Происхождение фотосинтеза и древнейшие ископаемые следы жизни
Расхождение бактерий и архей
Последний всеобщий предок (LUCA) дал начало двум весьма разным группам микробов — бактериям и археям. Скорее всего, их различия связаны с тем, что они исходно приспосабливались к разным условиям обитания. Например, среди бактерий есть пять групп, способных к фотосинтезу, а среди архей таких нет. По-видимому, основной формой жизни бактерий в архейском и протерозойском периоде были цианобактериальные маты. Это слоистые колонии из многих видов микробов, общей толщиной до 1 см. Верхний слой мата состоит из цианобактерий, осуществляющих кислородный фотосинтез. Под ними находятся другие бактерии, которые ведут фотосинтез без выделения кислорода (например, серный) и используют свет с большей длиной волны, чем цианобактерии верхнего слоя. Также под цианобактериями находятся гетеротрофные бактерии, которые питаются отмершими клетками фотосинтезирующих соседей и выделяемой ими слизью. Они могут использовать кислород для дыхания. В самой глубине мата находятся микробы-бродильщики, которые разрушают органические вещества без участия кислорода. Хотя мат называется «цианобактериальным», он состоит из сотен видов разнообразных бактерий, цианобактерии доминируют в нем только по общей продуктивности.
Такой бактериальный мат на морском мелководье накапливает и упорядочивает осаждающиеся из воды минералы. В разных условиях это могут быть карбонаты, фосфаты или кремнезем. При этом образуются строматолиты («каменные ковры») — характерные слоистые минеральные отложения. В наше время строматолиты очень редки, и для ученых было большой удачей обнаружить их на мелководье залива Шарк (Австралия). Но в отложениях архейского и протерозойского периодов содержится множество строматолитов, и похоже, что в те эпохи цианобактериальные маты занимали большую часть морских мелководий и наземных водоемов.
Археи же предпочитают питаться неорганическими веществами, выходящими из глубин Земли, такими как водород и соединения серы. Они не используют энергию света и, за исключением одной группы метаногенов, не входят в состав цианобактериальных матов.
Различие в составе мембран бактерий и архей отражает разные исходные условия их жизни. Бактерии сменили терпеноспирты на жирные кислоты, так как двойные связи в молекулах терпенов уязвимы к ультрафиолетовому излучению Солнца. Липиды с двойными связями, в свою очередь, сохраняют прочность и текучесть мембраны в широком диапазоне температур, что важно для обитателей горячих источников. Выходы горячей воды в них могут неожиданно исчезать и появляться, поэтому их обитатели должны быть готовы в любой момент попасть из кипятка в холодную воду и обратно. Простая эфирная связь в архейных липидах более устойчива при высоких температурах, чем сложноэфирная. У бактерий, приспособившихся к высоким температурам, сложноэфирные связи в липидах заменяются на простые эфирные, т.е. вместо жирных кислот используются жирные спирты.
Скорее всего, предки бактерий и архей с самого начала разделились по направлениям приспособления. Одна группа, давшая начало бактериям, расселялась по поверхности суши и моря и совершенствовала механизмы использования энергии света. Другая группа «выбрала темную сторону», т.е. стала осваивать подземные местообитания. Ей пришлось научиться обходиться без света, и она дала начало археями.
Способы получения энергии у бактерий и архей
Из школьного учебника биологии можно узнать, что есть три основных способа получения энергии живыми организмами. Первый, основной для человека и животных, — аэробное дыхание, в котором сахара, жиры и другие вещества из пищи окисляются кислородом до воды и углекислого газа. Второй способ — брожение, которое превращает сахара в этиловый спирт (у дрожжей) или молочную кислоту (у животных). Брожение обходится без кислорода, но дает почти в 20 раз меньше энергии на 1 г сахара, чем аэробное дыхание. Кроме того, от молочной кислоты болят мышцы после нагрузки. Третий способ, фотосинтез, используют растения, и в конечном счете от созданных в нем сахаров зависят все, кто дышит кислородом или довольствуется брожением.
В мире прокариот (бактерий и архей) способы добычи энергии из внешней среды гораздо разнообразнее. Для начала: дыхание может быть не только кислородным. Например, на дне морей в слое ила живут бактерии, которые окисляют органические вещества сульфатом из морской воды. Этот процесс называется «сульфатное дыхание», или «сульфатредукция», и в нем сульфаты превращаются в сероводород. В почве множество бактерий используют в качестве окислителя соединения азота: нитраты и нитриты. Есть две разновидности нитратного дыхания: денитрификация и аммонификация. При денитрификации образуется свободный азот или оксид азота N2O («веселящий газ»), а при аммонификации бактерии превращают нитрат и нитрит в аммиак. Если аэробное дыхание позволяет из одной молекулы глюкозы получить 38 молекул АТФ, а брожение — только 2, то сульфатное дыхание дает 10–12 молекул АТФ на одну глюкозу, а нитратное — до 20, что гораздо лучше брожения.
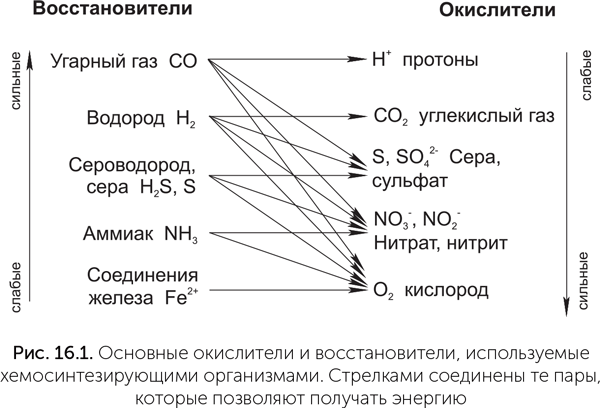
В дыхании прокариот окисляться могут не только органические вещества, созданные другими организмами, но и неорганические. Такой способ питания называется хемосинтезом. Бывает множество разновидностей хемосинтеза в зависимости от используемых окислителей и восстановителей (рис. 16.1). Например, водород, выходящий из глубин Земли и выделяемый бродильщиками, можно окислять сульфатом, при этом образуются вода и сероводород. Такой способ питания называется «гидрогенотрофная сульфатредукция», и распространен он у архей. Если нет сульфата, можно взять слабый, но вездесущий окислитель — углекислый газ. Соответствующие процессы называются «метаногенез» и «ацетогенез» по конечным продуктам: метан либо уксусная кислота. Элементарная сера в хемосинтезе уникальна тем, что может одновременно служить окислителем и восстановителем: этот процесс называется «диспропорционирование серы», его продуктами являются сероводород и сульфат. Наконец, при наличии сильных восстановителей можно использовать непривычный окислитель, который существует везде, где есть вода, — протоны (ионы водорода), восстанавливая их до газообразного водорода. Достаточно сильными восстановителями для этого процесса являются угарный газ и муравьиная кислота, а соответствующие способы питания — карбоксидотрофия и форматотрофия.
