Книга: Достающее звено. Книга 2. Люди
Назад: Глава 4 Звено между звеньями: “ранние Homo”, или Загадочный миллион: заря и первая половина истории Homo
Дальше: Глава 5 Homo erectus: непрямые дороги человека прямоходящего
Мозаика из Малапы
Южноафриканская пещера Малапа прославилась в 2008 г., когда в ней были найдены останки четырех удивительных существ. Два млн лет назад в подземную ловушку угодили взрослая самка, подросток-самец и детеныш. От первых двух сохранились почти целые скелеты, от детеныша – фрагменты конечностей; найдена также изолированная большая берцовая кость взрослого. Изучение уникальных останков вылилось в описание вида Australopithecus sediba, строго промежуточного между австралопитеками и “ранними Homo”, так что некоторые антропологи склонны говорить о Homo sediba. Находки из Малапы позволили говорить о Южной Африке как о возможной прародине человека. Детальные исследования уникальных окаменелостей уточняют и конкретизируют картину столь важного этапа антропогенеза – возникновения собственно человеческого рода.Показательно, что прогрессивные и примитивные признаки разных частей скелета распределены у малапских гоминид крайне неравномерно и мозаично, прогрессивность в одной детали компенсируется архаизмом в другой: часть не отличается от состояния, типичного для современного человека, тогда как другая слабо отличима от обезьян или занимает более или менее промежуточное положение. Сравнение A. sediba с A. africanus и A. afarensis показывает, что крайне трудно по одному из фрагментов предсказать уровень продвинутости какого-либо другого. Замечательно также, что ряд признаков малапских гоминид оказался прогрессивнее, чем у известных и притом более поздних хабилисов! Это является сильным аргументом в пользу южноафриканского происхождения рода Homo. Поэтому снова и снова встает вопрос о принадлежности малапских гоминид к роду Homo. Если уж они прогрессивнее хабилисов, то либо хабилисы – австралопитеки, либо седибы – Homo. Сложность дефиниций усугубляется крайней недостаточностью сведений относительно морфологии хабилисов: их черепа более-менее известны, но о строении посткраниального скелета антропологи доныне знают немного. Известные факты свидетельствуют о значительной изменчивости хабилисов, по некоторым параметрам их можно даже разнести на два-три, а то и четыре вида. Реальная изменчивость H. habilis наверняка была еще больше известной нам, так что седибы вполне могут укладываться в ее рамки, хотя сейчас антропологам трудно это оценить достоверно.Закономерно, что по подавляющему большинству черт A. sediba похож на A. africanus, а не на восточноафриканских австралопитеков. Это, с одной стороны, логично биогеографически, с другой – свидетельствует о параллельной гоминизации на юге и востоке Африки. Шел ли процесс очеловечивания южноафриканских австралопитеков совершенно независимо, или же они “по пути” периодически смешивались с северными кузенами – неясно. Учитывая географию Африки, второй вариант представляется более вероятным. Видимо, с грацильными австралопитеками и “ранними Homo” сложилась примерно такая же ситуация, какая существует с нынешними павианами: крайние формы четко отличаются, но в промежутках есть переходные формы, а все они вполне могут скрещиваться.Морфология малапцев была подробно описана в двух циклах статей в 2011 и 2013 г. в журнале Science.Авторы исследования зубов на основании огромного количества описательных признаков (22 черты) пришли к выводу, что A. sediba, входящий в один кластер с A. africanus, ближе к H. habilis и более поздним гоминидам, чем A. afarensis, объединяющийся с массивными австралопитеками (Irish et al., 2013). Вроде как A. africanus и не слишком родственен A. afarensis'у. Из этого следует логичный вывод, что Восточная Африка как бы стояла в стороне от прогресса, тогда как в Южной он бурлил по полной. Кстати, размеры зубов седибы очень маленькие, уступают любым австралопитекам и “ранним Homo”, что вроде как очень прогрессивно.Хочется, правда, сказать три но. Первое: использованный кластерный анализ – не лучший инструмент для оценки сходств и различий высокоизменчивых групп. Уж очень он округляет и усредняет, упрощает реальность. Второе: одонтологические признаки – далеко не самые надежные для выявления близости в сильнородственных группах. Наглядным тому доказательством является кладограмма, приведенная в электронном приложении к статье: негроиды в ней зависли между A. sediba и H. habilis с H. erectus, тогда как современные “североафриканцы” оказались общими предками всех прочих рассмотренных групп; нельзя отказать авторам, опубликовавшим такую кладограмму, в юморе. Куда-то бы попали монголоиды? Ясно, что, используя иные алгоритмы, можно получить и прочие забавные распределения групп, а та, что вошла в основной текст, просто больше понравилась авторам. Третье: рассмотренные восточноафриканские A. afarensis сильно древнее прочих включенных в анализ видов, так что их сходство с парантропами и отдаленность от “ранних Homo” необязательно отражает реальное родство. Видимо, у парантропов рассмотренные признаки менялись медленно (не зря они считаются более примитивными), а в гомининной линии – быстро. A. africanus же просто несколько моложе геологически, чем A. afarensis, что и отразилось в их “большей прогрессивности”.Исследование нижней челюсти второго индивида из Малапы показало, что он весьма отличается от A. africanus (Ruiter et al., 2013) – вывод прямо противоположный тому, что сделан для зубов! Отличия комплексны, заключаются как в размерах и форме, так и в ростовых процессах. Приятно, что видовой статус этим подтверждается, но неясно, откуда взялась такая специфика. На графиках канонического анализа (вот она – победа разума – не кластерного, не главных компонент, а канонического!!!) малапская челюсть занимает обособленное положение скорее рядом с A. afarensis и H. habilis с H. erectus, чем с кем-то еще. Впрочем, в приложении приведены и иные варианты анализов, в коих A. sediba оказывается едва ли не массивным австралопитеком. Имея некоторый опыт построения и анализа аналогичных графиков, могу ответственно заявить свое но: парой графиков в подобном случае обойтись никак нельзя. Единичное наблюдение может гулять по всем возможным осям довольно вольготно в зависимости от множества случайных моментов. Можно оценить соотношение групп наблюдений, но индивидуальная диагностика таким способом почти невозможна. Вот если бы в статье были приведены индивидуальные значения на графиках, а не точки для абстрактных средних – смысла было бы заметно больше.Мозг гоминида Малапа 1 не очень большой – 420 см³ – и в целом соответствует вариациям грацильных австралопитеков (Carlson et al., 2011). Показательно сильное сужение лобных долей и заметная уплощенность их сверху. Вместе с тем так называемый “лобный клюв” – выступание вниз центральной части лобных долей – несколько меньше, чем у австралопитеков, и гораздо меньше, чем у обезьян. Совсем прогрессивно выглядит глубокая височная ямка между лобной и височной долей, отсутствующая у обезьян и многих австралопитеков. Несмотря на то, что по абсолютным размерам и простейшим пропорциям мозг Малапы 1 уклоняется скорее в сторону шимпанзе, многомерный анализ главных компонент формы надглазничной области лобной доли поместил Малапу 1 на край распределения современных людей. Как особенно прогрессивный признак отмечено смещение назад обонятельных луковиц. Строго говоря, метод главных компонент неприменим при межвидовых сравнениях, но такова уж традиция зарубежной антропологии, и более адекватные методы, например канонический анализ, пока не применялись для анализа эндокрана из Малапы. Серьезным недочетом исследования следует считать отсутствие в анализе эндокранов хабилисов; из австралопитеков для сравнения привлечены лишь южноафриканские грацильные Sts 5 и Sts 60. Таким образом, из опубликованной работы о мозге Малапы 1 нельзя сделать выводов о прогрессивности этого гоминида. Как назло, на всех четырех известных эндокранах хабилисов реконструирована именно та область, которая сохранилась у Малапы 1 (из них OH 7 реконструирован вообще по одним теменным костям). Все же можно отметить, что эндокран KNM-ER 1805 выглядит гораздо более шимпанзеподобно, чем Малапа 1; у KNM-ER 1813, вероятно, была слабее выражена височная ямка, а KNM-ER 1470 весьма и весьма похож на Малапу 1. Последнее обстоятельство особенно важно, учитывая новые датировки KNM-ER 1470 – около 2,03 млн лет назад – “всего” на 50–60 тыс. лет древнее, чем Малапа, а вспоминая о погрешности определения возраста – они вообще современники.Позвоночный столб австралопитеков изучался уже не раз, но почти всегда отдельно от остального скелета. В поясничном отделе седибы с наибольшей вероятностью было пять позвонков – стандарт для современного человека (Williams et al., 2013). Ранее у разных австралопитеков и даже неандертальцев регулярно обнаруживались отхождения от этого правила. Еще важнее, что есть все основания предполагать у A. sediba наличие поясничного лордоза – изгиба вперед. Поясничный лордоз отсутствует у шимпанзе, но формируется у человеческих детей в раннем детстве, когда ребенок начинает сидеть, ползать и ходить; он жестко связан с прямохождением (у горилл поясничный лордоз тоже хорошо выражен, но обусловлен огромным весом тела). Крестец седибы содержит пять позвонков – как у современного человека. Его ушковидная поверхность – место соединения с подвздошной тазовой костью – велика и обширна, что связано с двуногой походкой и соответственно возросшими нагрузками. Отличием от человека можно считать некоторую относительную удлиненность поясницы и ее большую гибкость – черты, вероятно полезные при регулярном лазании по деревьям. В целом позвоночник A. sediba оказывается прогрессивнее, чем у австралопитеков, и примерно столь же развит, как у “мальчика из Нариокотоме”, то есть Homo ergaster'а, жившего на полмиллиона лет позже.Далее – грудная клетка (Schmid et al., 2013). Первым делом стоит отметить, что эта часть скелета сохраняется всегда хуже прочих. Фактически до сих пор не было ни одного примера хорошо изученной грудной клетки австралопитеков или “ранних Homo”. Уникальная сохранность малапских находок позволила установить, что грудная клетка A. sediba больше похожа на шимпанзоидную, нежели человеческую. Она имеет удивительно обезьяньи пропорции в целом: низкая и очень широкая, причем резко расширяется сверху вниз. Очевидно, такое строение тесно связано с обезьяньим планом строения верхней конечности. Суженность верхней части грудной клетки, как предполагается, отражает неспособность ходьбы и бега на более-менее дальние расстояния, а расширенность нижней все же не столь резкая, как у человекообразных обезьян. То есть и по этому признаку мы имеем замечательно переходную форму.Следующая статья посвящена изучению костей верхней конечности (Churchill et al., 2013). Первым делом обращает на себя внимание крайняя архаика лопатки из Малапы. По подавляющему числу параметров она оказывается наиболее схожей с лопатками орангутанов. Плечевые кости тоже крайне примитивны и попадают в компанию с Люси, Нариокотоме, Дманиси и Лианг-Буа 1. Длина предплечья относительно длины плеча у Малапы почти идентична хабилису OH 62 и находится на границе между людьми и шимпанзе, с уклоном к первым. Общая мораль – A. sediba крайне примитивен в строении рук, он запросто мог лазать по деревьям… если бы не кисть. Впрочем, даже продвинутая кисть не мешала малапцам забираться на деревья, ибо и современные люди делают это очень хорошо.Кисть Малапы 2 сохранилась почти полностью, исключая дистальные фаланги всех пальцев, кроме большого (Kivell et al., 2011). В строении ее частей имеются как типично австралопитековые признаки: грацильность тел пястных с увеличенными головками и ориентация их оснований, сильно вогнутая форма основания первой пястной, маленькие головки, большие основания и сильный изгиб фаланг, мощные гребни сгибателей на нижней стороне фаланг, – так и почти совсем человеческие: относительно длинный большой и короткие прочие пальцы, укороченная и расширенная форма дистальной фаланги большого пальца, форма ладьевидной, головчатой и крючковидной костей. Замечательно, что по конфигурации некоторых костей, например ладьевидной и дистальной фаланги большого пальца, Малапа 2 оказывается ближе к человеку, чем хабилис OH 7. Это очевидно даже при внешнем сравнении и полностью подтверждается многомерными анализами ряда измерений: Малапа 2 попадает в область изменчивости современного человека, а OH 7 – шимпанзе и горилл. В целом общие пропорции кисти и кости запястья Малапы 2 почти человеческие, тогда как пястные и фаланги – австралопитековые. Авторами исследования это интерпретируется как следствие переходного статуса гоминид из Малапы, при котором одновременно и сохранялись способности к древолазанию, и появлялись способности к точечному захвату и изготовлению каменных орудий. Вместе с тем есть и специфические черты. Например, полулунная кость, хотя имеет в целом человеческую форму, обладает очень маленькой фасеткой для соединения с головчатой костью; впрочем, измерение фасетки, как это представлено в файле дополнительных материалов к статье, может быть поставлено под некоторое сомнение, вероятно, ее размер несколько занижен, а специфика, стало быть, завышена.Тазовые кости и крестец более-менее полноценно сохранились у Малапы 2; от таза Малапы 1 осталось меньше фрагментов, что не помешало исследователям выполнить реконструкцию целого таза (Kibii et al., 2011). Хотя достоверность некоторых деталей реконструкции может быть поставлена под сомнение, в целом таз гоминид из Малапы оказывается гораздо более похожим на человеческий, чем таз афарского австралопитека Люси. Впрочем, не обошлось и без архаичных особенностей, свойственных всем древним гоминидам от австралопитеков вплоть до неандертальцев: ширина таза очень велика, особенно существенна большая ширина между вертлужными впадинами, ибо именно она задает меньшую эффективность двуногой походки по сравнению с современным человеком. Напрямую связано с большой шириной таза удлинение лобковых ветвей. Примитивным признаком можно считать и малый размер крестцово-подвздошного сустава; впрочем, он может быть следствием аллометрической зависимости от малых размеров тела. Все же больше бросаются в глаза прогрессивные черты, отличающие малапских гоминид от австралопитеков: образование крестца из пяти позвонков, не слишком развернутые, относительно вертикальные, сигмовидно-изогнутые крылья, а также повышенная массивность тел подвздошных костей, форма и направление подвздошных остей, более вертикально ориентированная нижняя лобковая ветвь, укороченная седалищная кость и, соответственно, малое расстояние между седалищным бугром и вертлужной впадиной, ориентация седалищных бугра и ости несколько внутрь, отчего размер верхнего входа в малый таз оказывается относительно крупнее нижнего. В целом признаки таза гоминид из Малапы либо прогрессивны, либо занимают промежуточное положение между австралопитеками и людьми. Жаль, что мы ничего не знаем о строении таза хабилисов, отчего оценка прогрессивности гоминид из Малапы повисает в пропасти между австралопитеками и эректусами.Самая монументальная статья цикла посвящена нижним конечностям (DeSilva et al., 2013). Несть числа морфологическим особенностям, описанным в самой статье и приложении к ней. Можно выделить несколько самых интересных моментов. При прямом положении ноги стопа была очень сильно пронирована, то есть завернута внутрь. Не то чтобы малапец был косолапым, но при ходьбе он опирался на боковую сторону стопы, а не ставил ее прямо, как мы. Видимо, с перераспределением нагрузок при таком странном положении связано чрезмерное развитие нижней передней подвздошной ости на тазовой кости, поскольку туда крепятся соответствующие связки. Существенно, что у шимпанзе и прочих четвероногих этой ости фактически нет вовсе.Бедренная кость удивительно современна. Общая форма и детали пропорций почти не отличаются от человеческих и одновременно непохожи на те, что есть у шимпанзе. Конечно, без архаики и специфики тоже обойтись было никак нельзя. При взгляде сбоку форма мыщелков бедренной кости довольно округлая и укороченная, более схожая с обезьяньим вариантом. Специфичным признаком можно считать крайне низкий наклон шейки бедренной кости.Коленная чашечка из Малапы гораздо сильнее схожа с человеческой, нежели обезьяньей, пропорции малой берцовой кости оказываются строго промежуточными между человеческими и шимпанзиными.Таким образом, малапские A. sediba имеют удивительную мозаику примитивных, прогрессивных и уникальных черт в строении ног. У других же австралопитеков и “ранних Homo” сочетания признаков могут быть иными. Это позволило авторам предположить неоднократное возникновение прямохождения или как минимум независимое развитие нескольких его вариантов. Вывод смелый! Действительно, разнообразие вариаций в деталях строения у австралопитеков велико. Но говорит ли это о независимости путей их эволюции? Не принимаем ли мы индивидуальные особенности за глобальные эволюционные тренды? Не так много нам известно целых находок ног и тазов столь большой древности, чтобы можно было обосновать это статистически. Вариабельность подобных деталей строения у современных людей тоже весьма велика. Имеются варианты, связанные с полом и возрастом, болезнями и даже приобретаемыми привычками. Как различить среди этой чехарды магистраль эволюции? Видимо, еще и еще раз проверять и сравнивать, описывать и измерять, думать и сомневаться.Стопа Малапы 2 сочетает очень примитивные и очень продвинутые черты (Zipfel et al., 2011). Голеностопный сустав имеет более-менее человеческую форму и функции; вероятно, имелись своды стопы; как у человека, осуществлялось прикрепление ахиллова сухожилия на пяточной кости; таранная кость по массе признаков, включая такие важные, как торзион головки, форма блока, размер, выступание и наклон латеральной стороны, форма нижней части шейки, больше похожа на человеческую, чем кости австралопитеков, и напоминает вариант хабилисов. Но массивная медиальная лодыжка и форма заднего края голеностопной суставной поверхности большой берцовой кости, форма и грацильность пяточной кости, вертикальная ориентация и уплощенность передней стороны головки таранной кости, большой относительный размер этой головки, а также размеры, форма и наклон задней части таранной кости выглядят шимпанзеподобно. Многомерный анализ размеров и пропорций большой берцовой кости однозначно помещает Малапу 2 в область изменчивости людей, таранной кости – между людьми и шимпанзе или в стороне от них, а пяточной кости – в область шимпанзе. Предполагается, что это может быть следствием сохранения некоторой степени древесности гоминид из Малапы. Рельеф боковой стороны пяточной кости вообще крайне изменчив у людей и, вероятно, у обезьян, но у Малапы 2 он все же больше напоминает вариант шимпанзе. Удивительно, но гораздо более древние афарские австралопитеки оказываются по строению этого рельефа неотличимы от человека и, таким образом, более продвинутыми, чем Малапа 2. Это может быть использовано как аргумент в пользу специализированности гоминид из Малапы и исключения их из числа наших предков.Таким образом, детальное описание новых находок, как оно часто случается, увеличило число возможных интерпретаций филогении гоминид. С одной стороны, гоминиды из Малапы по ряду признаков прогрессивнее более поздних хабилисов; с другой стороны, в их строении масса типично австралопитековых и даже обезьяньих черт; несколько признаков могут считаться более примитивными, чем у австралопитеков, либо специализированными. Конечно, можно списать их на индивидуальную изменчивость; в этом случае малапские гоминиды должны расцениваться как древнейшие известные представители Homo. Однако можно считать особо примитивные и специализированные черты доказательством особого статуса гоминид из Малапы: по ряду черт они вырвались вперед в эволюционной гонке, но по другим резко отстали и в итоге оказались в эволюционном тупике. Кажется, последний вариант не совсем хорош, потому что репродуктивная изоляция среди приматов никогда не была слишком жесткой. Вряд ли изоляция между южно– и восточноафриканскими популяциями прогрессивных грацильных австралопитеков была особо сильной. Если не было изоляции страусов, носорогов, слонов, львов, леопардов, павианов и множества прочих видов, с какой стати должна была быть изоляция гоминид, активно осваивавших открытые пространства? Таким образом, гоминиды из Малапы, по-видимому, действительно олицетворяют собой еще одно “достающее звено” эволюции – звено между австралопитеками и людьми.Своеобразным дополнением ко всему вышесказанному явилось исследование диеты двух взрослых индивидов Australopithecus sediba (Henry et al., 2012). Микроструктура эмали голосует за твердые продукты в рационе, изотопный анализ – за растения лесов, а фитолиты – за разнообразие на столе. Из прочих ископаемых гоминид седибы по типу питания больше всего похожи на Ardipithecus ramidus. Самое интересное, в некоторые сезоны седибы, похоже, глодали кору и древесину; такого не замечено ни за одним человекообразным приматом. Нелегко, похоже, жилось в Южной Африке два миллиона лет назад!
