы и в Малой Азии жили родственные уранопитеки и грекопитеки. В Греции найдены череп и несколько нижних челюстей уранопитека Ouranopithecus macedoniensis (9,2–9,6 млн лет назад), а также одна описанная как Graecopithecus freybergi (6,6–8 или, менее вероятно, 9–9,5 млн лет назад). Трудно понять, представляют ли уранопитек и грекопитек один вид или два рода. Сложность в том, что грекопитек описан по плохо сохранившейся нижней челюсти с разрушенными зубами. Все же между ними есть некоторые отличия, тем более они с большой вероятностью жили в разное время.
Уранопитеки изучены гораздо лучше. Они были крупнее всех прочих европейских приматов – размером с самку гориллы, около 50–70 кг. Череп греческого уранопитека отличается от черепа прочих человекообразных очень низким расположением и массивностью скуловых костей, отчего лицо приобретает крайне тяжелый вид. Лицо несколько вдавлено. Верхняя челюсть умеренно выступает вперед, примерно как у дриопитеков и горилл – сильнее, чем у проконсулид и гиббонов, но слабее, нежели у орангутанов, шимпанзе и даже австралопитеков. Глазницы почти прямоугольные, межглазничное расстояние и носовое отверстие широкие, небо глубокое. Верхние резцы очень широкие, клыки толстые и не слишком длинные, моляры большие и имеют крайне толстую эмаль, что предполагает питание какой-то очень твердой пищей.
Не так давно был описан особый вид турецкого уранопитека Ouranopithecus turkae, жившего позже греческого, 7,4–8,7 млн лет назад. Турецкий отличается бóльшими размерами, менее выступающей челюстью и более короткими клыками.
Уранопитеки жили в более открытых местообитаниях, чем большинство человекообразных, так как в это время в Европе леса постепенно исчезали и сменялись саваннами. Некоторые исследователи считают, что уранопитеки могли быть предками австралопитеков, однако у тех и других есть свои специализации, не позволяющие им выстроиться в стройный ряд “отцы и дети”. Все же некоторый параллелизм, особенно с Australopithecus africanus, не может не радовать. Если бы не оледенения, окончательно сгубившие европейскую тропическую флору и фауну, как знать – возможно, человечество появилось бы из уранопитеков?..
Последний из обезьян
В наши дни Европа не может похвастаться разнообразием приматов. Один вид макак на одной скале в Гибралтаре, еще один двуногий безволосый (правда, он – повсюду) – вот, собственно, и всё. Обезьяны в зоопарках, конечно, не в счет. Но так было не всегда. Когда-то Европу населяли довольно многочисленные приматы. В эоцене и олигоцене тут резвились адаписовые и омомисовые полуобезьяны, в миоцене – плиопитеки, мезопитеки, ореопитеки, анойяпитеки, дриопитеки, уранопитеки, грекопитеки и пиеролапитеки, в плейстоцене – макаки и теропитеки. Но климат становился все холоднее, и обезьяны исчезали…Когда же Европа лишилась последних представителей человекообразных? В первом номере Journal of Human Evolution 2012 года международная группа исследователей опубликовала описание находки, которая может пролить свет на этот вопрос (Spassov et al., 2012). В Болгарии в карьере Азмака около Чирпана были найдены остатки ископаемой фауны, включающей позднемиоценовых представителей слонов, саблезубых тигров, газелей, примитивных носорогов, гиппарионов и других животных. Сочетание видов млекопитающих позволило оценить время отложения слоев в 7 млн лет.