Книга: Хроники тираннозавра: Биология и эволюция самого известного хищника в мире
Назад: Глава 7 Туловище
Дальше: Глава 9 Внешний вид
Глава 8
Конечности
Наконец, мы добрались до выступающих элементов тела тираннозавров, а именно рук и ног, более формально называемых передними и задними конечностями. У тираннозавров передние конечности не используются для ходьбы, так что обиходное название «руки» вполне для них подходит (учитывая, что слово «ноги» часто используется, чтобы показать, что животное с помощью них ходит). Конечностям для движения нужна какая-то опора, поэтому они соединяются с главной структурной и опорной конструкцией организма – позвоночником. Каждая конечность сочленяется с совокупностью костей, присоединяющих ее к остальному животному – эти структуры называются «поясами конечностей». В случае передних лап это кости плечевого (или грудного) пояса, а таз (или тазовый пояс) исполняет ту же функцию для задних конечностей.
Плечевой пояс начинается с лопатки – длинной тонкой кости, лежащей поверх многочисленных грудных ребер (она частично с ними соединена мускулатурой, удерживающей руку) и идущей вниз, к груди. Здесь меньшая кость, коракоид, соединяется с лопаткой, а между ними помещается сустав плечевой кости; к лопатке и коракоиду также крепятся мышцы руки. Посередине тела, связывая два плеча, располагается кость, называемая вилкой или вилочковой костью, которая состоит из сросшихся ключиц (у птиц она более известна как вилочка или дужка).
В истории изучения динозавров у вилки долгая и многотрудная судьба. В начале XX в. в фундаментальном научном труде о происхождении птиц тщательно и подробно была проанализирована доступная в то время информация и сделан вывод: тероподы имеют больше общего с птицами, чем с любой другой группой животных, но далее гипотеза о них как о предках птиц решительно отбрасывалась на том основании, что у них нет вилочковой кости. В то время господствовала идея, что если какая-то структура утрачена, то она не может вернуться, и у других рептилий ключицы были, а у динозавров нет. Следовательно, если динозавры утратили ключицы, то они не могли стать предками птиц, у которых вилочковая кость наличествует. Эта идея была неверна в целом (утерянные черты могут возвращаться), но возникла она также из-за недостатка данных. Прошло время, обнаружились хорошо сохранившиеся скелеты животных, и теперь мы знаем, что у многих динозавров, включая тираннозавров, вилка была. Более того, вилка расположена между коракоидами, и для тираннозавров это означает, что руки были расположены ближе к середине груди, а не по бокам от туловища, как часто изображалось на старых реконструкциях.
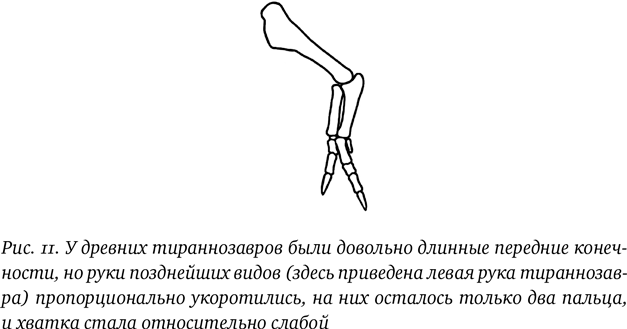
Если рассматривать переднюю лапу сверху вниз, то первой идет плечевая кость, потом локтевая и лучевая, и они составляют бо́льшую часть руки (рис. 11). Эти кости ничем не примечательны в сравнении с аналогичными костями других тероподов, однако со временем они значительно уменьшаются, и у более поздних тираннозавров стали пропорционально короче. Но они не изменились до такого состояния, когда теряется функциональность, в них сохранились основные сочленения и группы мышц, и руки должны были оставаться довольно сильными, несмотря на небольшую длину. Теории о функционировании передних конечностей у позднейших тираннозавров чрезвычайно противоречивы, и ниже мы более подробно рассмотрим некоторые из гипотез.