Глубинная биосфера
Одним из замечательных достижений микробиологии на рубеже веков стало открытие масштабной подземной биосферы. О том, что бактерии встречаются в земной коре на километровых глубинах, особенно в водах нефтяных месторождений, было известно и раньше, но только современные методы стерильного бурения, введение изотопных меток и анализа ДНК показали, что земная кора практически во всех районах заселена до глубины 3–5 км, а биомасса подземных микробов сравнима со всей биомассой суши, включая деревья.
Некоторые из подземных микроорганизмов, например обитатели нефтяных пластов, завершают разложение органики, созданной в процессе фотосинтеза и захороненной в толще земной коры. Другие могут строить все необходимые органические вещества из углекислого газа и получают энергию путем хемосинтеза: это сульфатредукторы, метаногены и карбоксидотрофы. Если они используют и восстановитель, и окислитель из недр Земли, то эти организмы могут быть полностью независимы от всей остальной биосферы. Одно такое сообщество, обнаруженное на глубине 3 км в Южной Африке, судя по составу воды, уже несколько миллионов лет существует в полной изоляции от жизни на поверхности планеты (http://elementy.ru/news/430367).
Глубинная биосфера, в отличие от жизни на поверхности планеты, почти неуязвима для различных катастроф. Падение астероида, глобальное оледенение, вспышка сверхновой в соседней звездной системе — обитатели глубин едва заметят эти катаклизмы. Даже если планета будет выброшена из своей системы в межзвездное пространство и ее поверхность остынет до температуры жидкого гелия, глубинная биосфера просто сдвинется глубже, ближе к теплому ядру планеты. Если на Марсе когда-то была жизнь, достигшая бактериального уровня сложности, то под поверхностью планеты она, скорее всего, сохранилась и может быть найдена при глубоком бурении. Есть лишь два способа уничтожить глубинную биосферу. Первый нам подсказывает Венера: неограниченный парниковый эффект прогреет поверхность и кору планеты до температур выше 200 ºC и надежно стерилизует ее. Второй способ — падение на планету крупного тела, сопоставимого с Луной. Выделение энергии при таком столкновении тоже прогреет верхние 10 км планетарной коры до нескольких сотен градусов, чего не переживут даже самые гипертермофильные микробы.
Поздняя метеоритная бомбардировка
3,9 млрд лет назад на планеты Солнечной системы обрушилось большое количество астероидов. В это время образовалось большинство кратеров Луны, Марса и Меркурия. Размеры этих кратеров указывают, что среди попавших на планеты небесных тел многие достигали размера 50 км, а самые крупные могли иметь размер до 200 км. Считалось, что удары крупнейших астероидов должны были приводить к испарению океанов и стерилизации поверхности Земли; соответственно, для зарождения жизни оставался короткий промежуток времени между окончанием бомбардировки и появлением известных следов жизни в гренландской формации Исуа возрастом 3,8 млрд лет. Но постепенно накапливались данные в пользу того, что жизнь возникла до метеоритной бомбардировки и как-то пережила ее. Были обнаружены включения углерода со смещенным изотопным соотношением в кристаллах циркона возрастом 4,25 млрд лет, а уточненные оценки с помощью молекулярных часов указывали на расхождение линий бактерий и архей, которое составляет более 4 млрд лет. Наконец, аккуратное моделирование метеоритной бомбардировки Земли и ее влияния на климат (Abramov, 2009) показало, что удары астероидов диаметром до 300 км стерилизуют только одно полушарие планеты, в то время как в другом полушарии температура не поднимается выше 70–80 ºC, и термофильные организмы выживают. Глубинная биосфера страдает еще меньше.
Хотя полной стерилизации поверхности Земли при поздней метеоритной бомбардировке не произошло, организмы, предпочитающие умеренные температуры до 50 ºC (они называются мезофилы, в отличие от термофилов, живущих при 50–80 ºC, и гипертермофилов, способных жить в кипятке), вряд ли выжили. После окончания бомбардировки холодные места обитания были вновь заселены потомками термофилов.
В нескольких очень интересных исследованиях (например, Akanuma et al., 2013) были сделаны попытки реконструировать последовательности отдельных ферментов LUCA. В этих работах последовательность предковых белков была вычислена из последовательностей множества их современных потомков как у бактерий, так и у архей; затем были синтезированы гены, кодирующие предковый фермент, встроены в клетки кишечной палочки и использованы для наработки и измерения свойств ферментов LUCA. Всякий раз оказывалось, что оптимальная температура для работы предковых ферментов — примерно 65–80 ºC. На родословных деревьях как бактерий, так и архей самые первые ветви состоят из термофилов. Это может быть как отражением термофильности LUCA, так и избирательным выживанием термофилов в эпоху поздней метеоритной бомбардировки. Если она привела к вымиранию всех микробов, кроме термофилов, то в этом случае методы реконструкции предковых последовательностей могут ошибочно приписать термофильность и LUCA.
Удары астероидов выбивают из планет осколки, способные преодолеть притяжение и выйти на околосолнечные орбиты. Более сотни марсианских метеоритов такого происхождения попали в руки ученых. Моделирование судьбы обломков, выбитых из Земли во время поздней метеоритной бомбардировки, показало, что они могут быть дополнительным убежищем для спор микробов. До 30% выбитых обломков возвращается на Землю в течение 5000 лет после столкновения, и содержащиеся в них споры могут вновь заселить остывшую после удара поверхность планеты (Wells, 2003). Более того, эти обломки имеют шанс перенести жизнь на другие тела Солнечной системы: 0,2% обломков попадают с Земли на Марс в течение 5 млн лет после столкновения, а самые первые достигают Марса уже через 100 000–150 000 лет. Обратный перенос с Марса на Землю требует больше времени — первые обломки совершают этот путь примерно за 300 000 лет, но зато их количество оказывается в 10 раз больше, чем перенесенных с Земли на Марс. Некоторые из обломков имеют шанс попасть даже на спутники Юпитера после 4 млн лет космического путешествия. Так что вполне возможно, что в ходе глубокого бурения на Марсе мы обнаружим жизнь общего происхождения с нашими земными организмами.