Промежуточность малапцев между всеми возможными австралопитеками и эогомининами создает немалую проблему. Интерпретаций может быть множество. Авторы первоописания вида склонны считать, что A. sediba является потомком A. africanus и предком Homo habilis (Berger et al., 2010). Если воспринимать древнейших восточноафриканских Homo rudolfensis как специализированную тупиковую ветвь, родственную Kenyanthropus или Paranthropus, то южноафриканские A. sediba могли быть источником Homo habilis, расселившихся в более позднее время к северу. Вместе с тем седибы примерно с теми же основаниями могут быть потомками A. afarensis, мигрировавшими на юг. В обоих случаях A. sediba может быть и тупиковой ветвью, и предком самостоятельной ветви южноафриканских эогоминин. По времени малапцы попадают примерно в момент конца вида H. rudolfensis и начала H. habilis, но похожи на последних. Не исключено, что A. sediba – это своеобразные “параллельные хабилисы” Южной Африки; учитывая слабую репродуктивную изоляцию приматов и отсутствие принципиальных географических барьеров между восточной и южной частями континента, можно предположить возможность обособления гоминид на уровне скорее подвидов с сохранением возможности их смешения в будущем.
Вероятно, именно эти “парахабилисы” представлены более поздними находками, описанными суммарно как Telanthropus capensis или Homo gautengensis. Они обнаружены в основном в отложениях пятого уровня Стеркфонтейна и имеют датировки около 1,5–2 млн лет назад. Замечательно, что в этой же брекчии появляются и оббитые гальки. К сожалению, большинство “ранних Homo” Южной Африки представлено изолированными зубами, но есть и более представительные образцы. Наилучшую сохранность имеет череп Stw 53. Он определялся как позднейший Australopithecus africanus, Homo habilis, неизвестный вид и, наконец, послужил голотипом вида Homo gautengensis (Curnoe, 2010). С одной стороны, череп невелик и по общей форме схож с Australopithecus africanus, с другой – челюсти еще меньше и грацильнее. Череп в целом и затылок в частности округлы, с очень слабым рельефом, однако височные линии сильно сходятся, почти сливаются между собой. В строении височной кости сочетаются черты Australopithecus africanus и Homo habilis, но с преобладанием последних. Центральная часть лица выступает вперед, а скулы раздаются слабо, что сближает Stw 53 с загадочными восточноафриканскими KNM-ER 1805 и KNM-ER 1813. Типична для эогоминин уплощенность передней стороны альвеолярной дуги, но небо с альвеолярной дугой сильно вытянуты в длину. Вместе с тем имеются “передние лицевые валики”, характерные для южноафриканских австралопитеков и парантропов. Объем мозга Stw 53 достоверно неизвестен, поскольку черепная коробка разрушена; реконструкции дают возможный размах от 450 до 680 см³. Зубы чрезвычайно большие, крупнее даже, чем у Homo rudolfensis, достигающие нижних пределов изменчивости южноафриканских парантропов. Таким образом, Stw 53 может быть расценен либо как предок или ранняя форма Paranthropus robustus (чему противоречит слабое развитие скуловых дуг), либо как самка Homo rudolfensis.
Показательно, что на основании правого скулового отростка верхней челюсти Stw 53 имеются следы зарубок от каменных орудий: кто-то отрезал нижнюю челюсть от черепа (Pickering et al., 2000). Таким образом, один из древнейших представителей Homo оказывается и древнейшей жертвой каннибализма: темные стороны человеческой натуры появились раньше самого человека.
Ряд ценных находок сделан в Сварткрансе. В брекчии, отличной от брекчии с парантропами, были обнаружены останки, первоначально описанные как Telanthropus capensis. Их датировка – от 1,63 млн лет назад и позже (Curnoe et al., 2001), вероятно до 1,2 млн лет назад или еще меньше. В Сварткрансе обнаружены костяные орудия, использовавшиеся, судя по всему, для вылавливания термитов (Backwell et d'Errico, 2001); правда, остается вопрос – кто этим занимался?
Из так называемых телантропов наибольшую известность получили нижние челюсти SK 15 (Телантроп I) и SK 45 (Телантроп II), а также череп SK 80/846/847 (Телантроп III). На самом деле они не синхронны: SK 15 моложе двух прочих.
Наибольший интерес представляет, конечно, череп. Сохранился он плохо и собран из массы обломков. Также по размерам подходит к черепу нижняя челюсть SK 74a, но по морфологии она соответствует некрупному парантропу. Поэтому неудивительно, что SK 80/846/847 определяли и как грацильного австралопитека, и как самку мелкого массивного австралопитека, как Telanthropus capensis, Homo habilis, женщину Homo leakeyi и, конечно, Homo erectus. В пользу каждой версии находились свои аргументы. Наконец, вместе с прочими южноафриканскими эогомининами череп послужил основой описания вида Homo gautengensis. Черты действительно своеобразны: лицо массивное и сильно прогнатное, но не очень большое; средняя часть лица выступает вперед в поперечной плоскости, но плоская в вертикальной; скулы высокие и массивные, но не раздаются в стороны; небо и альвеолярная дуга широкие относительно лица и уплощены спереди, но к тому же резко вытянуты в длину, так что небо получается относительно суженным; надбровный рельеф резко выступает вперед, но надбровные валики тонкие при взгляде спереди; лоб очень узкий и плоский, потому что височные линии резко сходятся между собой, хотя, судя по всему, не сливались в сагиттальный гребень. Детали височной кости скорее эогомининные, чем австралопитековые, в частности, имеется шиловидный отросток, да и сосцевидный развит достаточно сильно. Объем мозга предположительно составлял 450 см³, то есть был очень мал, примерно как у Australopithecus; впрочем, возможны и иные реконструкции, поскольку мозговой отдел крайне разрушен. Зубы SK 80/846/847 чрезвычайно крупные, первый верхний премоляр по мезиодистальному диаметру заметно превосходит и парантропов, и всех эогоминин.
Интересные данные получены палеодиетологами: соотношение Sr/Ca у SK 80/846/847 находится вне пределов изменчивости Australopithecus robustus, тогда как соотношение 87Sr/86Sr идентично этому виду; следовательно, различия в соотношении Sr/Ca отражают различия в питании (Sillen et al., 1995).
Нижние челюсти телантропов SK 15 и SK 45 в целом схожи. Обе особенны очень малой высотой тела в задней части. Нижние моляры велики, но все же меньше, чем обычно у H. habilis.
В Сварткрансе найдены и другие кости, относящиеся к тому же виду. В частности, это фрагменты черепа детеныша SK 27, нижние концы маленькой левой плечевой кости SKX 10 924 и большой правой SKX 34 805, верхние части очень современно выглядящих лучевых костей SK 18b и SKX 2045, маленькая IV пястная кость SK 85, проксимальная фаланга большого пальца кисти SKX 27 431, проксимальная фаланга кисти SKX 22 741, фрагмент довольно крупной и прогрессивной правой бедренной кости SK 1896, а также набор изолированных зубов.
Существенно, что по ряду признаков Homo gautengensis выходят за рамки изменчивости H. habilis и H. ergaster, причем отличаются от обоих больше, чем они друг от друга (Curnoe, 2010). В частности, верхние челюсти телантропов сильно вытянуты в длину, нижние челюсти, как уже говорилось, имеют очень низкое в задней части тело, премоляры и моляры отличаются сильно суженной формой. Судя по всему, телантропы отличались сильным схождением височных линий, хотя и в Восточной Африке можно найти сопоставимые примеры.
От прочих эогоминин Южной Африки сохранилось намного меньше. В частности, это фрагменты двух скелетов детей из Дримолен-Кейв с датировкой около 2 млн лет назад, определенные как Homo habilis, но также иногда включаемые в вид Homo gautengensis. Молочные и постоянные нижние зубы KB 5223 из Кромдраая также предполагались останками “раннего Homo” (Braga et Thackeray, 2003), но с большей вероятностью принадлежали Paranthropus robustus (De Ruiter, 2007), в пользу чего свидетельствует редукция передних зубов, мегадонтия моляров и их морфология. Столь же большие сомнения существуют и относительно обломка моляра GDA 1 из Гондолин – он относился как к гипермассивным парантропам, возможно даже родственным бойсовским восточноафриканским (Menter et al., 1999), так и к Homo (Curnoe, 2010).
Южноафриканские гоминиды отличаются от восточноафриканских. Но являются ли эти отличия хронологическими, видовыми или аналогичны расовым? Была ли Южная Африка тупиком, в котором гоминиды специализировались и вымирали, или горнилом эволюции, выдающим на север прогрессивные формы? Возможно, ответы появятся с обнаружением и исследованием большего количества сохранных южноафриканских гоминид с датировками около 3 млн лет и восточноафриканских с древностью от 3 до 2 млн.
Странные Homo Южной Африки: naledi выходит на люди
Иногда, когда солнышко скроется за тучкой и изжога одолеет, лезут в голову грустные мысли, что миновали, дескать, времена Индиан Джонсов, когда можно было зайти в пещеру и узреть целый скелет неведомого существа, картинно раскинувшийся на полу.
Ан нет, не прошли!
Разве что теперь для этого надо быть тощим.
13 сентября 2013 года два спортсмена-спелеолога – Стивен Такер и Рик Хантер – залезли в пещеру Райзинг Стар в Южной Африке. Она уже была известна узким “шкуродером” “путь Супермена”, по которому могут проползти лишь исключительно субтильные личности. Но Стивену и Рику повезло: в конце изведанного ранее туннеля они обнаружили продолжение, ведущее в манящие недра земли. И они не зря последовали диггерскому зову – в тупике (который теперь зовется Диналеди) их ждала сенсация. На дне небольшой камеры эффектно лежали чьи-то кости. Стивен и Рик оказались не просто спелеологами, а грамотными и образованными людьми. Они сфотографировали неожиданную находку и принесли фотографии Ли Бергеру, известному во всем мире южноафриканскому палеоантропологу. Несколько лет назад он прославился, открыв в пещере Малапа скелеты Australopithecus sediba. Очевидно, звезда удачи продолжает светить над его головой.
Ли Бергер организовал масштабную экспедицию в Райзинг Стар. Тут-то и обнаружилось, что иногда все же полезно есть мало каши: очутиться на месте сенсации дано не каждому. Только самые мелкие исследователи способны протиснуться в каменные тиски и достичь вожделенных костей.
Но это все присказка. Главное, конечно, не история открытия, а сами окаменелости. И тут сенсация окончательно стала сенсацией (Berger et al., 2015; Dirks et al., 2015).