Кисти передних конечностей также обычны для тероподов, в них имеется группа маленьких косточек запястья, потом длинные кости пясти (метакарпальные), потом кости пальцев (фаланги). У большинства животных, включая тираннозавров (но не человека), количество костей в пальцах различается. Люди также несколько необычны тем, что имеют по пять пальцев на каждой руке – таково было исходное количество пальцев у древних предков тетраподов, позже это число уменьшилось у множества групп. Самые первые тероподы обладали четырмя пальцами, большинство тремя, а знаменитый тираннозавр и другие тираннозаврины, алиорамины и альбертозаврины – только двумя. У некоторых экземпляров имелась лишняя пястная кость (а не лишний палец, как иногда пишут), так что в кисти могло быть три длинных кости, но пальцев оставалось только два. Дополнительная кость, по-видимому, появлялась и исчезала, это предполагает, что данный признак не находился под сильным эволюционным давлением и был, по сути, рудиментарным. Пальцы тираннозавров несколько более необычны, чем пальцы большинства тероподов, у которых первый палец довольно толстый и немного отставленный; хотя вряд ли он был таким, как человеческий большой палец, но по крайней мере чуть больше походил на него, чем другие пальцы. Такое строение наблюдалось у животных типа гуаньлуна, но у позднейших тираннозавров два пальца похожи друг на друга по общей форме и пропорциям и первый палец не отставлен.
Идентификация пальцев тероподов, вероятно, является областью наибольших разногласий и споров, чем любая другая. У птиц пальцы, формирующиеся на эмбриональной стадии, – это II, III и IV (т. е. три средних пальца предковой пятипалой кисти тетрапода, если начинать считать с большого), однако позже они уменьшаются и срастаются, и у большинства взрослых птиц остается два пальца, и те зачастую представляют собой только лишь небольшие выросты. Этого в целом и следует ожидать: когда у животных пальцы уменьшаются или утрачиваются, первым неизменно уходит пятый, а за ним первый. Однако у тероподов, хотя первым утрачен пятый палец (как можно увидеть по наличию четырех пальцев, а также укороченной пястной кости у самых ранних форм), следующим исчезает, по-видимому, четвертый. Это указывает на то, что в линии тероподов, ведущей к птицам, наличествовали пальцы I, II и III, что отличается от птичьих II, III и IV.
Этим парадоксом иногда пользуются как аргументом, ставящим под сомнение происхождение птиц от тероподов, но такому строению есть несколько возможных объяснений. Во-первых, могло произойти то, что называется сдвигом рамки, когда программа развития кисти фактически сдвигается на один палец, и второй развивается как первый, третий как второй и т. д. Такой феномен развития встречается в природе, а в пальцах случаются всевозможные аномалии развития. Их можно увидеть, например, у людей с лишними пальцами или с «зеркальной кистью», когда на каждой стороне кисти имеется по большому пальцу. Такое событие вполне могло произойти в какой-то момент на эволюционном дереве тероподов или древнейших птиц, однако это невозможно установить, пока мы не найдем идеально сохранившихся динозавров в эмбриональной стадии или просто очень маленьких (что теоретически вполне допустимо). Возможно и другое объяснение данной черты: у тероподов действительно наличествовали пальцы II–IV, и они утратили I палец, а II превратился в нечто, более похожее на I. У странного древнего цератозавра, названного лимузавром (Limusaurus), из среднеюрских отложений Китая II палец был похож на большой, но также имелся и значительно редуцированный I палец. Можно предположить, что данные изменения в эволюции тероподов произошли относительно рано, и I палец в самом деле был утрачен, так что кисть тероподов соответствует строению, наблюдаемому у птиц: пальцам II–IV.
Вне зависимости от того, какая из этих схем (если вообще работала какая-то из них) ответственна за различия между большинством тероподов и птицами, в данной ситуации, очевидно, возникают проблемы при описании пальцев таких животных, как тираннозавры, – какие у них были пальцы: I–III или II–IV? Большинство исследователей сходится на первом варианте, хотя у двупалых видов должен был исчезнуть также и III палец и остаться только I и II. Как уже упоминалось, число составляющих пальцы костей варьирует. У тираннозавров I палец состоял из двух костей, II – из трех, а III – из четырех. Первый палец был заметно короче других, а второй и третий имели почти одинаковую длину – в третьем есть дополнительная кость, но очень короткая и мало прибавляющая к длине. Эти оценки включают последнюю – специализированную – кость пальца, которая формирует основу для когтя и называется когтевой фалангой.
Когтевые фаланги имеют крюкообразную форму и поддерживают кератиновый коготь (более подробно рассмотренный ниже). Сама кость дает представление о форме когтя в целом, но, возможно, она не так тесно связана с внешним когтем, как могло бы показаться, и между ними могут обнаружиться существенные различия и в общем размере, и в точной форме. Некоторые когти, вероятно, имели значительно больший размер и были сильнее изогнуты, чем можно предположить на основании подлежащей когтевой фаланги. По бокам когтевой фаланги до кончика когтя проходят желобки, однако их функция неясна. Существует предположение, что они могли быть местами крепления кератина или что по этим желобкам проходили кровеносные сосуды, питающие кератиновый когтевой чехол, что позволяло когтю расти.