Устройство фотосинтеза современных растений и цианобактерий
Разнообразные реакции хемосинтеза, упомянутые выше, составляют лишь малую долю в энергетическом балансе биосферы. Подавляющее большинство живых организмов на планете зависит от энергии Солнца напрямую, как растения, или косвенно, как животные, получающие от растений пищу и кислород. Большинство бактерий тоже прямо или косвенно зависит от органики растительного происхождения либо от кислорода. Биосфера подключена к природному термоядерному реактору, Солнцу, благодаря фотосинтезу.
Процесс фотосинтеза, как он описан в школьных учебниках, состоит из световых и темновых реакций. Световые реакции сводятся к синтезу АТФ и восстановителей, таких как НАДФН, при помощи света и какого-либо донора электронов. Темновые реакции — это восстановление СО2 при помощи НАДФН и АТФ в цикле Кальвина или, реже, в других реакциях, и они уже обсуждались в главе о происхождении обмена веществ. Темновые реакции не уникальны для фотосинтеза. Тот же цикл Кальвина работает и при хемосинтезе, и свет не принимает участия в его реакциях.
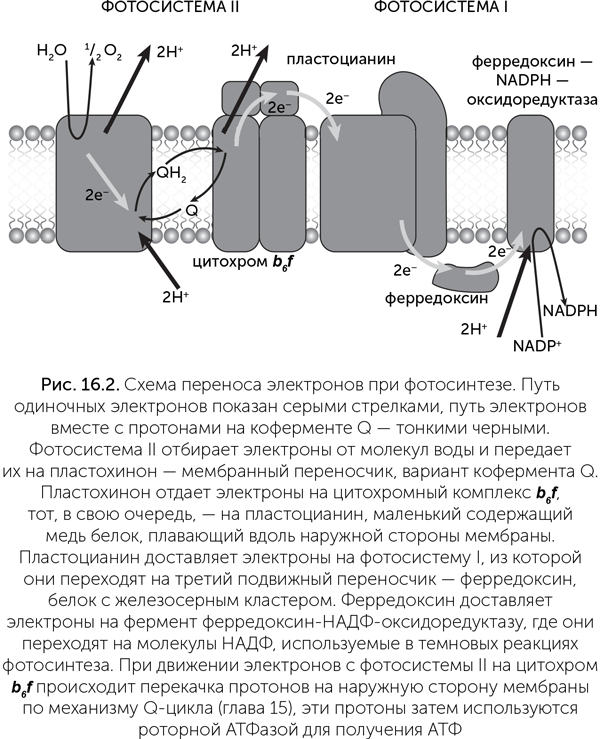
Напомним структуру фотосинтетического аппарата зеленых растений. Его функциональное ядро составляют реакционные центры — встроенные в мембрану белки, с которыми связаны молекулы хлорофилла — по две в каждом реакционном центре и несколько вспомогательных, передающих энергию света в центр. Возбужденные молекулы хлорофилла передают электроны на молекулы-переносчики, и затем они попадают в электрон-транспортную цепь. В отличие от дыхательной цепи, в которой электрический ток создается «химической батареей», полюса которой — НАДН и кислород, здесь электрический ток создают реакционные центры фотосистем, работающие подобно солнечным батареям. Они питают две нагрузки: во-первых, за счет энергии света происходит электролиз воды с выделением кислорода. Во-вторых, как и в дыхательной цепи, ток электронов через механизм Q-цикла преобразуется в ток протонов через мембрану и синтез АТФ на роторной АТФазе. Напряжения, создаваемого одной фотосистемой, недостаточно для электролиза воды, поэтому две фотосистемы соединяются последовательно. Как мы помним, модули электрон-транспортных цепей соединяются подвижными переносчиками электронов, поэтому для последовательного соединения нужны разные типы переносчиков и несовместимые «разъемы» для них.
Реакционные центры делятся на два типа по используемым переносчикам электронов: фотосистемы I типа (ФСI) передают электроны на ферредоксин — маленький водорастворимый белок с железосерным кластером, а фотосистемы II типа (ФСII) — на хиноны, плавающие в мембране. Цианобактерии и хлоропласты растений обладают обоими типами фотосистем, а различные фотосинтезирующие бактерии — только одним типом из двух. Электроны ФСI через ферредоксин переносятся на НАДФ и затем используются в реакциях восстановления разных органических веществ. Электроны ФСII проходят через хиноны на цитохромный комплекс b6 f, отдавая часть энергии на протонный ток и синтез АТФ, и далее на ФСI. В цианобактериях и хлоропластах растений окисленная ФСII с помощью водоокисляющего комплекса восполняет недостающие электроны из молекул воды, выделяя кислород. Кроме того, если клетке требуется больше АТФ, возможен циклический транспорт электронов по маршруту ФСII — хинон — цитохром b6 f — ФСII (рис. 16.2). Реакционные центры окружены большим количеством вспомогательных антенных белков, также связывающих хлорофилл, которые поглощают световые кванты и передают возбуждение на реакционные центры.
Разнообразие систем, запасающих энергию света
Очевидно, что столь сложная система из хлорофиллов и других пигментов и электрон-транспортных цепей не могла возникнуть сразу. У нее должны были быть более простые предшественники, а возможно, и альтернативные системы фотосинтеза, не использующие хлорофилл. Как мы помним из предыдущих глав, в состав живых систем с самого начала входили молекулы с особыми отношениями со светом. Например, все азотистые основания эффективно рассеивают энергию ультрафиолетового света в тепло, защищая соседние молекулы. Адениновые нуклеотиды в растворе также способны за счет энергии света присоединять фосфатные группы, синтезируя АТФ. Древние окислительно-восстановительные коферменты, такие как НАД и ФАД, тоже могли пройти отбор на особые фотохимические свойства. Как мы помним из главы про мир РНК-коэнзимов, ФАД (флавинадениндинуклеотид) и в современном мире проводит одну реакцию при помощи энергии света. Это реакция восстановления тиминовых димеров — одного из продуктов повреждения ДНК ультрафиолетом. В ходе экспериментов было обнаружено, что силикатно-протеиноидные микросферы, содержащие флавин, могут за счет энергии света синтезировать АТФ из АДФ и фосфата (рис. 16.3).
