Книга: Достающее звено. Книга 2. Люди
Назад: Глава 3 Щелкунчики, побежденные мышами: массивные австралопитеки
Дальше: Странные Homo Южной Африки: naledi выходит на люди
Глава 4
Звено между звеньями: “ранние Homo”, или Загадочный миллион: заря и первая половина истории Homo
Около 2,5 млн лет назад, одновременно с парантропами, появились и более прогрессивные существа – так называемые “ранние Homo”. Они отличались от австралопитеков многими чертами, главная из которых – крупный мозг. В промежутке от 2 до 1 млн лет назад заметно изменились пропорции тела – от коренастых типично австралопитековых у хабилисов до вытянутых человеческих у эргастеров, что было связано с окончательным выходом в саванны и повышением мобильности, что, в свою очередь, стало одним из важнейших условий последующего расселения гоминид по планете и их приспособления к самым разнообразным условиям. Впрочем, не стоит преувеличивать прогрессивность “ранних Homo”; самые ранние их представители с трудом отличимы от грацильных австралопитеков, а некоторые ученые и вовсе считают их позднейшей разновидностью австралопитеков. К тому же “ранние Homo”, судя по всему, вовсе не были однородной группой, и далеко не все они – наша прямая родня.
Одна из основных особенностей “ранних Homo” – уменьшенный жевательный аппарат. Челюсти и зубы становились все меньше, жевательные мышцы ослаблялись, рельеф на черепе понижался. Навсегда исчез сагиттальный гребень – даже у самых крупных самцов его не было.
Кстати, о пользе височной мышцы…
У человека есть четыре пары жевательных мышц; из них самая обширная – височная, она имеет веерообразную форму, так как начинается широкой дугой вдоль височной линии лобной и теменной костей, а кончается узким хвостом на венечном отростке нижней челюсти. Плоское брюшко височной мышцы закрывает мозговую коробку сбоку. У крупных приматов правая и левая мышцы сходятся на макушке на сагиттальном гребне, так что весь череп оказывается покрыт ей. Так как мышца по умолчанию толстая и мощная, она обеспечивает дополнительную защиту головы. От этого боковые стенки черепа оказываются тонкими, особенно район птериона – места соединения лобной, теменной, височной и большого крыла клиновидной кости, а чешуйчатый шов между височной и теменной костями сделан внахлест, довольно-таки непрочно. У человека же жевательная мускулатура особенно ослаблена, да и весь череп истончен в сравнении с обезьянами. Посему боковая часть черепа – висок – оказывается самой уязвимой. Поэтому и стреляться надо в висок: кость тут тонка, да к тому же расположена почти вертикально – не срикошетит.
История человеческого рода насчитывает более двух миллионов лет. Но первая половина этого срока покрыта завесой не то чтобы полной неизвестности, но таинственности и загадочности. Что происходило до и позже – более-менее ясно, но в промежутке от примерно 2,5 до приблизительно 1,5 млн лет назад становление рода Homo сопровождалось бурными и неясными событиями. Часто эволюция человека представляется как последовательность стадий, но людям “загадочного миллиона” не повезло. Австралопитеки, парантропы, архантропы, палеоантропы, неоантропы – все получили свои законные обобщающие названия несмотря на то, что неоантропы, например, появились всего лишь – и это при самой оптимистичной оценке – 200 тыс. лет назад. А люди, жившие целый миллион лет, удостоились только невнятного обозначения “ранние Homo”, причем спорно и отнесение их к Homo, и то, можно ли называть ранними тех, кто жил, скажем, 1,5 млн лет назад – почти в середине общего родового срока. Хорошими названиями могут быть “эогоминины” или “преархантропы”, они уже использовались в литературе, хотя применяются редко.
Границы “загадочного миллиона”, как обычно в палеоантропологии и вообще палеонтологии, могут быть определены лишь приблизительно; конечно, палеонтология не равняется на круглые цифры. Резкой границы не было ни между австралопитеками и преархантропами, ни между последними и собственно архантропами. В целом, “загадочный миллион” укладывается в промежуток 2,5–2,2–1,5–1,2 млн лет назад.
“Самый загадочный миллион”: жизнь до Homo
На самом деле “загадочных миллионов” было два. Первый из них лежит между 3 и 2 млн лет назад. Это самое интересное время, когда в Африке закончили свое существование грацильные австралопитеки и начали – парантропы и люди. В это же время появляются первые каменные орудия труда. Но этот промежуток времени минимально освещен палеоантропологически.
Во времена древнее 3 млн лет Африку населяли образцовые австралопитеки нескольких видов. В частности, в Хадаре большинство афарских австралопитеков имеют датировки от 3,9 до 2,9 млн лет назад. Например, классический череп AL 444–2 датирован 3 млн лет; AL 822–1 и AL 438–1, вероятно, чуть древнее – 3,1 млн лет назад; похожий возраст у находок AL 437, 439 и 440 (например: Kimbel et al., 1997, 2003).
В Омо в слоях формации Усно с датировками 3,0–3,3 млн лет назад обнаруживаются только кости австралопитеков, аналогичных хадарским и лаэтольским, то есть Australopithecus afarensis; останки из слоев B, C, D, E и F были описаны как Australopithecus africanus, что странно для Восточной Африки, но это в любом случае австралопитеки. В более поздней – 2,3 млн лет назад – формации Шунгура от слоя B до нижнего G по-прежнему следуют кости Australopithecus africanus, а вот в верхней части слоя G – уже Homo habilis, хотя фрагментарность находок не позволяет утверждать это с полной уверенностью.
В Западной Туркане, в Локалеи, правая бедренная кость KNM-WT 16002 была описана как Australopithecus sp. indet. (Brown et al., 2001); ее возраст 2,7±0,3 млн лет назад. В близких отложениях обнаружен “Черный череп” KNM-WT 17000 древнейшего парантропа – Paranthropus aethiopicus.
Кости из эфиопских местонахождений Боури, Матабайету и Гамеда с древностью 2,5 млн лет назад были описаны как Australopithecus garhi (Asfaw et al., 1999; Heinzelin et al., 1999). Это самые поздние достоверные грацильные австралопитеки, ряд специализаций которых свидетельствует, что они не были предками преархантропов. На австралопитеках гари род грацильных австралопитеков прекратил свое существование. Но небесследно; были как минимум две линии потомков: массивные австралопитеки (или парантропы) и преархантропы. О парантропах уже говорилось выше, они тоже оказались тупиком, а вот Homo стали новой цепочкой звеньев.
Самое интересное – момент происхождения рода Homo. Интервалом 2,7–2,3 млн лет назад (а это, между прочим, полмиллиона – немалый срок!) датируется до обидного немного находок. Имеющиеся же фрагментарны (что печально) и обладают противоречивым сочетанием признаков (что логично для переходных форм). Важно, что до 3 млн лет назад включительно обнаруживаются только и исключительно австралопитеки, а после 2,5 млн лет назад достоверных грацильных австралопитеков нет. Очевидно, в этом промежутке и надо искать основание нашего родового побега на древе гоминид.
Леди из Леди-Герару: первая леди среди людей?
Истоки нашего рода кроются во мраке времен. До сих пор в нашем распоряжении были афарские австралопитеки, жившие более 3 млн лет назад, и уже более-менее настоящие Homo с возрастом чуть более 2 млн. Внутри же “самого загадочного миллиона” – в интервале от 3 до 2 млн лет назад – мы знали африканских австралопитеков и австралопитеков гари, которые с наибольшей вероятностью являются тупиковыми ветвями эволюции. Останков же наших предков известно крайне мало – каждый зуб на счету! Теперь же досадный пробел хоть немного, да заполнился!На местонахождение Леди-Герару в Эфиопии неспроста возлагалось много надежд. Тут сохранились слои как раз искомой древности. Исследователи ныне знают толк в поисках: они организуют экспедиции именно в те места, где с наибольшей вероятностью могут найти то, что им надо. Впервые область Леди-Герару была отмечена как перспективная еще в 1970-х годах, но исследовалась недостаточно, так как ученых мало, а Африка большая; все силы уходили на изучение Хадара и других, более богатых местонахождений. Тем не менее палеоантропологический проект в Леди-Герару тоже был запущен, и уже много лет доблестные охотники за первым человеком целенаправленно выслеживают тут свою добычу. И потому великой и одновременно ожидаемой сенсацией стал их законный трофей – фрагмент нижней челюсти LD 350–1 с древностью 2,75–2,8 млн лет назад (Villmoare et al., 2015).Челюсть LD 350–1 замечательна своим комплексом черт. Если рассматривать отдельные признаки (подробнейше разобранные в приложении к статье), то можно даже разочароваться: вроде ничего особенного и нету, просто мелкий афарский австралопитек. В принципе, таких в Хадаре найдено уже немало. Но если поставить LD 350–1 в соответствующее место хронологического ряда афаренсисов, африканусов, седиб, рудольфенсисов и хабилисов, то картинка наполняется глубоким смыслом. Гоминид из Леди-Герару оказывается, во-первых, промежуточным между афаренсисами и “ранними Homo”, а во-вторых, весьма грацильным. Первое вполне ожидаемо из датировки, но всегда приятно, когда теоретическое предсказание оправдывается и подтверждается реальностью. Важно, что LD 350–1 отличается от афарских австралопитеков даже не размерами, а тонкой морфологией челюсти – всяческими бугорками и вмятинками; также первый премоляр у LD 350–1 симметричный, что предполагает малые размеры верхнего клыка. Грацильность же интерпретирована авторами открытия как свидетельство того, что LD 350–1 представляет собой некую особо продвинутую версию предков человека, тогда как многие последующие “ранние Homo” оказываются специализированными тупиками.Авторы описания подробно и добросовестно разобрали параллели гоминиду Леди-Герару: KNM-ER 5431 из Кооби-Фора, зубы из Шунгуры в Омо и AL 666–1 из Хадара. Все они обладают схожим комплексом и могут быть включены в одну переходную группу от австралопитеков к Homo. Одновременно сопоставление LD 350–1 с A. africanus, A. garhi и A. sediba показывает специализированность последних и их неучастие в становлении рода Homo. Авторы считают, что Леди-Герару имеет более примитивную форму альвеолярной дуги, чем Homo habilis и Homo rudolfensis, так что его невозможно отнести ни к одному из этих видов.Как итог, авторы открытия склонны классифицировать LD 350–1 как древнейшего Homo и, более того, как очень особенного Homo, нашего Великого Предка.Ура, товарищи!И тут мой критиканский разум выдает два но.Во-первых, едва ли не основным отличием Леди-Герару от афарских австралопитеков назван рельеф челюсти: отсутствие понижения в месте подбородочного отверстия, наличие понижения между вертикальным валиком передней поверхности симфиза и клыковыми ямками. Но ведь в этом смысле нижняя челюсть – кость вообще весьма изменчивая! Открываем шкаф, достаем оттуда несколько челюстей (у каждого приличного антрополога есть шкаф с черепами) и элементарно убеждаемся в этом! Симметричность первого нижнего премоляра – особенность не только Леди-Герару, она встречается даже у Ardipithecus kadabba из Гоны, не говоря уж о афаренсисах. Еще два отличия от афаренсисов – отсутствие базального выступания гипоконида на M1 и M2 (у A. afarensis оно выражено), а также наличие седьмого бугорка на M1 (у A. afarensis он ни разу не отмечен) – в действительности могут варьировать индивидуально или популяционно, чему свидетельством современный человек. То есть отличия-то действительно есть и они действительно отличают LD 350–1 от австралопитеков в человеческом направлении, челюсть из Леди-Герару вправду промежуточна. Но можно ли придавать этим отличиям таксономический статус? Достаточно ли их, чтобы ввести Леди-Герару в элитный клуб Homo? Какой процент выступания гипоконида, какая площадь моляров и какой индекс массивности челюсти еще австралопитечьи, а какие – уже человечьи? Это вопрос вопросов! А Леди-Герару по многим показателям упорно попадает хотя и между австралопитеками и людьми, но скорее уж в пределы размаха австралопитеков, но остается за пределами изменчивости “ранних Homo”. Так не логичнее ли классифицировать его как очень прогрессивного австралопитека?Во-вторых, продвинутость LD 350–1 во многом следует из малых размеров. Но в статье отчего-то не рассматривается возможность, что пол этого индивида может быть женским, – этот вопрос обойден стороной. Ясно, что определение пола по половине челюсти – дело ненадежное, но и небезнадежное, ведь палеоприматологи делают это даже по изолированным клыкам. Корень клыка в LD 350–1 сохранился, и он небольшой, скорее всего, гоминид из Леди-Герару был-таки действительно леди, так что предположение о малых размерах как следствии женского пола, а не прогрессивности – вполне законно. В некотором роде это замечание позволяет иначе взглянуть и на морфологические особенности – ослабленность рельефа, сравнительную грацильность кости и симметрию премоляра.Сии критические замечания, конечно, не стоит воспринимать как попытку принизить величие открытия – оно действительно замечательно! Их смысл – немножко снизить градус пафоса, дабы избежать крайних суждений, которые потом будут с таким же накалом опровергаться.