Если рассматривать сочленения руки, то плечо позволяло достаточно свободное вращение, но другие суставы были более ограничены в движениях. Как у человека (а также большинства животных), локоть являлся простым шарнирным суставом и позволял нижней части руки прижиматься к плечевой части. Однако вращение в локте у динозавров в целом и тероподов в особенности было ограничено, а кисти обращены друг к другу, как будто животное собиралось аплодировать. Многочисленные реконструкции отображают руки неверно, показывая ладони обращенными вниз и сильно согнутыми в запястьях (что часто называют «кроличьими лапками»). У многих более поздних тероподов запястья действительно обладали большой степенью гибкости, но в другой плоскости – они как будто пытались заставить внешние пальцы указывать назад вдоль локтевой кости в сторону локтя. Это было ключевой модификацией для полетного взмаха у птиц, однако у тираннозавров в этом месте движения были значительно более ограниченными.
Пальцы могли сгибаться, как у человека, и каждый сустав обладал вполне приличной гибкостью. Тем не менее в пальцах тираннозавров структурных элементов меньше, чем в пальцах человека, а при том, что в каждом пальце имелось разное количество суставов, тираннозавры не были способны сложить кисть в нечто вроде кулака. Первые тираннозавры обладали хорошей гибкостью суставов и мощной хваткой. Это подтверждается глубокими ямками по бокам фаланг, к которым крепились связки, присоединявшие следующую кость пальца. Чем глубже ямки, тем более прочно крепились связки и тем сильнее была хватка. Со временем ямки стали менее глубокими, и это показывает, что у более поздних видов хватка стала слабее. Основание когтевой фаланги часто расширено, что дает место для крепления мышц и в особенности связок, усиливает эту область и позволяет когтям протыкать то, что зажато в кисти (вероятно, добычу).
Эти длинные ноги
Второй пояс конечностей – тазовый – состоит из трех основных элементов: подвздошной, лобковой и седалищной кости, причем первая у взрослых особей полностью срасталась с крестцом, создавая структурно жесткую станину для ног. Тираннозавры довольно необычны своим «сплюснутым» тазом: если смотреть сверху, то кажется, что кости почти сходятся посередине, около невральных отростков крестцовых позвонков, отчего таз приобретает характерный вид. Почему это так, не вполне ясно; данная черта обнаруживается даже у таких ранних форм, как гуаньлун, в те времена, когда животные были еще небольшими и обладали менее специализированными ногами, чем позднейшие гиганты, и все же она сохранилась и даже развилась у более поздних тираннозавридов. Подвздошная кость служит местом крепления для разнообразных групп мышц, управляющих бедром, особенно перемещающих его вперед во время передвижения; главными же отводящими мышцами, как отмечалось ранее, являются те, что присоединялись к основанию хвоста.
Стержневидные лобковые и седалищные кости сходились с обеих сторон на средней линии тела и срастались, лобковая кость при этом была направлена вперед, а седалищная – назад. К подвздошной кости крепилось несколько групп мышц, отводящих бедро назад, а лобковая кость помогала поддерживать тело и служила местом крепления некоторых других ножных мышечных групп. У более поздних тираннозавров конец лобковой кости стал чрезвычайно большим, а седалищная кость немного уменьшилась, но причины этих изменений также неизвестны. Там, где три кости таза встречаются, они образуют край вертлужной впадины, представляющей собой большое отверстие в тазу, куда входит головка бедренной кости. На самом деле это отверстие заметно крупнее головки бедренной кости, но оно должно было быть выстлано большим количеством хрящевой ткани, предотвращающей истирание костей друг о друга при движении животного. Наверху вертлужной впадины имеется похожее на полку расширение нижней части подвздошной кости, помогавшее удерживать головку бедренной кости на месте.
Сама бедренная кость довольно типична для тероподов и имеет Г-образную форму за счет длинного стержня с полусферической головкой, почти перпендикулярной стержню и точно входящей в вертлужную впадину. Головка обеспечивает сочленение и позволяет бедру двигаться, а прямой стержень принимает вес животного. Именно такое устройство кости ожидается от животного с вертикальной посадкой корпуса (т. е. с ногами, расположенными прямо под туловищем), и также оно наблюдается у птиц и млекопитающих, но отличается у ползающих животных, таких как ящерицы и тритоны, у которых вес не приходится на продольную ось бедренной кости. Это один из ключевых признаков, показывающих, что динозавры в основном были животными с высокой посадкой тела, а не ящерицеподобными пресмыкающимися, и это ясно видно у тираннозавров.
