II. Нейрональные корреляты сознания (НКС)
Введение: что такое «нейрональные корреляты сознания» (НКС)?
Цель когнитивной нейронауки – обнаружить нейрональные корреляты и биологические механизмы, лежащие в основе когнитивных феноменов. Чтобы собрать эмпирические данные, которые помогли бы это сделать, когнитивная нейронаука использует методы, позволяющие измерять различные аспекты деятельности мозга. Такие измерения проводятся в то время, когда испытуемый решает те или иные интеллектуальные задачи. Поэтому они отражают активность мозга, связанную с той или иной умственной деятельностью. Методы когнитивной нейронауки (ЭЭГ, МЭГ, функциональная магнитно-резонансная томография (фМРТ), позитронно-эмиссионная томография (ПЭТ) и транскраниальная магнитная стимуляция (TMS)), можно использовать и для выявления нейрональных механизмов сознания. В последние годы поиск нейрональных коррелятов сознания стал популярной сферой исследований на стыке когнитивной нейронауки и науки о сознании.
Когда в нашей субъективной ментальной жизни что-то происходит – мы переживаем взрыв эмоций, чувствуем боль, видим цвета – одновременно что-то обязательно происходит и в мозге. Кажется, в нашей сознательной жизни не может быть феноменальных переживаний, которые бы не сопровождались соответствующей нейрональной активностью в мозге. Описывая такую взаимосвязь между разумом и мозгом, философы говорят о дополнительных отношениях, связывающих между собой активность сознания и активность мозга: для каждого события в сознании есть соответствующее событие в мозге. Это – принцип ковариации между сознанием и мозгом. Но обратите внимание, что эта связь действует только в одном направлении: обратное утверждение – что для каждого события в мозге есть соответствующее событие в сознании – не истинно. Изменения в активности мозга, отвечающие за неосознаваемую деятельность, могут происходить без участия сознания, но изменения в сознании обязательно сопровождаются какими-то изменениями в активности мозга.
Кроме того, такие зависимые отношения включают в себя также и предположение о том, что события в сознании так или иначе являются результатом событий в мозге и поэтому неотделимы от них. Это принцип онтологической зависимости сознания от мозга. Не может быть никакого сознания, если нет мозга, но мозг, конечно, может существовать без всякого сознания.
Специалисты по когнитивной нейронауке, очевидно, воспринимают подобные дополнительные отношения как нечто само собой разумеющееся. В соответствии с принципом онтологической зависимости они предполагают, что сознание без мозга невозможно, тогда как мозг без сознания возможен. Если бы сознание могло существовать независимо от мозга, мы наблюдали бы своего рода картезианский дуализм. Но у нас нет никаких веских доказательств того, что события в сознании могут существовать независимо от событий в мозге. Поэтому нам приходится верить, что события в сознании являются результатом событий в мозге.
Однако обратите внимание, что зависимые отношения ни в коем случае не объясняют, почему сознание и мозг связаны между собой таким особым образом. Кстати, ковариация вместе с онтологической зависимостью совместимы с теориями редуктивного и эмерджентного материализма. И одно возможное объяснение того, почему зависимые отношения признаются истиной, состоит в том, что либо редуктивный, либо эмерджентный материализм адекватно описывает реальность. Но этот философский вопрос, конечно, остается открытым.
Зависимые отношения могут лечь в основу программы неврологических исследований сознания. На основании принципа ковариации исследователи в области когнитивной нейронауки считают, что каждый субъективный феномен в сознании обязательно должен коррелировать с какими-то объективными неврологическими явлениями в мозге. Поэтому имеет смысл попытаться обнаружить с помощью объективных измерений все, что только возможно, о нейроанатомических и нейрофизиологических деталях единения сознания и мозга.
Что же мы имеем в виду, когда говорим, что события двух разных типов коррелируют друг с другом? Если два события коррелируют, они обычно происходят одновременно. Если одно из них появляется, исчезает, качественно или количественно изменяется, то же самое происходит и с другим событием, примерно в то же время. Поэтому возможно обнаружить, с помощью объективного измерения деятельности мозга, какие неврологические события точно коррелируют с теми или иными событиями в сознании. Это был бы первый разумный шаг к созданию теории отношений между сознанием и мозгом, основанной на эмпирических данных. Поэтому поиск нейрональных коррелятов сознания стал основным подходом к исследованиям сознания в нейронауке.
Долгосрочная цель таких исследований – выяснить, какие же неврологические события коррелируют с теми или иными состояниями и содержанием сознания. В конечном счете такие исследования должны определить, каковы минимальный достаточный объем нейронов или активность нервной системы, которая обязательно сопровождает то или иное осознаваемое переживание. Это стандартное определение концепции «нейрональных коррелятов сознания» (или НКС) (Кох, 2004).
Чтобы достичь этой цели и обнаружить НКС, необходимо описать как характер осознаваемых событий, так и характер событий в мозге, а затем попытаться соединить эти описания с помощью подробного словаря или карты. Во-первых, чтобы по отдельности изучать корреляты каждого типа феноменов сознания, необходимо создать таксономию или подробную классификацию различных уровней и содержания сознания. Во-вторых, нужно разработать эксперименты, в которых нейрональные корреляты сознания каждого типа можно было бы измерить независимо от других типов. В когнитивной нейронауке активно проводятся такие исследования.
Глава 7
Методы и эксперименты в сфере НКС
Введение: как разработать эксперимент по исследованию НКС
Методы функциональной томографии мозга: фМРТ и ПЭТ
Электромагнитное исследование мозга с помощью ЭЭГ и МЭГ
Выводы
Вопросы для обсуждения
Введение: как разработать эксперимент по исследованию НКС
Основной принцип, лежащий в основе экспериментов НКС, очень прост. Если вы хотите обнаружить нейрональные корреляты осознанного состояния или содержания сознания, следует разработать эксперимент, где есть два разных условия. Одно называется контрольным (или базовым ) условием. При этом условии состояние сознания, или содержание сознания, полностью отсутствует в психике испытуемого. Это условие служит нейтральной базовой линией , которому противопоставляется другое условие. Другое условие называется экспериментальным : определенное состояние сознания, или содержание сознания, четко присутствует в психике испытуемого. В идеале все остальные параметры одинаковы или не меняются в обоих условиях: единственное отличие – наличие или отсутствие содержания в сознании испытуемого.
Такая схема создает необходимую управляемую вариативность на уровне феноменов сознания. Чтобы получить информацию об одновременных событиях в мозге, активность мозга испытуемого нужно измерить тем или иным способом. Самые распространенные методы измерений можно разделить на две группы: методы функциональной томографии мозга и электромагнитное исследование мозга .
Методы функциональной томографии мозга: фМРТ и ПЭТ
Методы функциональной томографии мозга включают функциональную магнитно-резонансную томографию (фМРТ) и позитронно-эмиссионную томографию (ПЭТ). В экспериментах с НКС раньше использовались оба метода, но сейчас фМРТ используется чаще, потому что он дешевле и проще. При фМРТ испытуемый находится в специальной капсуле, где создано сильное статическое магнитное поле, и его мозг бомбардируют короткие последовательности высокочастотных (радиочастотных) электромагнитных импульсов. Магнитное поле и импульсы как таковые безопасны для мозга, но сочетание магнитного поля и импульсов влияет на субатомные частицы (протоны) в ядрах атомов водорода в мозге, побуждая их «отражать» эти импульсы. Такое «эхо» можно зарегистрировать с внешней поверхности головы испытуемого.