8 изолированных зубов KNM-ER 5431 с площадки 203 в Кооби-Фора имеют возраст 2,7–3,0 млн лет. Они диагностировались как Australopithecus afarensis, Australopithecus garhi или, честнее всего, Australopithecus/Homo gen. et sp. indet. В частности, морфология бугорков премоляров у KNM-ER 5431 австралопитечья, а седьмой бугорок на молярах – человечий. Но неопределенность морфологии и отсутствие специализаций позволяют предположить, что это зубы непосредственных предков Homo (Brown et al., 2001).
В Лаэтоли, в верхней части слоя Ндоланья с датировкой 2,5–2,7 млн лет назад, проксимальная часть большой берцовой кости была определена как Australopithecus garhi или Homo sp. (Harrison, 2002), но скорее по дате, чем по морфологии. Кость маленькая, сопоставимая с таким же элементом у Люси. Впрочем, тут же найдена верхняя челюсть EP 1500/1, имеющая все признаки Paranthropus aethiopicus, так что не исключено, что и нога принадлежала древнейшему парантропу, хотя бы и мелкому.
Таким образом, 3 млн лет назад человечьим родом еще даже не пахло, да и 2,7, и даже 2,5 млн лет назад существование Homo никак не может считаться доказанным.
Отсутствие находок приличной сохранности тем более обидно, что обитатели “самого загадочного миллиона” первыми начали изготавливать каменные орудия труда. Самые древние галечные орудия найдены в кенийских местонахождениях Ломекви – 3,3 млн лет назад – и Канжера – 2,2 млн лет назад, эфиопских Гона – 2,5–2,7 млн лет назад, Омо (Шунгура) – 2,4–2,5 млн лет назад, Хадар – 2,5 млн лет назад, Олдувай в Танзании – 1,5–2 млн лет назад, Сенга и Семлики (Ишанго) в Заире – 2 млн лет назад Первые орудия крайне примитивны: это булыжники всего с несколькими кривыми сколами. Однако эти орудия могут быть классифицированы на повторяющиеся типы. Это значит, что создатель, приступая к работе, имел в голове план своего творения, он знал, что хочет получить, и знал, что может получить разный результат. Есть и более специфические, чисто технические признаки, отличающие случайно расколовшийся камень и орудие: ударный бугорок, образующийся в месте сильного целенаправленного удара, оббивка с одного или двух краев при отсутствии ее на большей части камня, концентрация орудий в местах, где такие артефакты не могли образоваться и скопиться естественным путем. Кто автор сих творений – не совсем ясно; потенциально их могли делать австралопитеки гари, парантропы и “ранние Homo”. Родственная солидарность вызывает желание верить, что это были именно последние, но с большой вероятностью мастерами в разных случаях были разные гоминиды.
Заря Homo: древнейшие свидетельства
На звание древнейшего человека рода Homo претендуют несколько находок. Самой полной, без сомнения, является знаменитый череп KNM-ER 1470 – по совместительству голотип вида Homo rudolfensis. Его передатировали множество раз – то удревняли, то омолаживали, но по последним расчетам владелец черепа жил 2,03 млн лет назад (McDougall et al., 2012). Череп по совокупности признаков смело может быть опознан как “неавстралопитековый”. Главные его человеческие особенности – большой объем мозга и отсутствие сагиттального гребня.
Достоверный рекорд объема мозга грацильных австралопитеков принадлежит крупному самцу A. afarensis AL 444–2 – 550 см³. Но 500 кубических сантиметров грацильные австралопитеки достигали очень редко. У парантропов мозг был побольше: у TM 1517 из Кромдраая, возможно, 650 см³, а у большинства прочих – около 500 кубических сантиметров. Иногда приводимые бóльшие цифры основаны на реконструкциях объема мозга по размерам зубов или посткраниальных костей, а стало быть – крайне недостоверны. Для KNM-ER 1470 же минимальный реконструированный объем мозга – 752 см³, а максимальный – 775 см³. Крайне важно, что столь существенный прирост размеров мозга сопровождается полным отсутствием сагиттального гребня, служащего у его обладателей для прикрепления жевательных височных мышц. Все взрослые самцы крупных человекообразных обезьян и всех австралопитеков и парантропов имели развитый сагиттальный гребень. Судя по всему, он был одним из главных тормозов развития головного мозга, ибо формирование гребня в обязательном порядке означает срастание сагиттального шва – продольного между двумя теменными костями, а стало быть прекращение роста черепа и мозга.
Конечно, KNM-ER 1470 обладает и иными отличиями от австралопитеков. В частности, при всей уплощенности лица и его расширенности в нижней части, пропорции заметно отличаются от австралопитековых стандартов. Переносье KNM-ER 1470 хоть и уплощено, но далеко не в такой степени, как у австралопитеков; лицо даже в самой прогнатной реконструкции близко не соответствует варианту, скажем, AL 444–2; лоб несравненно выше и выпуклее, заглазничное сужение не такое резкое, затылок – круглее, а лобный и затылочный рельеф – слабее. Небо KNM-ER 1470 короткое и почти квадратное, со спрямленной передней стороной, тогда как задние зубы увеличены – мегадонтны, хотя и не в такой степени, как у парантропов. Конечно, не стоит идеализировать KNM-ER 1470: даже в сравнении с архантропами он крайне архаичен. Но в рамках концепции существования двух родов – Australopithecus и Homo – KNM-ER 1470 все же вполне достоин называться человеком.
Вопрос в том, к какому виду относится KNM-ER 1470. Вроде бы он заметно крупнее и массивнее “типичных” и при этом более поздних Homo habilis, с другой стороны, он выглядит заметно отличным и от “образцовых” архантропов. Посему В. П. Алексеев в 1978 году описал его как Pithecanthropus rudolfensis (Алексеев, 1978). С этого момента началась череда подтверждений и опровержений самостоятельности этого вида. “Человечность” KNM-ER 1470 очевидна при сравнении его с грацильными австралопитеками, но вовсе не столь явна на фоне современного человека. Посему он является отличным примером “достающего звена”. Впрочем, в немалой степени дело в количестве находок: мы имеем много австралопитеков и поздних Homo, а потому расцениваем KNM-ER 1470 как связку между ними. Если бы у нас было много ардипитеков и рудольфенсисов, но мало афарских австралопитеков, мы говорили бы о последних как о связующем звене между первыми двумя группами. Количество же находок в большой степени определяется сохранностью и доступностью слоев, а в немалой – везением палеонтологов и даже политической обстановкой в конкретных странах, определяющей возможности научных изысканий.
От других претендентов на звание древнейшего человека осталось не так много.
В Южной Африке, в верхней части четвертого уровня Стеркфонтейна, обнаружены обломки черепа с нижней челюстью и набор зубов детеныша примерно пяти лет Stw 151. Находка имеет датировку между 2,0 и 2,6 млн лет назад. Поэтому не странно, что Stw 151 обладает своеобразными особенностями: на общем фоне признаков Australopithecus africanus выражены явные черты Homo habilis (Moggi-Cecchi et al., 1998). В частности, размеры молочных и постоянных зубов в целом промежуточны между этими видами, хотя существенно отличаются от параметров Paranthropus robustus. Характеристики черепа определимы с трудом. Все же стоит отметить смесь австралопитековых и эогомининных черт височной кости, округлость верхней и нижней альвеолярных дуг, а также сравнительно мощное развитие скуловых дуг. Как уже говорилось, некоторые антропологи подчеркивают неоднородность выборки, обычно определяемой как Australopithecus africanus: часть находок проявляет повышенную массивность и крупнозубость и может представлять пращуров Paranthropus robustus. Stw 151 относится ко второй группе, предположительно предковой для “ранних Homo”.
В кенийском местонахождении Туген-Хиллс на берегу озера Баринго, в формации Чемерон, найден обломок правой височной кости KNM-BC 1, имеющий возраст 2,393–2,456 млн лет назад (Deino et Hill, 2002). Совокупность морфологических признаков позволяет диагностировать сей фрагмент как Homo, поскольку все они укладываются в пределы изменчивости этого рода (Sherwood et al., 2002). Отдельные черты совпадают с вариациями парантропов и грацильных австралопитеков, но полный комплекс кости из Чемерона отличается от характерного для них. Впрочем, по ряду признаков KNM-BC 1 отличается от “образцовых” Homo habilis: например, его нижнечелюстная ямка слишком длинная, широкая и глубокая, “австралопитековая”. Однако та же ямка смещена к центру черепа, прямо противоположно варианту парантропов, у которых височно-нижнечелюстные суставы были широко разнесены в стороны; различаются и многие частные особенности морфологии типа формы гребней и бугорков, а также расположения барабанной пластинки. Крайне любопытно, что число совпадений признаков KNM-BC 1 с вариантом парантропов больше, чем с вариантом грацильных австралопитеков. К великому сожалению, трудно напрямую сравнить KNM-BC 1 с KNM-ER 1470 из-за фрагментарности височной кости у последнего, но стоит обратить внимание на схожесть размеров их нижнечелюстных ямок, равно как, кстати, и у KNM-ER 1805. Неспроста целый ряд антропологов относили KNM-BC 1 к виду Homo rudolfensis. Правда, есть еще мнение, что кость из Чемерон принадлежит Australopithecus garhi (Asfaw et al., 1999), но у известных находок этого вида височная кость не сохранилась, отчего это предположение остается сугубо гипотетическим.
