Книга: Хроники тираннозавра: Биология и эволюция самого известного хищника в мире
Назад: Глава 10 Физиология
Дальше: Часть III Экология
Глава 11
Изменения
Мы рассмотрели различные виды тираннозавров, при помощи филогении поместили их в эволюционный контекст, а также описали их анатомию и вероятное функционирование в связи с анатомическими особенностями. Такие аспекты эволюции тираннозавров, как уменьшение количества пальцев и зубов у более поздних форм, вкратце уже упоминались. Здесь мы рассмотрим подобные изменения более детально и выскажем предположения о том, что они могли означать для различных линий тираннозавров в разное время.
Начнем с самого начала: как выглядел первый тираннозавр? В любом случае перед нам животное гипотетическое, потому что найти представителя истинной исходной популяции, ответвившейся от других тероподов, практически невозможно. Считается, что древнейшие популяции предков тираннозавров были небольшими в смысле количества особей и в целом просуществовали по эволюционным меркам недолго, так что вряд ли они появятся в палеонтологической летописи. Более того, кладистика не позволяет идентифицировать подобные группы; если бы мы когда-либо нашли такое животное и поместили его в филогению, оно бы считалось просто ближайшим родственником всех прочих тираннозавров, но не предком. Отчасти досада палеонтологов в общении со СМИ связана с тем фактом, что каждое открытое животное либо объявляется чьим-либо предком, либо недостающим звеном между двумя существами, в то время как оно не является ни тем, ни другим (точнее, мы не знаем, является ли оно тем или другим). При определении того, каким мог быть предковый тираннозавр, мы неизбежно представляем некий обобщенный образ, созданный на основании наших знаний о соседних с ним линиях.
Предковый тираннозавр должен был весьма походить на известные нам ранние формы – животных вроде гуаньлуна, стоксозавра и эотираннуса (рис. 12а), – но при этом был близок к древнейшим представителям групп, ближайших к тираннозаврам. К ним относятся компсогнатиды и, возможно, либо орнитомимозавры, либо аллозавроиды (поскольку не совсем понятно, отделились тираннозавры до компсогнатидов или после). На самом деле это может оказаться не так уж важно, поскольку мы ожидаем, что животные были очень похожи друг на друга: как-никак эти группы только начали расходиться друг с другом. И действительно, если посмотреть, скажем, на дилуна и китайского компсогнатида синокаллиоптерикса, то на первый и даже на второй взгляд между ними нет почти никаких разительных или даже явных различий, и было бы простительно посчитать их представителями одной клады. Это связано с тем, что они являются ранними представителями своих групп и еще не слишком дивергировали относительно предковой формы.
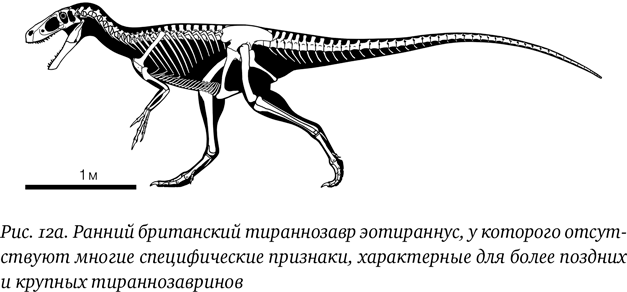
Предок тираннозавров должен был быть небольшим: возможно, всего около 2 м длиной (половину которых составлял хвост), и в этом нет ничего удивительного. Небольшой размер – обычная черта для ранних представителей многих эволюционных групп, поскольку эволюция зачастую идет более быстро у мелких животных (у которых жизнь поколения короче, а численность выше), а вымирание обычно настигает первыми более крупные виды (давая возможность оставшимся образовывать новые виды). Предка тираннозавра должно было отличать от других тероподов при ближайшем рассмотрении то, что он, несомненно, обладал отчетливыми признаками тираннозавров, или же это могла быть некая переходная или промежуточная между другими тероподами форма и даже самый ранний из «настоящих» тираннозавроидов.
