Книга: Хроники тираннозавра: Биология и эволюция самого известного хищника в мире
Назад: Глава 4 Родственные связи тираннозавров
Дальше: Часть II Морфология
Глава 5
Тираннозавры во времени и пространстве
Хорошо разобравшись, кто является тираннозавром, а кто нет, а также составив представление о разных группах тираннозавров и их родственных связях, можно углубляться в то, что происходило с этими животными на протяжении миллионов лет, пока они бродили по земле. Позже мы отследим эволюционные тенденции в изменениях формы и размера, продолжавшихся со времени первого появления тираннозавров и вплоть до последних представителей этой группы, но здесь рассмотрим картину их распространения.
Мы не знаем, каким был первый тираннозавр. Филогении позволяют нам определить виды, которые, по нашим предположениям, были ближайшими к точке отделения тираннозавров от других тероподов, но мы не можем сказать, являлось ли это животное исходным «анцестральным» или предковым видом. Следовательно, даже если нам посчастливится когда-либо найти поистине «первого» тираннозавра, мы даже не узнаем, что нашли его: ведь эволюция не просто отделяет вид как готовую новую группу, но постепенно разводит эволюционирующие популяции, и потому поиски подобного животного, скорее всего, тщетны.
Однако мы все-таки можем попытаться определить, как должно было выглядеть это животное, а также когда и где на Земле оно могло появиться, на основании того, что мы знаем о самых древних тираннозавроидах и базальной для них кладе целурозавров, от которых они отделились. Может показаться, что сложнее всего установить фактическое время появления этого животного, но в некотором смысле это один из самых простых вопросов.
Вопреки популярному представлению, возраст горных пород и окаменелостей нельзя определить методом радиоуглеродной датировки, т. е. по изотопам углерода. Дело в том, что радиоактивные углеродные часы точны в пределах нескольких десятков тысяч лет, а потому бесполезны для любых материалов старше 50 000 лет и, следовательно, применимы к очень малому числу окаменелостей, но зато очень полезны для древних останков, еще не успевших минерализоваться и превратиться в камень. Тем не менее сам принцип, на котором построен метод, не слишком далек от того, который используется при датировке значительно более древних останков.
Многие химические элементы существуют в нескольких формах, называемых изотопами: у изотопов одинаковое количество протонов и электронов, что придает им характерные свойства, но число нейтронов отличается, поэтому изотопы имеют разную массу. Эти формы пишутся с числами, которые показывают их атомный вес, так что часто можно увидеть упоминания об 238U как о форме урана или об 14С как форме углерода. Изотопы с дополнительными нейтронами нестабильны и радиоактивны, они избавляются от лишнего багажа, чтобы перейти в более стабильную форму. Поскольку этот распад происходит с одинаковой скоростью, характеризующейся периодом полураспада, то, если мы знаем соотношение радиоактивного изотопа и нормальной формы в начале временного периода и плюс сейчас нам известна скорость полураспада, мы можем рассчитать, как давно начался распад. Словом, мы можем приблизительно определить возраст данного материала. Что удобно, некоторые изотопы остаются стабильными чрезвычайно долгое время, поэтому мы можем датировать даже очень древние горные породы по таким изотопам с большим периодом полураспада. В настоящее время периоды полураспада для изотопов измерены очень точно и откалиброваны друг относительно друга, а лучшие датировки проводятся по нескольким изотопам, которые распадаются с разной скоростью, так что их можно сравнивать. Уже нет ничего необычного в том, что породы возрастом 100 млн лет датируются с точностью до пары десятков тысяч лет. Это очень впечатляющие результаты.
Но, как ни досадно, мы можем датировать таким методом только вулканические породы, а в них залегает мало ископаемых останков живых организмов (что неудивительно). Поэтому обычно мы датируем вулканические породы, залегающие выше и ниже горизонта, содержащего изучаемую окаменелость, и это дает диапазон, в пределах которого должно было жить данное животное. Иногда – когда вулканические слои лежат очень близко к нашей окаменелости – этот диапазон довольно узок, но в других случаях они могут располагаться сильно выше или ниже, отчего диапазон получается большим. Иногда породы датированы не очень точно или для них вовсе нет датировок (сопоставляя их с другими породами, мы все же можем получить общее представление, когда они сформировались, но не точные значения), и это неизбежно ведет к еще большему разбросу. Геологи также проделали огромную работу, соотнося слои горных пород в разных местах залегания, а палеонтологи помогали им, используя определенные ископаемые, которые могут относиться только к ограниченным временным периодам, что позволяет сопоставить отложения ископаемых останков по всему миру. В результате, если вдруг обнаруживается новое местонахождение окаменелостей, можно довольно просто определить, из каких породных формаций они происходят или как минимум к каким они ближе всего, и высчитать возраст.