Двигаемся вниз по конечности: далее идут крупная большеберцовая и несколько меньшая малоберцовая кости. У птиц малая берцовая кость практически отсутствует (как и у поздних птерозавров), а некоторые кости щиколотки срослись с большеберцовой костью и значительно упростили устройство сложной нижней части ноги, но у тираннозавров и у других нептичьих тероподов эти кости сохранились как отдельные элементы. На передней поверхности основания большеберцовой кости есть большое углубление для таранной кости. Это самая крупная кость в щиколотке тероподов. У нее имеется огромный треугольный вырост, располагающийся перед большеберцовой костью и придающий нижней части ног тероподов характерный вид. Как и запястье, предплюсну образует группа костей. Основание таранной кости похоже на роликовый шарнир, позволяющий стопе сгибаться и разгибаться, но в остальном довольно жесткий – в отличие от человека тираннозавры не особенно рисковали вывихнуть лодыжку.
Дальше идут пять плюсневых костей (метатарзальных). Первая, соответствующая нашему большому пальцу, очень мала и расположена примерно посередине второй плюсневой кости – это крошечный треугольный выступ, к которому крепится маленький палец, состоящий всего из одной обычной и одной когтевой фаланги. Это, по сути, эквивалент рудиментарного «прибылого» пальца, имеющегося у многих животных, и он представляет собой сильно редуцированный большой палец. Пятая плюсневая кость уменьшена еще сильнее – это короткая, тонкая и слегка изогнутая косточка на наружной стороне стопы, и на ней совсем нет пальца. Притом что первый палец сильно редуцирован, а пятый практически утрачен, на стопе остается три функциональных пальца для ходьбы, и поэтому тероподы оставляют трехпалые отпечатки ног.
Главные плюсневые кости представляют собой длинные прямые стержни, внешние кости (II и IV) немного расходятся к концу, а потому пальцы расставлены в стороны. У позднейших тираннозавров, однако, стопа довольно сильно изменена – центральная из трех плюсневых костей имеет очень тонкую проксимальную (верхнюю) часть, и она «зажата» между нормальными II и IV плюсневыми костями. Такое строение стабилизирует стопу, поскольку средняя плюсневая кость оказывается ограниченной двумя другими в боковом движении при ходьбе и особенно беге. При быстром перемещении на такое движение расходуется большое количество энергии, так что данная адаптация помогает сделать животное немного быстрее, но, что более важно, оно становится более ловким и эффективным. Такое строение наверняка помогало животному тратить меньше усилий при переходах на большие расстояния, и это хорошо сочетается с уже рассмотренными нами более длинными бедренными костями и длинными плюсневыми костями в целом: у тираннозавров были широкий шаг и экономичная походка.
Плюсневые кости тем не менее в норме не соприкасались с землей. Люди в этом довольно необычны: мы ходим на полной стопе, и с землей у нас контактируют и пальцы ног, и плюсневые кости. Тираннозавры же были похожи на большинство животных, которые ходят только на пальцах (или в особых случаях, как у лошадей, на кончиках пальцев), используя плюсневые кости для эффективного увеличения длины ноги.
Как и на передней конечности, в каждом пальце ноги имелось разное количество фаланг, и все пальцы оканчивались когтевой фалангой. Второй состоял из трех элементов, включая когтевую фалангу, третий – из четырех, а четвертый – из пяти. И снова, как и в кисти руки, хотя количество костей различалось, пальцы оказывались примерно одинаковой длины – только третий был немного длиннее остальных. Когтевые фаланги по форме были в основном коническими и совсем не так сильно изогнутыми, как в кисти: ведь они в первую очередь должны были обеспечивать взаимодействие с землей.
Как они ходили
Чтобы узнать, как передвигались тираннозавры, требуется сложная совместная работа различных областей науки. Мы можем реконструировать скелет и основные группы мышц тираннозавра с некоторой степенью точности, и представление об общем строении этих животных (включая такие вопросы, как расположение и размеры легких и воздушных мешков) позволяет понять, где находился центр масс. Для сохранения равновесия движущемуся тираннозавру было необходимо удерживать центр масс практически над ногами, иначе бы он опрокинулся вперед или назад. На этом основании мы можем при помощи компьютерных моделей животных попробовать определить схемы движения и сопоставить результаты с данными по отпечаткам ног.
Отдельные отпечатки ног динозавров и цепочки следов – довольно обычные находки, но проблема в том, чтобы определить, какому виду животных мог принадлежать этот след. Отпечатки различаются в зависимости от оставившего их животного, а также от того, как оно двигалось, какой в тот момент была поверхность, и от любых изменений вроде эрозии, случившихся впоследствии. В результате имеется множество следов, по которым нельзя с уверенностью отличить гадрозавра от теропода, а отнести их к какой-либо кладе, не говоря уже о роде, практически невозможно. Но даже при этом существует пара-тройка следов, которые, как мы точно знаем, были оставлены тираннозаврами, поскольку это явно огромные следы трехпалых тероподов, относящиеся к тому времени и месту, когда из тероподов в данном месте обитали только тираннозавриды.