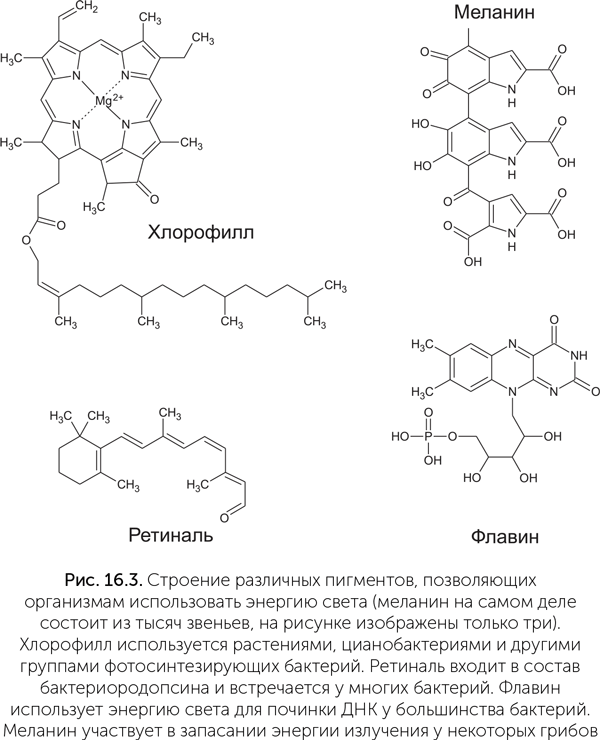
У животных и растений на основе бактериальной фотолиазы возникли криптохромы — ФАД-содержащие светочувствительные белки, регулирующие суточные и сезонные ритмы. Как ни удивительно, у животных криптохромы также участвуют в восприятии магнитного поля при помощи магниточувствительной реакции с анион-радикалом кислорода (Solovyov, Schulten, 2009).
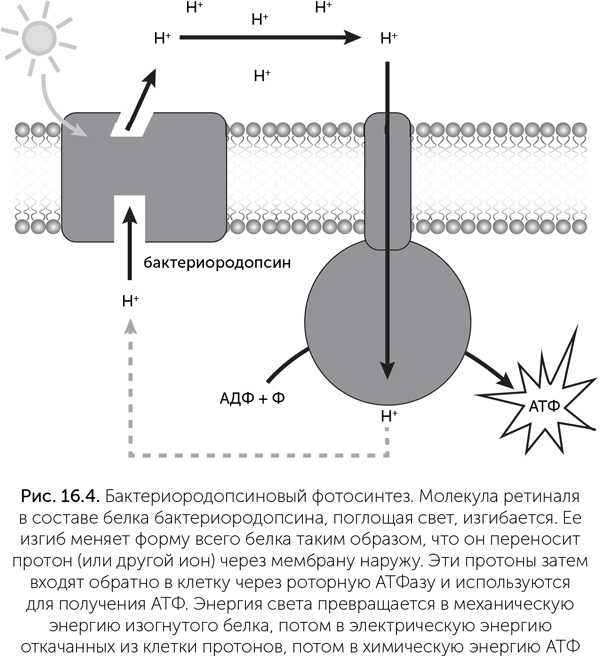
Другая группа светопоглощающих пигментов живых клеток — каротиноиды — тоже имеет разнообразные интересные функции. Один из каротиноидов, ретиналь, поглощает синий и голубой свет и превращает их энергию в механические изгибы молекулы, меняющей форму с прямой на изогнутую и обратно (рис. 16.4). В комплексе с мембранными белками бактериородопсинами ретиналь может переносить протоны, ионы натрия и даже ионы хлора через мембрану. Пропуская эти ионы обратно через вращающуюся мембранную АТФазу, клетка может получать АТФ за счет энергии света при помощи лишь одного дополнительного белка. Такая система гораздо надежнее и устойчивее к экстремальным условиям, чем сложные хлорофиллсодержащие фотосистемы. Бактериородопсины были впервые найдены у Halobacterium halobium, населяющей пересоленные лагуны, но в последнее время обнаружено, что они широко распространены у самых разных морских и пресноводных бактерий и позволяют им подолгу активно плавать без пищи. Родственные ретинальсодержащие белки — родопсины — работают в органах зрения всех животных, и благодаря им вы можете читать этот текст. Однако у одной глубоководной рыбы в глазах обнаружен и хлорофилл, позволяющий ей видеть в ближнем инфракрасном диапазоне! (Douglas et al., 1998).
Каротиноиды относятся к терпеновым веществам, гидрофобны и часто находятся в мембранах среди липидов. Самые распространенные каротиноиды, такие как бета-каротин, лежат в мембране параллельно хвостам липидов, проходя от одной стороны мембраны до другой. Скорее всего, исходно они защищали двойные связи древних липидов от повреждения ультрафиолетом, снимая с них возбуждение и рассеивая его в тепло, на самых ранних этапах эволюции мембран.
Третья группа веществ, поглощающих свет и, похоже, превращающих световую энергию в биохимическую, — это меланины, черные и желтые красители, определяющие цвет кожи и волос человека. Меланины состоят из нескольких тысяч одинаковых звеньев и в клетках обычно находятся в виде зерен. Кроме человека и животных меланины широко распространены у грибов и в разных группах бактерий. История открытия меланинового фотосинтеза начинается как фантастический роман: при очередном обследовании состояния саркофага Чернобыльской АЭС внутри него, на бетонных стенах, были найдены пятна совершенно черной плесени, растущей при очень высоком уровне радиации. Плесень была доставлена в лабораторию, где оказалось, что она относится к хорошо известному роду Cryptococcus, но, в отличие от известных родственников, накапливает в клеточных стенках огромное количество меланина. Дальнейшие эксперименты в медицинском колледже Альберта Эйнштейна в Нью-Йорке показали, что повышенный уровень радиации (в 500 раз выше естественного фона) ускоряет рост этой плесени в полтора раза на богатой среде и в три раза — при недостатке питательных веществ (Dadachova et al., 2007). Под действием ультрафиолета, рентгеновских лучей и гамма-лучей меланиновые гранулы восстанавливают НАД, что, видимо, используется в клетках плесени для запасания энергии в виде АТФ (Turick et al., 2011). Подобные черные штаммы были найдены и для других видов плесневых грибков. Они встречаются в высокогорье, где много ультрафиолета, и в зараженных радиацией почвах и способны расти по направлению к источнику радиации. Остается только пожалеть, что в организме человека меланин не обладает такими же замечательными свойствами.