Огромное множество породных формаций, содержащих останки динозавров, теперь датированы довольно подробно, поэтому в основном у нас имеется вполне удовлетворительное представление о возрасте изучаемого тираннозавра, и в некоторых случаях мы можем очертить временные границы существования вида.
Древнейшие тираннозавры
Самые древние тираннозавры – килеск и процератозавр, обоим примерно по 167 млн лет, но и тот и другой известны по довольно небольшому количеству ископаемого материала (относящегося к среднеюрскому периоду). Гуаньлун из Китая, однако, известен по нескольким почти полным экземплярам и позволяет составить значительно лучшее представление об этих древних тираннозаврах. Он ненамного младше двух первых: породы, в которых он был обнаружен, датированы 160 млн лет. Даже при отсутствии находок более ранних тираннозавров мы можем сделать вывод, что данная клада, вероятно, возникла раньше этого времени. Другие тероподы, такие как альваресзавры, которые, как мы знаем, отделились от основной ветви тероподов после тираннозавров, известны из тех же слоев, что и гуаньлун, и даже более поздние тероподы, очень близкие к птицам, залегают в породах того же возраста, как те, где обнаружили останки процератозавра. Если эти разные линии тероподов разошлись на тот момент и к тому же достигли такого разнообразия, то, скорее всего, они уже существовали некоторое время, и, следовательно, тираннозавры должны были отделиться еще раньше.
По моим предположениям, в итоге выяснится, что тираннозавры появились в раннеюрском периоде, возможно, около 190 млн лет назад. Незадолго до этого, в конце триаса, произошло большое вымирание, которое значительно изменило наземную жизнь и в результате, вероятно, помогло динозаврам – одной из многих групп рептилий – стать доминантами суши. Мы часто видим значительное увеличение видового разнообразия после таких вымираний, и оно вполне могло частично поспособствовать разделению тероподов на группы в это время, приведя в итоге к появлению тираннозавров.
В палеонтологии существуют некоторые фундаментальные закономерности, которые оказывают ограничивающее и искажающее влияние на то, какие ископаемые останки мы можем найти. Процессы образования окаменелостей таковы, что некоторые останки сохраняются лучше (и, следовательно, обнаруживаются чаще), а другие – значительно хуже, поэтому все находки мы видим через такой фильтр; это надо иметь в виду при интерпретации имеющихся у нас зачастую ограниченных данных. Самый очевидный пример такой погрешности – то, что твердые ткани (кости, зубы, раковины и т. п.) минерализуются со значительно большей вероятностью, чем мягкие (такие как мышцы, кожа и внутренние органы), поскольку первые практически не подвержены гниению, а вторые разлагаются довольно быстро. Поэтому, например, у нас имеется практически неограниченное количество окаменелых раковин аммонитов, но только в редких случаях сохранилось что-то от животного, обитавшего в этой раковине. Аналогично у молодых животных кости обычно менее минерализованы, чем у взрослых, поэтому сохраняются не так часто (здесь, правда, участвуют еще более важные факторы, как будет рассматриваться позже). Маленькие окаменелости труднее найти, чем большие, так что возможны погрешности не в пользу мелких находок, но очень большие окаменелости вряд ли были полностью засыпаны грунтом, поэтому, скорее всего, будут неполными.