Сигналы «эха» отражают точное расположение и магнитные свойства различных биологических тканей и структур мозга. Обычный или структурный фМРТ со здает трехмерное анатомическое изображение мозга в высоком разрешении; фМРТ, кроме этого, отражает изменения в количестве свежей, наполненной кислородом крови в мозге. Это называется BOLD-сигналом (то есть «сигналом, зависящим от уровня кислорода в крови (Blood Oxygen Level Dependent)»). Свежая кровь быстро поступает в те области мозга, где увеличивается активность нейронов, и изображение косвенно отражает, в каких областях мозга растет нейроэлектрическая активность нейронов. Таким образом, в НКС-исследованиях с помощью метода фМРТ, сравнивая различия в деятельности мозга в экспериментальных и контрольных условиях, можно видеть, в каких областях мозга нейроны стали более активными, отражая изменения, происходящие в сознании.
Минимальное время, в течение которого описывается отдельное изображение, называют временны м разрешением . Нужно несколько секунд, чтобы кровоток среагировал на всплеск локальной нейрональной активности; таким образом, в лучшем случае фМРТ отражает изменения в активности мозга с задержкой примерно в 5–10 секунд. Первые изменения в сигнале фМРТ возникают через несколько секунд после начала возбуждения, но реакция фМРТ достигает максимума примерно через 10 секунд после начала активации.
Это означает, что у фМРТ относительно невысокое временно е разрешение) . Активная интеллектуальная обработка и нейрональная активность происходит в первые две секунды после начала возбуждения, но фМРТ слишком медлительна, чтобы точно увидеть, что происходит. Наоборот, пространственное разрешение фМРТ относительно высоко. Пространственное разрешение – это минимальный размер элемента изображения (он называется «пикселем», или «voxel») в отдельной области изображения с точки зрения количества полученных сигналов (они интерпретируются как «активность мозга»). Пространственное разрешение изображений фМРТ составляет около 2–3 мм2.
В исследованиях по методу ПЭТ радиоактивные изотопы, испускающие позитроны, присоединяются к заранее выбранным исследователем молекулам (например, радиоактивный кислород присоединяется к молекуле воды H2O, что создает радиоактивную воду), а затем поставляются в кровоток испытуемого. Оттуда радиоактивные молекулы быстро распространяются по всему телу, и в том числе попадают в мозг. Радиоактивные изотопы непостоянны и распадаются с известной скоростью, испуская позитроны. В мозге позитроны сталкиваются с электронами, обе частицы уничтожаются и превращаются в энергию в форме двух гамма-лучей, исходящих из мозга в прямо противоположных направлениях.
ПЭТ-устройство – это кольцо датчиков гамма-лучей, которое надевают на голову испытуемого. Оно обнаруживает гамма-лучи, исходящие из мозга, и на их основании вычисляет, в каких областях мозга находились молекулы с радиоактивными элементами в момент распада. В зависимости от того, какие молекулы используются (воды, глюкозы, производные дофамина и т. д.), их распределение в мозге отражает определенный аспект метаболизма или активности мозга (например, кровотока, метаболизма глюкозы или взаимодействия нейромедиаторов). В большинстве НКС-экспериментов с использованием ПЭТ маркером для кровотока была радиоактивная вода.
При этом отражаются те же аспекты активности мозга, что и при фМРТ: кровь приливает к тем областям, где нейроны электрически и метаболически более активны. ПЭТ еще медленнее, чем фМРТ, и в лучшем случае отдельное изображение может показать лишь общий рисунок изменений, которые произошли в течение 30–60 секунд. Пространственное разрешение, в лучшем случае, составляет несколько миллиметров, обычно это немного ниже, чем при фМРТ.
Электромагнитное исследование мозга с помощью ЭЭГ и МЭГ
Электроэнцефалография (ЭЭГ) и магнитоэнцефалография (МЭГ) – методы электромагнитного исследования мозга. Они регистрируют сигналы, возникающие при электрической активности нейронов. Нейрональные импульсы создают диполярную электрическую циркуляцию в мозге. Диполь – это источник электричества с двумя полюсами, отрицательным и положительным. Внутри нейрона электрический ток идет от дендритов к телу клетки (это называется постсинаптическим потенциалом) и дальше, от тела, вдоль аксона (это называется потенциалом действия). Такие внутриклеточные потоки невозможно обнаружить за пределами мозга, но, к счастью, извне в клетку идет обратный, или объемный, ток, который завершает электрическую цепь. Обратный ток течет вне нейронов, назад, к противоположному электрическому полюсу. Обратные токи свободно текут в мозге и непредсказуемо распространяются по его тканям, потому что идут по нерегулярным путям, определяемым наименьшим электрическим сопротивлением внутри мозга .
Иногда обратные токи достигают черепа и проходят сквозь него в скальп.
Присоединив к скальпу электроды ЭЭГ, можно обнаружить эти слабые электрические токи в виде волн электрической активности мозга. Изменения в электрическом потенциале волн можно очень точно измерить во временном отношении, до миллисекунд (временно е разрешение ЭЭГ – одна тысячная секунды). К сожалению, чрезвычайно сложно точно определить, в какой области мозга находятся нейроны, активность которых создала зарегистрированные волны, ведь прежде чем дойти до электродов на скальпе, токи, как правило, проходят в мозге длинный и непредсказуемый путь, и в этом путешествии токи, исходящие из множества разных источников и областей, взаимодействуют, сливаются друг с другом, ослабляют друг друга или настолько сплетаются, что источник сигнала расшифровать уже не удается.
Точное предположение можно сделать только с помощью исходной модели локализации , вычислив наиболее вероятные области возникновения источников, которые создали зафиксированный паттерн ЭЭГ на скальпе. С точки зрения нейроанатомической локализации пространственное разрешение ЭЭГ измеряется в квадратных сантиметрах, а не в миллиметрах, – у ЭЭГ очень низкое пространственное разрешение по сравнению с фМРТ или ПЭТ.
В отличие от ЭЭГ МЭГ измеряет магнитные поля , создаваемые электрической активностью нейронов. Нейромагнитное поле генерируется той же самой электрической активностью, что и нейроэлектрическое поле, определяемое с помощью ЭЭГ, но нейромагнитное поле ведет себя совершенно иначе. С одной стороны, измерить его легче, а с другой – труднее, чем ЭЭГ. Легче, потому что нейромагнитное поле распространяется непосредственно из источника своего возникновения, оно не подавляется, не искажается мозгом или черепом. Соответственно точное место его происхождения можно обнаружить намного легче, чем при ЭЭГ. Но при этом нейромагнитные сигналы вначале настолько слабы, что лишь некоторые из них выходят за пределы мозга.
