Книга: Достающее звено. Книга 1. Обезьяны и все-все-все
Назад: Глава 40 Дальняя родня: тупайи и шерстокрылы
Дальше: Глава 42 Обезьяны
Глава 41
Примитивнейшие приматы
Корни человечества обычно ищут среди человекообразных обезьян, человеческую линию отсчитывают от возникновения прямохождения. Но, собственно, почему? Ведь приматы тоже имеют своих предков. Они прошли большой и сложный эволюционный путь, а непосредственными предками человека могли быть разные обезьяны. Поэтому необходимо вкратце рассмотреть разнообразие и историю возникновения отряда – его систематику и филогению.
Приматы (отряд Primates) в большинстве современных таксономических схем делятся на полуобезьян и настоящих обезьян. Некоторые группы – долгопяты и многие ископаемые формы – занимают промежуточное положение между этими большими подразделениями.
Заря: поступь Древнейшего и Великого Предка приматов
Первое известное приматоподобное млекопитающее – пургаториус Purgatorius. Это был небольшой, размером с мышь, зверек, живший на деревьях и питавшийся насекомыми. Пургаториус является наиболее вероятным прямым пращуром всех приматов, хотя его систематическое положение пока неопределенно – не так много от него осталось.
Эти крошечные зверюшки жили на самой границе мелового и палеогенового периода 65 млн лет назад на территории нынешних Канады и США. Один зуб был даже найден в меловых отложениях, содержащих кости динозавров. Зубы пургаториуса крайне примитивны, но некоторые “приматные” черты в них угадываются. Однако до недавнего времени было неясно, какой образ жизни вели эти первопредки. Более поздние плезиадапиформы были довольно разнообразны: часть из них была древесной, но некоторые вроде бы жили на земле, в лесной подстилке. А ведь древесность – одна из важнейших отличительных черт приматов как отряда! Когда же появилась эта особенность?
Новейшие находки, сделанные в США, проливают свет на первые шаги прародителей приматов. В Монтане были найдены кости ноги – таранные и пяточные – в отложениях древностью 65 млн лет (Chester et al., 2012, 2015; Clemens et Wilson, 2012). Они оказались приспособлены к активным поворотным движениям, типичным для древесных существ; детали строения указывают на принадлежность владельца к плезиадапиформам и родство с приматами. А ведь в это переходное от мела к палеогену время нам известен лишь один род приматоподобных существ – пургаториус. Именно поэтому исследователи предположили, что новонайденные плюсневые кости и обнаруженные ранее зубы принадлежали одному виду существ.
Пургаториуса трудно назвать приматом – слишком уж он примитивен даже для полуобезьян. Тем более интересно, что древесный образ жизни, определивший облик и эволюционную судьбу всех приматов, появился задолго до них самих.
Попытки стать приматом – плезиадапиформы
Плезиадапиформы (отряд Plesiadapiformes или подотряд Plesiadapiformes отряда Primates) известны из палеоцена и эоцена Европы, Северной Америки и Азии (65–42 млн лет назад). Обликом и образом жизни они были во многом похожи на грызунов. Размеры с мышь или крысу, вытянутая мордочка, маленький и просто устроенный мозг, глаза, расположенные по бокам головы, короткие массивные лапы, длинный хвост – все это общие признаки всех некрупных млекопитающих. Плезиадапиформы, с одной стороны, в общем плане строения имеют много от приматов, с другой – отличаются рядом параметров. Например, плезиадапиформы имели когти на всех пальцах, по крайней мере у части видов отсутствовало окостенение слуховой капсулы, многие обладали специализациями в зубной системе. Благодаря отпечаткам из французского местонахождения Мена известно, что плезиадаписы владели пушистым хвостом – не слишком характерным для приматов.
При этом из палеоцена и эоцена неизвестны другие млекопитающие, которые могли бы быть предками приматов. Посему включение плезиадапиформов в приматов или исключение из этого отряда – вопрос совести, а не таксономии. В настоящее время их все же чаще выделяют в самостоятельный отряд Plesiadapiformes, но близость к приматам подчеркивается объединением тех и других в рамках надотрядной группировки Primatomorpha.
Несмотря на древность, известно весьма много разных плезиадаписовых, в основном по зубам, но есть находки и целых скелетов. Замечательно, что даже на ранних этапах эволюции они были чрезвычайно многообразны. Это тем более так, поскольку мы знаем лишь о малой доле из общего числа столь древних животных. Густой букет родов и видов, возникший чуть ли не мгновенно из единого предка, называется первой радиацией приматоморфов.

Рис. 26. Схема эволюции приматов
Относительно типа передвижения и образа жизни плезиадапиформов велись споры. Одни считали их наземными (Gingerich, 1976, 1984), другие – древесными (Szalay et Delson, 1979), однако ж отсутствие срастаний костей, гибкость конечностей и пропорции кисти однозначно свидетельствуют в пользу второго варианта. Впрочем, древесность древесности рознь: плезиадапиформы не умели бегать и скакать по веткам так же резво, как делают это сейчас обезьяны. Может, потому они и приобретали специализации зубов, подобные грызуньим, – при пониженной мобильности надо извлекать из доступных ресурсов максимум выгоды? Неспроста зубная система, аналогичная системе многобугорчатых – с резко выступающими резцами, крупным режущим премоляром и перетирающими или давящими молярами, – возникала неоднократно среди примитивных приматоподобных существ: у саксонеллид Saxonellidae, карполестоидов Carpolestoidea и фенаколемуриновых Phenacolemurinae. Наблюдая, сколь неряшливо едят обезьяны – без конца роняя что-то, выедая лишь самые вкусные кусочки и оставляя массу объедков, – можно задуматься. Обезьяны всегда могут быстро переместиться и найти новые вкусняшки, а медленные плезиадапиформы, грызуны и многобугорчатые вынуждены довольствоваться тем, что есть на месте, и грызть до победы.