Во-первых, находок оказалось не просто много, а невероятно много: в камере метр на девять обнаружилось более полутора тысяч фрагментов! По итогу тут нашелся почти целый скелет и останки еще четырнадцати особей – взрослых мужчин, женщин, подростков и детей. Часть костей покоилась в собранном положении. Например, на поверхности красовалась совершенно целая кисть, прямо как в малобюджетных фильмах ужасов. Такого еще не бывало. Во всей Африке за век исследований найдено сопоставимое число окаменелостей. Более того, раскопки пока затронули лишь небольшой участок пола. Сколько еще останков хранит пещера? Трудно себе представить…
Во-вторых, кости лежали просто на полу. Они были лишь слегка припорошены каменной шугой, которая тысячи лет оседала на них со стен и потолка. Всего насыпалось пятнадцать сантиметров осадка. Судя по всему, кости не были никем потревожены, разве что некими неведомыми заблудшими спелеологами, которые были не столь сознательны и никому не сообщили о находке, а только пошебуршили окаменелости. Геологические изыскания привели исследователей к мысли, что пещера всегда была темным закутком, в нее всегда вел лишь тот же “шкуродер” – тесный туннель, стены которого к тому же местами покрыты острыми выступами. В пещере нет никаких следов человеческой деятельности и ни одной кости крупных животных. Найдены лишь шесть птичьих костей и несколько грызунячьих резцов. Нет и отпечатков зубов хищников на останках, нет и следов орудий. Нет и следов переноса водой. Как же так? Ничего тут нет, кроме огромной кучи человеческих костей, попавших в пещеру в собранном виде. Как? Откуда? Осторожные исследователи полагают, что раньше в камеру вел какой-то другой ход, через который люди (а также птицы и мыши) проникали сюда. Но как? Провалились случайно, как в Малапе? И так пятнадцать раз? И при этом ни один зверь крупнее мыши не повторил их печальной судьбы? К тому же состояние осадков говорит за то, что накопление костей происходило некоторое время, оно не было одномоментным. Так что вряд ли сразу целая группа древних людей попала в западню.
Но геологи утверждают, что альтернативного пути вообще нет и не было.
И тут открывается великое поле для сенсационных предположений. Как и зачем люди заползали сюда? Конечно, они были мелкими, в этом смысле им было проще, чем нынешним спелеологам и антропологам, но не все же определяется размерами. Они ползли на ощупь? Или – возможно ли такое?! – первобытные диггеры освещали себе путь факелами? Протискивались ли они в смертельный тупик, чтобы умереть тут спокойно? Или – а такое могло ли доселе прийти кому-то в голову?! – затаскивали трупы почивших соплеменников? Чтобы оценить невероятность сих предположений – не просто смелых, а почти на грани фантастики, – стоит ознакомиться с самими посетителями пещеры, обретшими тут вечный покой.
На этом месте во весь рост встает проблема, прямо вытекающая из вышеуказанного отсутствия в Диналеди чего-либо, окромя человеческих костей: нет датировки. Обычно геологический возраст определяется, исходя из стратиграфии (последовательности осадков), состава фауны и флоры, радиометрическими способами – по вулканическим туфам, обожженным костям или хотя бы песку. А тут нет ничего из этого. Даже песка!
Остается “в-третьих”: морфология. Великое изобилие костей для любого антрополога – солнечный праздник с каруселями и мороженым.
Морфология не подвела. В принципе, в Диналеди могли обнаружиться останки хорошо известных австралопитеков или эректусов. Это было бы тоже здорово, но не так пафосно. А тут все оказалось как надо – сенсационнее некуда. По итогу исследования группа антропологов описала новый вид древних людей – Homo naledi.
Чем же он уникален?
Диналедцы имели маленький рост – около полутора метров. Это не выделяет их из массы австралопитеков и “ранних Homo”. Голова была тоже очень небольшой; мозгов было не то чтобы мало, но и не много – 465 см³ для двух маленьких черепов и 560 см³ для двух больших. Для австралопитеков бóльшая цифра – индивидуальный рекорд, для хабилисов меньшая – тоже, только теперь в минимальную сторону. По итогу диналедцы занимают строго промежуточное положение. Это даже круче, чем в случае с седибой, чей объем – 420 см³, то есть скучно-австралопитековый.

Рис. 17. Череп Homo naledi.
Мозг Homo naledi выглядит весьма примитивно. Лобные доли низкие и короткие, хотя область “зоны Брока” – моторного речевого центра – вроде бы достаточно выпуклая. Особенно дремуче устроена височная доля: она фактически австралопитековая и особо не отличается от обезьяньего варианта. Лобная доля сходится с височной под тупым углом, как у шимпанзе.
Череп при этом не сказать чтобы был чересчур примитивным. Мозговая коробка имеет наибольшую ширину в нижней части, а боковые стенки черепа при взгляде сзади несколько сходятся кверху, но не так уж резко. Относительная высота черепа вполне прилична по меркам “ранних Homo”. Особенно интересно мощное развитие надбровных валиков и, соответственно, сильное заглазничное сужение. Такое строение больше подходит для классических эректусов, а не хабилисов. Впрочем, на женском черепе DH3 надбровные валики сравнительно тонкие. Лоб напрочь скошен, особенно на крупных черепах DH1 и DH2 – самцам и положено иметь брутальный вид. При этом на лобной кости прослеживаются лобные бугры: у обезьян и австралопитеков их не найти, а вот для Homo они вполне типичны. Височные линии выглядят даже прогрессивнее, чем у H. rudolfensis и H. habilis: они довольно слабые и следуют скорее по боковой, а не верхней стороне лобной кости. В задней части они не смыкаются с рельефом височной кости; зато имеется угловой торус – специфический элемент рельефа задней части теменной кости, типичный для эректусов и гейдельбергенсисов, но нехарактерный для “ранних Homo”.
Затылок вполне австралопитековой конфигурации – укороченный и почти вертикальный, но с мощным наружным затылочным бугром, больше подходящим для эректусов.
На височной кости слуховые отверстия и сосцевидные отростки маленькие, примитивные. Впрочем, подавляющая часть тонкостей строения височной кости не отличает новый вид от ранее известных “ранних Homo”.
О форме глазниц и носа остается больше догадываться. Возможно, новые материалы “откроют личико”, но пока оно остается скромно завешенным тафономической вуалью. Впрочем, известно, что у наледи не было “передних лицевых валиков” – утолщений по бокам носового отверстия, типичных для южноафриканских австралопитеков.
Челюсти, очевидно, сильно выступали вперед, особенно резко выражен альвеолярный прогнатизм на верхней челюсти. Передняя сторона нижней челюсти при этом не скошена, а почти вертикальна. Небо было мелким, чем Homo naledi больше похож на австралопитеков и H. habilis, нежели на более продвинутых людей, зато оно же было широким, – а это уже продвинутая черта. Альвеолярная дуга нижней челюсти самой что ни на есть умеренной формы: возможно, и не сапиентной, но точно и не австралопитековой. Удивительно прогрессивно выглядит высокое расположение подбородочного отверстия: таковое типично едва ли не только для современного человека.
Зубы Homo naledi – одна из самых продвинутых его частей. Особенно удивительны маленькие размеры моляров. По ним диналедцы сопоставимы разве что с современным человеком. Они уступают не только всем австралопитекам и “ранним Homo”, но вообще всем гоминидам, включая седиб и “хоббитов”. Таких мелкозубых человечков с таким строением черепа никто найти не ожидал.
Столь же противоречиво и строение посткраниального скелета.
Позвоночник в большей степени напоминает вариант плейстоценовых Homo. Особенное внимание обращает на себя большой диаметр позвоночного канала. Грудная клетка широко расходится книзу – эта черта типична для австралопитеков. Столь же примитивны и лопатки. На плечевых костях почти отсутствует скрученность. Они очень грацильны, со слабо развитым мускульным рельефом, а локтевая с лучевой весьма прямы: вряд ли диналедцы лихо скакали по деревьям.
Кисть Homo naledi, с одной стороны, сохраняет удлиненность и примитивную изогнутость фаланг (даже бóльшую, чем у австралопитеков), с другой – имеет прогрессивное строение запястья и продвинутые пропорции пальцев. Например, большой палец действительно большой. Концевые фаланги расширены – это признак трудовой кисти. Одновременно сустав кости-трапеции и первой пястной маленький, а на третьей пястной нет шиловидного отростка – это примитивные, отнюдь не трудовые черты. Есть и странности, скажем, форма первой пястной кости – с узким основанием и широкой головкой.
Тазовые кости сохранились неидеально, но ясно, что крылья подвздошной кости сильно расходились в стороны, как у австралопитеков. Вместе с тем расстояние от седалищного бугра до вертлужной впадины уменьшено, как у A. sediba и Homo. Суммарное сочетание выглядит необычным.
Вдоль шейки бедренной кости тянутся какие-то странные гребни и бороздка – такого доселе не встречали ни у каких гоминид. Что это – локальная популяционная вариация или видовая специфика? Все образцовые признаки прямохождения: форма, длина и наклон шейки, выраженные ягодичная бугристость и шероховатая линия – развиты по полной программе.
На бугристости большой берцовой кости (Читатель может нащупать ее у себя ниже коленной чашечки) имеется отдельный бугорок для сухожилия полусухожильной мышцы. Мышца эта сводит колени. Диналедцы часто держали пенальти?
Стопа Homo naledi – едва ли не самая человечная его часть: на ней отлично развиты продольный и поперечный своды, нет никакого намека на хватательные способности. При этом пяточный бугор выглядит уменьшенным, не вполне стандартны и некоторые частные пропорции вроде суженности основания первой плюсневой.
В целом мы видим беспрецедентный набор странностей. Успокаивает хотя бы то, что разные индивиды минимально отличаются друг от друга, так что мы можем быть уверены, что все они принадлежат к одному виду, а их особенности действительно имеют видовой или популяционный характер, но не индивидуальный.
Как же в сумме объяснить всю эту мозаику?
В отсутствие датировки может быть несколько вариантов.
Во-первых, Homo naledi может быть действительно предком всех прочих людей. Комбинация примитивных и продвинутых черт запросто объясняется промежуточностью его эволюционного положения, а датировка может располагаться в интервале от 3 до 2 млн лет назад. Однако некоторые признаки наледи слишком прогрессивны. Скажем, трудно представить, чтобы крупные зубы австралопитеков превратились в столь маленькие Homo naledi, а после – опять в большие H. habilis и H. erectus (не говоря уж о мегадонтных H. rudolfensis и H. gautengensis), после чего опять стали уменьшаться. Не слишком ли много вывертов? В этом смысле линия австралопитеки – хабилисы – эректусы выглядит куда более последовательной, тогда как наледи и рудольфенсисы оказываются специализированными боковыми ответвлениями от генеральной линии.
Тут мы переходим к “во– вторых”: Homo naledi может быть экзотической тупиковой ветвью эволюции, наряду с седибой, гари и парантропами. Одних уносило в увеличение зубов, других – в уменьшение. В этом случае Южная Африка представляется как место, где эволюция шла своим чередом, но этим людям не повезло: по совокупности признаков они проиграли гонку своим восточноафриканским родственникам.
Логично и “в-третьих”: можно пофантазировать, что Homo naledi в действительности жили довольно поздно, одновременно с гораздо более продвинутыми гоминидами. Тогда они могли представлять реликтовый пережиток ранней стадии эволюции. В этом случае можно объяснить и мозаичное строение, и предполагаемое знакомство с огнем, и даже существование погребального обряда, хотя бы и крайне примитивного: дескать, подсмотрели у продвинутых кузенов, вот и собезьянничали, насколько позволяли австралопитековые мозги.
А может, правы скептики и люди из Диналеди – это просто те же самые хабилисы, только мы наконец-то видим их собранный скелет? Ведь до сих пор такой радости мы были лишены. Даже принадлежность кисти и стопы OH 7 и OH 8 – голотипов вида Homo habilis – к виду Homo habilis, как ни парадоксально, остается недоказанной. А вдруг OH 8 – это стопа бойсовского парантропа? Для подтверждения или опровержения нужны полные скелеты хабилиса и парантропа, а у нас нет ни того ни другого. А в Диналеди мы можем иметь пусть своеобразно-мелкозубого и мелкоголового, но все же просто хабилиса. Или это вариант того же седибы? Ведь он тоже сочетает признаки австралопитеков и хабилисов, пусть и в других комбинациях. Кстати, седиба также знаменит именно своей мозаичностью, он тоже весьма малоголов и мелкозуб, а по разным признакам таза, рук и ног может быть равно отнесен к австралопитекам или людям.
Все же комплекс черт Homo naledi выглядит достаточно уникальным, чтобы признать его самостоятельным видом.
Еще не все кости из Диналеди подняты на поверхность. Еще не все их признаки описаны и осмыслены. Еще многое будет о них сказано. Одно точно: наша генеалогия в очередной раз оказалась интереснее и богаче, а при рассказе о роде “Люди” отныне нельзя не вспоминать о “Человеке наледи”.
Руки, ноги… Главное – наледи!
Не прошло и месяца с торжественного вступления Homo naledi в статус официального древнейшего человека, как вышли две большие статьи с подробным описанием их рук и ног.Руки и ноги – очень важные части человека. Строение стопы очевидным образом отражает способности к разным типам передвижения. Строение кисти – к лазанию по деревьям или трудовой деятельности. Неопределенность датировки наледи, отсутствие орудий и загадочность попадания костей в пещеру позволяют богато фантазировать об этих существах. Но на то и наука, чтобы не фантазировать, а устанавливать твердые факты.Первая статья посвящена строению кисти Homo naledi (Kivell et al., 2015). Кисть сохранилась идеально – имеются все косточки, исключая лишь гороховидную, чего еще не бывало для находок мало-мальски приличной древности. Более того – в отложениях она лежала в полностью собранном виде, что избавило исследователей от тягостных раздумий о количестве индивидов. От других индивидов осталось меньше, но тоже не так уж мало – всяко больше, чем от всех хабилисов, известных доселе.Кисть отличается крупным и толстым большим пальцем и широким трудовым запястьем, что позволяет утверждать, что наледцы пользовались руками для изготовления орудий. Одновременно пальцы с указательного по мизинец длинные и изогнуты сильнее, чем у большинства австралопитеков, что явно указывает на отличные способности к древолазанию.Кроме того, кисть наледца очень маленькая: миниатюрнее ее только малапец MH2 и самые мелкие современные люди. Пропорции уникальны: большой палец относительно третьего просто огромный, по этому параметру с наледцем сближаются опять же только особо одаренные современные люди, Кафзех IX (который тоже почти сапиенс), а превосходит лишь все тот же малапец MH2.Кости запястья почти не отличаются от современных. Существенно, что они же заметно разнятся от австралопитековых, “хоббичьих” и даже неандертальских. Фактически если бы в пещере нашли только запястье, то его без долгих сомнений определили бы как человеческое.Крайне необычна форма первой пястной кости: ее основание сужено, а тело и головка расширены. На теле к тому же имеются мощные гребни для мускулатуры. Что такое делали наледцы своими мускулистыми большими пальцами? Рыли вход в Райзинг Стар? Гиппарионов на скаку останавливали? Писали слишком много эсэмэсок по сотовому? Такое строение совершенно не похоже ни на каких человекообразных обезьян, ни на людей. Причем костей в Райзинг Стар найдено немало – аж семь штук, и все они одинаковы. То есть это точно видовой признак, а не индивидуальная особенность.Видимо, связанный признак – утолщенная пятая пястная кость. Можно говорить о латерализации кисти Homo naledi – усилении ее по краям, что имеет смысл, если кисть держит что-то очень тяжелое.Однако на основании третьей пястной кости нет шиловидного отростка. Этот отросток отличает современных людей и горилл от шимпанзе и австралопитеков (включая седибу). Он входит между головчатой и трапециевидной костями запястья и соединяется с похожим выступом на основании второй пястной. За счет этого кисть становится прочнее и может выдерживать большие нагрузки. Для гориллы это актуально, потому что она сама очень тяжелая и надо опираться на кулаки. Человек постоянно что-то сжимает в руках, его средний палец не должен бесконтрольно болтаться в стороны. Древесные же обезьяны и австралопитеки-бездельники могут свободно шевелить пальчиками; на деревьях это даже весьма актуально – при быстром перехвате веток кисть должна легко перекашиваться набок и никакие шиловидные отростки не должны этому мешать.А вот фаланги Homo naledi оказались не то крайне примитивными, не то крайне специализированными. Изгиб первых фаланг такой же, как у австралопитеков, хабилиса OH 7, да, в общем-то, и шимпанзе, а вот средних – вообще как у гиббонов, сильнее, чем даже в среднем у орангутанов, не говоря уж о шимпанзе, гориллах и австралопитеках с людьми. Тут, правда, стоит оговориться, что изгиб посчитать не так легко и сделать это можно по-разному. Но как ни измеряй, странность Homo naledi все равно очевидна.Как объяснить сии сочетания? Насколько они парадоксальны или же логичны? Древесные черты сочетаются у Homo naledi с трудовыми и пересыпаны специализированными. А может, если задуматься, все так и должно быть у первых людей, еще не до конца слезших с деревьев, но уже начавших делать орудия труда? Спасаться от хищников на ветвях все же здорово – для этого пальцы изогнуты, а третья пястная подвижна. Но необходимость крепко держать орудия труда вызывает усиление запястья, первой и пятой пястных костей. Более того, для компенсации слабости третьего луча первый и пятый усилены даже больше, чем у современных людей.Не может не обратить на себя внимание сходство по ряду признаков кисти Homo naledi и Australopithecus sediba. По общим размерам и пропорциям они фактически идентичны. Но вот детали строения запястья у седиб выраженно австралопитековые, а у наледи – человеческие. Учитывая датировку седибы в 2 млн лет и наличие шиловидного отростка на пястной кости KNM-WT 51260 из Каитио с датировкой 1,42 млн лет назад, можно предположить датировку Homo naledi между 2 и 1,5 млн лет назад.Несколько иная картинка вырисовывается при рассмотрении стопы наледцев (Harcourt-Smith et al., 2015). Она сохранилась чуть менее идеально, нежели кисть, но тоже отлично в сравнении с другими ископаемыми предшественниками человека. Всего откопано 107 костей, в том числе одна почти целая стопа.Почти по всем параметрам ножки Homo naledi неотличимы от современных. Общие пропорции, неоттопыренность большого пальца, продольный и поперечный своды – все как у людей. Впрочем, и тут внимательный глаз морфолога найдет специфику: таранная кость при взгляде сбоку все же скорее австралопитеко– и шимпанзеподобная, фаланги пальцев кривоваты, опора таранной кости на пяточной кости ориентирована совершенно не так, как у людей, хотя и не как у шимпанзе, зато идентично австралопитекам и гориллам. В итоге ходить наледцы должны были чуточку не как мы, хотя и совсем отлично от обезьян. Все же от древесного прошлого их отделяло меньше времени, да и сами они при случае запросто могли вскарабкаться по сучьям подальше от зубастых врагов.Руки и ноги составляют два из трех важнейших человеческих комплексов “гоминидной триады” – черт, отличающих человека от обезьяны. Третий комплекс – мозг. Очевидно, вкусное исследователи приберегли на десерт. Судя по первой публикации, мозг наледи очень примитивен. Очевидно, тут стоит ждать еще много любопытных поворотов.
