Книга: Хроники тираннозавра: Биология и эволюция самого известного хищника в мире
Назад: Глава 6 Череп
Дальше: Глава 8 Конечности
Глава 7
Туловище
Туловище тираннозавра представляет собой самую большую часть животного и по длине, и по массе. Позвоночный столб в виде длинного ряда позвонков проходит от задней части головы до самого кончика хвоста и служит главным опорным элементом тела и местом прикрепления конечностей, а также поддерживает голову. Внутри туловища располагается большинство основных органов, коротко рассмотренных ниже. Поскольку череп (описанный в предыдущей главе) связан с шейным отделом позвоночника, мы начнем описание туловища с первого из шейных позвонков.
Первый из ряда шейных позвонков – атлант, и он, как подсказывает название, поддерживает череп. Это сильно видоизмененный и необычный позвонок, по виду совсем непохожий на другие. За ним следует эпистрофей, еще один причудливый позвонок, который по большей части обеспечивает свободное движение головы по отношению к остальной шее. У большинства тираннозавров были довольно длинные шеи, но по мере увеличения размеров тела и особенно объема и массы черепа шея у тираннозавринов укорачивалась, уменьшая таким образом расстояние между головой и ногами и предотвращая чрезмерное утяжеление передней части и опрокидывание животного вперед. У всех тираннозавров имелось по 10 шейных позвонков, включая атлант и эпистрофей. Шея в положении покоя должна была изгибаться в виде буквы S, поднимаясь от туловища и скругляясь так, что голова оказывалась направлена немного вниз.
Помимо атланта и эпистрофея остальные шейные позвонки вполне типичны для позвоночных животных. Нижняя часть каждого представляет собой невысокий широкий цилиндр, называемый телом позвонка, он обеспечивает связь и сочленение позвонков в цепочку. В верхней части шейного позвонка располагается группа костных отростков, вместе называемых дугой позвонка или невральной дугой. Она является местом крепления мышц, удерживающих кости и позволяющих им двигаться, а также образует несколько дополнительных сочленений, вытянутых вперед и соединяющихся с дугой позвонка спереди, придавая конструкции прочность и устойчивость. Между телом позвонка и дугами есть отверстие, через которое проходит спинной мозг, соединяя головной мозг с различными частями тела. В промежутке между телами позвонков находится мягкая хрящевая прокладка, обеспечивающая амортизацию при движении костей, что предотвращает их истирание друг о друга. Каждая такая прокладка имеет дискообразную форму – это те самые диски, которые выскальзывают и вызывают у людей боли, если мешают костям двигаться правильным образом или, еще хуже, начинают давить на спинной мозг. Таким образом, позвоночник служит опорой для тела и сочленяющей системой, делающей возможным движение и обеспечивающей гибкость и прочный защитный футляр для спинного мозга.
Также в шейном отделе имеются несколько коротких шейных ребер, соединяющихся с позвонками и обеспечивающих поддержку дополнительных групп мышц. И мышцы, и разнообразные сухожилия скрепляют шейный отдел и обеспечивают ему поддержку, а также позволяют двигаться. На самом деле шея тираннозавра не слишком отличалась от шеи хищных птиц – кости соединялись сходным образом, и осуществляемые ими движения были похожи, особенно движение головой вверх и назад в процессе еды.
После шейных позвонков идут 13 грудных. В целом их можно отличить по небольшому изменению формы дуги позвонка и смещению места присоединения ребер. Эти ребра «настоящие», и они значительно больше шейных; они плавно изогнуты по всей длине и задают границу грудной полости. Грудные позвонки соединены друг с другом более крепко, чем шейные, обеспечивая меньшую гибкость и таким образом создавая прочное основание для туловища в целом.