Уголок занудства
Тип питания определяется по форме бугорков на молярах: высокие и острые типичны для насекомоядных животных, низкие и тупые – для растительноядных. Можно посчитать длину режущих гребешков эмали относительно длины самого зуба: у насекомоядных и листоядных гребешки будут относительно длинные (ведь надо пережевывать хитин или целлюлозу), у фруктоядных намного меньше, а у любителей нектара и древесных смол – совсем короткие. У насекомоядных эмаль обычно более гладкая, а у растительноядных – морщинистая.Еще точнее можно определять диету по микростертости эмали: тут учитывается регулярность или беспорядочность царапин, их частота, направление, глубина и форма краев, микросколы, видные под электронным микроскопом (например: Calandra et al., 2012; Scott et al., 2012). Для анализа этих особенностей используются фракталы – вот зачем будущим биологам могут понадобиться уроки геометрии в школе!У листоядов обычно очень крупные челюсти, так как листья малопитательны, их надо есть очень много; на углу нижней челюсти листоядов обычно развит большой дополнительный отросток.
Несмотря на внешнее сходство, зубная система специализированных плезиадапиформов не была идентична таковой грызунов. У представителей семейства плезиадапид Plesiadapidae верхние резцы имели бугорки и не были самозатачивающимися. У многих полностью исчезли нижние клыки и вторые нижние резцы, а у некоторых и верхние клыки, так что между резцами и премолярами имелся большой зазор – диастема. От ранних форм к поздним доля мягкой пищи – фруктов и особенно листьев – в рационе, вероятно, увеличивалась (Boyer et al., 2010a). Плезиадапиды были одними из самых успешных животных своего времени, в некоторых местонахождениях они встречаются чаще всех остальных млекопитающих. Это отразилось и в распространении: в Европе, Северной Америке, Пакистане и Китае с нижнего палеоцена по нижний эоцен. Размеры их сильно варьировали – самые мелкие были как белка, а некоторые были довольно крупными – размером с крупную кошку.
Не слишком родственны, но экологически схожи с плезиадапидами микросиопиды Microsyopidae, жившие с верхнего палеоцена по средний эоцен в Северной Америке; среди них встречались животные размером с бобра. Относительно микросиопидов, впрочем, есть сомнения: некоторые специалисты считают их грызунами, насекомоядными или группой в отряде шерстокрылов.
Паромомиоиды Paromomyoidea, включающие паромомиид Paromomyidae, одними исследователями считаются шерстокрылами, а другими – плезиадапиформами. В частности, род Ignacius на основании строения зубов относится к фенаколемуринам Phenacolemurinae – подгруппе паромомиид, типовой род которых Phenacolemur имел пальцы, почти неотличимые от пальцев современного шерстокрыла (Beard, 1990, 1993). Да и у самого Ignacius форма костей кисти тоже весьма шерстокрылья (Hamrick et al., 1999; Krause, 1991). Посему, пока были известны только изолированные кости конечностей, предполагалось, что эти существа имели летательную перепонку и были способны к планирующему полету. Однако обнаружение целых скелетов показало, что эти звери были больше похожи на белок, а не на летяг. Пропорции кисти к туловищу, строение основания черепа и внутреннего уха у Ignacius скорее как у современных обезьян (Bloch et Silcox, 2001; Bloch et al., 2007; Silcox, 2003). Такая мозаика признаков, запутывающая общую картину, типична для древнейших приматов и близких им групп. Паромомиоиды имели квадратные уплощенные моляры, приспособленные для перетирания фруктов, впрочем, в диете немалую роль могли играть древесный сок и насекомые. В последовательном ряду палеоценовых родов наблюдается быстрое удлинение и истончение нижних резцов с появлением промежутка – диастемы – между резцами и премолярами.
Карполестоиды Carpolestoidea почти полностью перешли на питание волокнистыми плодами, орехами и стеблями, благодаря чему приобрели плоские и высокие премоляры с пильчатым краем. У них имелись и продвинутые приматные черты: например, большой палец ноги был снабжен ногтем, а не когтем, что подразумевает и бóльшую чувствительность; впрочем, и карполестоиды были медлительными животными и не могли быстро прыгать по ветвям. Они существовали не так уж долго – в пределах среднего и верхнего палеоцена Северной Америки, а последние представители доживали свой век в нижнеэоценовых лесах Китая и среднеоэценовых – Пакистана. Карполестоиды никогда не были многочисленны, и исчезновение их может быть связано с климатическими колебаниями или конкуренцией с более крупными животными, поскольку карполестоиды были одними из самых небольших плезиадапиформов – размером с мышь или крысу.
Пикродонтиды Picrodontidae, с их морщинистыми многобугорковыми, очень широкими – особенно первыми – и совсем не стертыми молярами, стали очень специализированными фруктоядами, а может, даже питались преимущественно нектаром, пыльцой и древесным соком. Череп Zanycteris резко сужался спереди, этим пикродонтиды были похожи на современных мышиных лемуров Cheirogaleinae и хоботноголовых кускусов Tarsipedidae, питающихся нектаром. Пикродонтиды, видимо, дальше всех из плезиадапиформов ушли от исходной насекомоядности. Забавно, что зубы пикродонтид столь необычайно похожи на зубы летучих мышей, что в 1935 году Дж. Симпсон даже предполагал, что пикродонтиды относятся к рукокрылым (Simpson, 1935).
Все эти специализированные плезиадапиформы, конечно, не были предками приматов, но известны и более генерализованные их варианты, например средне-верхнепалеоценовые североамериканские палехтониды Palaechthonidae. У них сохранялась исходная для приматов зубная формула, не было диастемы между резцами и премолярами, отсутствовали гипертрофии каких-либо зубов. Строение черепа Palaechton nacimienti было самым примитивным среди всех плезиадапиформов: глазницы маленькие, ориентированы вбок и широко расставлены, заглазничное сужение не выражено, обонятельные луковицы большие; судя по увеличенному подглазничному отверстию, имелись длинные вибриссы. Ориентировались палехтониды в основном с помощью осязания, слуха и обоняния, а не зрения. Судя по всему, вели они в основном наземный образ жизни. В немалой степени все эти черты были связаны с малыми размерами тела и насекомоядностью палехтонид (Kay et Cartmill, 1977). Показательно, что группа палехтонид – вовсе не древнейшая среди плезиадапиформов; с одной стороны, она сохраняла исходную насекомоядность и экологически схожа с тупайями, с другой – по времени близка к настоящим приматам границы палеоцена и эоцена. Вряд ли настоящих приматов дали палехтониды с их особенностями, но истинными предками должны были быть очень похожие на них существа, только более древесные и всеядные.