Что творилось в Южной Африке два миллиона лет назад? Новые находки в Стеркфонтейне
Происхождение человеческого рода Homo никогда не перестанет волновать умы антропологов. За последние сто лет стало окончательно ясно, что родиной была Африка. Но Африка велика. А стран в ней много. Много и антропологов с самыми разными идеями. С описания первых австралопитеков в Южной Африке появилось мнение, что именно этот регион был горнилом гоминизации – процесса очеловечивания. Открытия в Восточной Африке сместили внимание специалистов туда. Некоторые задумываются и о Центральной Африке, и Северной, и Западной. XXI век принес свои новости.Обнаружение Australopithecus sediba в Малапе с датировкой 2 млн лет и Homo naledi в Райзинг Стар снова нагрузило южноафриканскую чашу весов. Конечно, не могло обойтись без странностей и споров – и седибы, и наледи имеют необычайное сочетание признаков: маленькие мозги совмещаются с маленькими зубами, продвинутые пропорции пальцев – c их чрезвычайной изогнутостью, длиннющие руки – со вполне человеческими стопами. Такую кутерьму можно интерпретировать всяко. Часть антропологов, особенно южноафриканских, склонны считать, что седибы и наледи – прямые предки рода Homo, просто еще не избавившиеся от некоторой дремучести. Другие мнят их весьма специализированной ветвью, сформировавшейся в изоляции на краю Земли, в отрыве от центра прогресса. Как обычно, решение – в новых находках и новых исследованиях.И вот они – новые находки!Во всемирно известной пещере Стеркфонтейн найдены важные окаменелости (Stratford et al., 2016). До сих пор в Стеркфонтейне обнаруживались в основном кости Australopithecus africanus, немного было парантропов и “ранних Homo”. Две новые находки откопаны в Милнер-Холл – центральной части пещеры, в слое, имеющем предположительный возраст около 2,18 млн лет назад, то есть несколько более древний, чем в Малапе. Такая датировка – уже хорошая новость, ведь известно не так много гоминид в интервале от 3 до 2 млн лет. И самое важное – морфология.Зуб Stw 669 – правый верхний первый постоянный моляр – радует глаз своими маленькими размерами, в самых нижних пределах изменчивости Homo habilis и большими, чем у Homo naledi. То же выявляется при анализе площади основных бугорков, если их измерять по отдельности, а совокупность общих размеров и площади паракона и метакона делает Stw 669 форпостом прогресса среди всех “ранних Homo”.Проксимальная фаланга кисти Stw 668, напротив, чрезвычайно велика. Таких огромных фаланг нет среди “ранних Homo”, имевших вообще мелкие ручки. Длина Stw 668 превосходит длину фаланг современных людей и даже некоторых шимпанзе и горилл, толщина – больше, чем у большинства современных людей и шимпанзе, и приближается к горилльим значениям. Замечателен и изгиб: он практически равен шимпанзиному и бонобьему, больше средней горилл, выходит за границы изменчивости всех древних и современных людей, включая седиб и наледи. Даже грацильные австралопитеки имели не столь кривые пальцы. Лишь две фаланги Australopithecus afarensis из Хадара изогнутее.Как же это понимать? Конечно, первое, что приходит в голову, – аналогичное сочетание маленьких зубов и кривых пальцев у седиб и наледи. Более того, морфология строго соответствует хронологии: новые останки из Стеркфонтейна самые древние и самые примитивные, седибы из Малапы чуть моложе и, что логично, чуть продвинутее, а наледи из Райзинг Стар – самые мелкозубые и чуть менее кривопалые. И все это в сравнительно небольшой области Южной Африки. Более наглядную эволюционную линию надо еще поискать!Но! Конечно, есть проблемы. Ведь на фоне других гоминид эта стройная линия выглядит стоящей совсем особняком. Таких мелкозубых и кривопалых гоминид нет ни в Восточной Африке, ни – что совсем интересно – в других частях Южной. Ведь описанные несколько ранее южноафриканские Homo gautengensis имели огромные зубы и куда как более короткие и прямые фаланги. Что любопытно, один из черепов Homo gautengensis найден в том же самом Стеркфонтейне…Таким образом, пока картина рисуется таким образом: в области, прилегающей к нынешнему Йоханнесбургу, со времени как минимум 2,18 млн лет назад шел эндемичный процесс: гоминиды стахановскими темпами теряли размеры зубов, но сохраняли крайне архаичные пальцы. Что ставило их особняком – пока загадка. Может, ответ дадут более тщательные палеоэкологические реконструкции? В других областях параллельно возникали свои варианты людей. Думается, все же именно эти прочие варианты стали нами.Слишком же странные люди Южной Африки сохранились лишь в виде окаменелостей – антропологам на радость, любопытным на удивление и задумчивым в назидание.
Выход из Африки: № 1
Около 2 млн лет назад свершилось великое – люди вышли из своей колыбели и побрели по просторам Евразии.
Иногда при обсуждении этой темы звучит сакральная фраза: “Люди в своей эволюции достигли нового уровня развития, который позволил им покинуть пределы Африки и освоить новые просторы”. Однако представляется, что все было не совсем так и дело обошлось без великих достижений. Естественно, миграция за пределы Африки не была хотя бы минимально осмысленной. Не было пафосного момента, когда бы этакий первобытный Колумб эффектно встал на пороге Евразии, картинно приложил руку к глазам, орлиным взором обозрел открывшиеся бескрайние просторы, залитые золотыми лучами восходящего солнца, и широким жестом указал оробевшим и притихшим сородичам, сбившимся за его спиной тесной кучкой: “В путь, друзья! Откроем же Евразию! Нас ждут великие свершения! Весь мир у наших ног! Мы пройдем его и заселим бесчисленными потомками, мы устремимся за горы и океаны, мы покорим космос, мы будем бороздить…” Нет, такого момента, к сожалению, не было. Не исключено, что люди даже не заметили, что находятся уже на соседнем материке, и уж точно не оценили величия момента. Собственно, событие это было рядовое, и даже не факт, что оно заслуживает описания в отдельной главке, но традиция обязывает.
Из предыдущих глав Читатель уже знает, что многие и многие приматы за миллионы лет до этого благополучно вселялись и выселялись с Черного континента и прекрасно обходились без особо продвинутых уровней развития. Более того, фауна Аравии и Африканского Рога до крайности похожа (в недавние времена в Аравии многие виды животных были истреблены, так что нынешние различия во многом искусственны). Страусы, леопарды, львы, гепарды, павианы, слоны, ослы и прочие звери преодолевали грань между континентами. Человек был частью той же фауны и мигрировал вместе с ней. Леопарды без всякой культуры заселили всю Африку, Азию, Европу и даже переплыли как-то в Америку, заделавшись там ягуарами. Без огня, языка, мифологии и прочих человеческих наворотов. А львы поначалу гораздо успешнее заселили северные части Евразии, чем люди, хотя люди их потом там и извели.
Скорее уж надо удивляться, что выход гоминид из Африки не свершился намного раньше, непонятно, что мешало сделать это австралопитекам.
Африку теоретически можно покинуть тремя путями: через Гибралтар, по Суэцкому перешейку или переплыв Баб-эль-Мандебский пролив. Гибралтар преодолеть нелегко, это самый маловероятный маршрут. Египет, Суэцкий перешеек и Синай большую часть времени были жаркими негостеприимными пустынями, не вполне понятно, как бы там могли жить охотники-собиратели. Однако климат менялся, и в какие-то моменты территория могла быть вполне проходимой. А Баб-эль-Мандебский пролив в периоды понижения уровня моря становился сравнительно узким – в пару километров шириной – и условно мелким, хотя и не настолько, чтобы можно было перебрести его вброд. Но с пляжей Джибути видны горы Йемена, так что африканским жителям было очевидно, что на востоке есть новые земли. А самое главное – экологические условия по обе стороны пролива ничем особо не отличаются. Переправившимся странникам не надо было привыкать и хоть как-то приспосабливаться, они отряхнулись и стали жить прежней жизнью прибрежных собирателей.
Так или иначе, люди переправились в Евразию. Возникает вопрос: когда?
Расселение людей за пределами Африки, к сожалению, в основном известно лишь по археологическим памятникам, не содержащим останков людей. К еще большему сожалению, почти все древнейшие стоянки чем-то, да неидеальны. То стратиграфия спорная, то орудия не факт что орудия, то датировки неточны.
Древнейшим вероятным внеафриканским местопребыванием была, разумеется, Земля обетованная. В Израиле раскопана стоянка Йир'он, слои которой датированы аж 2,4 млн лет назад или даже бóльшим временем (Brunnacker et al., 1989; Ronen, 1991), хотя далеко не все исследователи согласны как с цифрой, так и с определением найденных камней как орудий. Гораздо лучше изучена палестинская многослойная стоянка Эрк-эль-Ахмар, древнейшие фазы заселения которой относятся ко временам около 1,8 млн лет назад (Verosub et Tchernov, 1991; Bar-Yosef, 1995).
С Ближнего Востока путь раздваивался: можно было идти на север или двигаться на восток, навстречу восходу. Первое направление многим может показаться нелогичным: “Как же так, зачем же они шли в холода и полярную ночь?!” – воскликнет иной недоумевающий читатель. Однако те люди ничего не знали о морозах и льдах. Каждая перекочевка – на несколько километров – не изменяла ничего в их окружении. Только спустя сотни и тысячи лет потомки оказывались действительно в новой климатической зоне, но для каждого конкретного поколения изменений вроде бы и не было. А уж около 2 млн лет назад – да и намного позже, вплоть до 400 тыс. лет, – тропические по происхождению люди были еще не настолько оторваны от природы, чтобы освоить зоны даже с умеренным климатом, максимум, куда они совались, – это субтропики.
Северный маршрут упирался в очередные водные преграды – Босфор и пролив, соединявший Черное море с Каспийским, лежавший на месте нынешней Кубани. На краю тогдашней Ойкумены люди оставили свои следы: орудия с датировками 1,6–1,2 млн лет назад найдены на стоянках Богатыри / Синяя Балка и Родники на Тамани (Shchelinsky et al., 2010).
Восточный путь отмечен галечными орудиями на пакистанских стоянках Риват и Пабби Хиллз, датированных временем около 1,9 млн лет назад (Dennell et Hailwood, 1988). Дальше на восток огромные цифры назывались для китайских местонахождений Жензидун (2,25 млн лет назад; Zhang et al., 2000), Лунгупо (1,9 млн лет назад), Сяочанлянь (1,67–1,87 млн лет назад), Сихудю (1,8 млн лет назад), Данау, или Шангнабанг, в Юаньмоу (1,7 млн лет назад) и других. Но во всех этих случаях либо есть сомнения в датировках, либо подозрительны артефакты. Более достоверны находки и даты в Маджуангоу в долине Нихевань: 1,66 млн лет назад для древнейших слоев с орудиями и 1,32 млн лет назад – для самых молодых (Zhu et al., 2004).
Но все эти стоянки мало привлекают антропологов, ведь там нет костей. Другое дело – Дманиси!..
Странные Homo Грузии: Homo georgicus
Наконец, эогоминины известны и за пределами Африки. Конечно, речь идет о находках в Дманиси. Их датировка постоянно обсуждается, но по последним расчетам первое заселение совершилось около 1,78–1,85 млн лет назад (Ferring et al., 2011), тогда как кости людей могут иметь несколько более поздний возраст. Для человеческих останков наиболее достоверной обычно считается цифра 1,77 млн лет назад, но, поскольку они залегают в трещинах, иногда датируются заметно более поздними временами – до 1,07 млн лет назад (Calvo-Rathert et al., 2008; Pares et Goguitchaichvili, 2001). Состав фауны не противоречит обеим версиям. Кроме того, скелеты разных индивидов могут быть несинхронны: D2700/2735 залегает выше, чем D3444/3900 и D2600/4500. Впрочем, удивительная комплектность скелетов, а также компактное расположение трех из них в одном месте скорее отвечают версии о более-менее одновременном захоронении (конечно, непредумышленном). Найдены останки как минимум пяти индивидов. Замечательно, что в Дманиси мы имеем хороший набор полов и возрастов: череп D2700+D2735 принадлежал подростку, вероятно девушке, D2282+D211 – очень молодой, на грани взрослости, особи, возможно женщине, D2280 – молодому взрослому мужчине, D4500+D2600 – взрослому мужчине в полном расцвете сил, а D3444+D3900 – старику или, быть может, старушке.
Как это обычно бывает с гоминидами “загадочного миллиона”, признаки дманисцев мозаичны. По размерам черепа дманисцы попадают ровно в зазор между хабилисами и эргастерами, в наибольшей степени соответствуя KNM-ER 1470. У этого рудольфенсиса лишь чуть меньше мозговая коробка и чуть больше лицо (что соответствует и большей датировке), но он укладывается в пределы изменчивости дманисцев. Размеры зубов дманисцев тоже находятся в рамках вариаций рудольфенсисов, хотя меньше их средней и, тем более, таких рекордсменов, как UR 501 или, того пуще, парантропов.
Череп D4500 очень похож на прогнатную реконструкцию KNM-ER 1470 и, в ряде аспектов, на KNM-ER 1805, в сумме представляя что-то среднее между ними. Нижняя челюсть D2600 метрически и внешне весьма близка к KNM-ER 1482, KNM-ER 1801, KNM-ER 1802 и KNM-ER 60000, а также, хотя и несколько меньше, к UR 501. Невозможно пройти мимо необычайного внешнего сходства D2700+D2735, KNM-ER 1813 и KNM-ER 62000, которое подтверждается и исследованием многих деталей. Несмотря на то, что дманисцы часто диагностируются как эргастеры, по всем параметрам они примитивнее, чем, например, KNM-ER 3733. От классических же эректусов, к которым относили дманисцев поначалу, отличия уж слишком велики.