Предковое животное должно было иметь простые нитевидные перья. Как отмечалось ранее, перья или какая-то их разновидность могли быть унаследованными от предков для всех тероподов вообще, но, учитывая их наличие у таких животных, как дилун, и во всех линиях, возникших позже, они наверняка появились еще до отделения тираннозавров. Однако здесь оказываются важны зубы – необычная зубная система тираннозавров, в том числе более толстые предчелюстные зубы – могли служить отличительными признаками этих самых первых представителей группы. Их истинное происхождение – загадка, тем не менее их развитие, вероятно, послужило основой эволюции под действием естественного отбора: форма и расположение зубов изменились в результате мутации внутри какой-то небольшой популяции, состоящей из немногих особей. Вариации в биологии естественны и неизбежны: как бы вы ни походили на своих родителей или братьев и сестер, всегда имеются какие-то отличия. Новые комбинации генов и изменения в них при мутациях приводят к тому, что все индивиды немного отличаются от своих ближайших родственников. Потом какое-нибудь событие (засуха, наводнение или появление нового хищника) начинает оказывать давление на хищника, и модификация зубной системы дает ему преимущество, способствуя распространению и умножению, или, возможно, изменение происходит в какой-то изолированной популяции и потому быстро распространяется. Давление отбора (конкуренция с другими хищниками или недостаток пищи) приводит к передаче и распространению этого признака, потому что особи, им не обладающие, выживают хуже. Такие изменения редко происходят изолированно и могут сопровождаться появлением нового поведения, и другие элементы анатомии тоже вполне могут адаптироваться к меняющимся условиям.
Популяция, в которой таких изменений происходит все больше, отделяется и обособляется от своих родственников (особенно если она изолирована или если какая-то катастрофа полностью уничтожает сородичей) и со временем, вероятно, станет полностью отдельной в эволюционном смысле. Мы считаем такую популяцию филогенетически отдельной линией и можем выделить анатомические особенности, которые отличают и обособляют ее от родственников. Представители разделившихся групп фактически перестают скрещиваться с исходной популяцией в связи с ее вымиранием, географической отдаленностью, каким-либо генетическим отличием, которое мешает скрещиванию, или какими-либо изменениями в поведении, приводящими к тому, что две группы животных больше не признают друг в друге потенциального брачного партнера.
Мы можем отследить изменения в анатомии этих животных по ископаемым останкам и соотнести их с определенными функциями. Как нам известно, наличие арктометатарзуса (сплюснутой средней кости стопы) и более жесткая щиколотка хорошо коррелируют с эффективным и даже быстрым бегом, а поскольку этот признак развивался многократно в различных группах тероподов, предполагается, что в каком-то смысле его было «просто» приобрести с эволюционной точки зрения, хотя мы точно не знаем, с чем был связан отбор по этому признаку. Привело ли преследование быстроногой добычи к появлению более быстрых и эффективных хищников, требовалось ли им мигрировать по своим территориям, чтобы не отставать от стад добычи, и это запустило отбор на эффективность движения или им угрожали другие хищники и преимущество получали те, кто бегал быстрее?
На эти вопросы ответить нелегко, отчасти потому, что мы не можем точно идентифицировать предков и поэтому вынуждены смотреть на изменения у животных, близких к разделению групп или к ранним представителям, определяя, что могло происходить, когда эти изменения возникали. Если нам удастся объединить это знание с информацией о тенденциях в других областях (изменения климата, появление новых групп животных, другие анатомические приспособления, миграции между континентами) и нашим знанием дат и времен для событий эволюционного ветвления, мы сможем начать складывать цельную картину.
Отрастить большую голову
Легче всего идентифицировать изменения, связанные с хищным образом жизни. Почти все элементы организма выполняют более одной функции (рога барана, к примеру, в первую очередь используются самцами для драк за самок, но также могут служить для отпугивания хищников и играть небольшую роль в охлаждении мозга благодаря проходящей через них крови), но, по-видимому, для большинства элементов в начале их эволюции основной или по крайней мере доминирующей является одна функция. Когда речь идет об отличительных признаках, тесно связанных с хищничеством, они относительно однозначны. Что верно, то верно, ноги носят хищника одинаково, охотится он или нет, но можно предсказать, что самой важной частью его жизни будет ловля добычи, где каждый промах может стать последним. Следовательно, этот фактор будет оказывать более заметное влияние на выживание данного животного, чем многие другие. Изменения в ключевых элементах тела, таких как голова, передние и задние конечности, и поддерживающих структурах (например, шея) могут, таким образом, быть показательными в этом отношении.