Рис. 27. Черепа Plesiadapis (а) и Palaechton nacimienti (б).
Насекомоядность вполне сочеталась с древесностью у микромомиид Micromomyidae, которые были одними из самых мелких плезиадапиформов – весом 20–30 г (самыми лилипутскими были представители подсемейства тинимомиин Tinimomyinae), вдвое меньше современных мышиных лемуров. При их размере другая диета, нежели насекомые, млекопитающим резко противопоказана. Эти крошечные зверюшки жили в конце палеоцена и начале эоцена в Северной Америке.
Впрочем, и размеры микромомиид не предельны: поздние пикромомииды Picromomyidae побили все рекорды – Picromomys petersonorum весил ничтожные 10 г. Эти животные известны из среднего эоцена Северной Америки.
Позднейшей группой насекомоядных плезиадапиформов были толиапиниды Toliapinidae (Hooker et al., 1999). Они появились лишь в конце палеоцена и дожили до среднего эоцена. Большинство видов, включая примитивнейшего Sarnacius gingerichi, найдено в Европе, но Altiatlasius koulchii мигрировал в Марокко, а самый поздний – Seia shahi – обнаружен в Пакистане. Altiatlasius koulchii первоначально был описан как древнейший настоящий примат семейства Omomyidae, потом считался даже антропоидом-эосимидом (это было сенсацией, так как получалось, что человекоподобные приматы появились уже в конце палеоцена, 57–60 млн лет назад!), но позже пересмотрен как плезиадапиформ. Толиапиниды были весьма схожи с микромомиидами по образу жизни и являются их полным “старосветским” аналогом; к сожалению, от тех и других найдено очень немного останков.
Среди плезиадапиформов особняком стоит семейство Adapisoriculidae. Особенности его представителей позволяли разным исследователям включать его в сумчатых, насекомоядных, лептиктид или миксодектид, а также считать предковым для тупай. Адаписорикулиды назывались и как возможные предки афросорицид Afrosoricida, включающих современных златокротов Chrysochloridae и тенреков Tenrecidae (Seiffert, 2010). Несмотря на существенные различия с плезиадаписовыми в зубной системе, строение посткраниального скелета (то есть всего скелета, кроме черепа) у плезиадапиформов, шерстокрыловых и адаписорикулид весьма схоже (Smith et al., 2010); судя по всему, это были древесные насекомоядные животные. Более того, плечевая кость адаписорикулид имеет промежуточное строение между вариантами меловых “кондиляртр” и кайнозойских эуархонт, причем в ряду самих адаписорикулид обнаруживается постепенный переход от более примитивного варианта мелового Deccanolestes к более эуархонтовому у палеогеновых форм (Boyer et al., 2010b). Если считать адаписорикулид приматами или приматоморфами, а Deccanolestes hislopi – адаписорикулидом, то эта группа становится самой древней среди приматоморфов, поскольку указанный вид обнаружен в позднемеловых отложениях Индии. Учитывая, что Индия в то время была островом, удаленным от всех других земель, а Purgatorius в то же время жил в Северной Америке, пути миграций становятся совсем непонятными.
Замечательно, что адаписорикулиды найдены в нижнем и верхнем палеоцене и нижнем эоцене Европы, а также верхнем палеоцене и нижнем эоцене Северной Африки, но ни малейших их следов нет в хорошо изученных фаунах Северной Америки и главной части Азии. Таким образом, либо адаписорикулиды были до крайности приматоподобными сумчатыми или насекомоядными, либо приматы зародились не в Северной Америке. Сами адаписорикулиды могли возникнуть в разных местах: древнейшие представители найдены в верхнем мелу Индии (несколько видов одного или двух родов, отсюда они могли попасть в Европу через Восточную Африку или острова моря Тетис), но самое большое разнообразие обнаруживается в нижнем палеоцене Европы (они явно появились тут раньше, но меловые европейские млекопитающие известны плохо, так что в Индию и Африку могли попасть отсюда), а примитивнейший вид Afrodon chleuhi известен из верхнего палеоцена Марокко (хотя более ранних в Африке пока не нашли, потенциально они могли распространиться отсюда и в Индию, и в Европу; Smith et al., 2010; De Bast et al., 2012). Не исключено, что предки адаписорикулид жили в Европе и оттуда попали в Северную Америку, только там стали пургаториусами, а позже – плезиадапиформами. С другой стороны, адаписорикулиды могут быть предками современных шерстокрылов; из Индии они могли попасть в Юго-Восточную Азию, когда эти две части суши соединились (Smith et al., 2010). С третьей стороны, даже отнесение рода Deccanolestes к плацентарным вызывает сомнения; согласно некоторым кладистическим расчетам, он вместе с прочими адаписорикулидами может представлять очень архаичную линию (намного примитивнее, чем Cimolestes, Zalambdalestes и Purgatorius), восходящую к корням всех эутериев, поздние представители которой сохранили примитивные черты до весьма поздних времен (Goswami et al., 2011). Впрочем, адекватность результатов кластерного анализа, применявшегося в подобных исследованиях по выявлению филогении, мягко говоря, спорна, что можно видеть, например, при сравнении выводов двух групп палеонтологов: в одной работе адаписорикулиды оказались предками тенреков (Seiffert, 2010), во второй – ответвлением древнейших и примитивнейших эутериев, даже близко не родственных ни тенрекам, ни эуархонтам (Goswami et al., 2011). Как бы то ни было, существование адаписорикулид свидетельствует, во-первых, о возможности межконтинентальных обменов фаун в позднем мелу и раннем палеогене, а во-вторых, о существовании примитивных групп, близких к предкам приматов, на осколках Гондваны – в Африке и Индии.