Как и где обитало животное, тоже имеет огромное значение: чтобы труп фоссилизировался, т. е. стал окаменелостью, он должен быть засыпан грунтом прежде, чем полностью разложится или будет съеден падальщиками. Это означает, что дождевые леса, несмотря на обилие жизни, – совершенно неподходящие места для образования ископаемых останков, потому что скорость разложения в них просто астрономическая. Пустыни же, несмотря на низкую населенность небольшим количеством видов крупных животных, могут быть чрезвычайно продуктивны в плане окаменелостей, создавая ложную картину разнообразия и численности организмов. Аналогично любые животные, обитающие в прибрежных районах или местах вокруг речных пойм и озер, имеют намного больше шансов быть погребенными и сохраненными, чем живущие в высокогорных районах. Меня очень интересует, не пропустили ли мы динозавровый эквивалент горных козлов, поскольку в горах вряд ли образовывалось много окаменелостей. Здесь также вступают в игру основы биологии и соотношения численности групп: на каждого хищника могут приходиться сотни травоядных, а мелкие животные обычно более многочисленны, чем крупные. Кроме того, мы ограничены теми горными породами, которые нам доступны: к примеру, известны находки динозавров в Антарктиде и Северной Корее, но оба места довольно трудно исследовать, хотя и по разным причинам. Также для поиска окаменелостей нам нужны породы, выходящие на поверхность земли, так что любые «динозавросодержащие» породы на морском дне, или под землей, или даже под городами невозможно исследовать (к примеру, содержащие останки динозавров породы Парижского бассейна местами труднодоступны по очевидным причинам: над ними располагается город).
Добавим ко всему этому следующий факт: вообще-то немногие особи животных оказываются погребенными под грунтом, а также обнаруживаются палеонтологами (при полевых работах обычно находят значительно больше фрагментов, подвергшихся эрозии, и разрозненных окаменелостей, чем хорошо сохранившихся костей, не говоря о целых скелетах), и есть элемент чуда в том, что в музеях имеются хоть какие-то кости динозавров, не говоря о целых залах скелетов. Фактор, работающий на нас, – это, несомненно, время. Только одно из примерно миллиона животных становится окаменелостью, но за длительный период времени популяция даже крупных животных легко может насчитывать сотни тысяч или миллионы особей, и каждый год они производят миллионы потомков, а за пару-тройку миллионов лет, на протяжении которых, вероятно, существовал данный вид, общее число может вырасти до миллиардов и десятков миллиардов, что дает некоторый простор для изучения (если мы сможем их найти и выкопать).
Последний важный фактор, имеющий отношение к качеству ископаемых данных, – возраст горных пород. После того как организм становится окаменелостью, с ним может многое случиться. Даже если породы, содержащие окаменелость, не погребены под новыми слоями и не утянуты на дно океана континентальным дрейфом, они могут подвергнуться эрозии. Чем дольше они существуют, тем более вероятно, что это случится, и потому чем ближе мы подходим к современности, тем лучше становятся ископаемые данные. А значит, неудивительно, что у нас имеется лучшая летопись позднемелового периода, чем, скажем, среднеюрского, и что мезозой в целом беднее в плане находок, чем более близкие к нам периоды, но намного богаче значительно более древнего кембрийского (хотя, конечно же, отдельные места залегания окаменелостей могут быть весьма продуктивными и репрезентативными).
Все эти факторы влияют на тираннозавров так же, как на любую другую кладу, и в плане обнаружения древнейших форм мы подвержены почти всем этим тенденциям: первые тираннозавры должны были быть небольшими, немногочисленными, хищными и жить в средне– или даже раннеюрском периоде. Из имеющихся у нас видов гуаньлун – ближайший к истокам группы (по крайней мере есть целый экземпляр животного для работы), но, учитывая множество недавних находок останков динозавров из среднеюрского периода, есть неплохие шансы, что нам удастся обнаружить новые, лучше сохранившиеся экземпляры таких животных, как процератозавр, и они больше поведают нам об этом конкретном периоде возникновения и ранних этапов эволюции тираннозавров.
Датировка других наших тираннозавров показывает, что они существовали на протяжении широкого спектра временных периодов. Встречается некоторая скученность видов, потому что ряд окаменелостей были найдены в продуктивных районах залегания (например, алиорам и тарбозавр в позднемеловых отложениях Монголии, а также дилун с ютираннусом в раннемеловых породах Китая), но в целом наблюдается распределение форм по времени в течение 100 млн лет существования тираннозавров на Земле. Процератозавр и килеск являются примером самых древних известных тираннозавроидов и также самыми ранними процератозавридами, а первые тираннозавриды появляются лишь через весьма значительный промежуток времени. И альбертозавр, и горгозавр дают для альбертозавринов датировку примерно 70 млн лет, но эта ветвь должна быть еще старше, потому что они отделились от тираннозавринов, а самым ранним представителем тираннозавринов является недавно получивший название литронакс, которому около 80 млн лет. Фактически это означает, что данные дочерние группы тираннозавридов образовались и разделились относительно близко к концу всех линий тираннозавров, т. е. просуществовали «всего» 15 млн лет.