Таким образом, все обнаруженные сигналы МЭГ отражают активность только тех областей коры головного мозга, которые расположены близко к поверхности мозга и черепа. Кроме того, нейромагнитное поле направлено только в одну сторону, перпендикулярно к электрически активным нейронам. Только тем нейронам, которые ориентированы так, что их нейромагнитное поле направлено в сторону черепа, удается создать магнитное поле, которое можно измерить на поверхности черепа. Остальные нейромагнитные поля датчики МЭГ обнаружить не могут. Следовательно, метод МЭГ «слеп» к тем видам нейрональной активности, которые может регистрировать ЭЭГ. Но для кортикальных сигналов, которые МЭГ может регистрировать, его временное разрешение не меньше, чем при ЭЭГ (миллисекунды), а пространственное разрешение – несколько миллиметров.
Эксперимент НКС, проведенный с помощью ЭЭГ или МЭГ, может выявить, сколько времени нужно, чтобы сенсорно-перцептивная информация о стимуле достигла сознания. Эти методы позволяют точно отслеживать изменения, происходящие со временем: точность измерения составляет до тысячи непрерывных точек данных в секунду! Таким образом, можно увидеть, что в какой-то момент, при условии, когда информация входит в сознание и возникает то или иное переживание, электромагнитная реакция в мозге начинает отличаться от той, которая была зафиксирована в контрольном условии, когда информация не достигала сознания.
Это – критический момент времени, когда возникает нейрональная активность НКС; вся активность, имевшая место до этого момента, отражает обработку информации вне сознания. Кроме того, поскольку сигналы, которые регистрируют ЭЭГ и МЭГ, очень сложны и включают много различных частот электромагнитной энергии, можно выяснить, какой тип электромагнитной энергии связан с нейропсихологической активностью НКС. Точки на скальпе, где регистрируются реакции, также примерно отражают локализацию областей мозга, где, вероятно, возникла активность, но локализация реакции с точки зрения анатомии мозга весьма сложна, особенно для ЭЭГ.
Выводы
Кратко суммируем типичный эксперимент НКС: во время эксперимента испытуемый несколько раз оказывается под воздействием двух разных условий: наличие сознания (он переживает определенное состояние или содержание) и отсутствие сознания (испытуемый не переживает этого состояния или содержания). Во всем остальном условия эксперимента остаются идентичными. Активность мозга испытуемого одновременно измеряется с помощью фМРТ, ПЭТ, ЭЭГ или МЭГ. Методы функциональной томографии мозга ПЭТ и фМРТ отражают картину того, что происходит с метаболизмом или кровотоком мозга, и позволяют точно определить место этих изменений с точки зрения анатомии мозга, но не с точки зрения времени.
Методы электромагнитного исследования мозга ЭЭГ и МЭГ отражают нейрональные, биоэлектрические сигналы мозга и могут точно определить начало изменений в этих сигналах во времени, но не могут точно определить локализацию их возникновения с точки зрения анатомии мозга. Все эти методы позволяют обнаружить НКС. Эксперимент НКС всегда должен быть направлен на выявление какого-то определенного состояния или содержания сознания, давать информацию только об одном, совершенно определенном аспекте сознания. В следующих двух главах мы кратко опишем некоторые типичные эксперименты и их результаты, не вдаваясь (в довольно сложные) подробности.
Вопросы для обсуждения
1. Действует ли принцип дополненности в отношениях между сознанием и мозгом? Приведите примеры:
+ человеческого мозга без всякого сознания;
+ активности человеческого мозга без какого-либо соответствующего осознаваемого переживания.
Следующее, напротив, продемонстрировать невозможно. Вы согласны?
+ (человеческое) сознание (или любой тип сознания) без мозга;
+ событие в сознании (субъективное переживание в субъективной психологической реальности) без каких-либо соответствующих нейрональных событий в мозге.
2. Придумайте эксперимент для изучения нейрональных коррелятов сознания:
+ Какими будут контрольные условия?
+ Какими будут экспериментальные условия?
+ Какие методы томографии мозга вы будете использовать и что они покажут в результате эксперимента?
Глава 8
Исследования нейрональных основ сознания как состояния
Введение: сознание как состояние
Анестезия
Эпилептические припадки и глубокий сон
Синдром «запертого человека» (синдром изоляции)
Вегетативное состояние и другие глобальные расстройства сознания
Инвертированные зомби
Выводы
Вопросы для обсуждения
Введение: сознание как состояние
Сначала давайте поговорим о том, каким был бы идеальный эксперимент в исследовании нейрональных коррелятов сознания как состояния. В главе 3 мы определили состояние наличия сознания как фоновое состояние мозга или разума, которое позволяет испытывать субъективное переживание любого типа в целом и отсутствие которого (то есть бессознательное состояние ) абсолютно препятствует переживанию субъективного опыта любого типа. Идеальным экспериментом, позволяющим выяснить, что именно в мозге «освещает» сознание, был бы эксперимент, при котором сознательное состояние можно «включать» или «выключать» контролируемым образом.
Анестезия
Один из способов создания контролируемых сдвигов от состояния сознания к бессознательному состоянию – использование обезболивающих веществ для того, чтобы устранить сознание. Влияние обезболивающих веществ таково, что под воздействием анестезии человек сначала становится очень сонным (как будто он очень пьян), а затем прекращает реагировать на какие бы то ни было команды и даже на болевые стимулы (кстати, поведенческое определение бессознательного состояния в анестезиологии звучит примерно как «отсутствие реакций или неспособность двигаться в ответ на команду и амнезия определенных событий», а не как наше определение полного отсутствия субъективного переживания). Сигналы ЭЭГ человека, подвергшегося анестезии, также замедляются и становятся похожими на сигналы ЭЭГ глубокого сна, который также называют медленноволновым сном. При очень глубокой анестезии сигнал ЭЭГ может вообще исчезнуть (тогда возникает плоская ЭЭГ, прямая линия без каких-либо волн).
Объективно анестезия приводит к отсутствию реакции на внешние стимулы. Субъективно анестезия обычно (но не всегда) приводит к полному отсутствию феноменального сознания. Во время действия анестезии нет никакого переживания субъективного опыта, и даже чувство времени исчезает. Так что человеку, проснувшемуся после нескольких часов анестезии, может казаться, что он проспал всего несколько минут. Редкие исключения из бессознательного состояния это осознание анестезии – вызванные внешними стимулами переживания (ощущения, восприятие), отражающие то, что действительно происходит в операционной, и анестетический сон ( anaesthesia dreaming ) – внутренне порождаемое переживание , сопровождающееся галлюцинациями или сновидениями, никак не связанными с тем, что происходит с человеком.
Об этих двух типах субъективных переживаний больные иногда сообщают после пробуждения от анестезии. Но даже если пациент говорит, что ничего не помнит, возможно, какие-то переживания у него все-таки были. Иногда больные могут общаться с окружающими во время анестезии, двигая рукой (это называют методом изолированной руки), но потом говорят, что не помнят об этом. Некоторые пациенты могут это делать, даже несмотря на то что показатели ЭЭГ, отражающие глубину анестезии («bispectral индекс»), указывают, что их мозг находится в бессознательном состоянии. Возможно, ЭЭГ в этом случае недостаточно чувствительна к нейрональным процессам, лежащим в основе сознания (Alkire, Hudetz, & Tononi, 2008).