Специфические признаки и особенности являются главным препятствием для признания плезиадапиформов в качестве настоящих приматов. Ведь они обладали маленьким мозгом (гораздо меньшим, чем у приматов таких же размеров, хотя и большим, чем у других зверей) с хорошо развитыми обонятельными центрами; зрение, судя по направленным в стороны глазам, не было стереоскопичным; имелись когти на пальцах, их большой палец не противопоставлялся, а кисть была больше “цеплятельной”, нежели “хватательной”. Поэтому, несмотря на тот факт, что плезиадапиформы уже ушли от сугубой насекомоядности, а некоторые стали специализированными листо– и фруктоядами, многие систематики склонны считать первыми приматами только эоценовых адапиформов Adapiformes и омомиформов Omomyiformes.
Имевшиеся же прогрессивные особенности плезиадапиформов не уберегли их от вымирания. Традиционно и по умолчанию считалось, что в их исчезновении повинны более продвинутые настоящие приматы. Однако изучение частот встречаемости останков рисует другую картину (Fleagle, 1999). Резкий спад численности плезиадапиформов начинается совершенно синхронно со столь же резким подъемом численности первых грызунов, тогда как лишь через некоторое время появляются и начинают распространяться полуобезьяны. Стало быть, плезиадапиформы проиграли эволюционную гонку грызунам, зато своим исчезновением освободили дорогу полуобезьянам; спасибо мышам – без них нас бы не было! Таким образом, отличия плезиадапиформов от приматов оказываются крайне существенными: то, чего не было у первых, не позволило им успешно конкурировать с грызунами; то же, что приобрели вторые, вывело их на новый уровень, недоступный грызунам.
Дальнейшее развитие группы привело к первой радиации приматов – возникновению множества новых видов, давших начало разным линиям приматов.
Полуобезьяны
Полуобезьяны (подотряд Strepsirrhini или Prosimii) включают четыре группы. Древнейшая и предковая для современных – адапиформы Adapiformes. Современные лемурообразные Lemuriformes, лориобразные Lorisiformes и руконожковые Chiromyiformes раньше объединялись вместе, но думается, что различия между ними сильно недооценивались. Срабатывал закон проекции: чем дальше приматы от человека, тем хуже мы их различаем, тем более склонны смешивать в одну большую кучу; а вот ближайших родственников мы классифицируем тщательно, подробно и с чувством. Но ныне приматологи стали объективнее.
Полуобезьяны отличаются от обезьян массой примитивных особенностей. Например, у них не полностью бинокулярное зрение: глаза ориентированы несколько наискосок. Вообще зрение играет меньшую роль в жизни полуобезьян, чем обезьян; обоняние же развито гораздо сильнее. Поэтому и мордочка у них вытянутая, а нос слит с верхней губой, что, в свою очередь, снижает способности к мимике. Да оно и понятно, ведь полуобезьяны ведут преимущественно ночной или сумеречный образ жизни.
У современных полуобезьян есть и черты специализации. На указательном пальце стопы есть коготь, нужный для расчесывания шерсти; для той же цели служит щетка из вытянутых нижних резцов и клыков, сильно выступающих вперед. Прилагается и особое устройство для очистки этой гребенки от шерстинок: под языком имеется “подъязык” – треугольный вырост с пильчатым краем. Очевидно, полуобезьяны очень пекутся о своей внешности. Зубная щетка из резцов и коготь на втором пальце стопы объединяют галаго, потто, лори и мадагаскарских лемуров, так что все они наверняка имели общего предка.
Кроме более примитивной морфологии, полуобезьяны отличаются от настоящих обезьян поведением. Это почти исключительно ночные или сумеречные животные. Почти все полуобезьяны – древесные. Насекомых и прочих мелких животных они едят гораздо чаще, чем обезьяны, что и логично при малых размерах тела. Некоторые виды ведут одиночный образ жизни, некоторые живут группами.
Адаписовые (Adapiformes или Adapoidea) появились в начале эоцена около 56 млн лет назад. Они были распространены в основном в Европе и Северной Америке, но известны также из Северной Африки и Азии. Большая часть адаписовых вымерла на границе эоцена и олигоцена, видимо из-за общего похолодания и сокращения площади лесов. Однако в Южной и Восточной Азии последние адаписовые дотянули даже до конца миоцена. Тут климат менялся меньше, так что примитивные группы имели хорошие шансы на выживание, неспроста тут и сейчас живут, например, тупайи. Последних адаписовых добили, похоже, не погодные неурядицы, а потомки-конкуренты – злобные лори.
От современных полуобезьян адаписовые наглядно отличаются отсутствием зубной щетки: что поделать, дремучие времена – варварские нравы, парикмахерское искусство еще не было изобретено. Адаписовые имели размеры от мыши до большой кошки, были преимущественно древесными жителями, питались растениями, а некоторые и насекомыми, вели дневной или сумеречный образ жизни. Группа адаписовых очень разнообразна, во время ее образования – в раннем эоцене – происходила вторая радиация приматов.
Нотарктиды Notharctidae – весьма богатое семейство, появившееся в начале эоцена и исчезнувшее в начале олигоцена. Ранние представители подсемейства церкамониин Cercamoniinae (типа Donrussellia) были предками прочих групп нотарктид, а также семейств адапид Adapidae и азибиид Azibiidae. Таким образом, церкамониины – непосредственные, хотя и очень дальние предки лемуров. В 1984 году в Германии нашли целый скелет Darwinius masillae с отпечатком тела и шерсти вокруг; этот зверек жил 47 млн лет назад. В его строении можно усмотреть черты высших обезьян: мордочка короткая, половинки нижней челюсти сросшиеся, нет когтей на пальцах, на таранной кости имеется фасетка для малой берцовой кости. Впрочем, с наибольшей вероятностью эти продвинутые черты возникли конвергентно – независимо у дарвиниуса и обезьян. То же можно сказать про намного более позднего Afradapis longicristatus из Египта, жившего 37 млн лет назад, представителя особого подсемейства Caenopithecinae.