На 300 тыс. лет древнее KNM-ER 1470 верхняя челюсть AL 666–1 из Макаамиталу в Хадаре – 2,33 млн лет назад (Kimbel et al., 1996). Замечательно, что челюсть найдена в сопровождении фауны, которая указывает на более открытый, чем в более ранний период, ландшафт. Будучи на 700 тыс. лет моложе самых молодых Australopithecus afarensis и на 400 тыс. лет древнее “ранних Homo” из Олдувая, AL 666–1 является связующим звеном между этими группами. Приятно, что тут же обнаружены грубые галечные орудия, хотя справедливости ради надо напомнить, что древнейшие каменные орудия из Ломекви и Гоны имеют гораздо больший возраст – до 3,3 и 2,7 млн лет. Принадлежность AL 666–1 к людям, а не австралопитекам доказывается целым рядом сравнений, большей частью весьма специфических, типа формы дна носовой полости или верхнечелюстных пазух. Из внешних же признаков обращает на себя внимание ослабление альвеолярного прогнатизма и уплощенность альвеолярного отростка, отсутствие “передних лицевых валиков”, расхождение кзади линий заклыковых зубов, то есть параболоидность альвеолярной дуги, увеличение ширины и глубины неба. При этом небо крупнее и мельче, чем у более поздних хабилисов и эргастеров. Размеры зубов, что здорово, меньше, чем у австралопитеков, но больше, чем у Homo habilis, а морфология зубов вполне “хабилисская”. На основании столь скудных остатков трудно сказать, можно ли определять AL 666–1 как “образцового” Homo habilis. Некоторые его особенности могут объясняться мужским полом, некоторые – эволюционным уровнем.
Примерно такой же, если не больший возраст имеет нижняя челюсть UR 501 из Ураха на севере Малави – 2,3–2,5 млн лет назад (Betzler et Ring, 1995; Bromage et al., 1995a,b; Ring et Betzler, 1995). Здесь также найдены каменные орудия труда. Челюсть очень крупная, массивная и неоднозначно диагностируемая. В оригинальном описании она была определена как Homo rudolfensis с многочисленными чертами Paranthropus aethiopicus и даже Australopithecus afarensis. Такую мозаичность черт авторы объясняют тем, что Australopithecus afarensis был общим предком для Paranthropus aethiopicus и Homo rudolfensis, а около 2,5 млн лет назад их признаки еще не разошлись окончательно. Собственно, примерно с той же обоснованностью можно определить UR 501 как Paranthropus aethiopicus с некоторыми чертами Homo rudolfensis. Масса признаков UR 501 выходит за рамки изменчивости Homo habilis, а потому невозможно причислить челюсть к этому виду. Отнесение UR 501 к Homo rudolfensis тоже может вызывать сомнение вследствие малочисленности других материалов, относящихся к этому виду.
Авторы первоописания UR 501, основываясь на мозаичности признаков этой челюсти и схожих с ней KNM-ER 1482 и KNM-ER 1802, а также на связи их с фаунами и распределении других находок во времени и пространстве, предполагают, что Homo rudolfensis был эндемичным восточноафриканским видом, возникшим параллельно с Paranthropus aethiopicus и, несколько позже, Paranthropus boisei во время сравнительно холодного и сухого периода (Bromage et al., 1995b). Все эти виды объединяет повышенная массивность челюстей и зубов. Замечательно, что мегадонтия и усиление жевательного аппарата характерны и для других животных – не приматов – времени 2,5 млн лет назад (Turner et Wood, 1993). Около 2 млн лет назад условия вновь стали более влажными и теплыми, что вызвало миграции животных, в том числе расселение массивных австралопитеков в Южную Африку между 2 и 1,5 млн лет назад и появление Paranthropus robustus. Согласно этой концепции, Homo habilis представляют собой отдельный вид, возникший первоначально также в Восточной Африке, а затем – между 1,8 и 1,5 млн лет назад – мигрировавший в Южную.
Также на севере Малави, в местности Мвенирондо, в слоях Чивондо, был найден обломок правого нижнего моляра HCRP-MR-1106. Датировка слоев точно неизвестна, она лежит в широких пределах 1,8–2,7 млн лет назад, но с некоторой вероятностью ее можно конкретизировать до 2,33–2,52 млн лет назад (Kullmer et al., 2011). Таким образом, моляр из Мвенирондо также может быть древнейшим известным фрагментом Homo. Зуб был сильно стерт при жизни и обломан, обкатан, растрескан после смерти, так что видовая диагностика была проведена почти исключительно по микростроению эмали и форме поперечного сечения корней. Как и у схожего UR 501, у HCRP-MR-1106 имеются черты как Homo rudolfensis, так и Paranthropus, но первые преобладают.
Еще один “древнейший Homo” найден в Западной Туркане, в местонахождении LA1a формации Начукуи. Это нижний первый моляр KNM-WT 42718 с датировкой 2,34 млн лет назад (Prat et al., 2005). Опять же в ста метрах от моляра обнаружены каменные орудия. Молодость индивида способствовала сохранению всех деталей строения эмали, так что отличия от австралопитеков и парантропов определимы достаточно надежно; в частности, обращают на себя внимание относительно небольшие размеры зуба; одновременно размеры вполне вписываются в рамки изменчивости Homo rudolfensis. Конечно, одного зуба недостаточно для уверенного суждения о видовой принадлежности, но, по крайней мере, мы знаем, что около 2,3 млн лет назад в Восточной Африке существовали сравнительно мелкозубые гоминиды.
Фрагмент черепа KNM-WT 15001 из пачки Нату формации Начукуи в Западной Туркане имеет датировку 2,09 млн лет назад. Он определен как Homo habilis, но морфологические основания этого неочевидны.
Отличными находками недавно порадовала антропологов местность Колом-Одиет в Кооби-Фора с датировками 2,02–2,03 млн лет назад (Jungers et al., 2015). Тут был найден почти полный набор нижних зубов KNM-ER 64060, ключица, лопатка и кости рук KNM-ER 64061. Полное их исследование еще не опубликовано, но в анонсе отмечается, что длинные кости относительно тонки, а стенки их – толсты.
В Омо древнейшей находкой, обозначаемой как Homo habilis, является премоляр Omo L 33–3282 из основания слоя F формации Шунгура, чей возраст 2,36 млн лет назад (Suwa, 1990). Как и в других подобных случаях, фрагментарность находки позволяет только констатировать наличие “кого-то очень похожего на человека”, но лишает возможности проводить широкие сравнения.
Находки в Омо весьма многочисленны, но крайне фрагментарны. Здесь найдены практически только изолированные зубы, потому их видовая диагностика весьма спорна. Из более комплексных находок стоит отметить фрагменты очень древнего – 1,84 или даже 2,3 млн лет назад – черепа Omo L 894–1 из слоя G формации Шунгура. Он был определен как Homo habilis (Boaz et Howell, 1977), однако его скуловые дуги массивны, что свидетельствует о значительном развитии челюстей, а размеры зубов больше, чем у олдувайских хабилисов, но более-менее схожи со значениями Homo rudolfensis из Кооби-Фора. Интересно, что на одном из моляров обнаружен желобок, образовавшийся из-за использования палочки-зубочистки (Puech et Cianfarani, 1988), такая же привычка привела к серьезным проблемам со здоровьем у одного из дманисцев. Ковыряние в зубах было обычным делом у эогоминин?
Древность 2,0–2,02 млн лет назад имеет обломок нижней челюсти Omo L 75–14. Иногда он обозначается как Homo habilis, но хорошо выраженная мегадонтия заклыковых зубов позволяет определить его как Homo rudolfensis. Зубы все же заметно меньше, чем у парантропов, да и морфология нижних премоляров не похожа на таковую у Paranthropus boisei, хотя строение моляров близко к этому виду. Любопытно, что с правой стороны Omo L 75–14 врожденно отсутствовал третий моляр: сия вариация обычна у современных людей, но крайне редка у гоминид такой древности.
Находки Homo с древностью более 2 млн лет редки, фрагментарны и потому трудно поддаются сравнениям и интерпретациям. Не так много можно сказать по изолированным зубам и кускам челюстей. Парадокс в том, что более древние грацильные австралопитеки изучены намного подробнее. Именно поэтому сей временной этап – один из самых интересных для палеоантропологов. Многие вопросы могут быть решены только с новыми находками. Но важно, что задача поставлена, группы искателей бороздят африканские просторы, выведывая окаменелости, которые смогут пролить свет на наши родовые корни.
Очень древние Homo: Homo rudolfensis
После 2 млн лет остатки Homo становятся более частыми. Правда, ясности от этого не прибавляется, число вопросов, как обычно в таких случаях, только растет. Бóльшая часть находок сделана в Восточной Африке.
Особенно много окаменелостей обнаружено в Кооби-Фора – одной из главных палеонтологических сокровищниц Кении. Преархантропы датируются здесь временем примерно от 2 до 1,6 млн лет назад. Вместе с тем здесь же мы наблюдаем и самое большое разнообразие, с трудом поддающееся интерпретации. Практически все находки в разное время и разными авторами диагностировались и как Homo rudolfensis, и как Homo habilis, и как Homo ergaster, и как специфические виды. Иногда разница взглядов объясняется исследованием разных частей находок. Классический пример – KNM-ER 1590a-q, находка, включающая фрагменты свода черепа, верхнюю челюсть и набор зубов детеныша предположительно мужского пола 5,2–5,3 лет. Ее датировка – одна из самых больших в Кооби-Фора, порядка 1,85 млн лет назад. Округлость и грацильность свода, а также отсутствие мегадонтии позволили определить KNM-ER 1590 как Homo habilis, тогда как абсолютные и относительные размеры бугорков моляров – как Homo rudolfensis (например: Grine et al., 2009). Предполагаемый объем черепа – 870 см³ – тоже великоват для хабилисов.