Разнообразие и происхождение хлорофиллов и хлорофилл-связывающих белков
Большая часть фотосинтеза на Земле происходит благодаря хлорофиллу. По своему устройству его молекулы очень похожи на гем, о котором шла речь в предыдущей главе, но вместо железа в центре молекулы содержится магний.
В реакционных центрах встречаются три типа пигментов: хлорофилл a (цианобактерии, водоросли и наземные растения), бактериохлорофилл a (фотосинтезирующие бактерии, кроме цианобактерий), бактериохлорофилл g (только у гелиобактерий), а также феофитин — безмагниевый вариант хлорофилла а (только в паре с обычным хлорофиллом а). Кроме того, в разных растениях, водорослях и бактериях встречаются хлорофиллы b и с и бактериохлорофиллы b, c, d и e. Все эти вещества участвуют либо в передаче энергии света на реакционный центр, либо в движении электронов с реакционного центра на переносчики.
Зеленый цвет хлорофилла обусловлен тем, что он поглощает свет в двух диапазонах — красном и сине-фиолетовом. Свободный хлорофилл в растворе обладает красной флюоресценцией. Можно проделать простой опыт: нарезанную свежую зелень залить небольшим количеством спирта и оставить на ночь в закрытой посуде. Получится ярко-зеленый спиртовой раствор хлорофилла, который будет светиться красным при освещении ультрафиолетовым фонариком. У промежуточных продуктов синтеза хлорофилла — хлорофиллидов, протохлорофиллидов, протопорфирина IX — эта флюоресценция выражена сильнее, а полоса поглощения сдвинута в ближнюю ультрафиолетовую область. Хлорофилл в составе белков реакционных центров никакой флюоресценцией не обладает: вместо этого энергия поглощенного света превращается в электрическую.
Механизмы превращения энергии света в электрическую энергию в принципе одинаковы как в фотосистемах живых клеток, так и в солнечных батареях, созданных человеком. Солнечные батареи состоят из двух тонких слоев полупроводника (например, кремния) с разными добавками, из-за которых электрические свойства слоев отличаются (p-тип и n-тип проводимости). Поглощенный квант света вызывает возбуждение электрона в одном слое, после чего он быстро попадает в другой слой и не может вернуться обратно, кроме как через внешнюю электрическую цепь. Этот процесс называется «разделение зарядов». В реакционных центрах зеленых клеток содержатся молекулы хлорофилла. Они, как и гем, обладают обширной плоской системой двойных связей, электроны которых легко могут перемещаться по всей молекуле. По электрической проводимости хлорофиллы и гемы подобны графену, также обладающему обширной плоской системой двойных связей. Два полупроводника с разными свойствами, между которыми происходит разделение зарядов в фотосистеме II, — это обычный хлорофилл а и феофитин (хлорофилл без атома магния в центре). В фотосистеме I разделение зарядов происходит между «специальной парой» двух тесно сближенных молекул хлорофилла а и третьей молекулой того же хлорофилла а, проводящие свойства которого изменены соседними аминокислотами.
Хлорофилл-связывающие белки обоих типов реакционных центров, так же как главные антенные белки CP43 и CP47 цианобактерий и хлоропластов и светособирающие комплексы зеленых несерных бактерий (LH), состоят из повторяющихся похожих трансмембранных доменов, к части из которых прикрепляются молекулы хлорофилла. К молекулам хлорофилла близко прилежат остатки ароматических аминокислот — тирозина, триптофана, гистидина, которые могут передавать возбуждение на хлорофилл. Сравнение последовательностей этих белков друг с другом показывает, что все они могли произойти от одного предкового мембранного белка, имевшего 11 одинаковых доменов, с молекулами хлорофилла либо его предшественников на каждом домене (Mix et al., 2005; Mulkidjanian, Junge, 1997). В свою очередь, этот белок мог получиться путем последовательных удвоений доменов малого хлорофилл-связывающего белка с одним трансмембранным доменом, подобно современным светозащитным белкам HLIP цианобактерий.
Исходной функцией хлорофилл-связывающего белка, как и у каротиноидов, возможно, была защита от ультрафиолета: такой белок мог эффективно снимать возбуждение с соседних молекул и рассеивать его частью в тепло, частью — в красную флюоресценцию. Такая защита могла быть востребована еще на стадии РНК-белкового мира: протоорганизмы того времени уже имели мембраны, удаленные от минеральной подложки и не защищенные от ультрафиолета сульфидом цинка. Плавающие расселительные стадии протоорганизмов, не связанные с минеральной подложкой, выигрывали от наличия такого белка еще больше. Удвоения доменов и увеличение количества молекул хлорофилла повышали эффективность рассеивания энергии. В современных реакционных центрах сайты связывания переносчиков электронов (хинонов и ферредоксина) появились с утратой некоторыми доменами способности к связыванию хлорофилла.
Варианты хлорофилльного фотосинтеза
Способностью к фотосинтезу с помощью хлорофиллов или родственных им бактериохлорофиллов обладают пять неродственных групп бактерий: цианобактерии, пурпурные серные бактерии, зеленые серные бактерии (Chlorobi), зеленые несерные бактерии (Chloroflexi) и гелиобактерии (Heliobacteria). Только цианобактерии окисляют воду, выделяют кислород и обладают двумя типами фотосистем в одной клетке. Остальные фотосинтезирующие бактерии не выделяют кислород, более того, их системы фотосинтеза не могут работать в присутствии кислорода, за исключением Chloroflexi.