Среди костей обнаружен и один зуб – левый второй верхний премоляр – человекообразной обезьяны. Зуб целый, хотя и очень стертый. Морфологию его жевательной поверхности оценить практически невозможно, однако форма и размеры коронки оказались в наибольшей степени соответствующими Ouranopithecus macedoniensis, а малые размеры корней могут свидетельствовать о родстве с Graecopithecus freybergi. Эти два близких вида гоминоидов жили в Греции в позднем миоцене, около 8,7–9,6 млн лет назад. В то же время болгарский зуб отличается от грузинского удабнопитека и турецкого уранопитека, имеющих примерно те же датировки, что у греческих обезьян. Очевидно, виды гоминоидов закономерно распределялись географически: в Европе – на запад от Черного моря – жили одни обезьяны, а в Азии – на юг и восток – другие.Видимо, зуб из Болгарии принадлежал одной из последних человекообразных обезьян Европы. Примерно в то же время – чуть раньше или чуть позже – вымерли и ореопитеки в Италии. Их сгубило то же похолодание, что раздвинуло саванны в Африке и выгнало на их просторы предков австралопитеков. Европейские гоминоиды не смогли или не успели адаптироваться к новым условиям. Северное расположение не оставило им шансов на развитие.Человекообразные исчезли из Европы, и на долгих 6 млн лет она осталась вотчиной только мартышкообразных. Чуть более 1 млн лет назад человекообразные в лице человека взяли реванш, но это уже совсем другая история…
Итак, после всех вымираний крупные африканские человекообразные представлены ныне двумя родами с тремя видами. Гориллы Gorilla gorilla населяют Западную и Центральную Африку (иногда горная горилла выделяется в особый вид Gorilla beringei, но это, как и в случае с орангутанами, аргументировано лишь генетически и обусловлено в немалой степени нуждами охраны и спасения горилл). Шимпанзе делятся на два вида: обыкновенные Pan troglodytes живут также в Западной и Центральной Африке севернее от Конго, а карликовые, или бонобо, Pan paniscus – южнее этой реки в Заире.
Гориллы – самые крупные современные обезьяны. Конечно, измерять приматов можно по-разному. Рост горилл не такой уж запредельный, как многим может показаться, – в среднем у самцов 1,65–1,75, а у самок так и вовсе 1,4–1,5 м – ниже большинства европейцев. Самый высокий горилл был ростом 1,94 м – всяко меньше большинства баскетболистов. Другое дело – вес. Тут гориллам действительно нет равных: в среднем самки весят 60–100 кг, а самцы – 140–200 кг, с рекордом в природе 266 кг, а в неволе так и вовсе до 388 кг! Благодаря огромнейшим мышцам и здоровенной морде гориллы человек рядом с гориллой выглядит маленьким.
Гориллы настолько могучи, что не имеют в природе врагов, кроме людей. Лишь изредка леопард может напасть на детеныша. Излишняя безопасность привела к уменьшению стимулов к эволюции и развитию интеллекта, хотя потенциально гориллы крайне умны, что наглядно демонстрируют в экспериментах.
Череп горилл легко узнаваем по огромным общим размерам и мощнейшему развитию рельефа – надбровного валика, сагиттального и затылочного гребней. Богатырские жевательные мышцы сходятся у них на макушке, так что, когда мы смотрим на здоровенную башку гориллы, мы на самом деле видим немало шерсти, большие мышцы, толстый череп и не так уж много мозгов (в отличие от человека, размер мозга которого не слишком замаскирован всем, что покрывает его). Впрочем, “не так уж много мозгов” гориллы – это только в сравнении с нами, шибко умными, а на самом деле все равно рекорд для приматов: 535 см³ в среднем у самцов, с индивидуальным рекордом 752 см³. У горилл могучая нижняя челюсть, иногда довольно сильно выступающая вперед, для обозначения чего даже предложен специальный термин “нижнечелюстной прогнатизм”, впрочем неупотребительный.