Ряд других находок в Кооби-Фора часто идентифицируется как Homo rudolfensis. Некоторые из них близки по возрасту к KNM-ER 1470: 1,91–1,95 млн лет назад для верхней челюсти KNM-ER 62000 и фрагмента нижней челюсти KNM-ER 62003. Другие имеют датировки около 1,89 млн лет назад, но с уточнением возраста туфа KBS, под которым они залегают, их возраст может быть пересмотрен в сторону удревнения. Это, например, нижние челюсти KNM-ER 1482, KNM-ER 1801, KNM-ER 1802. Все они характеризуются схожим комплексом черт: большей грацильностью в сравнении с парантропами, но массивностью – с Homo habilis и H. erectus, суженностью альвеолярной дуги спереди, но широким расхождением назад, широкой и очень высокой восходящей ветвью, относительно большими молярами, не достигающими, впрочем, значений массивных австралопитеков, смесью признаков микроанатомии зубов Paranthropus и Homo.
Гораздо полнее черепная крышка с частью лица KNM-ER 3732 (все те же 1,88–1,90 млн лет назад). Промежуточные между Homo rudolfensis и Homo habilis размеры позволяют предположить, что это либо самка первого вида, либо самец второго. Другие же исследователи считают, что KNM-ER 3732 является древнейшим представителем вида Homo ergaster, поскольку он достаточно похож на KNM-ER 3883 (Schwartz et Tattersall, 2000b). Все же отличия от эргастеров показательны: KNM-ER 3732 меньше и грацильнее их, с менее развитым надбровным торусом, с объемом мозга около 700 см³, тогда как у эргастеров – около 800 см³.
Некоторые характерные черты Homo ergaster, возможно, формировались в недрах популяций эогоминин довольно рано. Хороший тому пример – затылочная кость KNM-ER 2598, чья датировка, возможно, приближается к 1,9 млн лет назад: значительные толщина, преломленность и развитие рельефа позволили диагностировать ее как принадлежащую Homo ergaster. Впрочем, учитывая данные из Дманиси, мы можем убедиться, что первые эогоминины вовсе не обязаны отличаться какой-то особенной грацильностью свода, так что нет особых оснований не включать KNM-ER 2598 в Homo rudolfensis. В челюсти KNM-ER 3734 с близкой датировкой разные исследователи также усматривают черты Homo rudolfensis или Homo ergaster.
Гораздо фрагментарнее череп KNM-ER 3891, несколько более молодой – около 1,8 млн лет назад. Он тоже был определен как Homo rudolfensis, в пользу чего свидетельствуют сильное развитие височной линии и сильный альвеолярный прогнатизм. Однако скуловой отросток верхней челюсти KNM-ER 3891 не слишком высок, альвеолярная дуга широкая и округлая спереди, а зубы не мегадонтны. Таким образом, можно видеть усиление черт, приписываемых обычно Homo habilis в более позднее время.
Рудольфенсис возвращается! Рудольфенсис возвращается?
Проблема изменчивости и разнообразия “ранних Homo”, видимо, еще долгие годы будет терзать умы палеоантропологов. Две противоборствующие концепции никак не могут одолеть друг друга. Согласно первой, среди первых представителей рода Homo имелась довольно большая половая или же межпопуляционная вариабельность. Согласно второй, наблюдаемые различия слишком велики и отражают существование как минимум двух видов: более мелкого Homo habilis и более крупного Homo rudolfensis. Установлению истины посвящены многие статьи и главы в умных книгах, мудрыми учеными применялись заковыристые математические анализы, оценки полового диморфизма и сравнения с изменчивостью обезьян и гоминид от австралопитеков до современных людей. Ничего не помогает. Как обычно, в конце таких работ звучит вечное упование на грядущие находки, которые таки прольют свет на темную загадку антропогенеза.И вот они, эти новые находки!В 2007, 2008 и 2009 годах в Кооби-Фора в Эфиопии были обнаружены три фрагмента черепов. Добрые антропологи подобрали им круглые номера – все любят красивые циферки с нуликами, которые и запоминаются легко. Наиболее полный и важный обломок – KNM-ER 62000 – почти полная верхняя челюсть. KNM-ER 60000 – практически целая нижняя челюсть. Наконец, KNM-ER 62003 – небольшой, но информативный кусок нижней челюсти. Датировки фрагментов – 1,91–1,95, моложе 1,87 и 1,90–1,95 млн лет назад соответственно. Сравнение этих находок с наличными коллекциями показало, что наилучшим образом они вписываются в рамки проблематичного вида Homo rudolfensis.Еще в 1972 г. Б. Нгенео нашел череп KNM-ER 1470. Сочетание крупных почти ортогнатных челюстей, заметно приплюснутых спереди, довольно большого мозга (большого сравнительно с австралопитеками и известными тогда Homo habilis, а не сапиенсами, как иногда в запале невежества пишут креационисты) и приличной датировки (которая сама по себе долгое время была предметом ожесточенных дебатов, а ныне устаканилась на 2,03 млн лет назад) сделало проблематичным отнесение KNM-ER 1470 к какому-либо из известных тогда видов гоминид. Подвешенное состояние порешил В. П. Алексеев в 1978 г., когда в своей великой книге описал вид Pithecanthropus rudolfensis (Алексеев, 1978). Правда, он сделал это не в полном соответствии с канонами и правилами международной номенклатуры, за что до сих пор подвергается нападкам критиков. Кроме того, книга была написана на русском языке, неведомом мировой общественности. В 1986 г., однако ж, вышел английский вариант книги В. П. Алексеева, а потому несклонные вникать в исторические детали зарубежные антропологи всегда пишут Homo rudolfensis Alexeev, 1986, чем раздражают и веселят наших антропологов. Но мы-то знаем правду: Homo rudolfensis Alexeev, 1978!Проблема, впрочем, не в бюрократических мелочах. Главной сложностью оказалось то, что долгое время Homo rudolfensis оставался видом, известным по одному-единственному черепу. Конечно, были попытки приписать туда же и другие находки, но все они были крайне фрагментарны, а потому образ не очень клеился.Челюсть KNM-ER 62000 замечательным образом напоминает лицо черепа KNM-ER 1470. Она столь же ортогнатна и уплощена, передняя сторона ее альвеолярной дуги так же ровна, отчего дуга становится, собственно, не дугой, а буквой “П”. Замечательно, что носовое отверстие KNM-ER 62000 узкое и почти треугольное, что является типичной чертой “ранних Homo” в целом и черепа KNM-ER 1470 в частности. Зубы KNM-ER 62000 вопреки крупным размерам самой челюсти не слишком велики в сравнении с прочими “ранними Homo”; кроме того, они относительно вытянуты продольно (мезиодистально на одонтологическом диалекте).Нижняя челюсть KNM-ER 60000 уникальна среди челюстей “ранних Homo” своей сохранностью. Удивительно, что полностью сохранились восходящие ветви – огромные, совершенно вертикальные, с эффектно развернутыми углами. Впечатляет и высота симфиза, на котором при некоторой фантазии можно даже углядеть зачаток подбородочного выступа. Как и на верхней челюсти KNM-ER 62000, альвеолярная дуга – не дуга, а прямоугольник. Собственно, именно эта черта позволила исследователям объединить KNM-ER 60000 и KNM-ER 62000. Зубы же их различаются, поскольку на KNM-ER 60000 они относительно укорочены, а не удлинены.Примечательно, что форма альвеолярной дуги отличает KNM-ER 60000 от канонических Homo habilis. Другое дело, что бы это значило? Действительно ли отличия пропорций зубов и альвеолярных дуг свидетельствуют о видовом статусе отличий? Имеющиеся цифровые данные, строго говоря, указывают скорее на уклонение более крупных форм от мелкого большинства.Можно обратить внимание и на более широкий набор находок. Вообще-то, форма нижней челюсти KNM-ER 60000 весьма напоминает таковую челюсти из Дманиси D2600, которая была не так уж давно описана в качестве самостоятельного вида Homo georgicus. Огромная величина восходящих ветвей и симфиза D2600 назывались в качестве эксклюзивных признаков эндемичного вида. Однако ж другие челюсти из Дманиси представляют спектр широкой изменчивости, сравнимый с размахом вариаций челюстей из Кооби-Фора. Весьма различается и форма лица дманисцев – опять же не в большей степени, чем восточноафриканские хабилисы и рудольфенсисы. Равным образом среди людей современных рас можно найти гораздо больший спектр вариаций. Правда, для этого придется сравнивать людей из разных экологических зон, зато на стороне ископаемых находок – интервалы в сотни тысяч лет.Помнится, долгий и нудный подсчет размеров и форм челюстей и прочих обломков из Восточной Африки, проведенный в масштабе изменчивости всех гоминид, привел меня некогда к выводу, что хабилисы и рудольфенсисы расходятся лишь на расстояние подвидов (текст диссертации можно найти на сайте www.antropogenez.ru). Именно так выглядят и графики, отражающие размеры зубов новых находок в Кооби-Фора. Любопытнее, что часть восточноафриканских находок – KNM-ER 1801, KNM-ER 1802, KNM-ER 1805, UR 501 – я тогда посчитал принадлежащими новому виду массивных австралопитеков, конвергентному с “ранними Homo”. Одним из ключевых признаков этого потенциального вида была повышенная массивность в сочетании с прямоугольной формой альвеолярной дуги. Знакомое сочетание! Не его ли мы видим на KNM-ER 60000? Замечательно, что размеры зубов этих индивидов – KNM-ER 1801, KNM-ER 1802, UR 501 – занимают промежуточное положение между Paranthropus boisei и “ранними Homo”, а KNM-ER 1805 почти совпадает с KNM-ER 60000 и KNM-ER 62000. Чехарда таксономических определений продолжается…
Датировка 1,9 млн лет назад позволяет предположить, что к виду Homo rudolfensis относятся тазовая и бедренная кости KNM-ER 5881 из Кооби-Фора. Они описаны как специфические и отличающиеся от восточноафриканских Homo erectus. Собственно, отчего бы им быть похожими на эректусов, если эректусы имеют гораздо меньший возраст? По измерительным же характеристикам кости KNM-ER 5881 закономерно занимают промежуточное положение между австралопитеками и более поздними “ранними Homo”, а из последних ближе всего к хабилису OH 62.
К виду Homo rudolfensis с некоторой вероятностью относятся обломки длинных костей левой ноги KNM-ER 1481a-d, найденные также под туфом KBS и имеющие датировку около 1,89 млн лет назад. Эти кости определяли и как “раннего Homo sp.”, и как Homo habilis, и как Homo ergaster, и как Homo erectus. Бедренная очень похожа на бедренную современного человека, маленькая и грацильная. Вероятно, у многих исследователей срабатывает стереотип: раз рудольфенсисы имеют бóльшие размеры челюстей и зубов в сравнении с хабилисами, то и их посткраниальный скелет должен быть крупнее. На самом деле такого закона в природе нет; например, синхронные рудольфенсисам дманисцы имели столь же крупные зубы, но очень низкий рост и маленькие кости. Замечательно, что коленный сустав KNM-ER 1481b выглядит прогрессивнее, чем у афарских австралопитеков AL 333-x26 и AL 333–4, но практически не отличается от варианта современного человека.