Серные бактерии, как пурпурные, так и зеленые, для восстановления углекислого газа окисляют сероводород и серу, выделяя сульфаты. Также они могут использовать для фотосинтеза молекулярный водород (выделяя воду) и соли двухвалентного железа (осаждая магнетит, Fe2O3). Пурпурные серные бактерии имеют фотосистему второго типа, отдающую электроны на хинон, и фиксируют CO2 в цикле Кальвина. Они особенно многочисленны в сернистых источниках и в озерах, где вода разделена на верхний пресный и нижний соленый слои. В таких озерах отсутствует перемешивание, и нижний соленый слой насыщен сероводородом.
Chloroflexi тоже имеют фотосистему второго типа, подобно пурпурным бактериям, но используют 3-гидроксипропионатный цикл фиксации CO2 (Zarzycky et al., 2009). Их историческое название «зеленые несерные бактерии» неверно, так как они могут использовать для восстановления СО2 сероводород, серу и водород (Tang et al., 2011). Впрочем, самый изученный вид этой группы, Chloroflexus auranticus, в природных условиях обычно не фиксирует СО2, а потребляет органические кислоты, выделяемые соседями по сообществу, и получает за счет света только АТФ.
Зеленые серные бактерии (Chlorobi) имеют фотосистему первого типа, отдающую электроны на ферредоксин, и используют восстановительный цикл Кребса для фиксации СО2 (Buchanan, Arnon, 1990).
Heliobacteria используют фотосистему первого типа с бактериохлорофиллом g в реакционном центре. Они населяют заболоченные почвы и особенно многочисленны на заливных рисовых полях. Гелиобактерии не способны к автотрофному росту (на СО2 в качестве единственного источника углерода) и нуждаются в готовой органике в виде сахаров или органических кислот, хотя в их геноме закодированы все, кроме одного, ферменты восстановительного цикла Кребса (Tang, Blankenship, 2010). Энергию света они используют для получения АТФ и фиксации азота.
Разные группы фотосинтезирующих бактерий отличаются по предпочитаемой интенсивности света. Фотосистема, настроенная на улавливание слабого света, под ярким полуденным солнцем становится опасной для клетки, вызывая фотохимические повреждения белков и мембран. Многие цианобактерии способны управляться с ярким светом и имеют для этого специальные приспособления: протеинкиназы для быстрой регулировки фотосистем (в течение секунд-минут) путем фосфорилирования белков и маленькие светозащитные белки HLIP (high light induced protein), синтез которых запускается ярким светом. Часть пурпурных бактерий тоже имеют подобные приспособления и переносят яркий свет. Другие фотосинтезирующие бактерии предпочитают укрываться от яркого света под слоем воды или под клетками цианобактерий в бактериальном мате. У Chlorobi и Chloroflexi есть приспособления к очень слабому свету: хлоросомы, зерна из очень плотно упакованных молекул хлорофилла. Благодаря им Chlorobi живут, например, в сероводородном слое Черного моря на глубинах до 100 м.
Эволюция хлорофилльного фотосинтеза
Хлорофилльный фотосинтез используют пять групп бактерий, не родственных между собой. Очевидно, что в распространении генов фотосинтеза большую роль сыграл горизонтальный перенос генов. Это подтверждается и данными по вирусам, заражающим цианобактерий: в их геномах часто встречаются гены компонентов фотосистем. Вирусы могут использовать эти гены для перестройки фотосинтеза зараженной клетки, чтобы быстро получить много энергии для размножения вируса ценой гибели клетки чуть позже. Гены, кодирующие систему фотосинтеза, часто образуют в геноме бактерий компактную группу (супероперон), что должно повышать вероятность переноса всего комплекта.
Система фотосинтеза цианобактерий устроена гораздо сложнее, чем у других групп бактерий: только в ней есть два типа фотосистем, дополнительные вспомогательные пигменты (фикобилины) и белки, связывающие их в компактные гранулы (фикобилисомы), а также защитные белки HLIP и протеинкиназы для регуляции фотосистем (рис. 16.5). Если сравнить набор генов, кодирующих детали системы фотосинтеза, у разных групп фотосинтетических бактерий, то получается, что и пурпурные серобактерии, и Chlorobi, и Chloroflexi, и гелиобактерии имеют больше общих генов с цианобактериями, чем друг с другом. Иными словами, горизонтальный перенос генов фотосинтеза происходил в основном либо от цианобактерий, либо к цианобактериям, но не между четырьмя остальными группами (Mulkidjanian et al., 2006). Лишь хлоросомы, служащие для приспособления к очень слабому свету, являются общим признаком Chlorobi и Chloroflexi и, видимо, были перенесены от одной из этих групп бактерий к другой.
Наиболее вероятно, что предки цианобактерий первыми освоили хлорофилльный фотосинтез, и у них же позднее появились две фотосистемы. Другие группы фотосинтезирующих бактерий приобрели способность к фотосинтезу благодаря событиям горизонтального переноса ряда генов от цианобактерий. Скорее всего, эти переносы произошли до того, как цианобактерии освоили кислородный фотосинтез. Фотосистемы Chlorobi и особенно гелиобактерий сохраняют некоторые очень примитивные черты (подробнее об этом — в главе 17).