Как минимум тираннозавры как группа прожили долго и пережили многие другие линии динозавров: со времени первого появления в палеонтологической летописи они просуществовали до самого конца мела. Тираннозавр на самом деле один из самых последних известных динозавров, и останки представителей этого вида находят в породах, образовывавшихся практически вплоть до великого вымирания 65 млн лет назад. Все эффектные образы тираннозавра на фоне горящего мира или обугленного скелета на выжженной равнине вполне имеют право на существование: ветвь тираннозавров закончилась только вместе с массовой гибелью множества их родственников.
Вокруг света за 80 млн лет
Тираннозавры не только просуществовали долгое время, но и широко распространились по миру. Хотя их останки наиболее многочисленны и лучше известны в Северной Америке, Восточной Азии и в меньшей степени в Европе, предполагаемые останки тираннозавров также нашли в Бразилии и даже в Австралии. Притом что палеонтологическое изучение южных континентов еще переживает период относительного «детства», там вполне могут быть найдены новые ископаемые – правда, принадлежность этих южных видов к тираннозаврам оспаривается, и они не однозначно тираннозавровые по признакам. Для австралийского экземпляра, к примеру, найдено немногим больше, чем часть таза, и, хотя у него имеется увеличенный лобковый «башмак» и другие классические тираннозавровые черты, при таком небольшом количестве данных его принадлежность остается несколько неопределенной.
Больший интерес представляет вопрос, где и когда появились в мире те или иные виды. Обобщая наши знания о возрасте различных окаменелостей, их географическом происхождении и эволюционных взаимосвязях между ними, мы можем проследить пути их распространения по миру. Добавив данные о времени разделения континентов, возможно составить представление о том, как могла идти эволюция некоторых групп в привязке к этим грандиозным событиям.
В этом вопросе важно постараться установить, связано ли данное распространение видов по континентам с дисперсией или викаризмом. Как виды попадали туда, где мы их нашли? При дисперсионном типе распространения представители группы переселяются на уже отделившийся континент или участок суши: например, первые животные прибыли на новообразовавшиеся вулканические Галапагосские острова с Южно-Американского континента. В случае викарирования виды перемещаются вместе с сушей: когда континент или массив суши отделяется, он забирает с собой виды, которые на нем обитают, и их потомки остаются там, даже если позже вымрут в других местах. Викаризмом объясняется присутствие лемуров на Мадагаскаре: когда-то они тоже обитали на Африканском континенте, а когда там они вымерли, осталась только изолированная популяция на Мадагаскаре.
Живые организмы могут путешествовать между континентами, даже далеко разделенными, хотя, конечно же, это значительно проще для тех видов, которые способны благополучно пересекать обширные водные пространства, подобно птицам и кокосовым орехам. В мезозойскую эру континенты располагались в целом ближе друг к другу, чем сейчас, но даже при этом существовали места, где обширные массивы суши, такие как Индия, были полностью отделены от всего остального; в то же время на другие территории, такие как Антарктида и Южная Америка, можно было попасть по перемычкам относительно легко (рис. 8).
Если рассмотреть стандартную филогению тираннозавров вместе с известными местами нахождения отдельных экземпляров различных тираннозавров, картина получается довольно сложная. Ранние формы встречаются на многих континентах: процератозавридов и других древних тираннозавроидов находили в Европе, Северной Америке и Азии. Начнем с процератозавридов, поскольку упоминавшийся выше гуаньлун происходит из среднеюрского периода и найден в Западном Китае, процератозавр относится к тому же периоду (пусть и несколько старше) и обнаружен в Великобритании, а стоксозавр – в позднеюрских отложениях в США, и ему около 150 млн лет. Таким образом, даже это первое раннее расхождение групп тираннозавров продолжалось по меньшей мере 10 млн лет, и эти животные обитали на трех континентах.