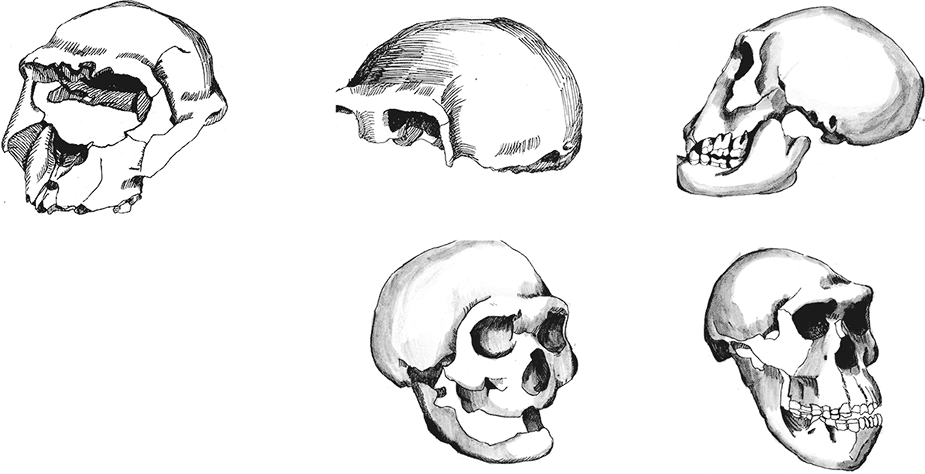
Рис. 18. Пять черепов Дманиси.
Особый интерес представляет сочетание самого массивного лица и самого маленького мозга – 546 см³ – у D4500+D2600. Это фактически максимум для афарских австралопитеков и близко к минимуму для эогоминин. Однако разница с самым “мозговитым” дманисцем D2280 (730 см³) составляет 184 см³, то есть аналогично индивидуальному разбросу у орангутанов, примерно вдвое меньше, чем у горилл, едва больше, чем у шимпанзе, на 100 с лишним кубических сантиметров меньше, чем среди мужчин-синантропов, и в два или два с половиной раза меньше, чем у современного человека (даже без учета рекордов). Таким образом, по такому важному показателю дманисцы вполне укладываются в пределы нормальной изменчивости самых разных человекообразных. То же касается их челюстей и прочих признаков.
Разница конкретных находок из Дманиси, конечно, не могла не породить гипотез о существовании тут нескольких видов гоминид, благодаря чему челюсть D2600 получила собственное наименование Homo georgicus. Однако эти же различия можно интерпретировать и иначе – как половозрастные и индивидуальные. Дманисцы очень здорово заполняют промежутки между такими резко различными находками, как KNM-ER 1470, KNM-ER 1805, KNM-ER 1813, OH 24, SK 80/846/847, KNM-WT 15000, KNM-ER 3733 и даже Сангиран 17. Крайняя точка зрения – сведение вообще всех гоминид “загадочного миллиона” и даже некоторых за его пределами в один полиморфный вид с сильнейшим половым диморфизмом и выраженной возрастной вариабельностью, причем эти аспекты изменчивости накладываются на плавную эволюцию во времени (Lordkipanidze et al., 2013). Дманисцы в такой системе являются своего рода эталоном и примером внутрипопуляционного масштаба.
Данные по половому диморфизму дманисцев позволяют по-новому взглянуть на эту проблему в отношении африканских гоминид. Например, спорный череп KNM-ER 42700 в принципе может быть рассмотрен как молодая самка архантропа (Spoor et al., 2008). Правда, половой диморфизм африканских Homo erectus тогда получается очень сильным, почти как у горилл, и, в общем-то, даже бóльшим, чем у дманисцев. Очевидно, все же не стоит забывать и о хронологических преобразованиях.
Грузинские гоминиды задали немало загадок. Сначала они считались эректусами, потом эргастерами, после был описан самостоятельный вид, и, наконец, те же авторы сошлись на том, что дманисское разнообразие позволяет объединить вообще всех “ранних Homo” вместе с архантропами воедино, а их различия полностью объяснимы половыми и возрастными особенностями. Думается, крайности излишни, но где лежит “золотая середина” – вопрос.
Споры о систематике не закончатся, пока на свете не останется меньше двух систематиков. Даже введение молекулярной систематики, казалось бы – супермегаобъективной и математически проверяемой, не избавило научный мир от споров, а разожгло их лишь с новой силой. Что уж говорить о систематике ископаемых гоминид, от которых остались окаменевшие кости. Главной проблемой любой из спорящих сторон является отсутствие универсального критерия выделения вида. Что, собственно, понимается под палеонтологическим видом? Единство морфологии? Способность к скрещиванию? Вечна проблема масштаба. И никто не победит в этом споре. В некоторых же случаях, как кажется, спор разгорается скорее из спортивного интереса, чем из необходимости. И случай с Дманиси, вероятно, именно таков.
На пятачке с небольшую квартиру найдены останки пяти гоминид. Попали они сюда едва позже 2 млн лет назад и являются древнейшими за пределами Африки. В самой Африке в это время достоверно жили лишь два вида гоминид – Paranthropus robustus и какой-то из “ранних Homo”. В зависимости от предпочтений конкретного антрополога можно называть его Homo rudolfensis, Homo habilis, Homo ergaster или даже Homo erectus, однако ж достоверное сосуществование двух хороших, надежных видов Homo в Африке не доказано никем ни для одного местонахождения. Про отнесение дманисцев к парантропам речь не идет. Остается два варианта: либо дманисцы принадлежат к синхронному африканскому виду, либо по пути успели дифференцироваться в свой особенный. Третий же вариант – что в Дманиси мы имеем представителей двух видов – выглядит довольно-таки фантастично. Просто из хронологических и экологических соображений. Нет, он не вовсе невероятен, но ежели даже в благословенной Африке во множестве местонахождений нет доказанных случаев сосуществования двух видов Homo, то появление их в однокомнатной квартире в уединенной Грузии, мягко говоря, странновато…
Вопрос о названии грузинских людей многим может казаться каким-то очень принципиальным, но так ли это? Объективно мы не можем проверить, могли ли дманисцы скрещиваться с африканскими современниками. Учитывая пластичность, типичную для приматов, наверняка могли, но доказать мы не можем ни “да”, ни “нет”. Остается морфологический критерий: насколько сильно дманисцы отличаются от африканцев? А тут все зависит от того, какие признаки мы возьмем, каким анализом обработаем и как результаты проинтерпретируем. То есть, в отсутствие надежной универсальной методики, можно переливать из пустого в порожнее сколько угодно. С этого места начинается спор о словах. Эти размеры совпадают – значит, один вид. Нет, в этом признаке видим расхождение – разные виды. Но кто сказал, что отличие именно по вот этому показателю не давало возможности дманисцам скрещиваться с африканцами? Да никто! А то ведь недолго по пяти черепам описать и шесть видов… А еще ведь есть кости скелетов – уже и десяток видов обеспечен! Здравствуй, типологизм XIX века! Кстати, абсолютно аналогичный спор разгорелся в последнее время по поводу малапских Australopithecus sediba: кое-кто предположил, что челюсти и скелеты относятся к разным видам, даже несмотря на то, что скелеты артикулированные!
Нельзя забывать, что систематика создана людьми и призвана помочь нам в упорядочивании информации о живых организмах, а не создавать сложности. Должно быть удобство, а великой истины в названиях древних организмов в общем-то нет. Систематика – это инструмент, а не цель.
Из всего вышеизложенного кто-то может сделать вывод, что все эти изыскания – сплошная демагогия, лишенная смысла. Но это, как ни странно, не так. Во-первых, такие споры активизируют разработку новых подходов и более детальное изучение окаменелостей. Во-вторых, проблема принципиально все же решаема. Надо только лучше и больше использовать материалы по современным людям и обезьянам. Ведь у нас есть универсальный масштаб внутривидовой, видовой, родовой и семейственной изменчивости человекообразных: два вида шимпанзе, гориллы, орангутаны, десяток гиббонов (все обезьяны – с подвидами), куча рас современных людей. Все карты на руках, надо только ими пользоваться. Карту рас западные антропологи разыграть неспособны в силу политкорректности, но обезьян-то пока никто исследовать не запрещает!
Важно, что находки в Дманиси позволяют с новой стороны взглянуть на становление рода Homo, дают новые уникальные данные, будят научную мысль, провоцируют дальнейшие исследования. Хорошо б еще без фанатизма…
Пятый череп из Дманиси: первые люди – на лицо ужасные, добрые внутри
Грузия – хорошая страна! Высокие горы, прозрачный воздух, гостеприимные люди. И уж особенно мила Грузия сердцу всякого антрополога. Плодородная земля горной страны не только родит виноград и чай, но и бережно хранит в себе древние кости. Что уж за счастливые причины тому виной, но сохранность останков в Дманиси просто поражает: даже хваленая Африка в подметки не годится этому местонахождению по части идеальности находок.Не уверен, знали ли авторы новой статьи и члены редакции Science о существовании портала www.antropogenez.ru, но по факту они сделали хороший подарок к его трехлетию и здорово подгадали к очередному заседанию “Клуба Antropogenez.ru”. Хочется верить, что они вняли страстному призыву нашего редактора, прозвучавшему за пару дней до издания статьи (краткое содержание: “Доколе?!”), и опубликовали-таки пятый череп!Об этом черепе давно ходили смутные слухи. В телевизионных ли новостях мелькнут интригующие кадры доставания чего-то серого из земли, в докладе ли баронессы де Люмлей на слайде появятся загадочные и будоражащие воображение цифры “пять черепов от четырех индивидов”, сорока ли протрещит в кустах… Слухами земля полнится, все знали о пятом черепе. Но никто не видел его. Череп был найден еще в 2005 г., а вот уж кончался 2013-й. Трудно сказать, чем была вызвана такая задержка в публикации: не то секретность (“учОные скрывают?!”), не то авторы открытия решили взять пример с исследователей Арди (тогда могли бы еще годиков пять подождать), не то коробка в хранении затерялась (и не на восемь лет теряли), не то бурная общественно-политическая жизнь Грузии внесла коррективы в ход научного прогресса. Много еще может быть причин. Нам сие неведомо, да оно и не важно, главное, факт остается фактом – открытие состоялось!Под торжественные звуки фанфар и барабанную дробь перед изумленными взглядами заинтригованной публики предстало…??? Да что же это?! Из мглы веков и глубины грузинской земли выглянула, не постесняюсь этого слова, страшная морда невиданной конструкции и несказанной первобытности (Lordkipanidze et al., 2013). Плоский лоб (есть ли он?), огромное надбровье (Брокен-Хилл и Бодо нахмурились пуще, но, что называется, нервно курят в сторонке), гигантские челюсти, торчащие вперед (парантропы угрюмо потупились), махонький мозг 546 см³ (горилла довольно скалит зубы и торжествующе бьет себя в мохнатую грудь). Приплюснутый свод, расширенный снизу, – в лучших традициях австралопитеков и самых дремучих питекантропов. Огромные скулы: рельефные, расставленные в стороны, продолжающиеся в огромные гребни на височной кости, – далеко ли до афарских австралопитеков? Позадиглазничное сужение – просто праздник какой-то! А уж если присоединить нижнюю челюсть – конечно, ту самую, самую огромную и страшную D2600, послужившую голотипом вида Homo georgicus, – то облик дманисца выходит поистине первобытным, какому может позавидовать даже Рон Перлман.Вообще, уже по экзотической форме и гигантским размерам этой челюсти было очевидно, что череп ее владельца должен быть по меньшей мере необычным, но гадать антропологи не любят, а потому мысли на этот счет в головах бродили, но на страницы не выплескивались. Да и дманисская девушка (второй череп D2282/D211) тоже не самая грацильная красавица: каких мужчин ждать в приложение к таким девушкам? И вот ожидания оправдались сверх меры. Как же интерпретировать такое чудо?Черепа из Дманиси благодаря удивительной сохранности и различиям пола и возраста позволяют оценить не только общее положение в эволюционной последовательности, но и разобраться с некоторыми любопытными деталями. Например, незадолго до публикации пятого черепа вышла статья, в которой различия нижних челюстей дманисцев хорошо были интерпретированы как производные возраста, стирания зубов и их смещения вдоль альвеолярной дуги. Можно ли повторить подобное объяснение и по отношению к черепам? Если сделать это, то выводы получаются неожиданными. Изменение морфологических признаков с возрастом у дманисцев происходило по модели, скорее типичной для современных шимпанзе, а не людей. У шимпанзе милый маленький малыш с круглой головенкой и маленьким милым личиком за десяток-другой лет превращается в огромное насупленное чудище с полкой над глазами, здоровенными челюстями, украшенными мощными клыками, у которого мозговой отдел как-то теряется за раздавшимся во все стороны лицом. Доселе считалось, что наша линия как раз тем и прогрессивна, что взрослые люди сохраняют многие детские черты, на этом в свое время была основана даже целая гипотеза неотенического происхождения человека. И вот дманисец правит нам эту благостную картину: оказывается, еще на стадии хабилисов – эргастеров возрастные изменения были скорее обезьяньими. Конечно, некоторые мысли на сей счет неоднократно выдвигались. Есть немало находок детенышей и взрослых австралопитеков разных видов, KNM-WT 15000 и KNM-ER 3733, Моджокерто и Сангиран. Разброс их детских и взрослых вариантов явно превышает современные значения. Но доныне подобные возрастные пары были сборными, то есть происходили из разных мест и имели не вполне одинаковые датировки. А в Дманиси мы имеем пять черепов, наиболее вероятно относящихся к одной синхронной популяции, да еще идеальной сохранности, да еще всех возрастов от подросткового до вполне себе старческого, да еще и разного пола для полноты картины. И взрослый мужчина в самом расцвете сил у дманисцев – это уж настоящий мужчина! Его отличия от подростков едва ли не больше тех, что мы видим у нынешних шимпанзе, и намного превышают сапиентный возрастной разброс.Любопытно, насколько онтогенетическое сходство с шимпанзе отражает поведенческую сторону? Крупные размеры лица – явный признак иерархичности и значительной разницы полов. Однако клыки у дманисцев весьма скромные, и в целом монструозный пятый череп D4500/D2600 по этому признаку совершенно не выделяется из общего гоминидного ряда. Так значит, в душе они были добрые и отзывчивые? Подтверждением этому служит четвертый череп D3444/D3900 – беззубого старичка, требовавшего заботы и пережеванной мягкосердечными сородичами кашки. Да и орудия, в изобилии обнаруженные в Дманиси, делать было непросто. Нужно было учиться и учить, а с нервозностью и непоседливостью шимпанзе такой уровень недостижим.Показательно, что на общем эволюционном фоне – от шимпанзе и австралопитеков до сапиенсов – дманисцы занимают весьма широкое поле. На графике многомерного анализа распределение конкретных черепов прелюбопытнейшее: пятый череп (взрослого мужчины-самца) располагается заметно ближе к загадочному черепу KNM-ER 1805 и южноафриканскому грацильному австралопитеку Sts 5, чем к эргастерам. Дманисская молодежь – черепа D2282 и D2700/D2735 – оказывается идентичной хабилисам (KNM-ER 1813 и, в меньшей степени, OH 24), рудольфенсисам (KNM-ER 1470), эргастерам (KNM-ER 3733 и, в меньшей степени, KNM-WT 15000) и питекантропам (Сангиран 17). Крепкий старик – четвертый череп D3444/D3900, – потерявший при жизни все зубы, а вместе с ними и альвеолярные края обеих челюстей, а потому несколько похожий на Бабу-ягу из мультика про пионера Ивашку, оказывается самым “сапиентным” и заметно ближе к неандертальцам и Брокен-Хиллу. Вот уж воистину – старость облагородила его черты!Такая картинка порождает множество замечательных идей.Во-первых, лишний раз становится ясно, что не стоит чересчур увлекаться конкретными признаками конкретных находок. Широкий взгляд на вещи дает более правдивый образ действительности. А всякие мелочи запросто могут быть преходящими – возрастными, например. Внутригрупповая же изменчивость у древних гоминид была не меньше нашей, а то и больше. Нельзя зацикливаться на тонкостях взаимного расположения точечек на графиках, хоть бы и многомерных анализов. То или иное соотношение этих точек может быть обусловлено сиюминутным состоянием отдельных индивидов, а не глобальным эволюционным процессом. Популяционный подход, давно победивший в расоведении, должен уже занять достойное место и в антропогенезе.Во-вторых, может, мы просто до сих пор мало видели по-настоящему взрослых мужчин эргастеров? Может, они просто редко доживали до самого эффектного возраста? Или были столь на лицо ужасные, что злые леопарды с гиенами далеко обходили их стороной, а потому мы не находим их останков? А рудольфенсисы – в который уж раз – не самцы ли хабилисов? И доселе таинственные черепа KNM-ER 1805 и SK 80/846/847 – необычные как раз сочетанием огромных прогнатных челюстей и даже сагиттальных гребней и более-менее крупного мозга – не самцы ли все тех же хабилисов-эргастеров? Не забыты и пресловутые мегантропы – это ли не настоящие взрослые мужчины эректусов? А некоторые мужчины-самцы эргастеров – вернее, считавшиеся доселе самцами – не взрослые ли женщины-самки? Например, пол черепа KNM-ER 3733 находится под вопросом, но версия о его женской принадлежности озвучивалась не раз, а с учетом дманисских находок она предельно логична. В этом случае дманисцы оказываются не столь уж архаичными, как казалось до сих пор; они занимают закономерное место эргастеров, более прогрессивных, чем хабилисы, но примитивнее эректусов.Особенно близка эта мысль, конечно, исследователям Дманиси. Ведь одно дело, когда изучаешь тупиковую уклонившуюся ветвь малорослых страшненьких недоэргастеров, забредших слишком далеко на север и вымерших от непосильных тягот и невзгод вдали от исторической родины. И совсем другое, когда это гордые представители генерального эволюционного тренда, охватывавшего Ойкумену от мыса Доброй Надежды до самых до окраин, то есть, собственно, до Дманиси.В таком случае не грех и таксономию подправить. И вот уже звучит предложение считать эргастеров хроноподвидом Homo erectus ergaster – не сказать чтобы оригинальное, но обретшее новое звучание и особый переливчатый тон. А чтобы не забывали уникальность именно родных дманисских черепов, не пора ли и на самого Карла нашего Линнея с его принципами биномиальности и триномиальности замахнуться и выдать тетраномиальное название Homo erectus ergaster georgicus (Lordkipanidze et al., 2013)? Что ж, замахнемся и на Линнея! Наконец, и хабилисы не так уж выделяются на фоне грандиозного внутридманисского размаха, а посему отчего бы и их не включить в тот же самый вид на правах молодых самок? Весы таксономистов опять кренятся в сторону “объединителей”…Замечателен пятый череп, что и говорить! Долго еще не утихнут водовороты ученых дискуссий, много еще бумаги и мегабайтов будет изведено на статьи и, чем черт не шутит, книги, посвященные ему. Но неутолима жажда познания. А потому, не желая изменять внутреннему зуду, вновь воззовем к грузинским коллегам: доколе?! В приложении к статье невозможно не заметить интригующую картинку распределения находок в Дманиси, а на ней – два скелета: от того же пятого индивида и подростка, да плюс к ним фрагменты от старика. Это же два с половиной скелета, товарищи! Конечно, часть костей уже была описана в 2007 году, но не пора ли представить миру и прочее? И коли уж так модно стало интерпретировать все возрастной изменчивостью – даешь анализ! Мы бы и сами не прочь им заняться, но кости в Грузии, а мы здесь. Полные скелеты эргастеров не такая банальщина, чтобы годами держать их на полках без описания. Грузинские коллеги, мир ждет вашего слова!