Ту же датировку имеет крайне фрагментарный скелет KNM-ER 3735. Его обычно определяют как Homo habilis, но основания этому крайне зыбки. От скелета осталось довольно много обломков, но каждый из них очень невелик. Малые размеры тела, как и в случае с KNM-ER 1481, вряд ли могут сами по себе служить диагностическим признаком. KNM-ER 3735, вероятно, обладал шимпанзеподобным соотношением длин лучевой и плечевой костей – даже более примитивным, чем у Люси, – и, возможно, увеличенной длиной рук относительно ног (Haeusler et McHenry, 2007). Пропорции KNM-ER 3735 могли быть примитивнее, чем у более позднего, но менее крупного олдувайского хабилиса OH 62 (Haeusler et McHenry, 2004). Вместе с тем на височной кости KNM-ER 3735 имеется шиловидный отросток – прогрессивный гоминидный элемент, который, впрочем, изредка встречается и у австралопитеков.
Еще хуже сохранность скелета KNM-ER 1500a-p, имеющего все ту же датировку 1,89 млн лет назад. Несмотря на наличие фрагмента нижней челюсти, видовая принадлежность остается спорной – Paranthropus boisei, Australopithecus sp., Homo rudolfensis или Homo habilis. Множество примитивных признаков сочетается на этих фрагментах с более-менее прогрессивными. Рост, как и в предыдущих случаях, был небольшим.
Обычно как Homo ergaster или даже Homo erectus определяется правая тазовая кость мужчины KNM-ER 3228. Однако ее стратиграфическое положение – ниже туфа KBS – определяет датировку более 1,87 млн лет назад, возможно 1,95 млн лет назад. Учитывая, что в этих же слоях встречены черепа и зубы только Homo rudolfensis, гораздо логичнее предположить, что и таз принадлежал особи этого вида. Замечательно, что пропорции таза и места прикрепления мышц на нем существенно отличаются от типичных для австралопитеков и приближаются к современному варианту. Кость крупная, так что рост мог достигать даже 1,8 м.
Одна из самых молодых находок, определяемая как Homo rudolfensis, – нижняя челюсть KNM-ER 60000, имеющая датировку 1,78–1,87 млн лет назад (Leakey et al., 2012). Она же является и самой целой из относимых к этому виду. Невозможно не отметить ее крайнего сходства с челюстью D2600 из Дманиси, что особенно впечатляет, учитывая их синхронность. Кстати, и размеры неба более древней находки KNM-ER 62000 в наибольшей степени совпадают с дманисскими D2282 и D2700, по ширине – и с D4500.
Впрочем, небольшой фрагмент тела нижней челюсти KNM-ER 819 тоже очень массивен, а потому иногда также определяется как Homo rudolfensis. Между тем его новейшая датировка – всего 1,5 млн лет назад (McDougall et al., 2012). Таким образом, либо вид Homo rudolfensis продержался дольше, чем обычно считается, либо есть какая-то погрешность в датировке, либо челюсть относится на самом деле к Paranthropus boisei, либо вся концепция самостоятельности Homo rudolfensis неверна, либо, что вероятнее всего, KNM-ER 819 отражает начало увеличения массивности челюстей у нового вида Homo erectus.
Несколько сотен тысяч лет существования вида Homo rudolfensis – длинный и недостаточно изученный этап нашей эволюции. Эти люди жили не там, где их кости могли успешно сохраняться, они еще не хоронили умерших и совершенно не заботились о потребностях будущих антропологов. Парантропы, обитавшие ближе к воде, известны намного лучше. Только упорные усилия современных исследователей могут пролить свет на эти “серые тысячелетия” нашей предыстории.
Не самые древние Homo: Homo habilis
В слоях выше туфа KBS в Кооби-Фора размеры костей эогоминин уменьшаются, а потому практически всегда их определяют как Homo habilis. В частности, к ним относятся фрагменты нижних челюстей KNM-ER 1501 и KNM-ER 1502, чья датировка порядка 1,6 млн лет назад. Более полные находки из поздних слоев Кооби-Фора – KNM-ER 1805 и KNM-ER 1813 – специфичны, а потому будут рассмотрены позже.
Классические Homo habilis найдены в Олдувае. Они не синхронны и достаточно разнородны, но, к сожалению, крайне фрагментарны: не обнаружено ни одного целого черепа. Находки гоминид сопровождаются примитивными галечными орудиями, отчего вся культура получила название олдувайской. К древнейшим эогомининам Олдувая относится маленький фрагмент нижней челюсти OH 4 с датировкой 1,9 млн лет назад. Сохранившийся второй или третий моляр большой, но все же меньше, чем у Homo rudolfensis из Малави и Кооби-Фора.
Строение эогоминин Олдувая времени порядка 1,79–1,86 млн лет назад известно по черепу OH 24, или Твигги, предположительно женскому. Обычно он считается образцовым представителем вида Homo habilis, но малые размеры и некоторые черты строения позволили ряду авторов отождествить его с Australopithecus africanus, другие же видят сходство с Homo rudolfensis. Все же ряд параметров позволяет отличить OH 24 от австралопитеков: это умеренное заглазничное сужение, поднятие затылочного отдела, вертикальное расположение барабанной пластинки, другие детали строения височной кости и основания черепа. Вместе с тем отсутствие шиловидного отростка – выраженно примитивный признак, лицо крайне уплощено и даже вдавлено, резко выступает вперед, носовые кости очень узкие, а по передней стороне лица идут вертикальные “передние лицевые валики”, аналогичные тем, что характеризуют южноафриканских австралопитеков и Stw 53. Все же скулы выдаются не так сильно, как у австралопитеков. Характерно строение верхней челюсти OH 24: небо очень короткое и почти квадратное, линии передних и заклыковых зубов ровные, сходящиеся под прямым углом. При этом нет никаких признаков заклыковой мегадонтии, а передние зубы относительно задних крупны. Объем мозга 560–590 см³, то есть больше, чем у австралопитеков. Строго говоря, чисто морфологически OH 24 вполне может быть расценен как австралопитек, но очень и очень продвинутый, фактически это очередное “достающее звено” между Australopithecus и Homo. Кстати, очень похожи на Australopithecus africanus и другие олдувайские гоминиды близкого возраста, например OH 68 и OH 70.
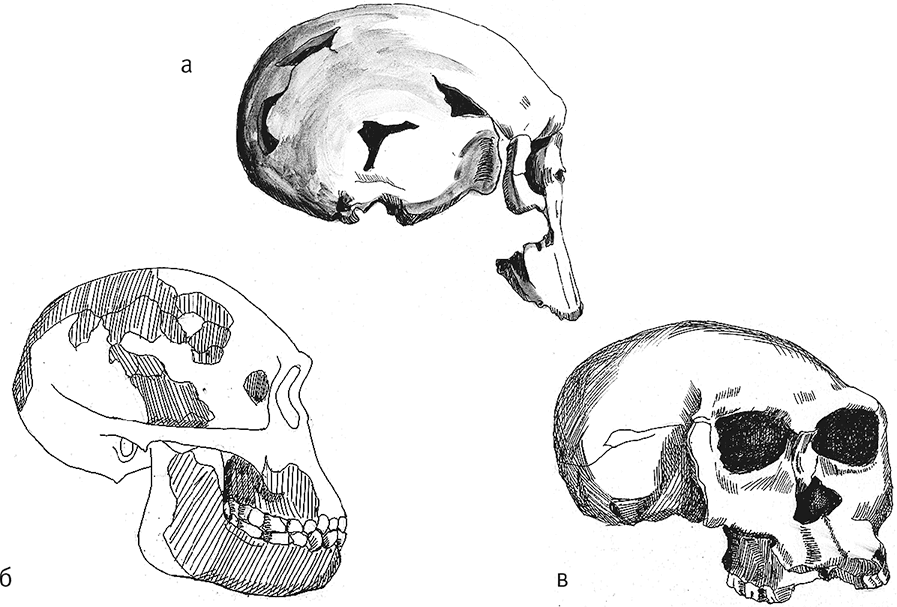
Рис. 13. Череп KNM-ER 1470 (а), черепа OH 13 (б) и OH 24 (в).
Довольно сильно от OH 24 отличается верхняя челюсть OH 65, имеющая аналогичную датировку. Ширина и высота OH 65 несколько меньше, тогда как размеры неба, альвеолярной дуги и зубов – заметно больше, чем у OH 24. Посему ряд антропологов склонны сближать OH 65 с эогомининами типа KNM-ER 1470, а OH 24 – с мелкими вроде KNM-ER 1813 (например: Clarke, 2012).
Голотип вида Homo habilis – обломки двух теменных костей, нижняя челюсть и кости кисти OH 7 – имеют датировку 1,74–1,82 млн лет назад (в 2006 году еще один зуб из той же челюсти был найден в вышележащем слое, так что ранее определявшийся возраст мог быть завышен; Leakey et al., 1964). Не исключено, что кости принадлежат разным индивидам. Забавно, что длина реконструированной теменной кости довольно велика, несмотря на предположительно подростковый возраст, почему некоторые авторы склонны сближать OH 7 с KNM-ER 1470 в противовес мелким индивидам, предположительно представляющим другой вид (Rightmire, 1993); в этом случае смысл термина Homo rudolfensis теряется, а часть хабилисов перестают быть хабилисами. Казалось бы, в пользу этой точки зрения говорит и большая датировка OH 7, однако теменная кость мелкого KNM-ER 1813 в реальности длиннее, чем у большого KNM-ER 1470, а OH 7 занимает между ними промежуточное положение, тем более что реконструкция OH 7 довольно условна. Предполагаемый объем мозга был 657–680 см³. Нижняя челюсть OH 7 не может быть названа массивной, так что ее отождествление с Homo rudolfensis Кооби-Фора и Малави спорно, но подростковый возраст не позволяет делать четких выводов.
Человек ли человек умелый?