Ископаемые останки различных особей и видов неопределенной филогенетической принадлежности осложняют картину, а такие находки, как уже упомянутые австралийская лобковая кость и бразильский экземпляр, дают основания считать, что тираннозавры могли распространяться по миру несколько раз, добираясь до южных континентов. В то же время основные данные все же предполагают, что группа в целом изначально обитала в Северном полушарии, и такие животные, как эотираннус, дриптозавр и сюнгуаньлун, появились соответственно в Европе, Северной Америке и Азии в раннемеловом периоде. Если продвинуться немного вверх по филогенетическому дереву, то альбертозаврины явно происходят из Северной Америки и, по-видимому, представляют собой местную эволюционную радиацию, которая не распространилась за пределы континента; аналогично алиорамины в настоящее время известны только из Восточной Азии. Тем не менее, хотя северные континенты в ранне– и позднемеловом периодах разошлись еще дальше, тираннозаврины встречаются и в Северной Америке, и в Азии.
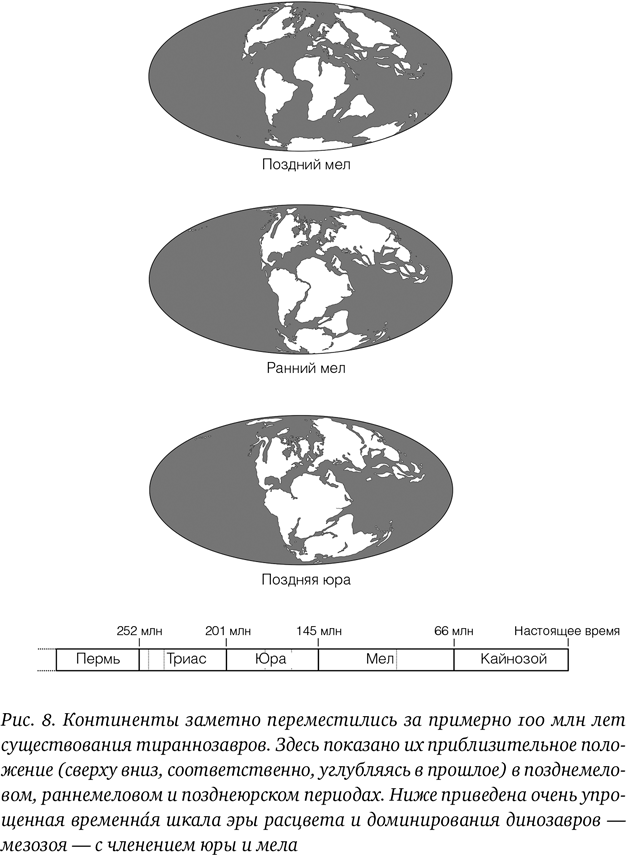
Глядя на аналогичную динозавровую фауну в Азии и Северной Америке, мы, что неудивительно, видим ряд параллелей: на обоих континентах есть цератопсы, гадрозавры, овирапторозавры, разнообразные дромеозавры, троодонтиды и альваресзавры – и один-два завропода. Однако, по-видимому, в целом на каждом континенте обитают собственные линии животных. Обычно в Азии нет близких родственников североамериканских животных, и наоборот (гадрозавр зауролоф – явное исключение, поскольку данный род известен для обоих этих континентов), и это предполагает, что в целом не так уж много животных переходило с континента на континент. Тем не менее тираннозаврины, судя по всему, совершили этот переход в какой-то момент, а может быть, и не один раз.
Обращаясь к филогении тираннозавров из предыдущей главы, мы видим, что все древнейшие тираннозавриды (и альбертозаврины, и тираннозаврины) происходят из Северной Америки, что дает основания считать этот континент вероятным местом их появления, а позже возникает всплеск видообразования в Азии (оттуда, например, алиорам, однако установление места появления алиораминов, возможно, поколеблет гипотезу североамериканского происхождения базальной клады). Это означает, что в какой-то момент представители данной группы перебрались в Азию и породили многообразные виды, но прямо на «вершине» дерева находится тираннозавр – животное вполне американское, так что его предки, по всей видимости, должны были вернуться через Берингов пролив в Америку.