Брюшные кости
Вдоль нижней части грудной клетки идет странный ряд костей, называемых гастралиями. Они в основном размещаются в теле свободно (т. е. не соединены непосредственно с другими костями) и обеспечивают некоторую поддержку передней части грудной клетки. У большинства динозавров они располагаются рядами перпендикулярно позвоночнику и представляют собой очень тонкие длинные кости – даже у гигантских завроподов они не толще нескольких палочек сухой вермишели, и потому редко сохраняются и воссоздаются в реконструкциях. Однако у поздних тираннозавров по сравнению с другими динозаврами гастралии становятся довольно странными по форме, размеру и креплению. Каждая почти равна ребру по толщине, а вся конструкция имеет приблизительно треугольную форму, и утолщенный конец располагается в середине туловища. Гастралии у них не лежат рядами параллельных полос, а перекрывают друг друга с разных сторон, образуя длинный ряд связанных V-образных, если посмотреть снизу, структур; некоторые могли даже срастаться, становясь еще прочнее. Это совершенно необычная конструкция, но как, почему и зачем она развилась, не известно. Предположительно она помогала поддерживать большую и тяжелую грудную клетку, особенно когда животное ложилось, однако у других столь же крупных и массивных тероподов не развилось ничего подобного, и еще только предстоит исследовать, почему тираннозавры настолько отличаются от всех.
Вернемся к позвоночному столбу: после грудных позвонков идут 5 костей крестца, состоящие из группы сросшихся позвонков. У многих животных на удивление маленький крестец, даже у таких крупных, как слоны, но у динозавров в этом отделе обычно имеется 5 или более позвонков, причем довольно больших. У молодых животных все крестцовые позвонки отделены друг от друга и с виду очень похожи на грудные, только располагаются между тазобедренными суставами. Однако по мере роста животного все позвонки срастаются телами (и дуги позвонков тоже часто срастаются друг с другом), становясь единой прочной костной структурой. Боковые отростки костей, называемые крестцовыми ребрами, тоже срастаются друг с другом, а также с соответствующими сторонами подвздошной кости. Таким образом, у взрослых особей таз в целом представляет собой один массивный прочный блок и обычно является самым крупным одиночным элементом скелета любого позвоночного. Крестец осуществляет передачу усилия. Вес массивного двуногого животного, такого как взрослый тираннозаврид, полностью приходится на ноги (а при ходьбе или беге – на одну ногу) и передается через бедренную кость. Если бы подвздошные кости не были так прочно скреплены с крестцом, то под весом динозавра наверняка проткнули бы спину животного, и туловище осело бы наземь.
Продолжаем двигаться дальше по позвоночнику. После крестца идут хвостовые позвонки. Динозавров часто реконструируют с очень длинным хвостом, как у ящериц, но на самом деле хвосты их чрезвычайно разнообразны и по количеству позвонков, и по общей длине. Некоторые динозавры явно обладали огромными хвостами: гигантский завропод апатозавр имел по меньшей мере 82 хвостовых позвонка, причем некоторые очень большие, а у овирапторозавра каудиптерикса (Caudipteryx) их было 22, и многие были очень короткими. Из-за сочетания факторов (связанных с тем, сколько в хвосте позвонков, каков их размер и как прочно они соединяются) даже частично сохранившиеся хвосты у динозавров очень редки, а полностью комплектные практически не известны. К счастью, одной из таких редкостей является экземпляр горгозавра, у которого сохранились все хвостовые позвонки до последнего – всего 37. Тем не менее бессмысленно, опираясь на это число, строить предположения о других особях горгозавра, не говоря уже о других тираннозаврах. Несмотря на малое количество данных по полным хвостам динозавров, нам точно известно, что и число позвонков, и длина хвоста значительно варьировали даже внутри одного вида. У тираннозавра нескольких первых позвонков довольно короткие, потом идут относительно длинные, далее они снова уменьшаются, становясь все короче и короче по направлению к кончику хвоста. Примерно на половине длины хвоста позвонки становятся очень простыми, и также чрезвычайно уменьшается сложность позвоночных дуг, пока наконец не остается почти ничего, кроме тела позвонка, а самый последний хвостовой позвонок представляет собой не более чем полукруглую шишку. Поскольку количество костей, мышц и кровеносных сосудов становится все меньше по направлению к концу хвоста, меньше структурного материала требуется, чтобы скреплять и придавать жесткость конструкции, так что все сочленения постепенно сходят на нет, поскольку мало что дают, кроме ненужного веса.