Другие ранние нотарктиды – азиадапины Asiadapinae – известны из Индии. Marcgodinotius indicus, живший 53 млн лет назад и весивший всего 130 г, настолько похож на примитивную обезьяну Anthrasimias gujaratensis из эосимид Eosimiidae, что до сих пор неясно – один это вид или два разных. Вместе с тем большинство черт Marcgodinotius indicus – наиболее примитивны среди всех адаписовых приматов. Такая комбинация, если вдуматься, логична: самые примитивные полуобезьяны должны быть похожи на самых примитивных обезьян.
Отлично сохранились останки нотарктуса Notharctus tenebrosus из подсемейства Notharctinae из Северной Америки. Этот почти полуметровый примат с длинным хвостом весьма напоминал лемуров, однако имел даже более продвинутое строение: его морда уменьшена, а глаза развернуты вперед и окружены замкнутым костным кольцом. Его родственник смилодектес Smilodectes gracilis имел еще более короткую мордочку; обонятельные луковицы смилодектеса были редуцированы, а зрительные центры мозга развиты, что свидетельствует о дневном образе жизни. Как и большинство адаписовых, он был древесным и листоядным.
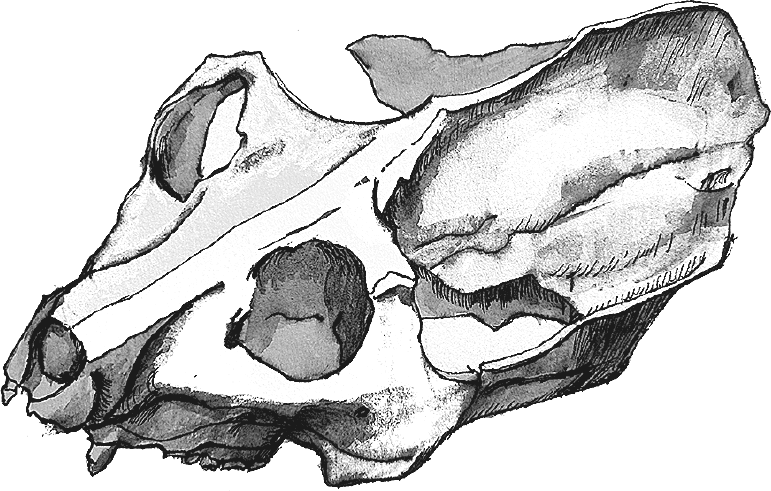
Рис. 27. Череп Adapis parisiensis.
Адапиды Adapidae известны преимущественно из Франции, Германии и Швейцарии, но в 1990-х гг. их представители были найдены в Китае (Adapoides troglodytes) и Северной Африке (Djebelemur martinezi и близкие ему формы, выделяемые в особое подсемейство Djebelemurinae). Лучше всего изучены парижский адапис Adapis parisiensis и более крупный Leptadapis magnus, от которых найдено довольно много целых черепов, в том числе более двадцати – еще в XIX веке. Обилие материалов позволило рассчитать уровень полового диморфизма, то есть степень различий самцов и самок. Выяснилось, что самцы весили примерно в полтора раза больше самок, а половой диморфизм по клыкам не больше или совсем немного больше, чем по размерам тела (Gingerich, 1981). Стало быть, межсамцовая конкуренция была незначительной, а в группах, скорее всего, присутствовало сразу множество и самок, и самцов.
Leptadapis magnus весил в среднем 8,4–9 кг, а отборные самцы аж 11,5 кг, тогда как чуть более поздний Adapis parisiensis – всего 2 кг (Gingerich et Martin, 1981). Мозг последнего имел размер лишь 8,8 см³ (втрое меньше, чем у кошки!); сравнение с современными полуобезьянами показывает, что адапис сильно уступал им в интеллекте. Маленькие глазницы свидетельствуют о дневном образе жизни, очень маленькие подглазничные отверстия – о редукции вибрисс. Слух у адаписов был отличный, но зрение все же играло роль главного органа чувств. Адаписы были древесными и преимущественно листоядными животными.
Североафриканские азибииды Azibiidae, жившие от 52 до 35 млн лет назад, были намного более мелкими животными: азибиус Azibius trerki – 115–160 г, другой вид того же рода – 630–920 г, алжирипитек Algeripithecus minutus – всего 65–85 г. Это были древесные ночные животные с очень большими глазами и длинными вибриссами. У нижне-среднеэоценового алжирипитека с большой вероятностью имелась зубная щетка из нижних резцов, так что этот примат – один из лучших кандидатов на роль общего предка лемурообразных и лориобразных.
Впрочем, на это почетное звание претендует и другой североафриканский примат – джебелемур Djebelemur martinezi, живший около 50 млн лет назад в Тунисе. Его выделяют в подсемейство Djebelemurinae, подгруппу адапид. Хотя у него нет зубной щетки, все же по комплексу признаков он оказывается отличным предком для африканских галаго, тем более что и по размерам – 100 г – он идеально соответствует карликовым галаго.
Последние адаписовые – сиваладапиды Sivaladapidae – дожили до конца миоцена (10 или даже 7 млн лет назад). Это были не очень крупные, но и не слишком мелкие – от 0,5 до 4 кг – животные, древесные, частично фрукто-, частично насекомоядные. В Таиланде найден среднемиоценовый Siamoadapis maemohensis. В Индии и Пакистане позднемиоценовые Sivaladapis nagrii и Indraloris himalayensis найдены вместе с останками сивапитеков Sivapithecus, а в Китае три вида Sinoadapis – с человекообразными юаньмоупитеками Yuanmoupithecus, люфенгпитеками Lufengpithecus и лаккопитеками Laccopithecus. Таким образом, последние представители примитивнейших полуобезьян передали эволюционную эстафету продвинутым гоминоидам. Современные орангутаны ловят лори; может быть, сивапитеки так же гонялись за их отдаленными родственниками? Вымирание последних адаписовых совпало с периодом похолодания и осушения климата; видимо, и конкуренция со стороны полуобезьян новых типов заметно возросла.
Некоторые палеоприматологи предполагают, что адаписовые могли быть предками и высших обезьян – это так называемая “адапоидная гипотеза”. В качестве переходных форм от адаписовых к обезьянам назывались Darwinius masillae, Algeripithecus minutus и Djebelemur martinezi, но сторонников этой точки зрения сейчас немного.