Огромный вес делает горилл почти полностью наземными животными. Однако вся их морфология вопиет, что спуск на землю произошел эволюционно совсем недавно; строго говоря, неизвестно когда, но достаточно уверенно можно предположить, что позже появления австралопитеков. По земле гориллы передвигаются, как и шимпанзе, на четвереньках с опорой на согнутые фаланги пальцев. Из-за этого для противостояния тяжести туши головки пястных костей и проксимальные фаланги кисти резко расширены, гороховидная кость огромная, сложно устроенная, укрепляющая запястье. Тыльные стороны средней и концевой фаланг пальцев кисти не только голые, но и имеют пупырчатую поверхность – почти папиллярные узоры.
Как и у орангутанов, половой диморфизм – различие самцов и самок – у горилл выражен очень резко: самцы почти вдвое крупнее. Старые доминантные самцы отличаются серебристым цветом спины, они пасут гарем из нескольких самок с детенышами, а также молодых самцов. Когда эти молодые самцы подрастают, они образуют свою группу, уводя самок у патриархов.
Шимпанзе – самые близкие наши родичи.
Шимпанзе в ископаемом виде представлены чрезвычайно скудно: найдены лишь три зуба в кенийской местности Туген-Хиллс с датировкой около 545 тыс. лет назад (McBrearty et Jablonski, 2005). Они ничем принципиально не отличаются от современных и ничем не могут помочь в решении вопроса о происхождении этого рода. Однако сопоставление данных генетики и палеонтологии позволяет оценить время расхождения шимпанзиной и человеческой линий в 7–11 млн лет, причем большинство ученых склоняется к наименьшей цифре.
Обыкновенные шимпанзе отличаются от карликовых шимпанзе бонобо размерами и поведением. Обыкновенные имеют рост около 1,4 м у самок и 1,5 м у самцов и вес 40 и 50 кг соответственно, хотя отборные особи могут достигать и 80 кг. Бонобо гораздо мельче и грацильнее: 1,2 м ростом и весом самок и самцов 33 и 43 кг, максимум – до 60 кг. Морда и клыки обыкновенных гораздо внушительнее, лицо обычно желтое, а уши безволосы и лопоухи, у бонобо же лицо более интеллигентное и черное, а уши сравнительно маленькие и пушистые. У обыкновенного шимпанзе объем мозга в среднем 394 см³, у бонобо – 349 см³ (вот она, магия чисел!).
Примечательно, что различия обыкновенных шимпанзе и бонобо по размерам челюстей и зубов, а также по уровню полового диморфизма вызваны не какой-то спецификой питания, а разницей в уровнях межсамцовой и внутригрупповой агрессии и возбудимости. Бонобо гораздо спокойнее, более уравновешенны и дружелюбны. Кстати, они довольно часто в природе ходят на двух ногах, причем иногда не для дела, а просто потому что им так нравится. Бонобо регулярно переносят в руках разнообразные предметы или детенышей.
Группы шимпанзе владеют определенными участками леса, однако состав самой группы не так стабилен, как у горилл. Отдельные особи – особенно самки – могут достаточно свободно переходить из стада в стадо. Внутри группы имеется иерархия, но она далеко не столь жесткая, как у павианов. Каждый индивид может завоевать авторитет сородичей не только силой и агрессией, но благодаря уму и сообразительности. Широко известен случай, когда самец шимпанзе, стоявший на низшей ступени социальной лестницы, добился места вожака, догадавшись напугать соплеменников громыханием пустых канистр из-под керосина, украденных у исследователей.