Изменения в размере и форме головы были очевидной и главной модификацией (рис. 12b). Не считая гребней на головах процератозавридов, влиявших, скорее, на внешний вид, череп у ранних форм был в целом довольно небольшим. У гуаньлуна череп выглядел пропорциональным по отношению к остальному телу: он не казался какой-то неестественно увеличенной чудовищной конструкцией, как у тираннозавра, но смотрелся бы уместно практически на любом тероподе. Однако у более поздних тираннозавроидов, например ютираннуса, голова стала больше по всем параметрам (в особенности по высоте) и начала выглядеть как доминирующая часть тела животного. Эта особенность была сходной у алиорама и дасплетозавра, а у тираннозавра и тарбозавра голова поистине заняла господствующее положение. Она не просто стала больше и выше (за исключением алиораминов), но также более массивной и прочной. У ранних представителей группы опорные кости, задающие очертания черепа, в особенности вокруг отверстий по бокам, были довольно тонкими – между ними много места, – но у тираннозавринов они стали куда более толстыми и прочными. Аналогично при переходе от ранних форм к поздним зубная кость и мандибула в целом сделались более глубокими и прочными, и особенно увеличились они у тираннозавринов. В общем, в голове стало значительно больше костной массы, чем раньше, и, хотя одно это не делало ее крепче, все же предполагается, что данные животные развивали все более мощный укус, а также череп, способный выдержать большие нагрузки.
Это изменение происходило вместе с другими, более явно проявляющимися в зубах. Когда мы переходим от первых тираннозавров к более поздним, становятся очевидны две тенденции: количество зубов на верхнечелюстных и зубной костях уменьшается, так что в челюстях оказывается меньше зубов, и эти зубы увеличиваются, в особенности в ширину. В целом зубы хищных тероподов сплюснуты с боков, так что оказываются довольно тонкими, и, хотя они определенно не «кинжалообразные», как периодически утверждается, в поперечном сечении они относительно узки. У альбертозавринов и особенно у тираннозавринов зубы на верхнечелюстных и зубной костях очень толстые, и если не полностью круглые в сечении, то близки к тому. Эти два изменения (более толстые зубы и меньшее их количество) также, по-видимому, коррелируют с увеличением силы укуса (как и более короткие шеи позднейших видов, служившие лучшей опорой для тяжелой головы) и с изменениями, произошедшими с черепом. Это предполагает, что отбору подверглись все эти черты, изменяя способ использования головы при укусе и увеличивая способность совершать более мощные укусы.
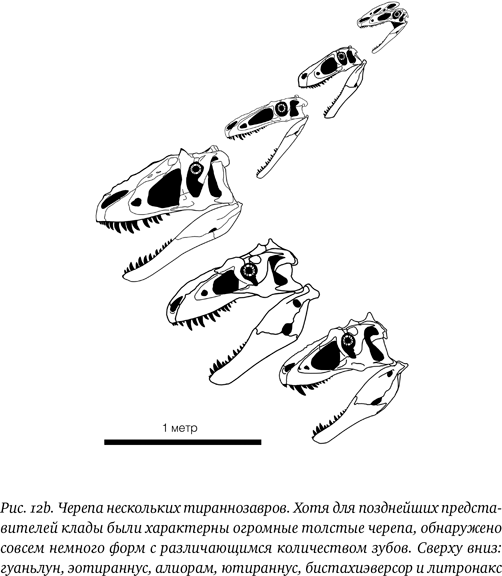
Что любопытно, черепа ранних тираннозавроидов по форме похожи, скорее, на черепа детенышей тираннозавридов, а это предполагает, что форма головы более поздних видов стала результатом изменений в схемах роста по мере того, как животные становились все крупнее. Также это подразумевает, что экология молодых тираннозавринов могла быть похожей на экологию взрослых тираннозавроидов, поскольку похожая форма, вероятно, коррелирует с похожим стилем укуса.
Что вытягивать – руки или ноги?