Что и говорить, вопросов относительно первых Homo немало. Антропологи неустанно ведут жаркие споры о статусе как целых видов, так и отдельных находок. Примером такой битвы за истину может служить очередная статья, опубликованная недавно в журнале Nature (Spoor et al., 2015). В ней авторы явили на суд ученой аудитории новые реконструкции теменных костей и нижней челюсти OH 7 – голотипа вида Homo habilis. Проблема в том, что нижняя челюсть хабилиса сломана и сплющена, а теменные кости разбиты и почти не смыкаются друг с другом. Вновь реконструированная нижняя челюсть получилась весьма примитивной, с вытянутой альвеолярной дугой. Авторы считают, что она оказывается примитивнее, чем даже на полмиллиона лет более древний Homo AL 666–1 из Хадара (это, правда, верхняя челюсть, но, имея одну, можно понять форму и противоположной). Авторы также аргументируют, что Homo habilis резко отличается от Homo rudolfensis, так что они не могут представлять один вид.Теменные кости OH 7 реконструировались неоднократно, но отчего бы не сделать это еще разок, благо в лаборатории простаивают томограф да ядреный компьютер? И вот очередное цифровое колдовство свершилось, и на глаза удивленной публики выплывают цифры 729–824 см³! А ведь это в полтора раза больше, чем считалось доселе! Как же это объяснить? Авторы реконструкции считают, что все логично: просто большой мозг эволюционировал раньше и активнее, чем форма челюстей.Если суммировать итоги реконструкции челюсти и черепа, получаются интересные выводы: Homo habilis – это не предковый для современного человека вид, а тупик эволюции, настоящим же предком был эфиопский AL 666–1. Эволюция мозговой коробки была более стандартной и ранней, тогда как челюсти менялись замысловатее; Homo habilis сохранял примитивный вариант долго после появления гораздо более продвинутых людей. Впрочем, и Homo rudolfensis – хоть и ближе к нам, чем Homo habilis, но тоже тупик.Вот такие пироги!Как обычно, есть пара-тройка но.Во-первых, форма альвеолярной дуги – штука до крайности изменчивая. Хотелось бы узнать – каких современных людей авторы брали за образец? А ведь среди особо прогнатных восточных экваториалов – австралийских аборигенов и меланезийцев – частенько встречаются индивиды с прямоугольной зубной дугой, резко вытянутой, с клыками в выраженных углах между спрямленной передней линией резцов и прямыми параллельными линиями заклыковых зубов. Что, их тоже теперь относить к австралопитекам? Политкорректность не треснет?Во-вторых, реконструкция объема мозга по раздолбанным теменным костям – дело неблагодарное. Доселе для OH 7 публиковался разброс вероятных размеров мозга от 560 до 724 см³. В. И. Кочеткова реконструировала целые эндокраны OH 7 в трех вариантах, и ведь самый крупный из них оказался как раз самым неправдоподобным! “Австралопитековый” имел размер всего 560 см³, а “умеренный” – 650 см³. Другие авторы предполагали величину около 687 см³. Но 800 см³ – не многовато ли? Томография и компьютеры – это, конечно, здорово, но классику ведь тоже забывать не стоит. Да и есть ли смысл убиваться по поводу размеров именно этого черепа – они всегда останутся спорными, – если есть несколько почти целых черепов: OH 24, KNM-ER 1470, KNM-ER 1805 и KNM-ER 1813.В-третьих, спор о принадлежности “ранних Homo” к одному или нескольким видам ведется уже полвека, и никому пока не удалось победить. Еще одна реконструкция старой находки вряд ли способна поставить окончательную точку в этой грандиозной битве мозгов.В-четвертых, если H. habilis – тупик, H. rudolfensis – тупик, то кто же тогда предок? (На этом месте должны появиться креационисты и радостно закричать: “АГА!!!”) Авторы статьи считают, что H. erectus, но относят к этому виду дманисцев, так что выходит, что эректусы совершенно синхронны и хабилисам, и рудольфенсисам. Точка зрения до крайности спорная.Кажется, путаницу и противоречия можно успешно разрешить, если больше внимания уделять хронологии и не смешивать группы, отстоящие друг от друга на сотни тысяч лет. Тогда будет видно, что H. rudolfensis – более древний и массивный вид, включающий в себя в том числе дманисцев, H. habilis – более молодой и грацильный, а H. ergaster и тем более H. erectus – это уже совсем другая история. Такая схема не избавляет от необходимости выискивать тупики эволюции, но в ней наблюдаемая изменчивость приобретает эволюционный смысл, а количество непонятных ветвей резко сокращается.Так что интрига сохраняется, битва за первого человека в самом разгаре, впереди еще много интересного! Как говаривала Алиса: чем дальше, тем любопытственнее и любопытственнее…
Особый интерес представляет кисть OH 7. По ряду признаков она может быть определена как специализированная – в частности, своеобразную форму и очень маленькие размеры имеет ладьевидная кость. Существенно, что сустав между костью-трапецией и первой пястной костью был седловидный – “трудовой”, хотя и сильнее уплощенный, чем у современного человека. Примитивное строение имеет IV запястно-пястный сустав. Основание II пястной кости по некоторым параметрам ближе к горилльему варианту, чем человеческому, а сама кость довольно грацильна. Соотношение широтных размеров пястных костей OH 7 больше напоминает обезьяний вариант и очень редко встречается у современного человека. На фалангах продольные борозды головок и борозды для сгибателей пальца на телах очень глубокие, а валики по бокам фаланг бугристые, так что тела фаланг расширены в средней части; все эти признаки скорее как у человекообразных обезьян, нежели человека. Вместе с тем головки концевых фаланг расширены, что является одним из важнейших признаков трудовой кисти. Особенно велика концевая фаланга I пальца кисти, хотя она может относиться к стопе, другому индивиду или даже другому виду. Совокупность признаков кисти свидетельствует, что 1,7–1,8 млн лет назад кисть еще не обрела всех человеческих свойств; вместе с тем это самая человеческая кисть из древнейших, намного более человеческая, чем была у австралопитеков.
Такая же мозаика примитивных и прогрессивных черт обнаружена на стопе OH 8, найденной тут же; не исключено, что ей обладал тот же индивид. С одной стороны, стопа принадлежала, очевидно, полностью прямоходящему существу и имела хорошо выраженные продольный и поперечный своды, с другой – своеобразна, а по ряду черт равно отличается от понгид и современного человека (Kidd et al., 1996). В частности, очень оригинальны таранная, пяточная, ладьевидная и кубовидная кости, большой палец, вероятно, был сильнее отведен, чем обычно у современного человека, но при этом первая плюсневая крайне массивна, да и другие длинные и мощные и, вероятно, чуть более подвижные, чем у человека. Таким образом, стопа хабилисов еще могла сохранять адаптацию к жизни на деревьях, но в силу ли эволюционной инерции или действительно использования по этому назначению – неясно. Справедливости ради надо сказать, что ярлычок с большого пальца стопы, если он и был, не сохранился, так что видовая принадлежность OH 8 вообще-то точно неизвестна. Некоторые антропологи считают, что это стопа бойсовского парантропа. Вопрос может быть окончательно решен только обнаружением более-менее целого скелета парантропа или хабилиса.
В слоях с датировками 1,7–1,8 млн лет назад или чуть больше вместе с останками Paranthropus boisei были найдены мелкие фрагменты черепа с большой и малой берцовыми костями OH 6, а также большая и малая берцовая кости OH 35. Судя по всем указанным посткраниальным костям, рост олдувайских эогоминин был невелик, порядка 1,6 м.
Не только размеры, но и пропорции можно установить по остаткам скелета взрослого индивида OH 62, известным также как Дик Дик Хилл или Ребенок Люси и имеющим датировку порядка 1,8 млн лет назад. Скелет состоит из 302 фрагментов, но в данном случае, к сожалению, “много” не значит “хорошо”. Все же можно понять, что по большей части признаков OH 62 соответствует другим мелким Homo habilis Олдувая и Кооби-Фора. Впрочем, и тут не обошлось без альтернативных мнений: отдельные исследователи склонны считать, что OH 62 больше похож на Australopithecus afarensis, чем на Homo habilis (Berillon et Marchal, 2002). В отличие от OH 24, этот гоминид не имел “передних лицевых валиков”, так что нет оснований предполагать его родство с южноафриканскими австралопитеками. Гораздо интереснее все же его посткраниальные особенности. Размеры костей небольшие, так что рост достигал всего 1–1,25 м. Существенно, что руки относительно ног были очень длинными, даже длиннее, чем у Australopithecus afarensis (Richmond et al., 2002). OH 62 и KNM-ER 3735 представляют два самых изученных скелета “ранних Homo”, и в обоих случаях пропорции конечностей выглядят весьма архаичными; судя по ним, эволюция от четвероногих приматов к людям не была совсем прямой, имела место специализация пропорций в первой половине “загадочного миллиона”.
В скором будущем мы узнаем о строении хабилисов намного больше, так как в Кооби-Фора найден скелет KNM-ER 64062 с датировкой 1,82–1,86 млн лет назад (Jungers et al., 2015). От него сохранились кости рук и ног. В предварительной публикации анонсирована смесь примитивных и прогрессивных черт в их строении, кажется с преобладанием именно прогрессивных.
Из несколько более поздних слоев Олдувая – 1,7 млн лет назад – происходит фрагментарный череп подростка OH 16, или Джорджа. Таксономически он определялся как Homo rudolfensis, Homo habilis, Homo ergaster, Homo sp. nov. или ранний Homo erectus, а первоначально даже как Zinjanthropus boisei. В принципе, OH 16 может считаться образцовым хабилисом; он мало отличается от OH 24. Размеры этого черепа оказываются самыми усредненными из известных для гоминид “загадочного миллиона”, в частности, объем мозга составляет 638 см³, типичны для хабилисов и детали строения височной кости. Надбровные дуги выступают несколько сильнее, чем у большинства хабилисов и рудольфенсисов, но слабее, чем у эргастеров и тем более эректусов. Примитивными чертами являются близкое схождение височных линий и очень большие размеры зубов, такие же, как у австралопитеков; особенно велики клыки.
Примерно то же можно сказать о находке OH 13 (Синди, или Синдерелла): как и OH 16, она определялась всеми возможными способами – Homo habilis, Homo sp. nov., Homo ergaster, Homo erectus. Ее возраст колеблется от 1,65 до 1,78 млн лет назад (Spoor et al., 2007). В действительности фрагменты свода черепа с наибольшей вероятностью принадлежат взрослой особи, а верхняя и нижняя челюсти – подростку. Размеры черепа, челюстей и зубов вполне соответствуют типичным для Homo habilis. Впрочем, сообразно поздней датировке, OH 13 обладает и прогрессивными чертами, например скругленностью альвеолярной дуги спереди и расхождением ее ветвей назад. Вместе с черепом и челюстями были найдены фрагменты плечевой и лучевой костей, свидетельствующие о малых размерах тела OH 13.
С большой натяжкой можно назвать скелетом фрагменты костей KNM-ER 164 с датировкой около 1,6 млн лет назад, впрочем столь незначительные, что даже видовая их принадлежность находится под большим сомнением. Это могут быть останки как Paranthropus boisei, так и Homo habilis, и Homo erectus.