Под хвостовыми позвонками находится ряд более мелких костей, называемых шевронами (или гемальными дугами). У каждого шеврона имеется пара сочленений с позвонками вверху, хотя из-за своей формы они располагаются скорее между сочленениями позвонков, чем прямо под телом одного позвонка; далее на каждом из них есть более длинный отросток кости, направленный назад, так что в боковой проекции они выглядят весьма похоже на бумеранги или латинскую букву L. Между телом позвонка и шевронами проходят некоторые из главных артерий, снабжающих кровью хвост и мышцы, связанные с задними конечностями; по сути, шевроны ведут себя как аналог дуг позвонков на нижней стороне позвоночника, только защищают кровеносные сосуды, а не нервный ствол. Форма этих шевронов меняется на протяжении всего хвоста: ближе к концу они становятся короче и проще по форме и в итоге совсем исчезают. У первых двух-трех хвостовых позвонков также нет шевронов, потому что там должна была находиться клоака (мочеполовое отверстие).
Сочленения между хвостовыми позвонками менее жесткие, чем между грудными позвонками, и хвост должен был обладать некоторой гибкостью. Им бы не получилось свободно вилять во всех направлениях, как кошачьим, но, вероятно, возможно было двигать из стороны в сторону с приличным диапазоном и при необходимости изгибать. В целом, однако, хвост являлся довольно жестким, и его приходилось держать горизонтально, судя по сочленениям и мышцам. Старомодные изображения динозавров, волочащих хвосты по земле, теперь решительно опровергаются по двум причинам: во-первых, как рассматривается позже, мы уверены, что они обладали более вертикальной осанкой, чем представлялось вначале, а во-вторых, потому что опорные сочленения хвоста и мышцы должны были удерживать хвост достаточно жестко. Кроме того, в отпечатках следов динозавров очень редко обнаруживаются следы от волочения хвоста (иногда только от кончика), так что представление о том, что по земле тянулась половина хвоста, определенно неверно.
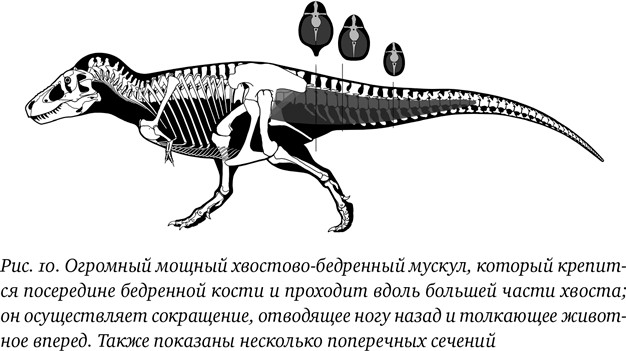
Это вполне логично, потому что хвост способствует двум важнейшим двигательным функциям, а в положении волочения он бы, скорее, им препятствовал. Во-первых, хвост двуногих животных является противовесом. Если тело в основном располагается параллельно земле, а стопы находятся под тазобедренными суставами, то его станет перевешивать вперед, если что-либо не будет его уравновешивать сзади: хвост и является таким противовесом. Он может показаться недостаточно большим для такой функции, но, как мы увидим, тело не настолько массивно, как кажется, и, поскольку хвост обычно несколько длиннее, чем голова и туловище вместе взятые, получается некоторый механический выигрыш, потому что хвост оказывается дальше от ног, чем голова. Во-вторых, хвост служит местом прикрепления огромных хвостово-бедренных мышц, доходящих до основания бедренной кости. Это главная сократительная группа мышц, которая отводит ногу назад и таким образом толкает животное вперед. Словом, это был важнейший мускул, позволявший динозавру ходить или бегать, и он не мог бы эффективно функционировать, если бы передняя часть хвоста располагалась почти параллельно бедренной кости и хвост волочился по земле, а грудной отдел позвоночника был бы вытянут почти вертикально. Не так давно было доказано, что этот мускул больше, чем считалось ранее (рис. 10), и, вероятно, образовывал некое утолщение у основания хвоста, где крепился.