Мы движемся дальше по телу, и руки демонстрируют тенденцию, противоположную голове, становясь со временем короче. У первых тираннозавров руки были относительно длинными и имели по три пальца, но при переходе к тираннозавридам руки уменьшались, пока не превратились в короткие отростки у тираннозавринов. Более того, притом что когтевые фаланги сохранились и вроде бы продолжали нести острые когти, они стали менее изогнутыми, чем у более ранних форм, и их основания редуцировались. Следовательно, эти животные обладали меньшей силой захвата, и места прикрепления связок у них стали меньше. В сочетании с относительной притупленностью когтей эта черта подразумевает, что руки стали использоваться меньше, чем у ранних представителей клады тираннозавров. И это, по-видимому, снова связано с возрастающей ролью головы: в конце концов, если голова выполняет всю работу по убийству добычи и питанию, руки становятся менее полезными. Отбор может действовать так, чтобы руки уменьшались, потому что неиспользуемые большие руки – нечто вроде мертвого груза: на их рост и поддержание тратятся ресурсы, при этом они болтаются без дела, не принося особой пользы.
Интересно, что нечто похожее произошло в другой группе крупных тероподов. У абелизавров с южных континентов мелового периода руки также редуцировались, в то время как появился более специализированный и необычно короткий плоский череп. У этих животных руки редуцировались даже сильнее, чем у тираннозавров, и, хотя на кистях сохранилось по четыре пальца, они стали крошечными, и на них даже исчезли или максимально упростились когти, а особенно простой и почти дегенеративной сделалась плечевая кость. Следовательно, это предполагает, что в обеих данных группах эволюционный процесс, который привел к изменениям в черепе, был связан с уменьшением использования рук и в итоге с их изменением.
Даже у самых первых тираннозавроидов были длинные по сравнению с туловищем ноги, и этот признак сохранялся во всей линии. Последние из тираннозавринов также сохранили его; они обладали пропорционально длинными бедренной и плюсневыми костями, и это отличает их от всех остальных нептичьих тероподов, кроме орнитомимозавров. Учитывая тесные родственные связи между орнитомимозаврами и тираннозаврами, это, возможно, характерный признак для исходного отделения этих линий от базальной ветви. Одной из ключевых предковых черт у тираннозавров было удлинение частей задних конечностей, что повышало эффективность и, вероятно, скорость этих животных. Это привело к тому, что орнитомимозавры стали высокоспециализированными в области больших скоростей, как предполагается, отчасти для обеспечения бегства, поскольку более поздние формы, по-видимому, в основном перешли к травоядности. У тираннозавров же подобный признак, вероятно, указывает на новую стратегию преследования добычи, связанную с изменениями головы и рук, хотя, несомненно, на ранних этапах своей истории некрупные тираннозавры также должны были быть уязвимы для более крупных тероподов и тоже могли нуждаться в способности быстро бегать.
В какой-то момент плюсневые кости у тираннозавров тоже изменились, породив характерную сплюснутую среднюю кость и странную треугольную форму этой части стопы. Этот признак не наблюдается у ранних тираннозавров и не развивается во всей полноте до появления тираннозавридов. Он явственно отсутствует даже у значительно более позднего ютираннуса, а это предполагает, что подобная черта возникла в эволюции тираннозавров относительно поздно и «заработала» только у крупных видов. Поскольку такая специализированная стопа, как известно, помогает уменьшать энергетические потери, возможно, это изменение было вызвано увеличением размеров тираннозавров; отбор в данном случае шел в направлении уменьшения усилия, требуемого для поворотов многотонного тела на относительно высоких скоростях.
Больше значит лучше
Еще одна очевидная тенденция – увеличение размера тела у тираннозавров. Процератозавриды и базальные тираннозавроиды были небольшими (менее 5 м общей длины), позднейшие тираннозавроиды стали заметно больше (около 8 м), альбертозаврины и алиорамины – еще больше (10 м), а тираннозаврины сделались совсем огромными. Увеличение размеров тела со временем вполне обычно для многих эволюционных линий: отчасти из-за того, что фокус вымирания сосредоточен на самых крупных видах, большинство клад начинают свое развитие с небольшого размера. Это означает, что чаще всего единственный возможный путь – становиться больше, хотя, разумеется, у динозавров увеличение размера было выражено сильнее, чем у большинства линий позвоночных, и в итоге такая тенденция дала самых огромных наземных животных всех времен среди крупнейших завроподов. Различные линии динозавров вырастали в гигантов из мелких исходных животных (упомянем лишь немногих: аллозавры, цератопсы, титанозавровые завроподы и овирапторозавры). Точные причины этих изменений в настоящее время неизвестны, но я подозреваю, что для тираннозавров по крайней мере такой переход является устойчивой эволюционной тенденцией, поскольку коррелирует с постоянно увеличивающейся силой укуса и соответствующими адаптациями.