Одни из позднейших находок в Кооби-Фора представляют очередной показательный пример смешанности признаков гоминид “загадочного миллиона” – правая верхняя челюсть KNM-ER 42703 с датировкой 1,44 млн лет назад и черепная коробка KNM-ER 42700 возрастом 1,55 млн лет назад. Первая была описана как Homo habilis, вторая – Homo erectus (Spoor et al., 2007). Из этого был сделан вывод, что два указанных вида сосуществовали в Восточной Африке на протяжении как минимум полумиллиона лет. Однако, как справедливо заметили другие антропологи, слишком большое количество признаков отличает KNM-ER 42700 от “типичных” Homo erectus, так что его с бóльшим основанием можно было бы определить как все того же Homo habilis или, более осторожно, Homo sp. (например: Baab, 2008). В частности, слишком маленькие общие размеры, малая толщина костей свода, округлость затылка, минимальное развитие надбровного рельефа никак не вписываются в стереотип архантропов, зато почти идеально соответствуют хабилисам. Размеры KNM-ER 42700, действительно, оказываются минимальными для архантропов, но одновременно максимальными для хабилисов. Объем его мозга был 691 см³, что больше индивидуального максимума хабилисов, но меньше, чем у всех классических архантропов, кроме дманисцев, кои, собственно, и не являются классическими. В некоторой степени двоякое истолкование положения этого черепа в филогении гоминид объясняется промежуточностью его строения, в некоторой – молодым возрастом и, с большой вероятностью, женским полом. Таким образом, время около 1,44–1,55 млн лет назад можно считать последним рубежом сохранения черт Homo habilis в Восточной Африке.

Рис. 14. Галечные орудия.
Кстати, о высоком…
За рассуждениями о зубах и барабанных пластинках не стоит забывать и о духовном. Хабилисы не зря называются “умелыми”: их время – это расцвет галечной, или олдувайской, культуры. Галечные орудия были, очевидно, универсальными, но тем не менее среди них есть несколько типов: чоппер – большое орудие из гальки с оббивкой с одной стороны, чоппинг – большое орудие из гальки с оббивкой с двух сторон. Кроме орудий, выделяются: нуклеус – не орудие, но заготовка, из которой изготавливалось орудие, отщеп – кусок камня, отколовшийся от нуклеуса при изготовлении орудия, чаще выбрасывался за ненадобностью, но иногда мог использоваться без дальнейшей обработки.У первых орудий есть четыре принципиальных отличия от более совершенных: во-первых, из одного камня изготавливалось одно орудие, во-вторых, орудие изготавливалось исключительно деструктивно – путем отсекания лишнего от заготовки (все по О. Родену!), но без комбинации отдельных элементов, в-третьих, обработка края имела целью его заострение (в более совершенных индустриях иногда, напротив, некоторое притупление), в-четвертых, орудие использовалось, видимо, только как прямое продолжение руки, метательных орудий достоверно неизвестно, хотя наверняка и австралопитеки, и “ранние Homo” умели бросать разные предметы (часто находимые сфероиды интерпретируют как метательные камни).Древнейшие примеры использования кости известны из южноафриканских пещер Сварткранс и Дримолен-Кейв с датировками около 1,2–1,8 млн лет назад; они выглядят как костяные обломки, использовавшиеся для расковыривания термитников, что было доказано экспериментально по характерной изношенности их концов. Иногда орудия изготавливались из обломков крупных костей по той же технологии, что и каменные орудия. Несмотря на простоту галечных орудий, их выделка требует определенных навыков и интеллектуальных способностей, точности взгляда и верности руки. Не стоит недооценивать древних творцов; примитивность примитивностью, а сложность сложностью. Читатель может найти в ближайшем овраге кремневый желвак и попробовать сделать хотя бы самый простецкий галечный чоппер. Лучше не надо – шуму будет много, а пользы, скорее всего, никакой; да и пальцы отшибить можно, колоть-то придется камень. Делать такие орудия надо уметь и надо учиться. Трудно сказать, перенимали ли детишки искусство чоппероделания простым наблюдением, или родители целенаправленно обучали своих чад. Думается, группы, где уроки камнеобработки были поставлены более целенаправленно, получали преимущества и в целом выигрывали. Разгильдяи же и лодыри оказывались на обочине прогресса и были обречены на прозябание и в конце концов вымирание.Чопперы достаточно эффективны как ножи для разрезания. Для проверки этого положения проводился опыт: в Африке археологи наделали чопперов, взяли лишнего, никому не нужного слона (уже “готового”, ни одно животное не пострадало!) и экспериментально разделали его тушу за пару часов. Если уж совершенно неподготовленные люди, никогда в жизни не свежевавшие слонов чопперами, смогли управиться за столь малое время, то что уж говорить об умудренных богатым опытом хабилисах!Между 2 и 1,5 млн лет назад появляются новые, более совершенные инструменты – ручные рубила. Это большие заостренные на конце орудия с более-менее ровной обработкой двух сходящихся краев; ручные рубила бывают в виде бифаса – обработанные с двух сторон – или унифаса – обработанные с одной стороны. Древнейшие рубила известны из кенийского местонахождения Кокиселеи 4 с датировкой 1,76 млн лет назад (Lepre et al., 2011), едва моложе – из эфиопского Консо (Beyene et al., 2013). Ручные рубила, найденные в танзанийском местонахождении Пенинж, были сделаны заметно позже – 1,4–1,7 млн лет назад, зато про них известно, что они использовались для обработки дерева (Dominguez-Rodrigo et al., 2001).По наличию типов орудий и их форме выделяют два периода галечной культуры: олдован A – ранний, 2–1,7 млн лет назад, в котором нет бифасов, и олдован B – поздний, 1,6–1,7 млн лет назад, в котором бифасы есть.Кстати, в поисках древнейших индустрий археологи описали довольно много “первых культур”. Например, в Европе выделялась “культура эолитов”, а в Южной Африке – “культура галек Кафу”, но сейчас они не признаются за реальные.Древнейшие стоянки расположены всегда в открытой местности недалеко от воды. По костям животных можно определить, в какой сезон люди жили в том или ином месте. Поэтому мы знаем, что “ранние Homo” совершали сезонные миграции (например: Peters et Blumenschine, 1995). Особенно интересны стоянки в Олдувае. В локальном местонахождении FLK North 6 найден почти полный скелет слона Река, погруженный в глину. Люди не могли вытащить слона из болота (ох, нелегкая это работа!), а потому съели ровно половину, торчавшую на поверхности. Непосредственно рядом с ним найдено 123 артефакта. Аналогичная картина рисуется в местонахождении FLK North II: расчлененный скелет Deinotherium, тоже застрявший в глине, и 39 орудий и манупортов около него. Слона Река съели и обитатели Барогали в Джибути, есть и другие подобные находки.Другое важное местонахождение Олдувая – DK1. Тут был обнаружено скопление базальтовых обломков в виде круга диаметром 3,7–4,3 м (Leakey, 1989). Интерпретация его может быть разной. Кое-кто предполагал, что это просто скопление лавы вокруг дерева, но когда такие же сооружения находят в голоценовых слоях, никто особо не сомневается, что это остатки примитивного жилища в виде шалаша из веток, основания стен которого были придавлены камнями. Но датировка олдувайского жилища – более 1,75 млн лет назад!
Научно-техническая революция: прорыв в будущее по-ашельски
Многие люди при словосочетании “научно-техническая революция” представляют себе космические ракеты, компьютеры, дымящие трубы заводов, паровоз с велосипедом, на худой конец. Однако ж не всегда НТР была синонимом пара и электричества, стекла и бетона. 1,75 млн лет назад наши предки совершили глобальный интеллектуальный прорыв, выйдя за тесные рамки галечной индустрии и на сотни тысяч лет обеспечив своим потомкам благоденствие в уютном совершенстве ашельской культуры.От галечной (или олдувайской – по месту первого описания) ашельская индустрия отличается целым рядом нововведений. Довольно бесформенные олдувайские орудия – чопперы и чоппинги – дополнились четко унифицированными и гораздо более качественно сделанными. Появились и быстро вошли в моду ручные рубила – тяжелые широкие орудия с двумя режущими краями и острым концом. Спрямленным рабочим краем вместо острия отличались кливеры, похожие на нынешние долота. Аморфнее острия, чья конфигурация, видимо, в большей степени зависела от фантазии, лени и надобности. Судя по повторяемости основных типов, каждый из них имел более-менее определенную функцию. Галечные орудия, конечно, тоже удовлетворяли немудрящим потребностям первых людей, но с ашельскими, безусловно, жить стало лучше, жить стало веселее.Cтатья, написанная, как водится, международным коллективом археологов и посвященная описанию очередного древнейшего ашельского местонахождения, была опубликована в “Известиях Американской академии наук” в 2013 г. (Beyene et al., 2013). Надо сказать, что за два года до этого древнейший ашель уже был обнаружен в местонахождении Кокиселеи 4, расположенном в Кении (Lepre et al., 2011), где слои с орудиями залегали на 4,5 м выше уровня, датированного 1,78 млн лет назад, так что имеют возраст около 1,76 млн лет.Новые датировки получены для местонахождения Консо (конкретнее – в KGA6-A1), расположенного на юге Эфиопии. Они практически совпадают с кенийскими – 1,75 млн лет назад. Из этого следует уже тот замечательный вывод, что либо истоки ашельской традиции лежат в еще более глубоком прошлом, либо технические инновации распространялись практически мгновенно по просторам африканских саванн. Видимо, как для современного человека очевидны преимущества компьютера перед печатной машинкой или гусиным пером, поезда и самолета – перед телегой, а ксерокса – перед монахом-переписчиком, так и перед первыми людьми не вставали вопросы: что лучше – чоппер или кливер, чоппинг или рубило? Сделать, конечно, сложнее, но пользоваться – несравненно приятнее. Ортодоксы и консерваторы проигрывали и угрюмо отсиживались по углам Ойкумены, а новаторы торжествующе разносили свет прогресса по долам и лесам.Важнее другой аспект открытия – преемственность технологий. До сих пор многие “наидревнейшие” индустрии частенько зависали как бы в хронологическом вакууме, а их развитие обнаруживалось в более-менее отдаленном будущем. Обычно проблема банально в том, что геологические слои не накапливаются в одном месте непрерывно, иногда они не формируются или даже разрушаются. Другое дело – материалы в Консо. Длительная последовательность отложений позволила проследить последовательность археологического развития от 1,9 до менее чем 1 млн лет назад. В ранних слоях лежат галечные орудия, индустрия в KGA6-A1 была определена как прото-ашель, а в KGA4-A2 с датировками 1,6 млн лет назад найден уже типичный ранний ашель. Разница заключается в качестве обработки орудий: у более поздних она тщательнее, форма вывереннее, симметричнее, режущий край тоньше, увеличивается число ударов, совершенных во время изготовления, уменьшается размер сбитых отщепов. Развитие прослеживается и дальше примерно в том же направлении. Примечательно, что ручные рубила совершенствовались интенсивнее, чем острия. Для последних, в частности, не обнаруживается изменения числа ударов, нанесенных при выделке.Нельзя не заметить – и авторы статьи заметили это, – что с новейшими датировками ашеля практически уравнивается время появления этой культуры и вида Homo ergaster. Ведь давно было обнаружено, что культура совсем уж тесно не привязана к биологическим видам, но в целом соответствие все же есть. Границы же Homo habilis – Homo ergaster и олдувай – ашель расходились примерно на 250 тысяч лет. Теперь же они совпадают почти идеально: 1,65–1,7 млн лет назад для KNM-ER 3733 и 1,75 млн лет назад для Кокиселеи 4 и Консо. С учетом того, что морфология KNM-ER 3733 уже резко отличается от Homo habilis, а камни сохраняются лучше черепов, – совпадение совсем точное. Конечно, это не исключает того, что ретрогады-хабилисы еще долго могли бродить по африканским закоулкам и клепать свои устаревшие чопперы. Но прогресс не остановишь! Мозги и рубила проторили дорогу эргастерам и их потомкам в светлое будущее, к следующей революции, вперед – к “среднему каменному веку”. Но это уже совсем другая история…
Статус вида “человек умелый” весьма зыбок. С одной стороны, его трудно отличить от предшествовавших “людей рудольфских”, с другой – от “людей работающих” и даже “прямоходящих”. Даже таксономическая принадлежность видового голотипа – OH 7 – ставится под сомнение. Homo habilis, несомненно, прогрессивны строением своего жевательного аппарата, но по ряду признаков посткраниального скелета выглядят “откатившимися назад” в эволюции в сравнении даже с афарскими австралопитеками, а по размерам мозга хотя и превосходят австралопитеков, но “отстают” от рудольфенсисов. Следствие ли это прихотливых загогулин эволюционного процесса, или свидетельство происхождения хабилисов от каких-то еще неведомых австралопитеков? А чтоб антропологам не показалось, что загадок маловато, из африканских недр на свет божий появляются совсем уж невразумительные находки.