Точно регулируя концентрацию обезболивающего вещества в мозге испытуемого, можно медленно угнетать сознание или медленно позволить сознанию снова появляться. Возникает вопрос: что происходит в мозге, когда сознание исчезает (или вновь появляется) в результате действия обезболивающих веществ? Это исследовали с помощью метода ПЭТ. Результаты нескольких исследований (в которых использовались разные обезболивающие вещества) оказались сходными: когда сознание исчезает, происходит снижение активности таламуса (Alkire & Miller, 2006). Таламус – это структура, расположенная в центре мозга, через которую сенсорная информация поступает в кору мозга. Таламус чрезвычайно тесным и сложным образом связан с корой, и эта связь представляет собой двунаправленные петли.
Таламус и кора отправляют сигналы друг другу, туда и обратно, в виде сложных паттернов. Иногда эти связи локальны: определенные участки коры или таламуса связаны с определенными участками друг друга. Иногда они диффузны и глобальны, так что один участок таламуса может устанавливать обширные и сложные связи с разными участками по всей коре. Уменьшение активности таламуса, вызванное обезболивающими веществами, возможно, устраняет нейроэлектрическую активность в таламокортикальных петлях, особенно в тех, которые связаны с более высокими частотами ЭЭГ (бета– и гамма-диапазона).
Области коры головного мозга, которые представляются наиболее важными для сознания и которые взаимодействуют с таламусом в момент возвращения сознания, расположены в задней коре: в ассоциативной области на пересечении латеральной височной, затылочной и париетальной коры и в центральной париетальной коре (Alkire, Hudelz & Tononi. 2008). Высокочастотная нейроэлектрическая активность в областях, связывающих эти области коры мозга с таламусом, по-видимому, необходима для состояния сознания, возможно, потому что они интегрируют отдельные содержания сознания в единое целое (Alkire, Hudetz & Tononi, 2008).
Эпилептические припадки и глубокий сон
Новые доказательства обязательного наличия высокочастотной электрической активности между корой и таламусом дают исследования эпилептических припадков и глубокого сна, лишенного сновидений (3-я и 4-я стадии медленноволнового сна). В обоих случаях все или почти все содержание сознания временно исчезает. При малом эпилептическом припадке это происходит резко и внезапно: пациент просто замолкает, иногда на середине фразы, и смотрит в пустоту в течение нескольких секунд. Когда припадок проходит, он может продолжить говорить с того же места, где замолчал. Во время такого припадка сознание, по-видимому, отключается и исчезает. В то же время на ЭЭГ можно увидеть выраженные медленные волны (3 Гц). Это означает, что таламокортикальные связи внезапно прерываются патологической медленноволновой активностью. Этот объективный феномен, заметный на ЭЭГ, на субъективном уровне выражается в отсутствии сознания.
Существует много различных типов эпилептических припадков, но есть гипотеза, что все припадки, серьезно ослабляющие или полностью устраняющие сознание, вызваны медленноволновой корковой активностью на на ЭЭГ, – подобные паттерны ЭЭГ возникают при анестезии, во время естественного глубокого сна или в коме. Полная потеря сознания происходит, когда патологическая активность охватывает большие сети между корой и областями подкорки (Cavanna & Mo naco, 2009; Yu & Blumenfeld, 2009).
Похожий паттерн ЭЭГ с большой амплитудой медленных волн (дельта-волн, 0,5–3 Гц) можно наблюдать на самых глубоких фазах сна (3-я и 4-я фазы медленноволнового сна). Эти фазы обычно не сопровождаются сновидениями: если после пробуждения человек сообщает о каких-то осознаваемых переживаниях, они обычно минимальны, статичны и очень просты по содержанию.
Метафорически говоря, когда феноменальные огни «включены», то таламокортикальные (или подкорково-корковые) двунаправленные нервные петли демонстрируют сочетание высоких частот сложной биоэлектрической активности; а когда эти огни «выключены», то эти петли захвачены медленными волнами большой амплитуды, указывающими на синхронизированную и менее сложную или менее дифференцированную таламокортикальную активность.
Синдром «запертого человека» (синдром изоляции)
Если для существования сознания как состояния нужна только нормальная таламокортикальная активность, возникает вопрос: можно ли быть в сознании, но при этом оказаться «запертым» в мозге, то есть не иметь возможности общаться с внешним миром? Да, это возможно. Такое состояние описано в литературе по неврологии и получило название синдрома «запертого человека» (синдрома изоляции). Пациент, находящийся в таком прискорбном состоянии, совершенно неподвижен и не реагирует ни на какие стимулы. В лучшем случае он может общаться с внешним миром с помощью движений глаз или моргания. При этом он находится в полном сознании.
Синдром «запертого человека» – это не расстройство сознания, а только отсутствие возможности двигаться. На ПЭТ-изображениях, отражающих уровни метаболизма мозга, видно, что у пациентов с таким синдромом совершенно нормальный уровень метаболизма мозга и метаболическая кортикальная активность распределена точно так же, как у здоровых испытуемых из контрольной группы, где самый высокий уровень активности отмечается в медиальной задней коре (Laureys, Owen, & Schiff , 2004). (Кстати, именно эти области коры мозга оказываются «выключенными» в результате анестезии!) Больной не проявляет внешних реакций, но у него сохраняются осознаваемые феноменальные переживания и он способность думать.
Вегетативное состояние и другие глобальные расстройства сознания
Глобальные расстройства сознания – это состояния, при которых травма головного мозга или болезнь привели к глубоко бессознательному состоянию (в клинической медицине его называют «отсутствием реакции на стимулы»). Смерть мозга, кома, вегетативное состояние и минимальное состояние сознания представляют различную степень серьезности глобальных расстройств сознания.
На снимках, полученных с помощью функциональной томографии мозга, отражающих метаболизм мозга у пациентов, переживших смерть мозга, никакой метаболической активности не регистрируется – ткань мозга мертва, – и внутренняя часть черепа выглядит на таких изображениях однородно черной (Laureys, Owen, & Schiff , 2004)!
Больные, находящиеся в вегетативном состоянии , страдают от серьезного повреждения головного мозга, вызванного остановкой сердца, инсультом, травмами тела или черепно-мозговыми травмами. Они впадают в кому, но смерти мозга не происходит. Затем они выходят из комы и входят в вегетативное состояние. При этом они начинают спонтанно открывать и закрывать глаза (своего рода «бодрствование» и «сон»), но при этом не реагируют ни на какие внешние стимулы. Изображения метаболизма мозга, сделанные с помощью ПЭТ, при вегетативном состоянии демонстрируют ненормально низкие уровни метаболической активности (Laureys, Owen, & Schiff , 2004).
Ужасное положение, когда сознание оказывается «заточенным» в неподвижном теле, не реагирующем на внешние стимулы, как при синдроме «запертого человека», привело к дальнейшим вопросам о том, действительно ли пациенты, не реагирующие на внешние стимулы, не обладают сознанием? Возможно, некоторые из них остаются в сознании, но не проявляют этого внешне? Недавние эксперименты показали, что у некоторых пациентов, находящихся в вегетативном состоянии, может сохраняться сознание и поэтому им нельзя поставить диагноз «вегетативное состояние».