Однако эта тенденция не абсолютна, поскольку, во-первых, такие животные, как тарбозавры, жили рядом с более мелкими формами вроде алиорама. Еще более примечательно то, что нанукзавр с севера Канады был намного меньше, чем мы предсказали бы, опираясь на размеры его родственников. Имея вполне солидные размеры (оценочно около 7 м в длину), по меркам других тираннозавринов он был маленьким, чуть ли не карликом. Что вызвало такое отклонение от тенденции увеличения размеров? Более того, уменьшение размера в данном случае – двойной сюрприз, поскольку нанукзавр обитал на Крайнем Севере, и мы могли бы ожидать проявления эволюционного феномена, называемого правилом Бергмана. Это правило говорит, что животные, обитающие в более холодных северных широтах, обычно крупнее живущих ближе к югу. В северных широтах более крупный размер тела помогает животным сохранять тепло (у них меньше отношение площади поверхности к объему), так что северные виды или популяции видов с широким ареалом в высоких широтах обычно крупнее, чем в низких. Нанукзавр являлся тираннозаврином и обитал на севере, так почему он был таким маленьким? Вероятная причина заключается в том, что он жил изолированно на относительно небольшом участке суши, а при ограниченных доступных ресурсах на небольшой территории и сама популяция должна была становиться меньше, и особи – уменьшаться в размерах под действием отбора. Это «островное правило», и здесь оно, по-видимому, пересилило ожидаемую тенденцию и привело в итоге к появлению небольшого тираннозаврина.
Встает вопрос об ограничениях эволюции. Размеры тела явно не могут увеличиваться бесконечно: фундаментальные ограничения, такие как физические свойства костей и мышц, и физиологические ограничения, связанные с достаточным снабжением тканей кислородом или передачей нервных сигналов, в итоге препятствуют животным становиться еще крупнее. В случае нанукзавра имели место конфликтующие направления отбора: крупные размеры дают возможность лучше сохранять тепло, но это преимущество уравновешивается и даже перевешивается ограничениями жизни на острове и небольшим количеством ресурсов. Также отсюда вытекают проблемы, связанные с изменением существующей формы: эволюция может работать только с тем, что есть, и если у нее имеется позднейшая форма тираннозавра с маленькими ручками и всего двумя пальцами, внезапное смещение отбора, скажем, в сторону благоприятствования тонким манипуляциям с добычей при помощи рук будет затруднено. В таких случаях специализация может затронуть какую-либо другую часть организма, в том числе возможно альтернативное решение проблемы, такое как перемещение в другую экологическую нишу или даже вымирание вида. Утерянные элементы могут восстанавливаться, но, если они отсутствовали в течение долгого времени, гены для них тоже могут оказаться утрачены, что сделает их возвращение без множества совместно возникших мутаций значительно менее вероятным. Отсюда вытекают наблюдаемые нами эффекты, такие как неоднозначность признаков: они могут быть приспособлены для выполнения многих функций или представлять собой компромисс между разнонаправленными давлениями отбора. Результатом могут быть поразительные приобретения и модификации, такие как бивни слонов, которые можно использовать в битвах с другими слонами или чтобы отбиваться от хищников, копать ямы для воды, снимать кору с деревьев и т. д.; на самом деле ни одно направление отбора не побеждает другие полностью, хотя половой отбор и внутривидовая конкуренция, по-видимому, доминируют, по крайней мере в отношении бивней.
В биологии всегда имеют место подобные удивительные исключения и отклонения. При достаточном времени и разнообразии где-нибудь в кладе почти неизбежно появляется эволюционное исключение: примером могут служить гигантские нелетающие птицы, безволосые млекопитающие, безногие ящерицы, гигантские сухопутные черепахи и лягушки, которые превращаются из головастиков во взрослых особей. Даже если изъять подобные случаи из уравнения, остаются некоторые очевидные эволюционные тенденции для тираннозавров, и выявление их создает наглядное представление о том, как они менялись и что означали эти изменения для их жизни. В следующей части мы переходим к поведению и экологии тираннозавров: как они вписывались в свои экосистемы и взаимодействовали с представителями собственного вида и других видов.
Назад: Глава 10 Физиология
Дальше: Часть III Экология

