Книга: Хроники тираннозавра: Биология и эволюция самого известного хищника в мире
Назад: Глава 3 Виды тираннозавров
Дальше: Глава 5 Тираннозавры во времени и пространстве
Глава 4
Родственные связи тираннозавров
Несмотря на все разногласия и споры вокруг идентификации видов и присвоения им названий, у нас все же сложилось неплохое представление о различных видах (и родах) тираннозавров – по крайней мере на сегодняшний день, – и мы рассмотрели признаки, объединяющие разные группы. Знание о том, чем характеризуются виды и какой ископаемый экземпляр к какому виду относится, – очень важная отправная точка, но, чтобы по-настоящему понять эволюцию группы животных, нам нужно представлять, как ветвилось эволюционное древо тираннозавров, образуя разнообразные линии и виды.
Беглый обзор некоторых общих признаков различных родов дает представление о том, какие из них могут быть ближе друг к другу, а какие дальше. Таков в целом подход традиционных таксономистов: объединять живые организмы в группы на основании нескольких общих признаков, но подобный метод имеет ограниченное разрешение, и всегда есть риск уже упоминавшейся проблемы «толстокожих-пахидерм», т. е. ошибочный выбор характерных признаков. Разумеется, тираннозавр, тарбозавр и чжучэнтираннус имеют достаточно общих признаков, чтобы мы могли счесть их близкими родственниками (они крупнее любых других тираннозавров, у них меньше зубов в верхнечелюстных костях и более высокий череп), но полезно и интересно точно знать, кто к кому ближе, а это требует более детального анализа.
Методы, используемые для составления эволюционных деревьев организмов и показывающие, какие между ними родственные связи, называются филогенетикой, а составленные «деревья» – филогениями. Их создание, по сути, сводится к оценке всех доступных данных по какому-либо виду и сравнение с данными по другим видам, чтобы определить, у каких видов больше общих черт. Виды, являющиеся более близкими родственниками, будут иметь больше общих признаков, поскольку у них более длительная общая эволюционная история и они меньше изменились с тех пор, как приобрели отличия от предков, а менее родственные виды будут иметь меньше общих черт.
Поначалу филогении строились вручную: небольшое количество видов или родов сравнивалось по паре десятков признаков и из этих данных собиралось дерево. У такого метода имелось две очевидные проблемы: он занимал чудовищно много времени, а объем учитываемой информации был в лучшем случае невелик. Сравнение вручную примерно десятка видов по 20 или 30 признакам могло занять целый день, и, конечно же, всегда оставался риск субъективной ошибки. Это стало явным улучшением старой системы простого выбора нескольких почти произвольных признаков и присвоение им важного статуса, но переход от пяти-шести черт к паре десятков на самом деле не был таким уж грандиозным скачком вперед, притом что сейчас (в отличие от прошлого) можно было бы обеспечить большее разрешение метода и в теории определить точные родственные связи между любыми тремя-четырьмя видами. Без какой-либо автоматизации этот процесс неизбежно оказывался очень медленным.
Будучи студентом, я именно так учился строить филогении в не столь уж далеком 1997 г. – составлял эволюционные деревья вручную. Это было довольно увлекательное занятие, давшее мне действительное понимание используемых данных и того, как изменения или дополнения к этим данным могут повлиять на итоговое дерево. Но сам процесс неминуемо должен был быть вытеснен компьютерным расчетом.