В одном эксперименте больных, находящихся в вегетативном состоянии, просили выполнить простые умственные задачи на воображение, например представить себе, что они играют в теннис или гуляют вокруг дома. Как ни странно, изображения фМРТ мозга некоторых пациентов показали точно такую же активность, как у здоровых испытуемых из контрольной группы, выполняющих ту же самую задачу! Казалось, пациенты могут намеренно создавать мысленные образы – субъективные сознательные переживания определенного типа – в соответствии с инструкциями, которые им дали, хотя при этом они никаким образом не могли общаться с исследователями (Owen et al, 2006). Этот эксперимент предоставляет убедительные доказательства того, что состояние сознания (а значит, и какое-то его содержание) может существовать даже у тех людей, которые не реагируют ни на какие внешние стимулы и которые, кажется, совершенно лишены сознания.
Когда больной начинает выходить из вегетативного состояния, он входит в нестабильное состояние, на грани между сознанием и его отсутствием. Поэтому иногда он может осмысленно реагировать на те или иные стимулы или на попытки общаться с ним, но эти реакции нестабильны и фрагментарны. Такое состояние называется минимальным состоянием сознания. Изображения ПЭТ при минимальном состоянии сознания показывают, что здесь метаболизм мозга выше, чем при вегетативном состоянии, но все еще ниже, чем у здорового мозга (Laureys, Оуэн, &Schiff ,2004).
Инвертированные зомби
В исследованиях сознания проблема, связанная с тем, сохраняется ли сознание при анестезии, в вегетативном состоянии и в других подобных ситуациях, в последнее время привлекает к себе все больше внимания. Для описания существ, обладающих сознанием, но при этом демонстрирующих все поведенческие признаки и реакции существ, не обладающих сознанием, была предложена новая концепция: инвертированный зомби (Mashour & LaRock, 2008). Это противоположность типичного философского зомби (который внешне кажется сознательным, но не обладает внутренним сознанием): инвертированные зомби внешне не проявляют признаков сознания, но обладают сознанием внутри. Пациенты, которые демонстрируют внешние признаки отсутствия сознания, как минимум, могут оказаться инвертированными зомби. Чтобы эмпирически доказать, являются они ими или нет, нам понадобилось бы устройство для томографии мозга, регистрирующее наличие (и отсутствие) феноменального сознания или квалиа в мозге.
Выводы
Нейрональные корреляты сознания как состояния можно исследовать, изучая влияние анестезии, эпилептических припадков, глубокого сна и глобальных расстройств сознания. Результаты таких исследований показывают, что таламус, таламокортикальные связи, корково-подкорковые петли и задние области коры мозга играют важную роль в возникновении сознания как состояния. Кроме того, в состоянии сознания обширные нейронные сети в этих областях создают высокочастотную электрическую активность. Если такая активность в этих сетях подавлена высокоамплитудными медленными волнами, сознание исчезает.
Когда мы исследуем испытуемых, на первый взгляд «не имеющих сознания», и разрабатываем эксперименты НКС, чтобы обнаружить НКС сознания как состояния, нужно соблюдать осторожность и отличать истинное отсутствие сознания (отсутствие феноменального сознания) от инвертированных зомби (отсутствие внешних реакций при наличии внутреннего феноменального сознания). Некоторые пациенты, не реагирующие ни на какие внешние стимулы, не лишены феноменального сознания.
Вопросы для обсуждения
Если анестезия просто лишает человека реакций (парализует) и приводит к амнезии, значит ли это, что человека, находящегося под воздействием анестезии, можно назвать «инвертированными зомби»: он полностью сохраняет сознание, но когда приходит в себя, то ничего не помнит о том, что происходило во время анестезии? Какая разница между истинным отсутствием сознания и только параличом в результате анестезии? Если бы вам предстояло перенести анестезию, хотели бы вы знать, что при этом будет с вашим сознанием?
Глава 9
Исследования нейрональной основы зрительного осознания
Введение: зрительная информация и зрительноео осознание
Исследования бинокулярной конкуренции (Binocular rivalry)
Зрительные галлюцинации
ЭЭГ и МЭГ исследования зрительного осознания
Транскраниальная магнитная стимуляция (ТМС)
Выводы
Вопросы для обсуждения
Введение: зрительная информация и зрительное осознание
В исследованиях НКС, посвященных определенному содержанию сознания, зрительное осознание, безусловно, изучают чаще всего. Поэтому здесь мы рассмотрим некоторые из самых показательных экспериментов в сфере нейрональных коррелятов зрительного осознания.
Зрительная информация поступает от глаз в мозг через ядра таламуса (латеральные коленчатые тела) и заканчивает свой путь в первичной зрительной коре (V1), которую еще называют стриатной корой. Оттуда она поступает в сложную сеть областей коры мозга, которые специализируются на обработке различных аспектов зрительной информации. Два основных пути обработки называются вентральным зрительным трактом (он идет от области V1 к экстрастриатной затылочной коре, а затем к височной доле) и дорсальным зрительным трактом (он идет от области V1 к задней париетальной коре (см. рис. 5.3). Вентральный зрительный тракт содержит области, обрабатывающие такие зрительные характеристики, как цвет, форма и тип объекта (например, лицо, животное или место). Таким образом, вентральный тракт, как предполагается, собирает воедино представления о связанных зрительных характеристиках, позволяющие их распознавать и объединять в целостный объект. Дорсальный зрительный тракт связан с пространственным расположением и движением объектов. Считается, что он помогает отслеживать движение объектов в пространстве и управляет сенсомоторным взаимодействием с объектами.
Возникает вопрос: в каких областях зрительной зоны коры мозга расположены нейрональные корреляты осознаваемого зрительного восприятия? Что происходит в мозге, когда мы видим объект или его элемент или когда зрительная информация входит в наше сознание в последовательной феноменальной форме, в виде красивых, ярких зрительных квалиа, например хроматических цветов? Чтобы исследовать этот вопрос, нужно разработать эксперименты, где в одних условиях зрительная информация не входит в сознание – субъективно мы не видим стимулов, а в других условиях та же самая зрительная информация входит в сознание, и у нас возникает субъективное зрительное восприятие стимула.
Исследование бинокулярной конкуренции
Для экспериментов, позволяющих отделить нейрональные корреляты субъективного зрительного переживания от активности, непосредственно вызванной зрительными стимулами, один подход оказался особенно плодотворным. Это исследования феномена, который называется бинокулярной конкуренцией. Ее можно вызвать, одновременно показывая два разных, зрительно несовместимых стимула, один для левого глаза, а другой – для правого (рис. 9.1). Обычно мозг объединяет информацию от обоих глаз в одно целостное трехмерное изображение, но в этом случае такая интеграция невозможна, потому что два представления не соответствуют друг другу. Скажем, левому глазу мы показываем изображение кролика, а правому глазу – изображение цветка.