Кстати, о гнездах…
Шимпанзе каждый вечер, укладываясь спать, делают себе гнездо, обычно на дереве. Это не слишком масштабное сооружение из заверченных веток, мало спасающее от чего-либо, но дающее ощущение комфорта и уюта. Однако показательно, что шимпанзе никогда не пользуются гнездами дважды. Кочевая жизнь приводит их каждый вечер на новое место, хотя перемещения и происходят в границах определенной территории. Думается, австралопитеки вели себя примерно так же. В этом смысле изобретение людьми дома, постоянного места жительства и уж подавно прописки с регистрацией – странная новация, вызванная ненормальной численностью. Пока было куда брести, а границы, таможни, визы и паспортный контроль не превратили планету в тюрьму строгого режима, гоминиды перемещались сколько хотели. Человек – прирожденный путешественник, страннические наклонности прошиты в его обезьяньей сути как минимум с палеоцена. Потому-то людям так нравится открывать что-то, даже если они знают, что это уже прекрасно известно другим. Скажем, в большом городе открывается новая станция метро – многие люди едут “открывать” ее и потом рассказывают об этом знакомым-родственникам так, как будто на станции до них не ступала нога человека и вообще они съездили на доселе неведомый континент. Нам интересно бывать в новых местах, хотя и про родную территорию мы, конечно, не забываем. Так в людях уживаются сбалансированные патриотизм-территориальность с первооткрывательством-бродяжничеством.
Шимпанзе во многих деталях поведения похожи на человека. И чем дольше изучают их этологи, тем больше выявляется это сходство. Шимпанзе иногда кооперируются для охоты на мелких животных. Они используют палочки для выуживания термитов из ходов термитников. При этом в некоторых группах принято специально обкусывать такие палочки. Иногда термиты выкапываются с помощью палки, используемой как лопата, даже с упором ногой в торчащие вбок ветки. Другие шимпанзе раскалывают орехи камнями. Примечательно, что камни для этого специально никогда не обрабатываются, но подыскиваются определенной формы. Поскольку в тропическом лесу подходящие булыжники могут быть в дефиците, шимпанзе иногда хранят их и, возвращаясь на место после отсутствия, отыскивают старые орудия, а не находят новые. Третьи шимпанзе изготовляют из широких листьев “кружки” для воды или делают из жеваных листьев губки для собирания воды из пней. Недавно описано изготовление шимпанзе “копий”, заостренных с помощью зубов и используемых для охоты на галаго. Шимпанзе могут отлично жить и без орудий – многие и живут, но с ними жизнь облегчается и разнообразится. Существуют особые традиции использования орудий, свойственные отдельным популяциям обезьян. Традиции существуют и в области общения. Например, отличаются способы приветствия. Есть даже группы шимпанзе, в которых принято “рукопожатие” – касание рукой к руке. Пристальный взгляд и оскал у обезьян, как и у человека, означают угрозу, а ласковое прикосновение и поглаживание – дружелюбие. Они могут дружить и враждовать, любить и ненавидеть, обманывать и ругаться. Наконец, они смеются, у них есть юмор.
Интеллект шимпанзе, по мнению тех, кто с ними работает, приблизительно соответствует таковому трех-четырехлетнего ребенка. Впрочем, нельзя и переоценивать способности обезьян. Так, настоящей орудийной деятельности у них не существует: преобладает деструктивное поведение, а конструктивное находится лишь в зачаточном состоянии. Шимпанзе неспособны долго и целенаправленно обучаться, передача культурных навыков и закрепление нововведений происходит во многом случайно, часто полезные знания утрачиваются. Уровень кооперации весьма низок – шимпанзе не так часто делают что-то сообща, с единой целью.