Это заняло больше времени, чем вы могли бы подумать. К этому моменту компьютеры в науке распространились повсеместно и программы для филогенетических построений были доступны, однако количество расчетных вариантов даже для небольших деревьев поражало воображение. Как в задаче с даром махараджи (предполагалось положить одно зернышко риса на первую клетку шахматной доски, два на вторую и т. д.), числа становились очень большими, причем чрезвычайно быстро. Для трех видов есть только три возможных результата: А и В ближе друг к другу, чем С; В и С ближе, чем А; и наконец, А и С ближе, чем В. Однако для четырех видов уже возможны 15 комбинаций (А и В, потом С, потом D; А и В, потом D, потом С; А и В, потом С и D как вторая пара и т. д.), а пять видов дают уже 105 возможных деревьев, когда же вы берете всего лишь десять видов, то получаете уже 34,5 млн комбинаций. А если рассматривать десятки или сотни видов, то проблематично даже подсчитать количество возможных деревьев. В итоге прошло значительное время, прежде чем вычислительная мощность среднего персонального компьютера позволила обрабатывать объемы информации, и прежде чем программное обеспечение стало достаточно совершенным, чтобы быстро исключать громадное количество заведомо неправильных деревьев перед тем, как тщательно изучить относительно небольшое число вариантов и выбрать лучший.
В наши дни обычным стал анализ ископаемых групп позвоночных, где десятки видов сравниваются по сотням признаков. Недавние анализы тираннозавров охватывают практически все известные виды и более 300 признаков, описывающих всё: от формы верхнечелюстной кости и дентикул на зубах до места прикрепления мышц таза, формы лодыжки и строения черепной коробки. Иными словами, было собрано и обработано колоссальное количество данных, и почти все особенности, какие только удалось обоснованно выделить, проанализировали и сопоставили. Сведе́ние всех этих данных воедино означает, что результаты такого анализа – лучшие, какие вообще возможно получить в имеющихся обстоятельствах (недостаток данных, например от неполных скелетов, может создавать сложности и говорить о том, что мы не в состоянии всегда учесть все, и, разумеется, поскольку детеныши могут выглядеть не так, как взрослые животные, им следует уделять особое внимание).
Поставим тираннозавров на место (эволюционное)
Давайте начнем с динозавров в целом и с того, где главные игроки располагаются на эволюционном дереве или по крайней мере на его участке в пределах ближайших к нам разветвлений группы рептилий. Тираннозавры – всего лишь одна группа (или клада – более точный эволюционный термин) тероподов, и важно понимать, как они связаны с другими формами. Это ключ к определению, какие черты уникально тираннозавровые, а какие унаследованы ими от предков. Как можно видеть на диаграмме ветвящегося дерева, тероподы в действительности находятся ближе всех к завроподоморфам: группе, в которую входят все крупнейшие и знаменитые гиганты с длинными шеями, такие как диплодок и апатозавр. Все прочие динозавры, включая рогатых динозавров, броненосных динозавров и гадрозавров, относятся к кладе птицетазовых (рис. 6). Динозавры, по сути, являются довольно близкими родственниками нединозавровых птерозавров (часто неверно называемых птеродактилями, а также иногда ошибочно считающихся предками птиц), и крокодиловых – группы, теперь значительно уменьшившейся, но прежде весьма разнообразной. Птицы в очень многих аспектах являются динозаврами и действительно произошли от группы птицеподобных динозавров, называемых дейнонихозаврами.