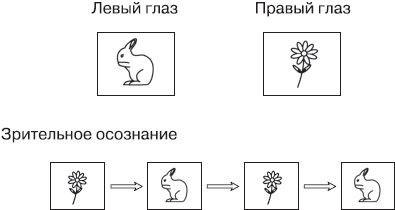
Рис. 9.1. Бинокулярная конкуренция
Левый глаз все время «видит» только кролика, а правый глаз все время «видит» только цветок. Но в зрительном осознании цветок и кролик возникают последовательно, друг за другом, и содержание сознания спонтанно меняется каждые несколько секундСначала мозг пытается объединить оба образа, но когда это не удается, изображения начинают конкурировать друг с другом за доступ в сознание. Таким образом, сначала, в течение нескольких секунд, испытуемый видит цветок и больше ничего, а затем цветок внезапно превращается в кролика. В течение нескольких следующих секунд испытуемый видит кролика, а потом образы снова переключаются.
Спонтанная конкуренция между двумя глазами и двумя стимулами может продолжаться довольно долго. Этот феномен дает идеальную возможность исследовать, что происходит в мозге, когда информация оказывается в сознании. Во время эксперимента оба стимула постоянно находятся перед глазами испытуемого – они остаются неизменными. Поэтому испытуемого просят сообщить, когда у него в сознании происходит переключение образов. Например, он может нажимать на кнопку.
В экспериментах с бинокулярной конкуренцией участвовали и обезьяны, и люди (хотя, как можно предположить, было не так просто научить обезьян сообщать , что они видят!). В экспериментах с участием обезьян активность единственной клетки в зрительной коре мозга регистрировалась с помощью вживленных микроэлектродов: это тонкие иглы, введенные в зрительную кору головного мозга, и острие иглы регистрирует электрическую активность нейронов, расположенных рядом с ней.
Таким образом удавалось точно определить, в каких участках мозга возникает активность, но сигнал отражал только реакцию одного или нескольких нейронов на изменения в субъективном переживании. Итак, исследователи регистрировали реакции клеток, «интересовавшихся» стимулами, которые видела обезьяна, – эти клетки реагировали на наличие или отсутствие физического стимула в поле зрения при отсутствии конкуренции. Некоторые клетки активировались, когда обезьяна видела изображение лица, а другие – когда она видела изображения каких-то других объектов, при отсутствии других стимулов.
Когда два изображения показывали одновременно, в ситуации конкуренции между ними, результаты показали, что в первичной зрительной коре (V1) лишь для немногих из клеток, активность которых регистрировалась (около 20%), эта активность коррелировала с изменениями в содержании сознания. Когда регистрировалась активность клеток других областей вдоль вентрального зрительного тракта, количество клеток, коррелирующих с изменениями сознания, увеличивалось вдоль всего вентрального тракта. Таким образом, в задней части височной до ли уже 90% клеток, активность которых регистрировалась, «интересовались» стимулами данного типа, при этом их активность коррелировала с изменениями в содержании сознания: они были максимально активны, когда предпочитаемый стимул появлялся в сознании, – в отличие от тех ситуаций, когда он просто находился перед глазами обезьяны и воспринимался мозгом, но не достигал сознания. Активность этих клеток снижалась, когда предпочитаемый стимул исчезал из сознания и в сознании появлялся конкурирующий стимул (Leopold & Logothetis, 1999).
После столь захватывающих экспериментов на обезьянах подобный эксперимент был проведен с участием людей, но на этот раз активность мозга регистрировалась с помощью фМРТ. Прежде всего, оказалось, что когда испытуемому показывают изображение лица, максимально активируется особая область вентрального тракта, «фузиформная лицевая область», а когда испытуемый видит изображение дома, активируется другая область, «парагиппокампальная область места». Когда и лицо и дом демонстрировали одновременно в условиях конкуренции, уровень активности этих областей коррелировал с содержанием сознания. Когда в сознании появлялся образ лица, активировалась область, отвечающая за восприятие лиц, а когда в сознании возникало изображение дома, активировалась область, отвечающая за восприятие места. Таким образом, возникает впечатление, что области, которые специализируются на обработке определенных типов объектов в мозге человека, влияют на зрительное осознание объектов того же типа (Kanwisher, 2001).
Зрительные галлюцинации
Независимые доказательства одного и того же явления были получены в ходе экспериментов, где наличие или отсутствие определенного содержания в зрительном осознании возникало спонтанно, вообще без каких бы то ни было стимулов. Неврологическое расстройство под названием «синдром Чарльза Боннета» характеризуется спонтанными зрительными галлюцинациями в виде объектов, лиц, людей или других зрительных иллюзий. Эти галлюцинации могут быть настолько яркими и реалистичными, что их легко перепутать с реальностью. Однако каждый больной обычно видит особые, но всегда одинаковые образы, и поэтому легко учится распознавать свои галлюцинации и не путать их с реальностью (Fychte и др., 1998).
Во время эксперимента проводилась запись активности мозга с помощью фМРТ. В контрольных условиях у пациента не было галлюцинаций, а в экспериментальных условиях содержание галлюцинаций явно присутствовало в зрительном осознании. Результаты показали, что наличие галлюцинации коррелирует с активностью определенных областей вдоль вентрального зрительного тракта.
В целом доказательства, полученные в ходе этих (и многих других) экспериментов. Указывают, что вентральный зрительный тракт – вместилище зрительного осознания, по крайней мере, здесь находятся нейрональные корреляты содержания. Если объединить эти данные с независимыми данными, полученными у больных с нейропсихологическими расстройствами, потерявшими способность видеть зрительные образы (мы рассматривали эти данные в главе 4), мы наблюдаем убедительное единство в результатах, которое подтверждает, что для зрительного осознания того или иного содержания необходима активность одних и тех же анатомических областей вдоль вентрального зрительного тракта.
Вентральный зрительный тракт – это сложная совокупность участков коры головного мозга. Поэтому можно сказать, что активность в какой-то ее части коррелирует со зрительным осознанием, не имея в виду совершенно точной локализации нейрональных механизмов сознания. До сих пор остается неясным, где и на каком участке вдоль вентрального тракта информация впервые всплывает в сознании. Почти все исследования указывают, что для зрительного осознания наиболее важны эволюционно более поздние области в височной доле. Но многие исследования также показывают, что активность в первичной зрительной коре (V1), куда сначала поступает зрительная информация извне, может отражать содержание зрительного осознания, а не физический стимул как таковой (Rees, 2007).
Роль области V1 в зрительном осознании до сих пор не выяснена. Есть данные, указывающие на ее участие, а также другие данные, указывающие на то, что она не является необходимой для зрительного осознания. Например, люди, у которых зона V1 оказалась повреждена вследствие травмы головного мозга и которые страдают кортикальной слепотой, все же могут представлять себе зрительные образы (Solms, 1997). Это доказывает, что зона V1 не является абсолютно необходимой для зрительного переживания. Даже если зона V1 так или иначе способствует зрительному осознанию, она делает это не одна. Вероятно, она взаимодействует с несколькими областями, расположенными дальше вдоль вентрального зрительного тракта, и зрительное содержание входит в сознание только после того, как пройдет весь этот сложный путь. (Эта идея подробнее обсуждается в разделе «Теория рекуррентной обработки» в главе 11).