Современные полуобезьяны делятся на лемурообразных Lemuriformes (лемуры Мадагаскара), лориобразных Lorisiformes (галаго и потто Африки и лори Юго-Восточной Азии) и руконожковых Chiromyiformes (руконожка Мадагаскара). Размер этих животных колеблется от мыши до небольшой собаки, некоторые ископаемые лемуры были с крупную собаку или даже гориллу. Практически все они ночные или сумеречные, отчего круглые глаза – самая яркая черта всех полуобезьян.
Современные лемуры – жители только одного острова, но на нем, к сожалению, не найдены древнейшие предки лемуров. Удивительным образом ближайший родственник – Bugtilemur mathesoni – обнаружен в нижнем олигоцене Пакистана (впрочем, он может быть древнейшим мышиным лемуром, адаписовым приматом или же вообще шерстокрылом подсемейства Ekgmowechashalinae). Как лемуров занесло на Мадагаскар – непонятно. Остров был отделен от Африки уже в мезозое 160–165 млн лет назад, так что приматы могли достичь своей земли обетованной только по воде – на плотах из переплетенных растений, скорее всего, из Восточной Африки.
Предки руконожки точно неизвестны, в наибольшей степени на нее похож особый примат плезиопитек Plesiopithecus teras, чьи останки найдены в египетских отложениях с датировкой около 34 млн лет назад. Его своеобразие столь велико, что он выделяется в собственное надсемейство полуобезьян Plesiopithecoidea. С руконожкой плезиопитека роднит общее сходство формы черепа и строение зубов, в особенности выступающие вперед огромные нижние резцы (Godinot, 2006).
Лемуры Мадагаскара часто упоминаются как “живые ископаемые”, поскольку они действительно очень похожи на полуобезьян эоцена. В популярной литературе обычно пишется, что лемуры смогли дожить до современности благодаря изоляции, поскольку высшие приматы на Мадагаскар так и не попали. Это верно лишь отчасти, ведь ближайшие родственники лемуров живут и в материковой части Африки, и в Азии в одних лесах с обезьянами. Правдивость же такого утверждения заключается в том, что на Мадагаскаре сохранилось и развилось необычайное разнообразие лемуров, поскольку они заняли все экологические ниши, потенциально годные для обезьян. В Африке же и Азии потто, галаго и лори живут в достаточно специфических экологических нишах.
Среди мадагаскарских лемуров наиболее известны лемуры-катта Lemur catta (они же – кошачьи или кольцехвостые лемуры), обладатели запоминающихся длинных хвостов в черно-белую полоску. Они живут территориальными общинами, весьма ревностно охраняя границы от соседних стай. Не самым стандартным для приматов является главенство самок – настоящий матриархат. Также необычен для приматов большей частью наземный образ жизни; объясняется он жизнью в засушливых районах юга Мадагаскара, где с деревьями бывают проблемы. Хотя катты отлично лазают и прыгают по деревьям, большую часть времени они проводят на земле. Из-за этого длина рук и ног у катт примерно одинаковая. Хвост от наземной жизни не исчез, а приобрел новое – сигнальное – значение. Показательно, что то же самое случилось с павианами в саваннах Африки. К сожалению, конкуренция с людьми оставляет кошачьим лемурам мало шансов на окончательный выход в саванну и дальнейшее развитие в сторону разумности, хотя в сравнении с другими лемурами катта находятся в самом благоприятном положении, их численность пока довольно велика.
Оригинальное отличие имеется между двумя подвидами черного лемура: Eulemur macaco macaco обладает желто-оранжевыми глазами, а Eulemur macaco flavifrons – голубыми. Кстати, у этих же животных сильно выражен половой диморфизм по окраске: самцы черные, а самки коричневые.
Индри Indri indri выделяются резко укороченным хвостом – всего пять сантиметров. Думается, что редукция хвоста у индри не случайность, ведь эти животные – самые крупные современные полуобезьяны, достигающие от полуметра почти до метра длины и весящие до 9,5 кг. Видимо, увеличение веса снижает значение хвоста как балансира совершенно аналогично человекообразным обезьянам. Зато большой палец и на кисти, и особенно на стопе совершенно огромный, резко противопоставленный прочим, сросшимся к тому же до середины длины. Индри живут маленькими постоянными семьями; в отличие от большинства лемуров они ведут дневной образ жизни.
Специализация к прыжкам по деревьям у индри и их ближайших родственников сифак Propithecus зашла настолько далеко, что по земле на четвереньках они перемещаться не могут, а вынуждены скакать боком на двух ногах, подняв руки. По деревьям же они галопируют с легкостью неимоверной. Даже индри, несмотря на размер, умеют неплохо прыгать, причем делают это боком из сидячего положения на вертикальном стволе. Сифака Верро Propithecus verreauxi, живущая в засушливых лесах Юго-Западного Мадагаскара, ничтоже сумняшеся сигает по таким колючим стволам, что больно смотреть. Как она при этом умудряется не разодраться в клочья – науке неизвестно.
Особый подвид серого полумаки Hapalemur griseus alaotrensis, обитающий по топким берегам озера Алаотра, – единственный примат, населяющий болота. Эти редкие зверьки живут в зарослях камыша и питаются его листьями, в отличие от ближайших родственников других подвидов и видов, которые живут в лесах из гигантского бамбука и едят бамбуковые листья.
Мышиные лемуры Microcebus murinus – одни из самых мелких приматов в мире, соперничающие за это звание с карликовыми игрунками: наименьшая длина их тела без хвоста – около 10 см, плюс 15 см приходится на хвост. А вот гигантский мышиный лемур Mirza coquereli имеет длину тела аж 25 см и вес 300 г! Родственные жирохвостые лемуры Cheirogaleus впадают в спячку на время сухого сезона, перед чем запасают жир в хвосте. Это единственные приматы, впадающие в спячку. Многие мышиные лемуры питаются нектаром цветов или древесными смолами.