Есть и другие группы мышц, проходящие через шею, тело и хвост и осуществляющие опору и движение животного, и эти мышцы дополнены сетью кровеносных сосудов, нервов и прочими тканями. Я, однако же, хочу перейти к важнейшим внутренним органам тираннозавров. Значительная часть информации по этой теме основывается почти исключительно на логических умозаключениях (в общем и целом у всех позвоночных имеется какая-то форма почек, и трудно представить, как тираннозавры могли бы без них обойтись, так что можно с уверенностью считать, что почки у них были) либо на данных экстраполяции (отверстия в костях показывают, где проходили кровеносные сосуды, и позволяют предположить, с чем они могли быть соединены), но в некоторых случаях нам очень помогают отдельные уникальные ископаемые находки.
Мягкие ткани
Несколько лет назад возникла большая шумиха по поводу находки окаменелого сердца динозавра. Такое возможно: внутренние органы могут сохраниться в том случае, если они не успели сильно разложиться и попали в достаточное количество грязи или других осадочных материалов или если образовался естественный слепок, и тогда, даже если сами органы сгниют и не минерализуются, их исходная форма сохранится в окаменелом виде (примерно так же, как отпечаток лапы). К сожалению, в итоге это «сердце» оказалось всего лишь необычным каменным желваком, но его размер, форма, положение в скелете и общий вид в самом деле позволяли посчитать его серьезной заявкой на сенсацию. И у современных крокодилов, и у птиц четырехкамерные сердца, которые несколько отличаются от сердец остальных рептилий, и это позволяет предположить, что у динозавров было нечто похожее. Однако между крокодильими и птичьими сердцами есть некоторые различия, поэтому трудно понять, были динозавры в целом больше похожи на крокодилов или на птиц.
Значительно лучше изучены легкие и система воздушных мешков. Как уже упоминалось раньше, тероподы и завроподы были по большей части довольно пневматическими существами. В придачу к парным легким и синусам, которые есть у почти всех наземных тетраподов (некоторые амфибии практически утратили легкие, а у змей легкое только одно), у этих животных имеются расширения легких, заходящие не только в полость тела, но и во многие кости. Отверстия проницают наружную поверхность кости, потом полости расширяются и занимают большую часть внутреннего объема кости. У таких пневматических динозавров (и также у современных птиц и вымерших птерозавров) в некоторых костях имеются огромные полости; даже гигантские шейные позвонки некоторых крупных завроподов могут более чем на 90 % объема заполняться воздухом и только на несколько процентов – костной тканью. Однако то, что эти кости были полыми, не делало их непрочными, потому что их укрепляло огромное количество крошечных костных балок, называемых трабекулами, – немного похоже на то, как спицы придают жесткость велосипедному колесу, – и кость в разрезе могла выглядеть как пчелиные соты. Многие длинные кости (такие как бедренная) у животных полые: если разрезать коровью бедренную кость, то вы увидите, что середина пустая, однако полость, во-первых, довольно маленькая по сравнению с пневматической полостью у динозавра, во-вторых, заполнена костным мозгом, и, разумеется, подобные полости не встречаются в таких костях, как грудные позвонки.