Многие исследования с помощью томографии мозга показывают, что кроме вентрального зрительного тракта активируются другие области мозга, прежде всего в лобной и теменной коре. Пока не ясно, действительно эти области необходимы для зрительного осознания или они просто активируются при выполнении зрительных задач, поскольку такие задачи требуют участия когнитивных функций более высокого уровня: селективного нисходящего внимания и рабочей памяти. Лобные и теменные нейронные сети, в любом случае, важны для пространственного внимания и пространственного осознания, потому что повреждение этих областей связано с игнорированием стимулов в одной половине пространства (Rees, 2007).
Гипотеза о том, что лобные и теменные области не являются необходимыми для зрительного осознания, подтверждена экспериментами, показавшими, что когда селективное нисходящее внимание не является необходимым для выполнения зрительных задач, эти области не активируются, хотя информация о стимуле входит в зрительное осознание (Eriksson, Larsson, & Nyberg, 2008). Другие эксперименты показали, что активация лобных областей коррелирует не только со зрительным осознанием, но также и со слуховым осознанием (Eriksson, Larsson, Ahl strom, & Nyberg, 2007). Именно это мы обнаружим, если лобные области больше связаны со вниманием, независимым от модальностей, рефлексивным осознанием и рабочей памятью, а не с феноменальным осознанием определенной модальности.
Однако противоречия между результатами разных исследований и разными гипотезами относительно НКС и зрительного осознания отнюдь не разрешены. Некоторые из этих противоречий связаны с более глубокими теоретическими и философскими предположениями о природе сознания: существует ли такая вещь, как чистое феноменальное сознание? Отражается ли весь осознаваемый опыт в рефлексивном сознании? То есть зависит ли все осознаваемое зрительное содержание от селективного внимания, рабочей памяти и способности выражать его словами? Мы вернемся к этим вопросам, когда будем обсуждать другие теории сознания, где этот спор возникнет снова.ЭЭГ и МЭГ исследования зрительного осознания
В экспериментах с ЭЭГ и МЭГ можно отследить во времени, когда информация от зрительного стимула входит в сознание. Сигналы ЭЭГ и МЭГ показывают, с временным масштабом в 1/1000 секунды, как развиваются электромагнитные реакции мозга. Сравнивая реакции мозга на увиденные (осознаваемые) стимулы со сходными стимулами, которые не были увидены, мы должны заметить, что в какой-то момент мозг начинает по-разному реагировать на стимулы, которые входят в зрительное осознание, и на стимулы, которые не осознаются. Когда это происходит? Когда и где мозг позволяет зрительному стимулу войти в сознание?
Основная идея экспериментов с ЭЭГ и МЭГ в сфере НКС нам уже знакома: сравнить, что происходит при наличие осознания стимулов и при отсутствии их осознания. Чтобы одни и те же стимулы иногда были невидимыми, а иногда видимыми, их часто демонстрируют на грани субъективного порога восприятия. Это значит, что время предъявления стимулов так невелико или их так трудно отличить от фона, что обычно испытуемый видит их лишь иногда и не видит в других случаях. При такой схеме эксперимента можно сравнить реакцию на увиденные и пропущенные стимулы при прочих равных (по крайней мере, очень близких к равным) условиях.
В подобных экспериментах одному и тому же испытуемому стимул предъявляют десятки или даже сотни раз, потому что реакция ЭЭГ на отдельный стимул настолько мала относительно всей остальной постоянной активности ЭЭГ, что ее почти невозможно различить. Нормальная постоянная ЭЭГ всегда содержит всевозможные виды спонтанной электрофизиологической активности мозга, не связанной ни с какими внешними стимулами. Слабые реакции ЭЭГ, каждый раз вызванные стимулом, усредняются, чтобы сформировать среднюю кривую реакций. Эта кривая нейтрализует всю активность ЭЭГ, не имеющую особого отношения к стимулу. Усредненную кривую реакций на стимул называют связанным с событием потенциалом (ССП), потому что она показывает только те электрические события в мозге, которые определенно связаны с интересующим нас событием, – в данном случае со зрительным стимулом.
Электрические реакции, связанные с событием, демонстрируют первые надежные различия между увиденными (осознаваемыми) и не увиденными (неосознаваемыми) стимулами. Эти различия возникают примерно на 150–200-й миллисекунде после начала предъявления стимула и обычно достигают пика через 250–300 миллисекунд. В исследованиях ЭЭГ эту реакцию называют «негативностью зрительного осознания» (visual awareness negativity) (НЗО), потому что обычно на связанных с событием кривых потенциала мозга она выглядит как отрицательно направленная волна и возникает только при предъявлении стимулов, которые достигают сознания (Koivisto, Kainulainen. & Revonsuo, 2009; Wilenius & Revonsuo, 2007). Эта реакция сильнее всего обнаруживается в задней части головы, в местах, где электроды присоединены к затылочной, височной и задней теменной долям, – в областях скальпа, находящихся как раз над зрительной зоной коры головного мозга.
В исследовании МЭГ подобные реакции на увиденные объекты были локализованы в правой затылочной доле через 250–300 миллисекунд после начала предъявления стимула (Vanni, Revonsuo, Saarinen, & Hari, 1996). Нейрональная активность, которая выявляет НЗО, как полагают, является в мозге электрическим коррелятом феноменального зрительного осознания. Если эта гипотеза верна, мозгу нужно около 0,2–0,3 секунды на то, чтобы обработать информацию о стимуле в зрительной зоне коры головного мозга, – до того, как эта информация сможет войти в сознание.
Может показаться, что это очень быстро, но на самом деле для мозга это довольно медленная реакция. Зрительная информация входит в первичную зрительную кору уже в течение первых 20–30 миллисекунд и быстро обрабатывается во всех зрительных областях в несколько первых миллисекунд. Считается, что такая обработка происходит совершенно автоматически, за пределами сознания. Но при этом эта информация может управлять нашими быстрыми моторными реакциями на зрительные стимулы (в дело снова вступает лишенный сознания зрительно-моторный зомби!).
Если вы когда-либо рефлекторно отбивали теннисную подачу или бадминтонный воланчик, хотя даже не видели мяча или воланчика, по которому ударили, то ваша быстрая и успешная реакция, скорее всего, была результатом неосознаваемой быстрой обработки. Для осознания стимула, наоборот, нужен сложный синтез обратной связи между разными областями коры мозга, а это требует времени. Поэтому осознаваемое восприятие и основанные на нем действия обычно происходят гораздо медленнее, чем 0,1 секунды, которая требуется для первой быстрой реакции. (Подробнее о различиях между быстрой и медленной обработкой зрительных стимулов мы поговорим в обсуждении теории Ламме в главе 11).
За реакцией НЗО на ЭЭГ обычно следует другая волна, «поздняя позитивность» ( late positivity ) (ПП). Это медленная, широкая волна, возникающая примерно через 400–600 миллисекунд от начала действия стимула. Она очень похожа на другие позитивные волны типа P3 на ЭЭГ, которые, как полагают, отражают активность внимания и рабочей памяти. В экспериментах с осознанием поздняя позитивная волна считается коррелятом рефлексивного сознания: активности селективного внимания, классификации, называния и самоотчета об осознаваемом зрительном восприятии объекта (Koivislo, Kainulainen, & Revonsuo, 2009). Таким образом, исследования ЭЭГ также указывают на совокупность реакций, которые вполне можно интерпретировать как наличие феноменального сознания (на что указывает НЗО) и рефлексивного сознания (на что указывает ПП). Кроме того, поздняя позитивность не возникает, если стимул, достигающий феноменального сознания и генерирующий НЗО, не требует особого внимания, определенной реакции или сознательной рефлексии (Koivisto, Kainulainen, & Revonsuo, 2009).