Минутка фантазии
В 2004 г. новостные агентства и даже научные журналы всколыхнула новость о том, что на севере Конго, в лесу около города Били, исследователи наблюдали неких странных обезьян – двухметрового роста, с черными лицами и огромными сагиттальными гребнями, ходящих на двух ногах, живущих в болотистой местности, строящих гнезда на земле, как гориллы, но питающихся фруктами подобно шимпанзе, убегающих при приближении людей, оставляя фекалии в три раза больше шимпанзиных и огромные следы, превосходящие горилльи. Местные жители добавили к этому, что обезьяны бывают двух типов: тех, что лазают по деревьям, можно убивать отравленными стрелами, а вот тех, что побольше и живут на земле, отравленные стрелы не берут, зато сами эти чудища убивают даже львов. Моментально появились предположения, что это не то новый неведомый вид обезьян, не то гибриды шимпанзе и горилл. Появилось и красивое название: “обезьяны Бондо”. Какая сенсация!Однако специальные экспедиции, отправившиеся в лес Били, показали, что все не так уж чудесно. Никаких следов горилл тут не нашлось, зато встретились шимпанзе самого обычного подвида Pan troglodytes schweinfurthii, хотя действительно весьма крупные и регулярно устраивающие гнезда на земле. Открытие нового вида и даже подвида не состоялось, зато исследователи сделали массу новых наблюдений за поведением наших ближайших родственников.Африка вообще полна тайн. С XIX века до наших дней появляются упорные слухи о существовании гибридов между шимпанзе и гориллами, обычно их называют коолокамба, иногда соко, коула-нгуйя, н'гуи-мун или чего. Действительно описано довольно много живых особей и черепов с промежуточными признаками. Показательно, что таковые происходят только из областей, где водятся и гориллы, и шимпанзе. Причем подавляющая часть таких особей – самки, что наводит на мысли о неравной выживаемости гибридов, ведь замечено, что при отдаленной межвидовой гибридизации млекопитающих самки оказываются жизнеспособнее самцов. Теоретически в метисации шимпанзе и горилл нет чего-то невероятного, тем более что неоднократно отмечались смешанные группы двух видов, пасущихся вместе и даже играющих друг с другом. Одна беда: стопроцентных доказательств гибридизации за полтора столетия так и не было получено. Основная часть сообщений о коолокамба публиковалась в XIX и начале XX века, когда африканские приматы были вообще слабо изучены. Сейчас же, когда обезьян становится все меньше, а исследователи уже не стреляют их почем зря, вероятность наткнуться на настоящего гибрида стремительно падает. Возможно, вопрос решат ДНК-анализы из старых образцов, хотя пока такие попытки положительных результатов не дали.Коли уж зашла речь о метисации, никак нельзя забыть опыты Ильи Ивановича Иванова – выдающегося отечественного биолога. В 1926 г. он пытался скрестить шимпанзе и человека, но безуспешно. Аналогичные эксперименты, кстати, планировались и в Италии, но там их запретил папа римский. По этому поводу существует множество домыслов и баек, но действительность банальна: гибриды не получаются. Очевидно, главной виной всему разница в количестве хромосом – у шимпанзе на одну пару больше. Правда, разным подвидам домовой мускусной землеройки Suncus murinus не мешает даже разница в пять пар хромосом (от 15 до 20), а бабочкам горошковым беляночкам Leptidea sinapis – и вовсе в пятьдесят (от 56 до 106)! Есть и другие примеры видов с разным числом хромосом. Да оно и понятно: если бы различие в числе хромосом было совершенно непреодолимым барьером, то у всех живых существ с половым размножением было бы одно и то же их количество – как у первопредка. Но человек отличается от шимпанзе не только числом хромосом, но и их строением. И. И. Иванов не мог знать этого в 1926 г., но без его работ червь сомнения – может ли быть метис? – поныне точил бы нас. Впрочем, он и так точит некоторых недоверчивых граждан (“ученые скрывают!!!”)…
Современные шимпанзе могут служить лишь общей моделью наиболее ранних ступеней эволюции предков человека. Этот уровень примерно соответствует выходу предков человека из тропических лесов в саванны и возникновению прямохождения. Кстати, есть группы шимпанзе, обитающие на границе леса и саванны. Характерно, что их поведение значительно отличается от поведения полностью лесных шимпанзе. Нельзя, однако, забывать, что и сами человекообразные обезьяны прошли длинный эволюционный путь: современные шимпанзе – это не копия наших предков и уж тем более не сами эти предки. Так, у них развилось костяшкохождение и огромные клыки, видимо несвойственные общему с человеком предку.
А что же наши предки? Где были они, как выглядели, что делали? Итак, люди…
Назад: Глава 42 Обезьяны
Дальше: Вкладка