Воздушные мешки заходили во многие кости динозавров, и в некоторых случаях практически по всему скелету обнаруживаются какие-нибудь пневматические расширения. В случае тираннозавров они сосредотачивались в позвоночном столбе и соединенных с ним костях. У тираннозавра, например, имелись пневматические отверстия в шейных позвонках и шейных ребрах, грудных позвонках и ребрах, крестцовых позвонках и первых нескольких хвостовых (хотя эти отверстия были довольно малы). Со временем у разных групп такие мешки перемещались все ниже по телу, и у некоторых нептичьих тероподов даже имелись пневматические участки плеч и передних лап, таза и ног, находившиеся на довольно приличном расстоянии от исходного местоположения в грудной клетке. Существует много групп воздушных мешков, и каждая пронизывает только определенные кости, так что мы можем проследить их развитие и продвижение по скелету.
Важно отметить, что у динозавров прохождение воздуха через легкие и воздушные мешки весьма отличалось от аналогичного процесса у млекопитающих. Мы вдыхаем, расширяя легкие, и воздух заполняет их. В наших легких есть многочисленные ветвящиеся трубки и трубочки, которые заканчиваются огромным количеством маленьких сферических образований. Здесь происходит газообмен, кислород поступает в кровоток, а углекислый газ уходит из него, потом мы выдыхаем и выталкиваем воздух обратно тем же путем, которым он вошел. Это двунаправленный воздушный поток: один и тот же «глоток» воздуха проходит внутрь и выходит наружу по одним и тем же путям легких.
У птиц эти разнокалиберные трубочки в легких соединяются друг с другом, и потому с виду напоминают не столько огромную кучу крошечных воздушных шариков, сколько лабиринт крошечных трубочек. Легкие также соединены с воздушными мешками, и все это вместе образует альтернативный дыхательный путь. При вдохе воздух затягивается в задние воздушные мешки, а оттуда переходит в легкие, потом в передние воздушные мешки и наконец выходит наружу. Следовательно, воздушный поток в легких оказывается односторонне направленным: воздух проходит по большей части дыхательной системы только в одну сторону, а воздушные мешки существуют исключительно для управления потоком, газообмен же происходит только в легких. В результате многие воздушные мешки представляют собой застойные тупики; тогда как мешки в полости тела используются в процессе дыхания, у ответвлений в костях нет таких функций, и обычно в них не наблюдается особенного потока воздуха, если он вообще присутствует.
Чрезвычайно интересное недавнее исследование показало, что аллигаторы и некоторые крупные ящерицы умеют дышать как птицы, несмотря на отсутствие воздушных мешков. Многокамерная структура каким-то образом позволяет легким функционировать и как легкие, и как группа воздушных мешков у птиц, и поэтому воздушный поток получается однонаправленным. Это является превосходным аргументом в пользу однонаправленного воздушного потока при дыхании у динозавров и предполагает, что такой тип дыхания развился еще до возникновения архозавров и, следовательно, был по меньшей мере в начале эволюции универсальным для динозавров. Наличие множества воздушных мешков у тероподов свидетельствует о том, что данный признак сохранился и с тех пор даже расширился: легкое птиц, таким образом, очень похоже на динозавровое.
О пищеварительной системе динозавров известно мало, но для тероподов есть хоть какие-то данные. Местоположение желудка можно приблизительно определить по положению проглоченных костей в верхней части грудной полости различных тероподов. Мы имеем некоторое представление о форме кишечного тракта благодаря найденному в Италии невероятному ископаемому экземпляру крошечного теропода под названием сципионикс (Scipionyx). Это компсогнатид (а значит, близкий родственник тираннозавров), и у него сохранились различные органы или по крайней мере их отпечатки, и в том числе бо́льшая часть пищеварительной системы. В целом она оказалась довольно короткой, как и ожидалось: и сципионикс, и тираннозавры были хищниками, поэтому должны были иметь относительно короткий и простой пищеварительный тракт, поскольку мясо в основном переваривается достаточно легко по сравнению с растительной пищей. Тот факт, что даже у тираннозавра копролиты (окаменелые фекалии) содержат различимые фрагменты костей, говорит нам, что тираннозавры не обладали сверхкислой пищеварительной системой, как у некоторых современных животных, т. е. были, вероятно, обыкновенными, а не исключительными животными в плане расщепления пищи.