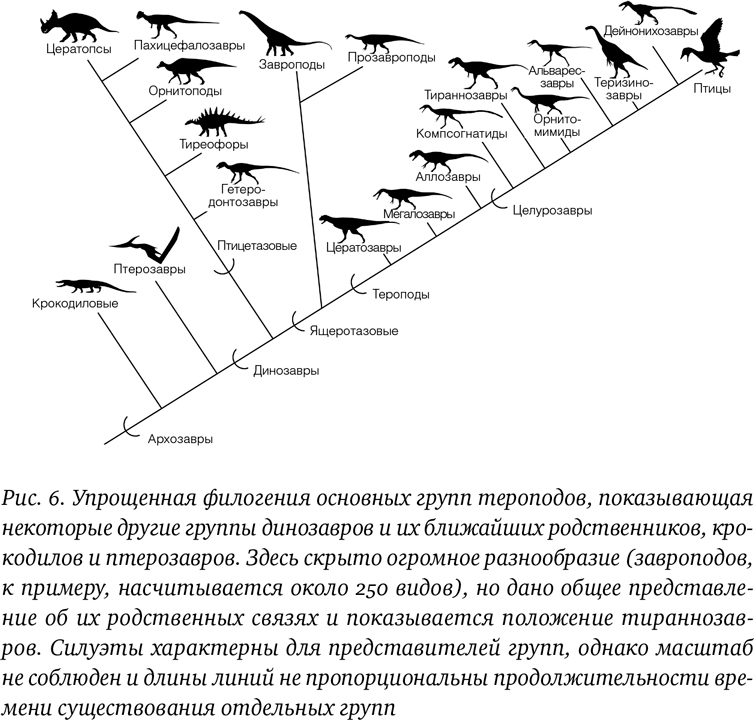
Эти деревья заведомо несколько упрощены: не все живые организмы располагаются аккуратными группами, и есть отдельные виды, которые размещаются между показанными на рисунке, а также виды, чье положение до конца не ясно. Тем не менее схема адекватно отображает общую согласованную позицию палеонтологов по поводу положения этих групп. Силуэты здесь и ниже дают общее представление о внешнем виде животных в соответствующих кладах, хотя во многих случаях разнообразие форм и размеров оказывается существенно преуменьшенным (например, среди птиц есть павлины, попугаи, киви, пингвины, страусы, колибри и альбатросы).
Внутри группы тероподов тираннозавры занимают среднее положение. Они были не первой появившейся группой, но и не последней. Названия клад часто даются по первому или самому известному ее представителю (тираннозавры, как понятно, названы по тираннозавру, аллозавры по аллозавру и т. д.), но остальные оказываются для непосвященного более непонятными: например, легендарный велоцираптор является дромеозавридом, и мало кому знакомо название теризинозавроиды. Как можно видеть, ближайшие родственники тираннозавров – компсогнатиды и орнитомимозавры, и тираннозавры близки к базальной группе, называемой целурозавры, что означает «полохвостые ящеры». Хотя теперь известно, что полые кости существовали задолго до возникновения этой группы, название дано вследствие обилия пневматических, как у птиц, костей, характерных для всех представителей этой крупной клады.
Возьмем тираннозавров «крупным планом» и рассмотрим схему, изображающую их «стандартную» филогению (рис. 7а). На ней видны три главных ветви, обозначающие отдельные клады, которые представляют собой результат эволюционного расхождения предшествующих клад. Раньше всех отходит ветвь процератозавридов, а далее внутри тираннозавридов появляются группы алиораминов, альбертозавринов и тираннозавринов. Несмотря на некоторую неуверенность в возрасте горных пород, сохранивших останки разных тираннозавров, в целом эволюционная схема данной филогении соответствует датам, когда эти группы появляются в палеонтологической летописи. То есть линии, которые, по нашим предположениям, возникли позже, действительно появляются в более поздних, более близких к современности породах. Кстати, это одна из причин считать, что кладистика дает в целом точные результаты. Исследования показали, что точки ветвления наших деревьев для всех видов ископаемых организмов в среднем совпадают со временем первого появления различных окаменелостей, чем повышают уверенность в анализе.
Тем не менее неизбежно возникают разногласия в вопросе точного расположения групп в разных анализах. Разные исследователи используют различные комбинации признаков и даже видов в своих работах и получают немного отличающиеся деревья. Например, некоторые деревья предлагали поместить бистахиэверсора в группу тираннозавринов – положение, довольно сильно отличающееся от показанного на рисунке выше. В целом, однако, все было довольно стабильно, по крайней мере до самых недавних пор.

Во-первых, в 2012 г. аргентинский палеонтолог Фернандо с коллегами предположил, что тираннозавры могли оказаться значительно большей группой, чем представлялось ранее. Они предположили, что еще несколько видов были или очень тесно связаны с кладой тираннозавроидов или даже входили в нее. Этот ряд видов включает загадочного мегараптора из меловых отложений в Аргентине и фукуираптора из Японии – животных, обычно считавшихся частью группы аллозавроидов и на настоящий момент в целом признанными представителями этой группы. Есть определенные анатомические особенности, говорящие в пользу данной гипотезы, но она довольно сильно контрастирует с предыдущими анализами и аллозавроидов, и тираннозавроидов. Разумеется, это предположение не является совсем невероятным, но мы пока отложим его в сторону.