Особым своеобразием отличается руконожка ай-ай Daubentonia madagascariensis. Количество ее уникальных черт столь велико, что ныне ее выделяют в самостоятельный инфраотряд руконожковых, равный по рангу лемурообразным и лориобразным, хотя и включающий лишь один современный вид. Руконожка выделяется строением своих зубов, поскольку у нее всего по одному резцу в каждой половине челюсти, нет “зубной щетки”, нет клыков, нет премоляров на нижней челюсти и лишь по одному премоляру на верхней. Резцы растут постоянно, как у грызунов; руконожка прогрызает ими кору деревьев, добираясь до пустот-ходов личинок насекомых. Конечности ее специализированы, средний палец на руке чрезвычайно вытянут и используется для простукивания деревьев в поисках тех самых пустот-ходов, а также для вытаскивания оттуда личинок. Современные руконожки размером с небольшую кошку, примерно 2,5 кг весом, тогда как ископаемая Daubentonia robusta могла весить аж 12,5 кг. Судя по немногочисленным известным останкам, этот гигант вымер в последнюю тысячу лет, скорее всего – с непосредственной помощью человека.
Разнообразны лемуры Мадагаскара, но в недавнем прошлом его населяли еще более удивительные животные. Когда первые индонезийцы высадились на берега острова и углубились в его первозданные леса, они нашли там фантастическую фауну. Мы не будем рассказывать тут об эпиорнисах – птицах Рух, нескольких видах карликовых бегемотов и аналогах трубкозубов Plesiorycteropus, составляющих уникальный отряд Bibymalagasia. Среди лемуров тоже хватало уникальных представителей. К сожалению, все они пали жертвами аппетита предков мальгашей – современных жителей острова. Ясно, что в первую очередь уничтожены были самые крупные животные. Из-за этого даже может сложиться впечатление, что ископаемая фауна Мадагаскара представлена исключительно гигантами, а нынешняя – карликами. На самом деле малютки были и раньше, но их останки имеют гораздо меньше шансов сохраниться.
Кстати, о чертогах, сокровищ полных…
В национальном парке “Циманампесотсе” на Мадагаскаре исследователи нашли три затопленные пещеры, битком набитые сокровищами: целыми черепами и скелетами рогатых крокодилов и птиц-эпиорнисов, гигантских фосс и карликовых бегемотов, летучих мышей и грызунов. Кости буквально лежат там грудами! Среди прочих богатств Циманампесотсе имеются многочисленные останки вымерших лемуров Megaladapis и Archaeoindris.Благодаря недоступности пещер кости сохранились идеально, как новенькие. Достать их можно, только ныряя с аквалангом. Только собирание и каталогизирование ценностей займет наверняка не один год, а их исследованием смогут потом заниматься многие поколения антропологов.
Например, современные вари Varecia весят 3–4 кг, а их ископаемые родственники Pachylemur – 8–10 кг, ласковидные лемуры Lepilemur – меньше килограмма, а Megaladapis – 50–88 или даже 140–200 кг, индри – меньше 10 кг, а его сородичи Mesopropithecus – 10–14 кг, Archaeolemur – 15–25 кг, Hadropithecus – 17–35 кг, Babakotia – 16–20 кг, Palaeopropithecus – 35–60 кг, Archaeoindris – от 130 до 200 кг.
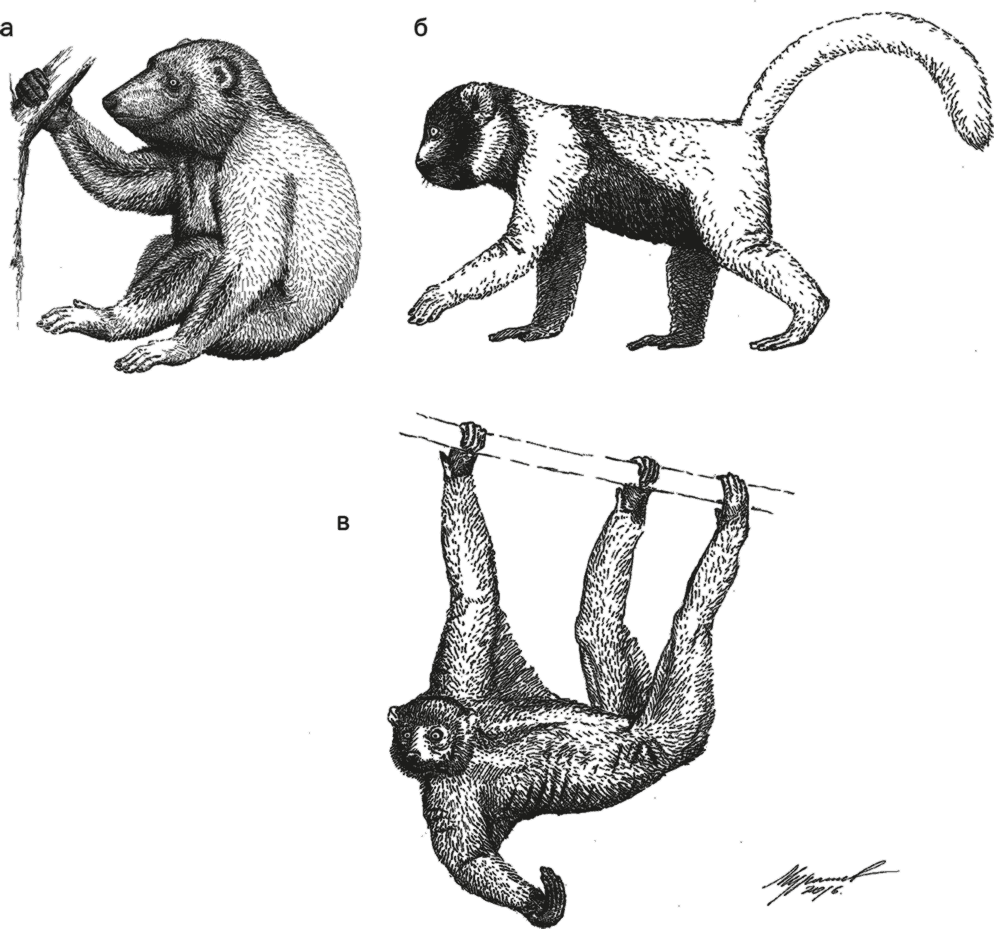
Рис. 29. Реконструкция Megaladapis (а), Hadropithecus (б) и Palaeopropithecus (в).