Некоторые из этих картин усложняются, если рассматривать иные возможные гипотезы филогении тираннозавров. Например, дерево Лёвена, упомянутое ранее, предлагает альтернативную картину переходов, с другими временны́ми рамками, но все равно в итоге получается, что тираннозаврины пересекали Берингов пролив неоднократно. Очевидно, эта группа состояла из своеобразных динозавров – кругосветных (или по крайней мере межконтинентальных) путешественников, и ее представители перемещались по миру больше, чем члены других групп.
Еще один интересный аспект – смена фауны в меловом периоде. В дальнейших главах мы рассмотрим изменения в фауне (включая и конкурентов, и возможную добычу), среди которой жили тираннозавры в разные моменты их истории, но в мезозое произошло ключевое событие, которое, как считается, повлекло за собой многие из фиксируемых нами перемен. В какой-то момент в юрском периоде среди растений появилось эволюционное новшество – цветковые. Как и многие группы организмов, цветковые растения поначалу были маленькими и редкими, но начиная с 125 млн лет назад (в раннем мелу) мы видим их массовое распространение наряду с изменениями во многих группах животных (включая, например, насекомых-опылителей). Классические образы гигантских завроподов, поедающих древовидные папоротники и саговники, уступили место гадрозаврам и цератопсам, ощипывающим молодые побеги хвойных и многие, более современные растения; в это же время распространяются птицы, появляются общественные насекомые, образуются новые виды млекопитающих, а другие группы в основном приспосабливаются к этим изменениям. Данный период известен как меловая революция, и считается, что массовая смена растительности на более современные цветковые растения, называемые ангиоспермами, привела к радикальному изменению жизни на суше (но не в море); в результате новые группы адаптировались и эволюционировали, обращая эти перемены к своей пользе, а другие постепенно сокращались или совсем вымерли.
Многое изменилось в меловом периоде и, разумеется, важные перемены произошли в динозавровой фауне по всему миру. Так, к позднему мелу в Северном полушарии осталось мало завроподов (хотя они продолжали процветать на юге), при этом анкилозавры, цератопсы и гадрозавры составляли огромную часть травоядной фауны, а разнообразные оперенные тероподы, называемые манирапторами, исчезли практически полностью. Тем не менее недавно было показано, что изменения, произошедшие в то время, были, возможно, не такими уж радикальными, как полагали раньше: хотя разнообразие многих новых групп в меловом периоде возросло и привело их к доминированию в позднем мелу, впервые они появились значительно раньше. Представители всех групп манирапторов уже присутствовали в среднеюрском периоде (кроме птиц, по данным на настоящий момент, однако археоптерикс известен из поздней юры), и другие важные группы, такие как цераптопсы, также существовали в это время. Возможно, меловая революция много сделала, чтобы ускорить изменения в этой фауне, но предположение, что она также вызвала новую вспышку видообразования динозавров (или что новые динозавры стимулировали подъем цветковых растений, как когда-то предполагалось), теперь кажется маловероятным. А вероятным кажется то, что развитие стало более устойчивым.
Несомненно, тираннозавры населяли значительные пространства в течение большей части своего существования, и хотя они так никогда по-настоящему и не укоренились на юге, в меловом периоде в целом и в позднемеловом в особенности их было множество – и видов, и особей. Несмотря на некоторые пробелы в наших знаниях, общая картина распространения вполне ясна, и хотя новые открытия могут дать больше информации, кажется маловероятным, чтобы мы вдруг обнаружили, скажем, некую неизвестную вспышку видообразования в Северной Америке или выяснили, что происходил некий масштабный обмен видами между Южной Америкой и Азией. Тем не менее, по-видимому, тираннозавры много путешествовали: другие группы также пришли в итоге к широкому распространению, однако в настоящее время мы не видим у них таких перемещений в разных направлениях, как у крупных тираннозавров.

Рассмотрев тираннозавров в эволюционном контексте, в следующей части мы углубимся в анатомию этих животных. Исследования в первую очередь основываются на самих костях, и глядя на них, мы можем что-то узнать и о мягких тканях, таких как мышцы, легкие, глаза и мозг, а также сделать выводы о росте и развитии этих животных, о том, как быстро они могли передвигаться, чем и как питались, и многое другое.
Назад: Глава 4 Родственные связи тираннозавров
Дальше: Часть II Морфология