Почему Дманиси – не парантроп?
Описание пятого черепа из Дманиси вызвало много споров, суждений и вопросов. Некоторые люди, не слишком близко знакомые с древними гоминидами, но все же несколько подкованные в вопросе, задаются вопросом: а действительно ли дманисский человек – человек? Покатый лоб, огромные челюсти, развернутые скуловые дуги, хмурое выражение – чем не австралопитек, чем не парантроп? Что тут, собственно, человеческого?Однако же человеческого в дманисце хватает. Австралопитеки и парантропы тоже выделяются не по наитию антрополога, а по комплексу важных диагностических признаков.От грацильных австралопитеков дманисец разнится катастрофически. Во-первых, австралопитеки существенно мельче. Конечно, дманисец тоже не гигант, но все же… Даже крупный по австралопитековым меркам самец Stw 505 имел мозг 505 см³, тогда как у дманисца все же 546 см³. Одно из разительных отличий дманисца от всех австралопитековых – степень выступания челюстей. Челюсти грацильных австралопитеков несравненно прогнатнее дманисских. От восточноафриканских парантропов дманисец отличается прямо обратным свойством – своей прогнатностью. Парантропы имели огромные челюсти, но эти челюсти у восточноафриканских парантропов никогда не выступали резко вперед. У ранних эфиопских и поздних южноафриканских бывало, но первые далеки от Дманиси хронологически, а вторые – географически.Совершенно различается конфигурация лица: у всех австралопитековых оно резко уплощенное, даже вдавленное внутрь, а у дманисца какая-никакая, но горизонтальная профилировка выражена; говоря человеческим языком – средняя часть лица выступает вперед относительно скул.Относительные размеры лица и мозговой коробки австралопитековых и дманисца несопоставимы: у первых – и грацильных, и массивных – челюсти крупнее относительно мозгового отдела – как в длину, так и в ширину. У парантропов лба при взгляде спереди практически не видно, а дманисец хотя тоже не Сократ, но все же подает надежды. У парантропов лицевой отдел буквально подавляет собой мозговую коробку, а у дманисца последняя все же больше, чем верхняя челюсть.Форма неба и альвеолярной дуги дманисца вполне человеческая – с достаточно плавно закругленной передней частью, в противоположность спрямленной у парантропов. У парантропов передние зубы очень маленькие, особенно в сравнении с огромными молярами. Размеры зубов дманисца далеко не достигают парантропьих значений. Например, у южноафриканских Paranthropus robustus, имевших зубы меньшие, чем восточноафриканские P. boisei, площадь коронки первого верхнего моляра по методике, примененной в статье о Дманиси, составляет 197,4, у Australopithecus afarensis – 163,5, у Australopithecus africanus – 177,5а у Дманиси тот же параметр – 159; второго моляра: у южноафриканских парантропов – 229,1, у Australopithecus afarensis – 191,1, у Australopithecus africanus – 224,4, у Дманиси – 197. Даже у хабилисов для первого моляра размер будет 162,6, для второго – 185,9, то есть первый моляр дманисца меньше не то что зубов парантропов, а даже мелких в целом хабилисов.У австралопитеков и парантропов более плавно закруглен свод, практически нет заваликовой борозды, а также совершенно отличен затылок – он почти вертикальный у грацильных видов, приостренный у южноафриканских парантропов и как бы приплющенный, чуть ли не скошенный у восточноафриканских. У дманисца же затылок крупный, высокий и далеко выступает назад, подчеркнут перегибом на границе с теменной костью и снабжен затылочным валиком. Форма затылочного рельефа совершенно различается у австралопитековых и дманисца.Весьма расходятся вообще все признаки рельефа. У австралопитеков и парантропов надбровье вполне умеренное или даже слабое по древним меркам, а у Дманиси – огромное, в этом отношении наши предки попервоначалу явно перехватили лишку.Прямо обратное соотношение височных линий: у Дманиси они хотя и расположены высоко, но все же не сходятся вместе аж на 4 см. У взрослых самцов грацильных австралопитеков и парантропов они всегда и в обязательном порядке сливались в мощный сагиттальный гребень. Только самки австралопитековых могли себе позволить неслияние височных линий. Этот признак принципиален, ибо соединение височных линий и, соответственно, схождение височных мышц на середине свода автоматически означает прекращение роста головного мозга. Все древнейшие гоминиды имели сагиттальный гребень, а род Homo тем и знаменит, что первым отказался от такого украшения, за счет чего смог позволить себе отращивать мозги намного дольше и, стало быть, больше. Правда, Дманиси не может похвастаться особой мозговитостью, но надо же было с чего-то начинать!Существенное ослабление жевательной мускулатуры дманисца сравнительно с австралопитеками и парантропами очевидно при оценке позадиглазничного сужения: у первого оно хотя и выражено, но не чрезмерно по эректоидным меркам, а вот у вторых сужение было запредельным в гоминидном масштабе.Скуловые дуги различны принципиально: у дманисца они, конечно, велики и широки, но при сопоставлении с парантропами кажутся просто жалкими. Любой уважающий себя парантроп, особенно взрослый самец, имел о-го-го какие скуловые дуги! Горилле такие не снились!Нижняя челюсть дманисца огромна, но по всем возможным параметрам отличается от челюстей австралопитеков и парантропов. Особенно бросается в глаза малая толщина и огромная высота тела дманисской челюсти, тогда как у австралопитековых пропорции диаметрально противоположны.Конечно, новый дманисский череп удивителен и необычен. Но все же это – человеческий череп; древнечеловеческий, если быть точным. Если приглядеться внимательнее, пятый череп из Дманиси отличается от четвертого – старческого – только чуть большими размерами за счет надбровья, большей приплюснутостью свода да прогнатной верхней челюстью. А ведь на четвертом черепе челюсти маленькие только оттого, что все зубы выпали при жизни, а альвеолы – ячейки для зубов – заросли задолго до смерти. Челюсти при этом всегда катастрофически уменьшаются. Второй же череп D2282 вообще по всем главным характеристикам: форме свода, рельефу, размерам и выступанию челюстей – соответствует пятому, даром что он женский.Конечно, Дманиси отличается не только от австралопитековых, но и от современных людей. Строго говоря, он занимает промежуточное положение. С нашей точки зрения он примитивен, австралопитеки бы сочли его чересчур новаторским. Но такова эволюция – в ней нет граней, но есть переходы. Где проводить линию между австралопитековыми и людьми? Находки в Малапе не так давно поставили этот вопрос ребром. Дманисский череп добавил в проблему нового колорита. Все же отсутствие сагиттального гребня и уменьшенные относительно мозгового отдела челюсти и зубы перевешивают все архаизмы и примитивности дманисца. Эти свойства были важнейшими, обеспечившими возможность прогрессивного роста головного мозга, а следственно – и культурного прогресса человечества вплоть до современности. Дманисцы находились у самых истоков этого рывка в цивилизацию. Они были еще примитивны: глядя со стороны Homo, мы видим огромные челюсти, покатый лоб, здоровенные зубы, приплюснутость свода и, самое главное, маленький мозг – переходные черты между австралопитековыми и “истинными” людьми. К тому же грузинским гоминидам не повезло с географией, и решающий рывок осуществили их африканские родственники, но зато дманисцы законно могут числиться в благородных рядах Homo.Таким образом, первые люди, благодаря пятому дманисскому черепу, оказались более первобытными и дремучими, чем казалось доселе, но все же можно радоваться, что они превзошли своих предков – грацильных австралопитеков – и не скатились в столь безвыходную бездну специализации, как парантропы – наши невезучие двоюродные братья по эволюции.
Уже не ранние Homo: Homo ergaster
Далее на африканскую сцену выступают Homo ergaster. Их главное отличие – заметный рост мозга и, соответственно, мозговой коробки с увеличением надбровного и затылочного рельефа, увеличением толщины костей свода, относительное уменьшение челюстей и абсолютное – зубов, округление альвеолярной дуги. Плавность перехода от хабилисов к эргастерам хорошо подтверждает происхождение вторых от первых. По этой же причине значительное количество фрагментарных находок интервала 1,5–1,6 млн лет назад не находит прочного места в систематике. К примеру, нижние челюсти KNM-ER 731, KNM-ER 820 и KNM-ER 1507 определялись и как Homo habilis, и как Homo ergaster, и как Homo erectus.
Голотипом для описания вида послужила нижняя челюсть KNM-ER 992 из Кооби-Фора (Groves et Mazak, 1975). Ее датировка колеблется примерно от 1,4 до 1,5 млн лет назад. Ее главное отличие от предшествовавших эогоминин – сравнительная грацильность. Кстати, по этому признаку KNM-ER 992 отличается даже от многих других эргастеров, но, будучи голотипом, она задает нам образцовый пример “эргастерности”. Другой “классический” Homo ergaster – затылочная кость и нижняя челюсть KNM-ER 730, обладатель коих жил 1,55 млн лет назад.
Ко времени 1,5 млн лет назад морфология черепа поменялась достаточно существенно. В частности, это выразилось в увеличении толщины свода, что отлично видно на примере фрагмента KNM-ER 1466 с датировкой ровно 1,5 млн лет назад. Но, слава палеонтологии, в нашем распоряжении имеется достаточный набор более-менее целых находок. Это, прежде всего, предположительно женский череп KNM-ER 3733, имеющий новейшую датировку 1,65 млн лет назад. Также это мужской череп KNM-ER 3883, обладатель которого жил 1,57 млн лет назад. Между этими гоминидами пролегает без малого сотня тысяч лет, так что не странно, что они отличаются друг от друга на уровне, близком к видовому. Но от хабилисов оба отличаются одним и тем же набором признаков: это заметно бóльший объем мозга – 848 и 804 см³, более мощное надбровье и выступающий затылок, более крупное абсолютно, но меньшее относительно мозговой коробки лицо, уменьшенные размеры неба, альвеолярной дуги и зубов, ослабленный альвеолярный прогнатизм. Аналогичные признаки, но бóльшая массивность обнаруживаются на более фрагментарном своде черепа KNM-ER 3892 возраста 1,43 млн лет.
Посткраниальные останки Homo ergaster не столь уж малочисленны, но весьма фрагментарны, а потому всегда диагностируются с большой долей сомнения. Это, например, обломки бедренных костей KNM-ER 737, KNM-ER 1809, большая берцовая и таранная кости KNM-ER 1476. Более комплектен скелет KNM-ER 1808 с датировкой 1,59 млн лет назад. Остатки черепа позволяют говорить, что это был именно Homo ergaster, однако кости сильно деформированы вследствие гипервитаминоза A. Из-за заболевания многие признаки могут быть искажены, но, по крайней мере, мы можем установить рост – порядка 1,7 м, что много, особенно учитывая женский пол KNM-ER 1808.