Транскраниальная магнитная стимуляция (ТМС)
Недавно к арсеналу когнитивных неврологов добавился новый экспериментальный метод, дополняющий вышеупомянутые подходы. Метод транскраниальной магнитной стимуляции (ТМС) не похож на методы томографии мозга, ЭЭГ или МЭГ. Скорее, это способ локального вмешательства в нормальное функционирование мозга контролируемым образом. С помощью ТМС в определенные участки на поверхности коры мозга можно направить короткие магнитные импульсы. Они вызывают необычный взрыв нейроэлектрической активности, ненадолго прерывающий нормальную активность мозга. В экспериментах с ТМС можно наблюдать влияние таких вмешательств на поведение и сознание.
Когда с помощью этого устройства стимулируют зрительную зону коры мозга, можно наблюдать несколько эффектов. При низкой интенсивности магнитных импульсов (что создает лишь небольшой искусственный взрыв активности в зрительной зоне коры мозга), непосредственно стимулируя кору, например зрительные области V1 и V5, можно фактически создавать зрительное переживание . При стимулировании этих областей испытуемые сообщают о том, что видят мгновенные зрительные образы (их называют фосфенами ) – короткие вспышки света и зрительные образы. Фосфены бывают разной формы, иногда они имеют тот или иной цвет или движутся. По контрасту, если интенсивность импульсов ТМС высока, то подвергающаяся воздействию часть зрительной коры мозга, похоже, на время прекращает выполнять свои функции. Эта область становится временно слепой к зрительным стимулам (возникает «скотома», или слепая часть поля зрения), или, как минимум, в соответствующей части поля зрения снижается восприимчивость к стимулам.
Результаты исследований с использованием ТМС показывают, что когда возникают зрительные феномены, различные зрительные области взаимодействуют друг с другом. Например, когда возникают движущиеся фосфены, активизируются области V1 и V5. Если импульсы ТМС воздействуют на область V1 и в то же время на область V5, в попытке вызвать движущиеся фосфены, испытуемый не видит фосфенов. Таким образом, кажется, области V1 и V5 взаимодействуют друг с другом в том случае, когда возникают движущиеся зрительные феномены (Silvanto, Cowey, Lavie, & Walsh. 2005).
ТМС помогает установить, где расположены нейрональные механизмы зрительного осознания. Этот метод имеет преимущество перед другими, о которых мы говорили выше: с помощью ТМС можно выяснить не только то, какие области коррелируют с сознанием, но и то, какие из коррелирующих областей действительно важны и какого рода их активность необходима для зрительного осознания. Все остальные методы просто коррелятивны – они показывают, что осознание стимула и активность той или иной области имеют тенденцию возникать одновременно. Но такая корреляция не говорит о том, что именно делает эта область. С помощью ТМС можно помешать области выполнять свою задачу, а потом проверить, как отсутствие активности в этой области меняет сознание. Угнетая активность той или иной области и сравнивая полученные данные с данными ее нормальной работы, можно выяснить, по крайней мере косвенно, как нормальное функционирование этой области способствует осознанию. В этом смысле эксперименты с ТМС больше напоминают изучение кратковременных, небольших повреждений или временных нейропсихологических расстройств, чем создание функциональных карт при томографии мозга.
Выводы
В целом исследования с помощью функциональной томографии мозга и электромагнитного исследования мозга выявляют сходные данные относительно нейрональных коррелятов зрительного осознания. Когда зрительная информация включается в феноменальное сознание, активируются области коры мозга вдоль вентрального зрительного тракта. Эта активность быстро распространяется по всему пути, но чтобы зрительная информация достигла зрительного осознания, необходимо сложное взаимодействие, обмен обратной связью между разными областями коры мозга.
Поэтому чтобы информация включилась в сознание, нужно какое-то время – как минимум около двух десятых секунды, и это отражается в хронологии того, как возникают первые надежные электрические реакции на сознание. Сразу же после того как информация впервые входит в феноменальное сознание, она направляется в рефлексивное сознание, особенно если эта информация необходима для выполнения текущей задачи, достижения цели или принятия решения. Этап рефлексивного сознания коррелирует с активацией лобно-теменных областей мозга, которые, как известно, необходимы для внимания и рабочей памяти. Также известно, что с данными когнитивными механизмами коррелирует определенная волна связанных с событием потенциалов на ЭЭГ («поздняя позитивность»). Она указывает, что зрительная информация достигла рефлексивного сознания.
Такое представление о нейрональных коррелятах зрительного осознания поддерживают данные исследований больных с нейропсихологическими расстройствами, у которых отмечаются повреждения в вентральном зрительном тракте. Если вентральные области не только коррелируют со зрительным осознанием объектов, но и необходимы для его функционирования, то нарушения в этих областях должны приводить к нарушениям зрительного осознания, когда человек не видит объектов или каких-то их особенностей.
Вот что при этом происходит: повреждение в области V4 приводит к ахроматопсии или исчезновению из зрительного осознания феноменальных цветов, повреждение латеральной затылочной коры (lateral occipital cortex) угнетает способность осознанно воспринимать целостные зрительные объекты (хотя действия под влиянием неосознаваемой зрительной информации все еще возможны), и т. д. (Подробнее см. более ранний раздел, посвященный нейропсихологии сознания, главы 4–6). Похожие эффекты можно также временно создавать у нормальных здоровых испытуемых с помощью ТМС зрительной зоны коры мозга.
Исследования в области нейрональных коррелятов сознания продолжаются, и в ведущих научных изданиях по когнитивной нейробиологии почти ежедневно публикуются новые результаты. Так что экспериментальный подход к исследованию НКС все ближе и ближе подводит нас к пониманию нейрональных механизмов сознания. Помогут ли все эти эмпирические данные окончательно решить философские проблемы, связанные с отношениями между сознанием и мозгом, пока неясно.
До сих пор нет никаких революционных данных или теорий, которые позволили бы хотя бы начать устранять «объяснительный разрыв». Даже если мы выясним где, когда и какая нейрональная активность участвует в создании зрительного переживания, поймем ли мы, как нейрональная активность возникает или как она создает субъективные зрительные феномены?
Вопросы для обсуждения
1. Придумайте самый лучший эксперимент НКС. Попробуйте представить себе, какой эксперимент мог бы выявить нейрональные корреляты сознания наиболее явным образом. Какой была бы задача испытуемого? Какие методы измерения вы бы использовали? Как результаты эксперимента помогли бы устранить объяснительный разрыв?
2. Если феноменальное сознание и рефлексивное сознание возникают почти одновременно, когда человек видит стимул, возможно ли измерить отдельно НКС для феноменального сознания и НКС для рефлексивного сознания? Какой эксперимент нужно провести, если мы хотим изучить НКС только для феноменального сознания?
Назад: I. Нейропсихология сознания
Дальше: III. Теории сознания