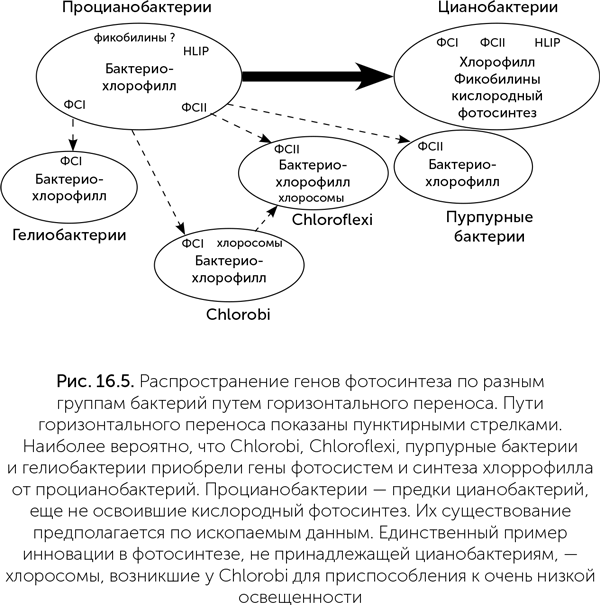
Геологические следы фотосинтеза и продуктивность древних экосистем
Первые следы фотосинтеза являются одновременно с первыми осадочными породами на Земле, в гренландской формации Исуа возрастом 3,8 млрд лет. Это джеспилиты, или полосатые железные руды (banded iron formations, BIF), а также включения графита со смещенным соотношением изотопов. Полосатые железные руды состоят из тонких (в доли миллиметра) темных слоев магнетита и более светлых слоев кремнезема или карбонатов. Считается, что они отлагались в морях в результате деятельности бактерий, осуществлявших фотосинтез с окислением железа. Магнетит откладывался днем, а карбонатные или кремнеземные слои — ночью. Первые ископаемые, очень похожие на современные нитчатые цианобактерии, появились в осадках Бак Риф (Южная Африка) 3,4 млрд лет назад. Все данные геохимии, однако, свидетельствуют о том, что появление кислорода в атмосфере произошло на 1 млрд лет позже. Изотопное соотношение углерода в осадках Бак Риф указывает на его биологическое происхождение, причем именно через цикл Кальвина. Другие пути фиксации СО2, такие как ацетил-КоА-путь, более чувствительны к различию в массе изотопов и поэтому сильнее обедняют биологический углерод тяжелым изотопом 13С. Отсутствие серы, сульфатов и магнетита говорит о том, что ни железо, ни сера и сероводород не использовались этими ископаемыми бактериями. Иначе говоря, методом исключения выявлено, что эти древние «процианобактерии» использовали в фотосинтезе водород. Их родство с современными цианобактериями подтверждается и положением в сообществе: они жили в самом верхнем слое бактериального мата на мелководье, а следовательно, были приспособлены к яркому свету.
Геохимики подсчитали количество водорода, железа и соединений серы, выделяемых из глубин Земли в архейском периоде (3,9–2,5 млрд лет назад). Исходя из этих оценок удалось сравнить продуктивность разных видов бескислородного фотосинтеза. По оценкам Canfield (2006), получается, что в глобальном масштабе вклад железоокисляющего фотосинтеза достигал 90%. Железоокисляющие фотосинтезирующие бактерии населяли поверхность открытых океанов. Их общая продуктивность могла быть примерно в 10 раз ниже продуктивности современных морских экосистем. Вклад водородного фотосинтеза оценивается примерно в 10%, а серного — не более 1%. Однако выход сероводорода из вулканических источников происходит локально, и фотосинтезирующие серобактерии могли образовывать высокопродуктивные «оазисы» в окрестностях вулканов.
Происхождение кислородного фотосинтеза
В течение архейского периода химический состав земной коры и верхней мантии постепенно изменялся: железо погружалось к ядру планеты, вулканическая активность слабела, и продуктивность всех типов фотосинтеза, зависящая от поступления водорода, серы и железа из недр Земли, снижалась. В этих условиях естественный отбор поддержал появление кислородного фотосинтеза, который использует бесконечный источник электронов — воду.
Системы защиты от токсического действия кислорода, необходимые для кислородного фотосинтеза, могли возникнуть даже раньше, чем кислородный фотосинтез. Показано, что на поверхности пирита (FeS2) при освещении в воде образуется перекись водорода — Н2О2. Ионы Fe3+ — продукт железо-зависимого фотосинтеза — тоже окисляют воду до перекиси водорода при освещении раствора, так что каталаза и другие защитные системы могли возникнуть вскоре после выхода бактерий из обогащенных цинком геотермальных водоемов в моря. Более того, показано, что водоокисляющий комплекс современных цианобактерий может использовать H2O2 в качестве источника электронов, причем для этого требуется меньше энергии света, чем для окисления воды. Возможно, обезвреживание перекиси водорода было первой функцией примитивного водоокисляющего комплекса. Существуют противоречивые данные о роли H2O2 в современном кислородном фотосинтезе — в зависимости от изучаемого вида и условий перекись может как подавлять его, так и стимулировать. Некоторым видам цианобактерий для выделения кислорода необходимо, чтобы заметное его количество уже содержалось в среде. Как это объяснить, пока непонятно (Olson, Blankenship, 2004).
Окисление воды — крайне сложная задача. Окислительно-восстановительный потенциал (его сокращенно называют редокс-потенциалом) бактериохлорофилла а (0,55 V) достаточен для окисления водорода, двухвалентного железа, H2S и серы. Редокс-потенциал хлорофилла а из цианобактерий выше, около 0,82 V. Однако если окислять воду, отбирая электроны по одному, то первая реакция, превращающая воду в гидроксильный радикал и протон, требует огромного редокс-потенциала в 2,75 V, недоступного ни для каких фотосинтетических пигментов. Поэтому необходим промежуточный переносчик электронов, способный окислить сразу две молекулы воды — редокс-потенциал хлорофилла будет достаточен, только если его равномерно делить на четыре электрона.
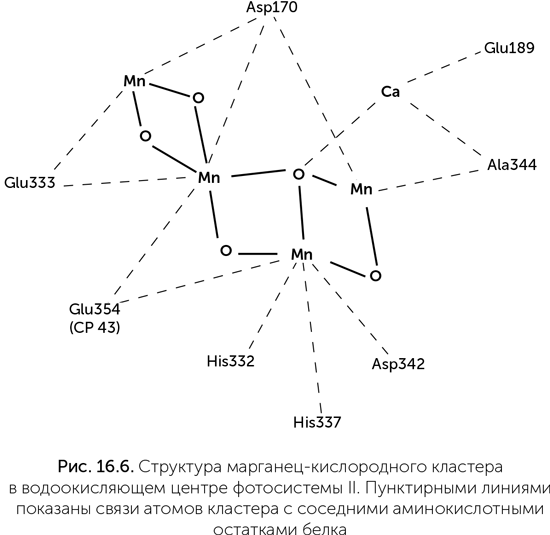
Водоокисляющий комплекс — это связанный с фотосистемой II марганец-кислородный кластер. Он содержит четыре иона марганца, меняющих степень окисления от +3 до +4, кроме того, с ним связано по одному иону кальция и хлора (рис. 16.6). Структура этого кластера напоминает элементарную ячейку таких природных минералов, как рансьеит (CaMn4O9 × 3H2O) и других слоистых оксидов марганца, подобно тому как FeS-кластеры окислительно-восстановительных ферментов похожи на ячейку пирита. Существовала гипотеза о происхождении водоокисляющего комплекса от защитного фермента супероксид-дисмутазы, одна из разновидностей которого содержит два иона марганца. Такой примитивный фермент, как предполагалось, мог быть способен к двухэлектронным реакциям — окислению H2O2 до кислорода. Окисление воды до H2O2 в такой системе было невозможно из-за недостаточного редокс-потенциала хлорофилла. К сожалению, никакого сходства между последовательностями супероксид-дисмутаз и белков фотосистемы II обнаружить не удалось.