Последняя важная система органов связана с выделением и размножением. Как отмечалось выше, тираннозавры (как все рептилии и птицы и даже некоторые млекопитающие, такие как утконос) имеют одинарную клоаку. Это общий выход мочеполового тракта и пищеварительной системы – проще говоря, все, что выводится из организма, выходит отсюда. Естественно, располагаться клоака должна была у основания хвоста и над самым задним концом седалищной кости. Мы знаем, что динозавры выделяли фекалии, потому что находим их копролиты, но, возможно, они производили экскременты, подобные птичьим и крокодильим, в которых фекалии и моча смешаны (отсюда белые потеки птичьего помета: в нем много мочевины), или же выделяли мочу отдельно, как делают разные птицы (по крайней мере иногда).
Клоака самок также должна была служить выходом для яиц. Яйца, скорее всего, формировались в двух яичниках и развивались в двух яйцеводах, в отличие от птиц, которые ограничиваются одним набором этих органов. Мы об этом знаем частично, потому что многие манирапторовые тероподы, по-видимому, откладывали яйца в гнезда по два, а также известен экземпляр троодонтида с парой яиц внутри тела – это показывает, что развивались оба яйца сразу. У тираннозавров многочисленные яйца, вероятно, формировались одновременно (что, похоже, было общим для многих тероподовых динозавров), причем это должно было происходить сразу в обоих яичниках.
Самец тираннозавра должен был доставить сперму в клоаку самки. Это может показаться невозможным при общем строении тираннозавров, но множество рептилий и птиц спариваются практически без каких-либо действий, кроме прижимания клоак друг к другу. Тем не менее размеры многих динозавров (и, конечно, форма их тела, когда речь заходит о бронированных стегозаврах) должны были делать спаривание неудобным, и могло требоваться нечто, помогающее преодолеть расстояние между клоаками.
Вообще говоря, некоторые животные с клоаками развили какую-либо форму «копулятивного органа». Этот термин не является некой данью излишней стыдливости, но в самом деле необходим, чтобы отличать такой орган от совсем иного пениса млекопитающих. У птиц и рептилий, обладающих подобными органами, они аналогичны (имеют ту же функцию), но не гомологичны (не имеют общего эволюционного происхождения) пенису и, следовательно, не должны называться тем же словом. Копулятивные органы могут быть короткими и простыми – или довольно длинными и сложными приспособлениями – в зависимости от того, как именно они функционируют, а также от имеющихся сложностей спаривания. У крупных тираннозавров вполне могло использоваться нечто, помогающее самцу дотянуться до самки, но мы вынуждены ограничиваться попытками так совместить два скелета друг с другом, чтобы копуляция могла считаться успешной (в одной музейной экспозиции два скелета динозавров смонтировали в позе спаривания). Чтобы получить некоторое представление, насколько причудливым может быть копулятивный орган, рекомендую поинтересоваться сексуальной биологией уток. Мгновенно раздувающийся орган, который, во-первых, длиннее своего обладателя, а во-вторых, имеет винтообразную форму – и это только начало. Утки – занятные, скажем так, животные в том, что касается секса.
Ключевыми элементами в совмещении этих животных друг с другом являются таз и задние ноги, а также важно, насколько многотонные тираннозаврины были способны объединять усилия, чтобы свести вместе свои репродуктивные органы. Одной из проблем должно было быть равновесие, но тут, вероятно, помогали гибкость и активное использование конечностей (и передних, и задних). Кстати, это удобная возможность перейти к следующей главе – к последней важной части скелета: конечностям.
Назад: Глава 6 Череп
Дальше: Глава 8 Конечности