Странные Homo Восточной Африки: Homo microcranous
Африканские саванны и пустыни хранят еще много откровений. Намеками на то, что не все так просто, являются некоторые окаменелости, не вписывающиеся в стройные стандартные схемы учебников. Некоторые находки из Восточной Африки стоят особняком среди всех эогоминин, даром что они одни из самых сохранных. Это прежде всего два черепа: KNM-ER 1805 и KNM-ER 1813.
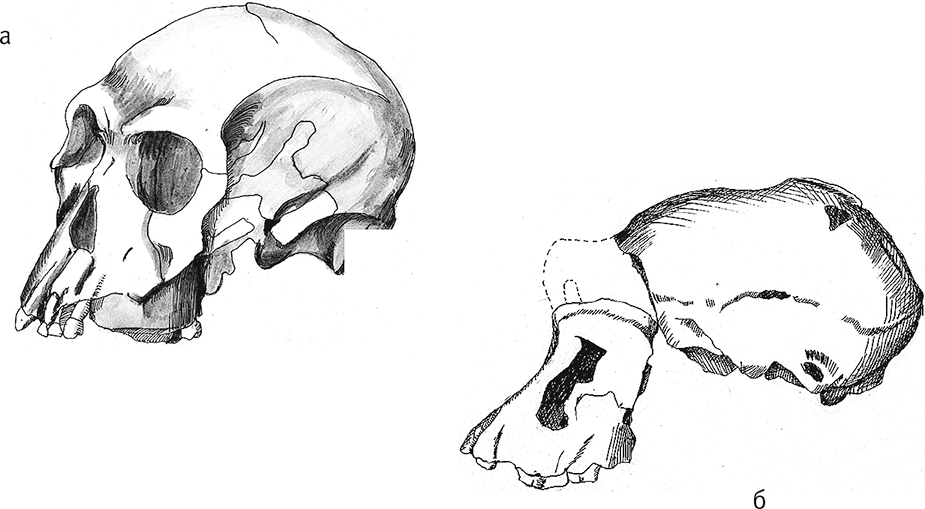
Рис. 15. Черепа KNM-ER 1805 (а) и KNM-ER 1813 (б).
KNM-ER 1805, известный также как “Загадочный череп”, согласно новейшим датировкам, жил 1,75 млн лет назад (McDougall et al., 2012). Мозговая коробка и челюсти разломаны так, что не стыкуются между собой, так что ориентация и даже размеры лицевого скелета точно неизвестны. Все же основные признаки вполне определимы. Признаки эти противоречивы, отчего находка диагностировалась крайне разнообразно: как Homo habilis, Homo rudolfensis, Homo ergaster, Homo sp., Paranthropus boisei. Строго говоря, положение KNM-ER 1805 среди прочих гоминид “загадочного миллиона” совершенно уникально. Череп довольно велик для австралопитеков, но вполне соответствует норме хабилисов: его объем 582 см³. Линейные размеры черепа даже больше, чем у хабилисов, поскольку затылочный рельеф выражен очень сильно (развитие надбровья неизвестно). При этом височные линии в задней части сходятся практически вплотную, образуя хотя и слабый, невыраженный и слегка недоделанный, но все же сагиттальный гребень. На затылке же выйный гребень сливается с височным в единое образование, чего не бывает у эогоминин и даже у Australopithecus africanus. Височная кость, как часто бывает, несет мозаику признаков. Особенно удивителен лицевой скелет: челюсти очень мощные, тяжелые, резко прогнатные, в продольной плоскости лицо вогнуто, а в поперечной средняя часть лица сильно выступает вперед. Судя по височному рельефу, скуловые дуги были немаленькими, но насколько они выступали в стороны, сказать сложно, согласно некоторым реконструкциям – не так уж и сильно, примерно как у KNM-ER 1813 и Stw 53. Альвеолярный отросток резко выдается вперед, выпукл продольно и поперечно и плавно переходит в дно носовой полости, что является выраженно примитивным вариантом. Нижняя челюсть сохранилась плохо, но она явно была массивной. На удивление, зубы KNM-ER 1805 невелики, намного меньше, чем у Australopithecus boisei и большинства Homo habilis, хотя моляры относительно передних зубов несколько увеличены. Сочетание мощнейшего жевательного аппарата и маленьких зубов очень необычно.
Как расценивать такую чехарду признаков – непонятно. Согласно одному из объяснений, KNM-ER 1805 был патологической особью; в пользу этого приводилось, например, наличие у него метопического шва. Однако каких-то специфических признаков болезней на самом деле нет, а метопический шов – вполне нормальная вариация у самых разных приматов, включая человекообразных обезьян в целом и человека в частности.
С другой стороны, можно обратить внимание, что размеры нижних зубов KNM-ER 1805 непринципиально отличаются от Дманиси D2600, хотя верхние заметно меньше, чем у Дманиси D4500. А ведь по времени “Загадочный череп” и Дманиси почти синхронны! Хотя по некоторым абсолютным размерам черепа KNM-ER 1805 заметно уступает дманисцам, зато по объему мозга и ширине лба даже несколько превосходит D4500. Может, это вариации на одну и ту же тему поздних мелкозубых рудольфенсисов?
Возможно также, KNM-ER 1805 был гибридом парантропа и эогоминина: от первого он унаследовал челюсти, а от второго – мозги и зубы. Конечно, биологические признаки не наследуются готовыми комплексами, но “метисная” гипотеза происхождения KNM-ER 1805 ненамного хуже альтернативных версий. Удивительно, что все авторы отмечали своеобразие KNM-ER 1805, но никто не решился дать ему собственное латинское наименование.
Второй “нестандартный” эогоминин Восточной Африки – KNM-ER 1813. Его датировка такая же, как у KNM-ER 1805 (1,75 млн лет назад), но признаки существенно отличаются. Главная особенность KNM-ER 1813 – очень маленькие размеры. Они были увековечены в названии Homo microcranous, предложенном специально для этой находки (череп послужил голотипом и для вида Homo antiquus, включавшего также афарских австралопитеков, но такое объединение оказалось слишком уж спорным, а название было уже раньше использовано для неандертальцев, а потому не может считаться валидным; Ferguson, 1995). Объем мозга KNM-ER 1813 равен 505–510 см³, то есть в верхних пределах изменчивости афарских австралопитеков и ниже нижних – хабилисов. Однако KNM-ER 1813 очень мало похож на Australopithecus afarensis и не очень – на Homo habilis: у него слишком слабо выступающие, хотя и высокие челюсти (выше, чем, например, у OH 24), слишком округлый затылок, слабое надбровье и довольно-таки выпуклый лоб, височные линии широко отстоят друг от друга, череп гораздо долихокраннее, чем у хабилисов; лицо узкое, а его средняя часть сильно выступает вперед, тогда как скулы заметно скошены; передняя поверхность верхней челюсти плоская, без “передних лицевых валиков”; зубы несколько меньше, чем у большинства Homo habilis, но относительно самого черепа не такие уж и мелкие. Особенности височной кости весьма прогрессивны. Вместе с тем скуловые дуги довольно сильно расходились в стороны, а нижняя челюсть должна была иметь очень высокую и узкую восходящую ветвь.
Представляет ли KNM-ER 1813 крайний вариант индивидуальной изменчивости Homo habilis или самостоятельный вид – большой вопрос. Если принять первый вариант, то вариабельность Homo habilis оказывается очень большой, хотя, собственно, почему бы ей и не быть таковой? Речь ведь не о средних, а о крайних значениях. Если же верна вторая версия, то крайне интересно: чем обеспечивалась изоляция между близкими видами эогоминин, особенно учитывая наличие еще и парантропов? И где останки других представителей этого вида?
Будем надеяться, что новые открытия дополнят наши знания.
Странные Homo Южной Африки: sediba и gautengensis
Особняком стоят находки из пещеры Малапа в Южной Африке, имеющие возраст 1,977 млн лет назад. Два скелета и остатки еще двух отсюда были описаны как Australopithecus sediba, причем в первоописании подчеркивалось, что малапские гоминиды имеют примерно поровну признаков Australopithecus и Homo (Berger et al., 2010), так что в принципе они могут называться и Homo sediba. Чрезвычайно любопытно, что целый ряд черт A.sediba указывает на их происхождение от южноафриканских A. africanus: это уплощенность передней стороны альвеолярного отростка верхней челюсти, значительная глубина неба, общая форма нижней челюсти, детали строения зубов. Вместе с тем черт эогоминин тоже немало: в числе их отсутствие расширенности лица, развернутости скуловых костей и “передних лицевых валиков”, относительно умеренный прогнатизм, детали височной кости, строение таза и некоторых других посткраниальных костей. Есть и такие признаки, по которым A. sediba больше похож на A. afarensis, чем на кого-либо еще. От кого A. sediba действительно заметно отличается, так это Paranthropus и Homo rudolfensis. Объем мозга малапцев маленький, типично австралопитековый – около 420 см³.
Рис. 16. Череп и скелет Australopithecus sediba.
Назад: Глава 3 Щелкунчики, побежденные мышами: массивные австралопитеки
Дальше: Странные Homo Южной Африки: naledi выходит на люди