Вторая гипотетическая возможность большого сдвига парадигмы возникла с обнаружением литронакса в 2013 г., и с этим крупным новым родом появился новый анализ эволюции тираннозавров. В статье, описывающей это животное и дающей ему название, авторы, Марк Лёвен с коллегами, представили дерево, чуть ли не радикально отличающееся от предыдущих интерпретаций. В этой работе многочисленные таксоны передвинулись со своих «стандартных» позиций, и расстановка тираннозавров стала заметно иной. У этого есть некоторые довольно важные следствия, в частности, говорящие о том, как разные тираннозавры и особенно тираннозаврины могли распространяться по миру и перемещаться между континентами. Этот вопрос рассматривается далее в главе о тираннозаврах во времени и пространстве, но с эволюционными следствиями мы разберемся здесь.
Глядя на филогению Лёвена, мы видим, что между группами произошли перестановки (рис. 7b). Основные ветви никуда не делись, и большинство их представителей остались на местах (к примеру, тираннозавр и дасплетозавр по-прежнему тираннозаврины, альбертозавр и горгозавр – альбертозаврины, гуаньлун – процератозаврид), но другие ветви переместились. Например, бистахиэверсор теперь стал тираннозаврином, алиорам перестал быть представителем тех же тираннозавринов, а тератофоней оказался значительно ближе к тираннозавру, чем считалось раньше.
Разумеется, интересно знать, как мы представляем себе связи разных родов и видов животных друг с другом, но филогении также дают нам возможность увидеть то, как эволюционировали конкретные признаки, когда они появились, степень их изменения, и ставить вопросы типа «каких общих черт у групп больше всего?». Это позволяет нам составить представление об эволюции групп, но при другом расположении родов наши интерпретации этих схем также изменятся. Один важный признак, к которому мы вернемся позже, – длина передних лап у различных тираннозавров, но нагляднее будет привести краткий пример.

В то время как тираннозавр и другие тираннозаврины знамениты очень маленькими передними лапами и кистями, у ранних форм этих животных передние конечности были сравнимы по размеру с конечностями других тероподов. Фактически существует весьма устойчивая тенденция уменьшения передних конечностей от первых тираннозавровых к тираннозавринам, но если, к примеру, новый анализ предполагает, что тарбозавр на самом деле был процератозавридом, нам придется считать, что передние лапы значительно уменьшались не однажды, а дважды. Мы внезапно получаем вид процератозавридов, который просуществовал на много миллионов лет дольше, чем мы думали раньше, и внешне был похож на тираннозавра. Такая резкая перемена чрезвычайно маловероятна (у тарбозавра слишком много общих черт с другими тираннозавринами, чтобы были опровергнуты их родственные связи), но этот гипотетический пример демонстрирует важность понимания родственных отношений между видами.
Эти и подобные анализы характерны для современных палеонтологических исследований, потому что позволяют проследить изменения во времени и пространстве, т. е. серьезные возражения против «стандартной» филогении, подобные филогении Лёвена, и интригуют, и вызывают сомнения. Верной может быть только одна модель эволюции тираннозавров (возможно, конечно, не верна ни одна из ныне известных), и для специалистов по тираннозаврам это богатейшая область исследования.
Далее в книге я буду для ясности придерживаться «стандартной» модели как основания для всего, касающегося эволюции тираннозавров, тем не менее в некоторых областях, где уместно, я ссылаюсь и на филогению Лёвена.
Теперь мы знаем, кто такие тираннозавры и чем характеризуются они сами и их эволюционная история, но эта группа просуществовала 100 млн лет на трех, а возможно, и на пяти континентах. Как именно эти клады распределялись по столь огромному промежутку времени и гигантской территории, какие виды и группы когда появились – наша следующая тема.