Разумеется, даже если вы знаете, что перед вами след тираннозавра, это не продвинет его идентификацию, учитывая сложности, описанные выше. И все же существует по крайней мере несколько относительно устойчивых взаимосвязей между размером и формой стопы динозавра, расстоянием между отпечатками и тем, как этот динозавр двигался. Длина стопы дает хорошее представление о размере животного, а расстояния между отпечатками демонстрируют, насколько широк был его шаг, и таким образом указывают на скорость передвижения. Добавим эти данные к нашей модели движения, и, понимая работу мышц, мы сможем выстроить для тираннозавров картину набора скорости, скоростей разворота и предельных скоростей, хотя точные значения всегда останутся недостижимыми.
Что касается фактического паттерна следов, несомненно, что тираннозавры были двуногими и, по-видимому, переставляли ноги поочередно. Это может казаться очевидным, но, хотя четвероногие животные имеют разнообразные схемы шагов при разных скоростях и в разное время (сравните, скажем, идущего жирафа, лошадь в легком галопе и кошачий шаг), даже двуногие способны передвигаться по-разному. Мелкие птицы, грызуны наподобие тушканчиков, кенгуру и валлаби прыгают, синхронно отталкиваясь задними ногами, а некоторые животные исполняют нечто вроде подскока, где после одного короткого шага идет длинный, отчего получается странный асимметричный бег (так, например, делают грифы), так что тираннозавры на самом деле ходили вполне нормально.
Бедренная кость у тираннозавров достаточно длинная по сравнению с другими тероподами, однако нижняя часть ноги и плюсневые кости пропорционально оказываются еще длиннее. Животные, которые быстро бегают, зачастую имеют короткую бедренную кость, поскольку, чтобы сделать полный шаг, бедро следует пронести вперед по дуге. Если бедро короткое, можно за меньший промежуток времени сделать больше шагов. Потеря в длине шага, следующая из укорочения бедра, компенсируется длинной большеберцовой костью и/или длинными плюсневыми костями. Посмотрите на ногу лошади или гончей – при общей значительной длине ног и скоростях, развиваемых этими животными, бедро у них относительно короткое и составляет небольшую часть длины ноги. У тираннозавра бедренная кость длинная, так что, возможно, он не мог очень быстро переставлять ноги, но это предположительно компенсировалось более длинными, чем обычно, нижними частями ноги, и такое строение указывает на то, что тираннозавры все же должны были быть относительно быстрыми ходоками и бегунами.
При любых скоростях тираннозавры, по-видимому, переносили одну ногу за раз, и хотя более крупные формы могли быть слишком массивными для бега, они все-таки были способны передвигаться достаточно быстро: большая длина шага может скомпенсировать многое. В нормальном стоячем положении ноги должны были ставиться бок о бок, а бедренные и большеберцовые кости – располагаться почти вертикально и образовывать опорный столб по отношению к земле, но при ходьбе эти кости, вероятно, переносились ближе к срединной линии тела животного. При каждом шаге нога должна была немного уходить под туловище, а стопа ставиться под тазом, а не сбоку, более эффективно принимая вес животного. Посмотрите, как ходит курица, а еще лучше – страус, и вы увидите, что его пальцы собираются при каждом шаге, чтобы пронести стопу мимо стоящей на земле ноги, а потом расправляются и ставятся под туловище. Если смотреть на следы птиц и тероподов (и даже на человеческие), становится ясно, что ноги не просто передвигаются спереди назад, но также из стороны в сторону, и такой характер движения становится более выраженным на больших скоростях.

Имеющаяся у нас информация дает наглядное представление о том, как тираннозавры могли использовать руки и ноги. Вписав эти знания в контекст других их анатомических особенностей, а также той окружающей среды, где обитали тираннозавры, и животных, с которыми они сосуществовали, мы можем воссоздать картину их жизни. Анатомия животного сообщает огромное количество информации о том, как оно функционировало, о его типичных действиях, возможностях и ограничениях и, следовательно, о том, как оно, вероятно, жило (а также умерло), даже если этот вид вымер десятки миллионов лет назад. Об этом мы будем говорить ниже, реконструируя жизнь тираннозавра, но сначала завершим описание этого животного рассказом о его покровах.
Назад: Глава 7 Туловище
Дальше: Глава 9 Внешний вид