Кстати, о морковке, белых мишках и пчелках…
Кости KNM-ER 1808 изуродованы, притом отнюдь не течением времени и не тягостью земных пластов. Скорее всего, болезнь KNM-ER 1808 вызвана очень сильным гипервитаминозом A (Walker et al., 1982). Свод черепа и кости скелета покрыты бесформенными костными наростами – поражены периоститом. Подобные деформации не так легко заработать. Самый прямой путь – съесть за раз пару тонн морковки. Но прямой – не значит простой. Есть другой способ – слопать печень какого-нибудь хищника, лучше всего белого медведя, на худой конец – моржа или кита (немало полярников погорело на незнании основ физиологии!). Правда, в Африке с ними напряженка, но и крупная кошка тоже подойдет; как говорится, на безмедвежье и саблезубый тигр – морж. Этот метод, понятно, тоже не лишен недостатков, ведь для развития периостита недостаточно разок выпотрошить леопарда, надо делать это регулярно. Мы, конечно, верим в способности нижнеплейстоценовых тарзанов, но все же день изо дня вырывать печень у саблезубых гомотериев (кстати, как раз в это время они вымирают в Африке – так вот она, причина?!) – это, наверное, перебор. Еще можно есть много печени морских рыб, но вот беда – от Кооби-Фора до моря немножко далеко: полтысячи километров.Существует, однако, более спокойное и надежное средство получить гипервитаминоз A: есть пчелиных личинок (Skinner, 1991). В них витамина A содержится тоже немало, главное – любить процесс. Мед и личинки в Африке – отличный источник витаминов, белка и жира. Посчитано, что пигмеи мбути в дождливый сезон могут до 80 % калорий получать из меда и пчелиных личинок. Кстати, пигмеи жалуются, что если переусердствовать, то начинают болеть кости и суставы. Не это ли происходило и с KNM-ER 1808?Возникают и другие вопросы. Например, как эргастеры могли разорять пчелиные гнезда? Современные конкуренты Винни-Пуха почти всегда обкуривают их дымом, да еще желательно при этом пользоваться сетками и перчатками. Думается, эргастерам последняя роскошь была недоступна, а вот запалить огонек они, вероятно, были уже способны.Кстати, гипервитаминоз A сопровождается массой впечатляющих симптомов: на сухой шелушащейся коже появляются высыпания и трещины, она чешется, лицо краснеет, ладони желтеют, губы сохнут, а уголки рта кровоточат, ногти истончаются и ломаются, человек лысеет, у него выпадают брови. Кроме того, несчастный теряет аппетит, у него нарушается сон и болит голова, кости и суставы, его тошнит, он становится раздражителен. Никому такого не пожелаешь… Зато какой простор для современных реконструкторов, стремящихся к гиперреалистичности в изображении первобытных времен!
Особый интерес представляет фрагментарный скелет эргастера KNM-ER 803a-t, имеющий датировку около 1,53 млн лет назад. Замечательно, что по ряду признаков – например, размерам и строению плюсневой кости – этот индивид очень напоминает дманисцев и KNM-ER 1500, а по некоторым – к примеру, слабому изгибу локтевой кости – оказывается довольно прогрессивным. Размер тела, вероятнее всего, был небольшим, около 1,6 м.
Намного представительнее знаменитый скелет KNM-WT 15000, известный также как Мальчик из Турканы или Нариокотоме III из пачки Нату формации Начукуи Западной Турканы. Его не раз передатировали, долгое время общепринятым считался вариант 1,53 млн лет назад, но самый современный – 1,47 млн лет назад (McDougall et al., 2012). Скелет замечателен своей уникальной сохранностью: следующие сопоставимые – только некоторые неандертальцы. Однако индивидуальный возраст примерно в 11–12 лет затрудняет сравнения с другими находками. Поэтому KNM-WT 15000 определяли и как Homo ergaster, и как Homo erectus, а по мнению некоторых исследователей, он отличается на видовом уровне как от KNM-ER 992, так и Сангирана и KNM-ER 3733 (Schwartz et Tattersall, 2000b).
Некоторые особенности KNM-WT 15000 первоначально были описаны как примитивные, но в реальности являются следствием патологии развития. К таковым относится сужение позвоночного канала (Latimer et Ohman, 2001).
Череп KNM-WT 15000 несет все признаки ранних архантропов, но выраженные в нерезкой форме благодаря молодому возрасту. Очевидно, поэтому же часть черт выглядит схожей с таковыми у хабилисов. Например, объем мозга не очень велик – 750–880 см³, надбровье развито не слишком сильно, затылок более-менее округлый.
Благодаря сохранности, возможно установить размеры тела KNM-WT 15000. Первоначально рост подростка определялся в пределах 1,6–1,68 м, а взрослого аппроксимировался до 1,8 и даже 1,95 м. Однако в последующем, во-первых, было показано, что реконструкция роста сильно зависит от того, на основании каких материалов и каким математическим методом мы ее выполняем (Hens et al., 2000), а во-вторых, целый ряд работ был посвящен обсуждению вопроса – а был ли ростовой скачок у гоминид того времени (Clegg et Aiello, 1999; Smith, 2004)? Если его не было, то длина тела взрослого KNM-WT 15000 могла быть всего около 1,63 м (Graves et al., 2010). В любом случае пропорции тела были резко вытянутыми, выраженно тропическими и одновременно вполне человеческими, в отличие от хабилисов.
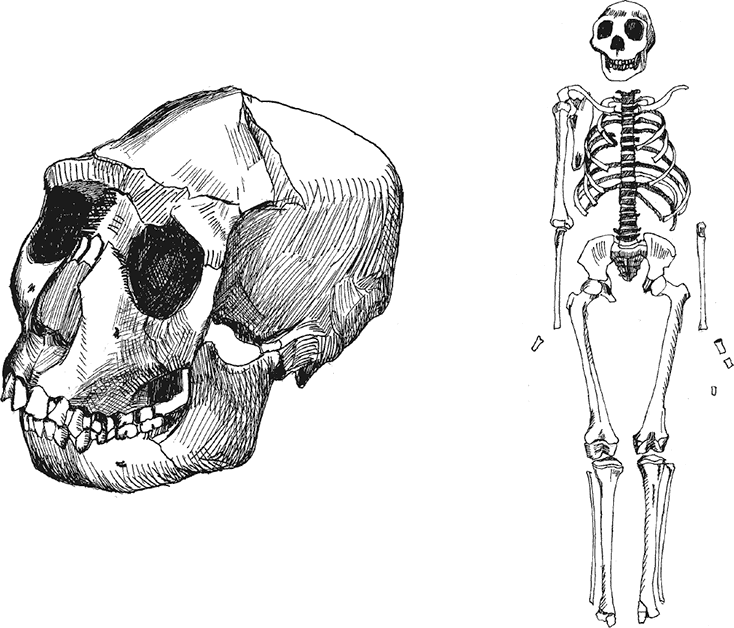
Рис. 19. Череп и скелет KNM-WT 15000.
К сожалению, от KNM-WT 15000 почти не сохранились кости кистей и совершенно не сохранились кости стоп. Этот пробел в некоторой степени заполняет третья пястная кость KNM-WT 51260 из Каитио, недалеко от Нариокотоме, имеющая датировку 1,42 млн лет назад. На этой кости есть такой важный признак, как шиловидный отросток. Этот отросток отсутствует у шимпанзе, Australopithecus afarensis и Australopithecus sediba; судя по основанию второй пястной кости, не было его и у Homo habilis OH 7. Зато отросток сильно развит у неандертальцев и современных людей, поскольку нужен для укрепления кисти при трудовой деятельности. Таким образом, на грани Homo ergaster и Homo erectus кисть уже была истинно “трудовой”, человеческой; неспроста незадолго до этого гоминиды изобрели новую – ашельскую – технику изготовления орудий.
Находки в Мелка-Контуре – фрагмент левой плечевой кости Гомборе IB-7594 с датировкой 1,6–1,7 млн лет назад и обломок нижней челюсти Гарба IVE с датировкой 1,4–1,5 млн лет назад – чаще определяются как Homo erectus, но по сути ничем не отличаются от Homo ergaster Кооби-Фора. В Западной Туркане, в слоях пачки Нату формации Начукуи с датировками 1,44 млн лет назад, найдены не очень представительные обломки черепа KNM-WT 16001, также сочетающие черты Homo ergaster и Homo erectus.
В Олдувае гоминиды из слоев моложе 1,6 млн лет назад обычно обозначаются как Homo erectus, но среди них также можно видеть существенную изменчивость. Самым комплектным и показательным, без сомнения, является мужчина OH 9, которого неоднократно передатировали с наиболее достоверным итогом в 1,33–1,48 млн лет назад. Его черепная коробка уже очень велика – 1067 см³ и морфологически ближе к классическим архантропам. Впрочем, от азиатских эректусов “шелльского человека” отличает ряд черт, отчего и он не избежал описания в качестве самостоятельного вида, причем сразу в трех ипостасях: Homo leakeyi (или Pithecanthropus leakeyi), Homo (Proanthropus) louisleakeyi и Pithecanthropus olduvaiensis (или Homo erectus olduvaiensis).
С OH 9 можно было бы отсчитывать начало архантропов-эректусов, если бы не OH 12. Особенности мозговой коробки OH 12 очень похожи на таковые OH 9, тогда как некоторые детали строения лица больше соответствуют варианту KNM-ER 3733 (Anton, 2004). Объем его мозга – 727 см³. При этом OH 12, вероятно, вдвое моложе: его датировка может быть около 700 тыс. лет назад, хотя другие авторы отстаивают цифры до 0,78–1,2 млн лет назад (McBrearty et Brooks, 2000). Из близких отложений происходят нижние челюсти OH 22 и OH 51, также “зависающие” между Homo ergaster и Homo erectus. Судя по тазовой и бедренной костям OH 28 все из тех же отложений, гоминиды этого времени были достаточно рослыми – порядка 1,65 м для женщины. Впрочем, фрагментарность всех этих находок и существование других “образцовых” Homo erectus с датировками порядка 1–1,2 млн лет назад не позволяет основывать на них слишком глубокомысленные выводы. Таким образом, граница двух этих видов, как и всегда в палеонтологии, оказывается нечеткой, растянутой на многие тысячи лет.
Нижняя челюсть KGA10–1 из Консо с датировкой около 1,4 млн лет назад чаще определяется просто как Homo erectus. Одна из самых заметных тенденций в хронологических изменениях нижних челюстей гоминид “загадочного миллиона” – понижение высоты восходящей ветви. Если у рудольфенсисов восходящая ветвь была очень высокой, то через эргастеров к эректусам она заметно “сдает”, так что у KGA10–1 может называться просто низкой. Очевидно, это отражение уменьшения высоты верхней челюсти и, следовательно, лица в целом. Параллельно мы видим и постоянное сокращение размеров моляров.
Особые находки сделаны в кенийских местонахождениях FwJj14E в Илерет и GaJi10 в Кооби-Фора с датировками 1,51–1,53 и 1,435 млн лет назад – это следы ног нескольких человек, ходивших по вулканическим туфам в компании с бегемотами и другими зверями (Bennett et al., 2009). Следы замечательны по нескольким причинам. Во-первых, они явно человеческие. Во-вторых, большой палец был прижат к прочим не в меньшей степени, чем у современных людей, всю жизнь ходящих босиком. В-третьих, пальцы были завернуты внутрь меньше, чем у австралопитеков Лаэтоли, но больше, чем у современных людей. Видимо, ориентация пальцев была одним из существенных признаков стопы и менялась уже на уровне Homo; это крайне интересно, ведь получается, что прямохождение продолжало эволюционировать даже 1,5 млн лет назад. В-четвертых, эти люди крупные. Они имели рост 1,53–1,86 м и вес от 41,5 до 60,3 кг (Dingwall et al., 2013). Это заметно больше, чем у австралопитеков и хабилисов, но соответствует ожидаемому для эргастеров – и время сходится, что приятно. Не исключено, конечно, что это отпечатки ног и вовсе парантропов, ведь об их стопе мы, честно говоря, вообще мало знаем, но хочется верить, что перед нами – следы наших предков, людей, проходивших как хозяева по необъятной Родине своей.
Кстати, об огне
Переход от Homo habilis к Homo ergaster ознаменовался величайшим открытием человечества – освоением огня! Этому вопросу посвящены бесчисленные страницы (отличные обзоры: Clark et Harris, 1985; James, 1989). Особенно много, как обычно, сказано неспециалистами. Мы же ограничимся немногими фактами. Древнейшие следы использования огня известны на восточноафриканских стоянках Чесованжа, Мелка-Контуре, Гомборе I, Гадеб, в двух местонахождениях в Кооби-Фора и одном в долине Среднего Аваша, в южноафриканской пещере Сварткранс. Они имеют возраст около 1,5, вероятно до 1,6–1,7 млн лет назад. Особенно замечателен очаг в Чесованже в слое с датировкой 1,38–1,46 млн лет назад. Исследования прокаленной почвы из него показали, что она нагревалась до температуры около 400 °C, как в обычном костре, причем огонь горел долго, чего не бывает при случайном пожаре природного происхождения (Gowlett et al., 1981). Более того, очаг обложен камнями, хотя и не так красиво, как делают современные туристы.Мы не знаем, как эргастеры добывали огонь и могли ли они его вообще целенаправленно добывать. Но несомненно, что использование огня стало поворотным пунктом человеческой цивилизации. Можно долго петь дифирамбы огню и его гениальным покорителям. Ведь люди смогли греться, пугать зверей, готовить еду, менять свойства природных материалов и изготавливать новые. Вероятно, обработанная на огне пища стала главной причиной изменения челюстей, зубов и вообще всего пищеварительного аппарата. Без огня люди вряд ли бы заселили север и попали в Америку. Без огня мы не полетели бы в космос, а эта книга не была бы написана. Слава огню!
В Южной Африке наличие вида Homo ergaster спорно. Однако к нему относились четыре зуба и мелкий фрагмент затылочной кости из Линкольн-Кейв. Останки крайне фрагментарны; диагностика их как Homo ergaster вызывает основательное сомнение с учетом датировки 115–253 тыс. лет назад при наличии орудий раннего ашеля (Reynolds et al., 2007). С другой стороны, может, это останки тех самых невероятно архаичных гоминид, которые якобы дожили вплоть до появления сапиенса и дали примесь в негроидов и о которых говорилось выше?
В пятом уровне брекчии Стеркфонтейна пара находок тоже определялась как Homo ergaster. Это разрушенные нижние челюсти Stw 80 (очень схожая с SK 15) и Stw 84 (имеющая несколько более продвинутые черты).
Примерно 1,3–1,4 млн лет назад сформировались древнейшие слои со следами присутствия людей в Убедийе в Израиле (раньше их датировали 1,9–2,5 млн лет назад; Belmaker et al., 2002). Найденные тут два небольших обломка теменной кости, фрагмент височной, кусочек свода, два резца и моляр относятся, вероятно, к более поздним временам (но наверняка древнее 680 тыс. лет назад), они определялись как австралопитек, Homo habilis, Homo ergaster или Homo erectus, хотя реальные признаки не позволяют уверенно говорить об этом.
Наконец, около 1,2 млн лет назад и меньше мы имеем “чистокровных” Homo erectus не только в Африке (Боури и Буйя, имеющие датировку ровно в 1 млн лет; возможно, Салданья), но и в Европе (Сима-дель-Элефанте), и в Азии (Сангиран и Гунванлин). Эти находки отмечают становление классических архантропов и одновременно расселение их по Ойкумене.
Отнесение всех “эректоидных” гоминид второй половины “загадочного миллиона” к Homo ergaster по-своему удобно, но и тут возможны сомнения. Даже KNM-ER 3733 – казалось бы, классический эргастер – может не принадлежать к этому виду, ибо, по мнению некоторых авторитетных исследователей, отличается от KNM-WT 15000 на уровне вида, а KNM-WT 15000, в свою очередь, столь же сильно разнится от KNM-ER 992 – голотипа Homo ergaster (Schwartz et Tattersall, 2000b). KNM-ER 3883 отличается от них всех. Посему KNM-ER 3733 определялся и как Homo leakeyi и даже послужил голотипом для выделения Homo kenyaensis, а KNM-ER 3883 – голотипом для Homo okotensis. Все же, кажется, нет смысла плодить слишком много экзотических названий. Понятно, что черепа подростков, женщин и мужчин, разделенные сотнями тысяч лет, да еще реконструированные из обломков, не могут быть идентичны. Но, по крайней мере, мы можем быть спокойны за отсутствие географических вариаций, пока не выходим за пределы Восточной Африки.
Из вышеприведенного обзора видно, что некоторые авторы определяют древнейших Homo ergaster еще во времена существования Homo rudolfensis – до 1,9 млн лет назад (KNM-ER 2598, KNM-ER 3228, KNM-ER 3732, KNM-ER 3734), хотя морфологическое сравнение показывает, что все такие находки не выходят за пределы изменчивости Homo rudolfensis. “Образцовые” эргастеры укладываются в промежуток времени 1,65–1,4 млн лет назад, хотя некоторые их черты могли сохраняться и позже.