Мегаладаписы Megaladapis были медленными листоядными древолазами, подобными огромным коалам. Судя по костям со следами разделки, они были желанным угощением на столах мальгашей вплоть до прибытия европейцев примерно полтысячи лет назад. Особенно впечатляющими габаритами отличался Megaladapis edwardsi. Не очень родственным, но очень похожим – как по габаритам, там и по пропорциям – был археоиндри Archaeoindris fontoynonti. У этих огромных существ – и мегаладаписов, и археоиндри – руки были заметно длиннее ног, предполагалось также, что они были наземными и вели образ жизни вроде современных горилл.
Гадропитеки Hadropithecus stenognathus характеризовались почти круглой головой с очень короткой мордой и мощнейшими челюстями. Их передние зубы были уменьшены, а моляры увеличены и имели бугристый вид, что позволяло пережевывать жесткие стебли и семена. Судя по коротким ногам с недлинными пальцами и малоподвижными суставами, гадропитеки были наземными животными. Таким образом, эти животные экологически были чем-то средним между современными геладами и гориллами.
Палеопропитеки Palaeopropithecus отличались отсутствием нижних клыков, вытянутым носом, крайне длинными руками (относительно ног – длиннее, чем у орангутанов и гиббонов!) с подвижными суставами, вытянутыми изогнутыми пальцами и уменьшенным большим пальцем. Очевидно, это были крайне специализированные древолазы, почти не спускавшиеся на землю. Возможно, они могли висеть на ветках вниз головой типа ленивцев или даже брахиировать подобно гиббонам, хотя бы и не так лихо.
Минутка фантазии
Некоторые криптозоологи не оставляют надежд, что какие-то из гигантских лемуров все еще хоронятся в дремучих лесах Мадагаскара. Упования эти основываются на свидетельствах местных жителей. Например, на севере Мадагаскара записаны предания о каланоро – маленьких “гоминоидах”, ныне вымерших, хотя один из них якобы был пойман в 1879 г. (Forth, 2012, p. 210). Трудно представить, что каланоро – какие-нибудь архаичные гоминиды типа “хоббитов”, поскольку древнейшие следы заселения острова оставлены всего около 4 тыс. лет назад (Gommery et al., 2011). Зато крупные лемуры как никто подходят на роль “лесных человечков”. Впрочем, леса Мадагаскара не такие уж бескрайние, как хотелось бы, они слишком быстро вырубаются, так что надежды на сохранение мегаладаписов или каких-то иных гигантов призрачны. Остается надеяться на их возрождение с помощью клонирования, ведь потенциально гены можно добыть из костей. Дело за технологиями.
Африканские галаго Galago – очень подвижные ночные животные, охотящиеся за насекомыми в кустарниковых зарослях. Галаго могут прыгать по ветвям на двух задних конечностях, балансируя длинным пушистым хвостом; из-за этого у них из пяточной и ладьевидной костей стопы развилась длинная цевка, хотя и не столь впечатляющая, как у долгопятов. Как и все немадагаскарские полуобезьяны, галаго – ночные животные, преимущественно насекомоядные, но уважающие и фрукты, и древесные соки.
Галаго уникальны тем, что переносят детенышей, держа их зубами, как кошки. Видимо, при столь бодром скакании цепляющийся за шерсть детеныш просто слетал бы с мамы.
Ископаемые галаго современного рода известны из миоцена и плиоцена – 3–11 млн лет назад – Северной и Восточной Африки. Там же жили похожие животные Laetolia, а несколько раньше – 15–20 млн лет назад – Mioeuoticus, Komba и Progalago, выделяемые в собственные подсемейства. Гораздо более древний сахарагалаго Saharagalago misrensis найден в отложениях конца эоцена Египта – 35–41 млн лет назад. Они уже древнее вадилемура Wadilemur elegans, жившего в том же Египте 34–35 млн лет назад, но классифицированного как представителя подсемейства Cercamoniinae (самого примитивного подсемейства среди адаписовых). Из всех адаписовых вадилемур и более древний джебелемур в наибольшей степени подходят на роль предков галаговых. Таким образом, корни галаговых, как и прочих современных полуобезьян, кроются среди изобилия эоценовых адаписовых. В те времена ареал таких животных мог расширяться и в более северные земли: среднеэоценовая хасселясия Chasselasia eldredgei, определяемая как примитивнейшее лориобразное, близкое к галаго, найдена в Швейцарии. То ли предки галаго мигрировали из Европы в Африку, то ли наоборот – новые находки должны прояснить ситуацию.
В противоположность быстрым галаго, африканские потто и азиатские лори – весьма медлительные животные с печальным выражением лица, не спеша подкрадывающиеся под покровом ночи к добыче и хватающие ее быстрым движением рук. Впрочем, все лориобразные едят и фрукты, а у африканских лори Perodicticus potto они составляют основу рациона.
Африканские золотые потто Arctocebus calabarensis – почти совершенные аналоги азиатских толстых Nycticebus и тонких Loris лори. Выше уже упоминалось, что толстые лори обладают ядовитыми железами на плечах и способны изображать очковую кобру, а потто обзавелись шипами на холке. Эти странные оборонительные стратегии, как бы чудно ни выглядели, достаточно действенны, чтобы обеспечить выживание своих обладателей.
Каранисия Karanisia из конца эоцена (35–41,2 млн лет назад) Ливии и Египта является древнейшим потто. Она была намного меньше своих потомков.
Лори современного рода Nycticebus linglom известны из миоцена Таиланда (18 млн лет назад). Родственные звери Nycticeboides simpsoni и Microloris pilbeami жили в Пакистане 9–10 млн лет назад, хотя неопределенные останки похожих животных есть там и в более древних слоях – до 15–16 млн лет назад.
Назад: Глава 40 Дальняя родня: тупайи и шерстокрылы
Дальше: Глава 42 Обезьяны