Однако было давно замечено, что выделение кислорода в фотосинтезе сильно стимулируется добавлением бикарбонатов (солей HCO3-). Добавление изотопно меченого тяжелым кислородом 18O бикарбоната показало, что при освещении в первые секунды выделяется кислород, содержащий только тяжелый изотоп 18О. Это значит, что водоокисляющий центр переключается на окисление бикарбоната, при этом выделяются кислород и углекислый газ. Кроме того, было обнаружено, что бикарбонат участвует в сборке марганцевого кластера. Водорастворимая форма марганца — ионы Mn2+. В физиологических условиях они находятся в клетке в основном в виде марганец-бикарбонатных комплексов, таких как Mn2 (HCO3) 22+. В процессе сборки марганцевого кластера эти комплексы присоединяются к фотосистеме II. А затем под действием света Mn2+ в составе комплексов окисляются ею до Mn3+, а бикарбонат — до кислорода и CO2.
Марганец-бикарбонатные кластеры по своему редокс-потенциалу могут служить донорами электронов для бактериальных фотосистем, при этом происходит окисление Mn2+ до Mn3+. Это было обнаружено в экспериментах с фотосистемой II пурпурных бактерий, в норме окисляющих железо (Khorobrykh et al, 2007). Из этого можно построить следующую схему происхождения кислородного фотосинтеза (рис. 16.7): сначала, по мере исчерпания железа в океане, предки цианобактерий стали переходить на марганец-бикарбонатные комплексы в качестве источника электронов для фотосинтеза. Затем мутации ФСII привели к тому, что ионы Mn3+ (продукт марганец-окисляющего фотосинтеза) стали задерживаться на белке и принимать участие в его работе. Из них собрался примитивный марганец-кислородный кластер, способный проводить фотоокисление HCO3– до кислорода и CO2. Эта реакция требует в полтора раза меньше энергии, чем окисление воды, и возможностей бактериальной ФСII для этого практически достаточно. Так у фотосинтезирующих бактерий появился доступ к новому источнику электронов — бикарбонату. По мере того как запасы марганца в океане были израсходованы и осаждены в виде MnO2, процианобактерии стали переходить на окисление того, что осталось, т.е. бикарбоната. Марганец, который до того был расходным материалом для фотосинтеза, стал катализатором для использования бикарбонатов, и потребность в нем уменьшилась в тысячи раз.
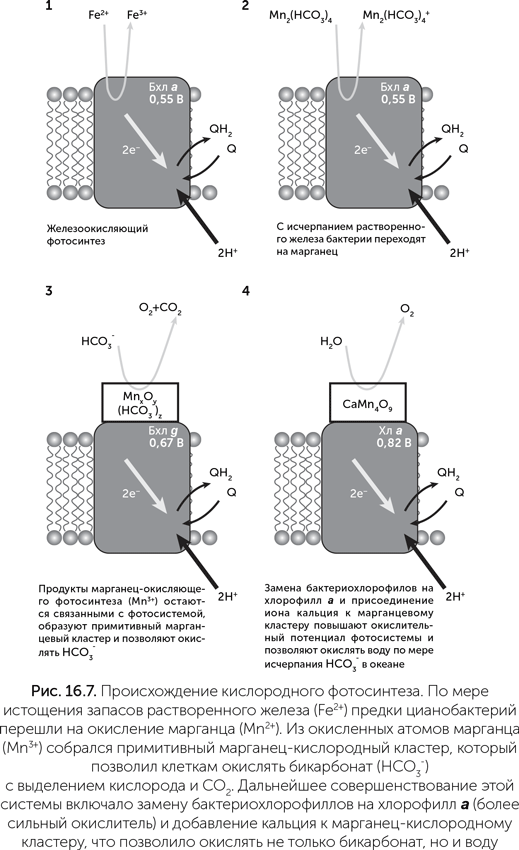
Далее бактериохлорофилл a в составе ФСII был заменен более редокс-активными пигментами, такими как бактериохлорофилл g, а затем и хлорофилл а, для повышения скорости бикарбонатного фотосинтеза. Одновременно совершенствовались системы защиты клеток от выделяемого кислорода. Наконец, по мере истощения запасов СО2 в атмосфере и бикарбонатов в море, к марганцевому кластеру присоединился ион кальция, повысивший его редокс-потенциал и сделавший возможным окисление воды. Так цианобактерии освоили неограниченный источник электронов для фотосинтеза и увеличили продуктивность всей биосферы в несколько раз (Dismukes et al., 2001).
Этот сценарий подтверждается новыми геологическими находками (Johnson et al., 2013). В Южной Африке были найдены морские осадки возраста 2,415 млрд лет, которые сильно обогащены оксидами марганца. Этот возраст примерно совпадает с началом оксигенизации атмосферы и гидросферы планеты. Там же присутствуют зерна легко окисляемого минерала пирита. Следовательно, в этих слоях осаждение марганца происходило не за счет реакций с кислородом. По-видимому, это следы деятельности марганец-окисляющих фотосинтезирующих организмов.
Итак, мы видим, что в течение 1,5 млрд лет после возникновения жизнь постепенно осваивала использование энергии Солнца и новые, все более распространенные восстановители для питания углекислым газом. С появлением кислородного фотосинтеза 2,4 млрд лет назад недостаток восстановителей (железа, водорода, соединений серы) более не ограничивал продуктивность экосистем. Появление в атмосфере кислорода, ядовитого для многих древних групп микробов, вызвало масштабное вымирание и перестройку экосистем. По-видимому, именно кислородная среда стала толчком к появлению новых организмов с более сложными клетками — эукариот, т.е. клеток с ядром. Только на основе эукариотной клетки стало возможно появление крупных многоклеточных растений и животных. В последней, 18-й главе мы рассмотрим движущие силы и механизмы появления эукариот.