Являются ли эргастеры самостоятельным видом? С момента первоописания споры по этому поводу не утихают. Некоторые восточноафриканские находки отстоят друг от друга на сотни тысяч лет, и впору удивляться, что они вообще бывают схожи между собой. От классических же азиатских эректусов Африка отделена тысячами километров, а потому ожидать идентичности их населения тоже странно. Но все же находок не так и мало, появляются новые, мы выявляем хронологические и, возможно, географические вариации, общие контуры африканской эволюции вполне понятны, и хочется верить, недалек тот день, когда фрагменты загадочной мозаики сложатся в окончательную и четкую картину.
Происхождение Homo: калейдоскоп мнений
Предки эогоминин, как ни странно, неочевидны, а этот вопрос имеет прямое отношение к их систематике. Хотя подавляющее большинство исследователей сходится, что предками были Australopithecus afarensis, некоторые сомневаются в этом, хотя бы частично. В частности, исследователи окаменелостей Омо указывают, что в слоях после афаренсисов, но до “ранних Homo” обнаруживаются зубы, удивительно схожие с южноафриканскими Australopithecus africanus. Таким образом, A. africanus оказывается не географически обособленным южноафриканским видом, а хронологической стадией (Hunt et Vitzthum, 1986). Такая точка зрения подтверждается и некоторыми исследованиями южноафриканских пещер, в которых основная часть находок классических A. africanus получает датировки всего 1,5–2,5 млн лет назад (Berger et al., 2002). Главная сложность этой концепции заключается в том, что она основана практически только на исследовании изолированных зубов. Парочка подходящих целых черепов могла бы выправить положение, но их пока нет.
Особый вопрос – место собственно южноафриканских грацильных австралопитеков во всей этой запутанной истории. Можно указать три основных варианта: либо африканусы являются тупиковой ветвью, либо они дали южноафриканских “парахабилисов” и, возможно, смешались с параллельно возникшими восточноафриканскими, либо они потомки афаренсисов и предки хабилисов, но вообще не привязаны к Южной Африке, просто там лучше сохранились слои с их останками. В пользу первой версии говорит масса специализаций A. africanus, в пользу второй – существование малапских A. sediba, в пользу третьей – особенности гоминид из слоев B, C, D, E и F в Омо, а также, например, OH 24, OH 68 и OH 70.
Другие палеоантропологи ищут экзотических предков для Homo rudolfensis. После описания вида Kenyanthropus platyops возникла версия, согласно которой именно этот плосколицый австралопитек дал рудольфенсисов, которых в этом случае можно называть Kenyanthropus rudolfensis, отделяя и отличая как от Australopithecus, так и от Homo (Collard et Wood, 2001). Однако самостоятельный статус самого кениантропа находится под большим сомнением. Даже уплощенность его лица, во-первых, не уникальна на фоне вообще плосколицых австралопитеков, а во-вторых, в значительной степени может быть следствием тафономической деформации.
Многие и многие исследования были посвящены попыткам разобраться во взаимоотношениях гоминид “загадочного миллиона”. Как обычно бывает в палеоантропологии, ученые разделились на два лагеря – “дробителей” и “объединителей”.
Первые (Blumenschine et al., 2003; Clarke, 2012; Grine et al., 1996; Kramer et al., 1995; Lieberman et al., 1996; McHenry et Coffing, 2000; Rightmire, 1993; Smith et Grine, 2008) считают возможным разделить виды Homo rudolfensis, H. habilis, H. ergaster и H. erectus. Крайние дробители добавляют еще виды H. gautengensis, H. microcranous, H. georgicus и предполагают еще и иные, например Australopithecus sediba, Homo kenyaensis, Homo okotensis, Homo leakeyi и неназванный вид, представленный черепом KNM-ER 1805. Крайняя морфологическая изменчивость “ранних Homo” позволяет метрическими и сравнительно-филогенетическими методами разделить их не только на отдельные виды, но роды и даже подсемейства (Дробышевский, 2003), что показывает огромный масштаб различий между группами. Иногда эти виды представляются как последовательно сменявшие друг друга (особенно для H. habilis и H. ergaster), иногда – как существовавшие параллельно или разнесенные географически (особенно для H. ergaster и H. erectus c H. georgicus). В частности, подчеркивается, что неспроста древнейшие люди довольно массивны (Homo rudolfensis), потом становятся мелкими (H. habilis), а после снова вырастают (H. ergaster).
Объединители (Lee et Wolpoff, 2005; Miller, 2000; Miller et al., 2002) обращают внимание, что слишком часто находки “разных видов” оказываются в одних слоях, что противоречит элементарным экологическим правилам о конкуренции родственных существ. Не многовато ли их? Если виды занимали близкие экологические ниши и не были разделены географически и хронологически (а у гоминид они вряд ли были специализированными, судя хотя бы по современным обезьянам и человеку), то как бы они вообще разделились? Ведь еще существовали Australopithecus garhi, Paranthropus aethiopicus, Paranthropus boisei, Paranthropus robustus, последних из коих дробители к тому же иногда делят еще на два вида. Таким образом, напрашивается логичная мысль о том, что по крайней мере некоторые из этих “разных видов” представляют собой просто самцов и самок или особей разных возрастов.
Дробители возражают на это тем, что нет достоверных случаев нахождения разных видов в одном слое, кроме надежно доказанного сосуществования эогоминин и парантропов. Ведь разница расположения в метр отложений значит тысячи, а то и сотни тысяч лет, а стало быть – и экологических условий. Вместе с тем среди дробителей тоже нет единства, конкретные виды и находки тасуются ими в самых разных сочетаниях.
Как представляется, одной из главнейших проблем, стоящих на пути решения вопроса о взаимоотношениях гоминид “загадочного миллиона”, является недоучет хронологического фактора. Слишком часто исследователи напрямую сравнивают находки, в реальности разделенные сотнями тысяч лет. Конечно, частично это объясняется неточностью самих датировок и обнаружением множества фрагментов на поверхности земли, а не в слое, но стоит все же стремиться к разделению материалов на хронологические группы. Иначе сравнение разных “ранних Homo” можно уподобить сравнению черепов питекантропов и современных людей – между ними тоже пролегают около 800 тыс. лет, как между древнейшими Homo и первыми достоверными архантропами. Никто ведь не усредняет значения по гейдельбержцам и сапиенсам, разделенным “всего” 200 тыс. лет, почему же мы можем говорить о “ранних Homo”, отстоящих друг от друга на бóльшие временные промежутки? Если за второй миллион лет существования рода Homo сменилось три “надежных” вида (H. erectus, H. heidelbergensis, H. sapiens), то почему бы трем видам не смениться и за первый – “загадочный” – миллион?
И действительно, можно видеть, что в целом гоминиды “загадочного миллиона” подразделяются на три морфологические группы, более-менее распределяющиеся и на три хронологических этапа: Homo rudolfensis с 2,4–2,3 по 1,9–1,8 млн лет назад, Homo habilis с 1,9–1,8 по 1,7–1,6 млн лет назад и Homo ergaster – до 1,5–1,4 млн лет назад. Как обычно бывает в палеоантропологии, четкие границы провести невозможно в принципе, но тенденции изменения прослеживаются достаточно хорошо.
Происхождение Homo: а может, все было так?
Как же можно попытаться свести все это многообразие находок и их интерпретаций?
В промежутке от 3 до 2 млн лет назад грацильные австралопитеки переставали быть грацильными. У некоторых это выразилось в увеличении передних зубов – Australopithecus garhi, у большинства – в увеличении задних. Как уже упоминалось, порядка 2,5 млн лет назад в связи с некоторым похолоданием и осушением климата среди разных групп африканских животных наблюдаются аналогичные изменения жевательного аппарата. В русле всеобщих преобразований из вида Australopithecus afarensis возникал вид Paranthropus aethiopicus и через него – Paranthropus boisei. Восточноафриканские парантропы адаптировались и даже специализировались к питанию травянистой растительностью, что особенно наглядно прослеживается в результатах палеодиетологических анализов (Sponheimer et al., 2013). Одновременно характер стертости эмали их зубов, палеоэкологические реконструкции и даже само обилие костей парантропов позволяют предположить, что жили они все же не совсем в открытых саваннах, а скорее рядом с водоемами, питаясь, вероятно, осокой (Cerling et al., 2011). Те же популяции, которые сохраняли “широкие взгляды на жизнь”, питались всем подряд и жили где попало, не столь специализировались своим жевательным аппаратом. Фактически эволюция гоминид интервала 3–2 млн лет назад – это появление массивных австралопитеков, тогда как Homo – отставшие неспециализированные группы, не нашедшие своего теплого местечка и бесприкаянно мыкавшиеся там и сям по африканским просторам. Однако появление специализированного варианта не могло произойти мгновенно и никак не сказаться на состоянии “серой массы”. На протяжении неведомого промежутка времени особи “препарантропов” и “прелюдей” могли скрещиваться между собой. Скорее всего, этот промежуток был не таким уж коротким, ибо у приматов репродуктивные барьеры вообще слабы. Отсутствие или ослабление конкуренции вследствие специализации и обитание в специфических условиях позволили первым парантропам вырасти в численности, судя по соотношению находок, успешнее, чем первым Homo. Таким образом, генные потоки в некоторый момент должны были преобладать от “препарантропов” к “прелюдям”, имея закономерным результатом увеличение у последних массивности в целом и мегадонтии в частности. Надо думать, именно этот эффект мы и наблюдаем в явных “парантропоидных” чертах Homo rudolfensis. Практически все гоминиды с датировками больше 2 млн лет обладают мегадонтными молярами и несколько уменьшенными передними зубами, их нижние челюсти массивны, с толстыми телами и высокими восходящими ветвями. Таким образом, Homo rudolfensis – это первые люди, несущие немалый “заряд парантропности”. Вероятно, скрещивание между парантропами и уже “готовыми” эогомининами могло происходить и позже, не такого ли метиса мы имеем в лице KNM-ER 1805? Или этот “Загадочный череп” представляет позднюю разновидность Homo rudolfensis – с уменьшенными зубами, но выраженными прочими признаками этого вида?
Несколько иная ситуация сложилась в Южной Африке. Там тоже появились парантропы, но другого, заметно менее массивного вида Paranthropus robustus. Этот вид резко отличался от восточноафриканского преобладанием в диете частей древесных растений. Впрочем, численность южноафриканских парантропов тоже была заметно большей, чем местных “ранних Homo”. Однако меньшая массивность южноафриканских “препарантропов” и их более позднее возникновение оказали закономерно меньшее влияние на “прелюдей”, что мы видим, например, в особенностях немегадонтных Australopithecus sediba и совсем уж мелкозубых Homo naledi. Впрочем, в последующем, с оформлением Paranthropus robustus, южные эогоминины также не избежали “робустизации”, причем едва ли не большей, чем восточноафриканские. Увеличение размеров зубов, массивности челюстей и резкое сближение височных линий у Stw 53, SK 80/846/847 и прочих телантропов или Homo gautengensis может отражать именно этот процесс.
Расстояние от Восточной Африки до Южной велико, но не непреодолимо, находки в Малави показывают, что гоминиды вполне могли проникать из одной части континента в другую. Вряд ли регионы были изолированы столь сильно, что формировались разные виды, скорее, ситуация была аналогична положению с современными павианами. Западноафриканский гвинейский павиан отличается от североафриканского анубиса, который отличается от гамадрила с севера Восточной Африки, тот, в свою очередь, – от бабуина юга Восточной Африки, а последний – от южноафриканского чакмы. Вместе с тем отличия не мешают всем им гибридизироваться в пограничных зонах, почему разные авторы склонны выделять от одного до пяти видов. Так и с гоминидами: мы можем статистически отличать южно– и восточноафриканских, но отдельные признаки отдельных находок иногда дают неожиданные специфические сходства на разных концах континента.
Другое дело – эогоминины, покинувшие пределы континента около 2 млн лет назад. Фактически это были поздние рудольфенсисы, но, уйдя с прародины, они оказались в изоляции. Поэтому по пути они, естественно, менялись, так что в Дманиси мы встречаем людей, которые по многим показателям не отличаются от Homo rudolfensis, но все же имеют некоторую специфику.
Со временем парантропы и эогоминины в Африке дифференцировались все больше; первые становились все массивнее, а вторые, изолировавшись генетически, стали грацилизоваться, что наглядно выразилось в уменьшении размеров челюстей и заклыковых зубов. С этого момента – порядка 1,85 млн лет назад – начинается вид Homo habilis. Впрочем, грацильность этого вида весьма относительна: во-первых, имеется целый ряд находок, связывающих Homo rudolfensis и Homo habilis непрерывной изменчивостью; это, например, KNM-ER 1590, OH 4 и даже OH 7 – голотип вида “Человек умелый”, во-вторых, размеры зубов и относительная величина челюстей хабилисов больше, чем у последующих гоминид. Homo habilis удивительны двумя странностями. Во-первых, у них уменьшился не только жевательный аппарат, но и размеры мозга. Во-вторых, пропорции их тела и конечностей оказываются едва ли не архаичнее, чем у афарских австралопитеков. Таким образом, мы видим, что понятие “прогресса” весьма относительно, он виден только на больших временных интервалах и немало зависит от конечного итога. Но ведь вопрос и в том – есть ли он, конечный итог? С точки зрения самих хабилисов, они, несомненно, были заметно прогрессивнее своих предков, так же как и мы считаем себя прогрессивнее неандертальцев и кроманьонцев, превосходивших нас мозговитостью.
Уменьшение размеров мозга у хабилисов по сравнению с более древними рудольфенсисами могло быть связано с повышением стабильности условий окружающей среды. Увеличение мозга рудольфенсисов, равно как и парантропов, было, очевидно, реакцией на перестройки африканского биома. Само возникновение новых родов и видов из пребывавших до того миллион лет в стабильности афарских австралопитеков говорит само за себя – среда бурно менялась. В эпоху же хабилисов условия могли вновь более-менее устаканиться. Припеваючи жить в саванне можно и с мозгом павиана, вот хабилисы и “расслабились”. Меньше неожиданностей – меньше надобности в генераторе нестандартных решений. Но жизнь берет свое: новые вызовы судьбы породили новые эволюционные подвижки.
Порядка 1,65 млн лет назад постепенные изменения морфотипа Homo habilis достигли такого уровня, что мы можем говорить о появлении нового вида – Homo ergaster. Его представители стали заметно крупнее, уменьшение зубов сопровождалось заметным ростом мозга и преобразованиями черепной коробки. Поскольку репродуктивно эргастеры были уже давно и полностью отделены от парантропов, а экологическая пластичность в сочетании с развитием культуры позволяла занимать все новые местообитания, Homo ergaster могли вступать с Paranthropus в прямую конкуренцию. Значительно больший интеллект и отсутствие пищевой специализации дали эргастерам заметные преимущества, а судьба массивных австралопитеков была предрешена. Предыдущий миллион лет трудовой деятельности не мог не сказаться и на строении рук эргастеров: они, похоже, стали первыми обладателями “трудовой кисти” и, видимо, именно благодаря этому смогли создать новую культуру – ашельскую, существовавшую после еще больше миллиона лет.
Так ли все было? Слишком много возможностей, слишком много допущений и предположений…
Мы точно знаем, что 1,2 млн лет назад архантропы уже расселились до Испании на западе и до Китая и Явы на далеком востоке. Начиналась заря нового этапа эволюции – времени истинных гоминид. Но что происходило до этого? До этого лежал бурный и насыщенный событиями миллион лет – загадочный миллион.
Назад: Глава 4 Звено между звеньями: “ранние Homo”, или Загадочный миллион: заря и первая половина истории Homo
Дальше: Глава 5 Homo erectus: непрямые дороги человека прямоходящего

