Книга: Лестница жизни: десять величайших изобретений эволюции
Назад: Глава 6. Движение
Дальше: Глава 8. Теплокровность
Глава 7. Зрение
Зрение — явление редкое. Глаза, по крайней мере, в общепринятом смысле, отсутствуют у растений, а также у грибов, водорослей и бактерий. Даже среди животных глазами обладают далеко не все. Царство животных делят на тридцать восемь типов, каждому из которых свойственна собственная модель плана строения, принципиально отличная от других, но лишь шесть из этих типов обзавелись настоящими глазами. Остальные продержались не одну сотню миллионов лет, вообще не пользуясь выгодами зрения. И естественный отбор не покарал их за слепоту.
Но на этом спартанском фоне эволюционные выгоды глаз все же смотрятся внушительно. Типы животных далеко не равны, и некоторые из них гораздо “равнее” других. Например, тип хордовые, к которому относимся и мы сами, а также все остальные позвоночные, включает более сорока тысяч видов. Тип моллюски, включающий слизняков, улиток и осьминогов, включает сто тысяч видов. А тип членистоногие, в который входят ракообразные, пауки и насекомые, включает более миллиона видов (8о % описанных к настоящему времени животных). При этом многие не столь известные типы, в том числе такие живые диковинки, как стеклянные губки, коловратки, черви-приапулиды и гребневики (в основном знакомые только профессиональным зоологам), включают сравнительно мало видов — десятки или сотни, — а тип пластинчатые — всего один. Если рассмотреть все глазастые и все безглазые виды вместе взятые, мы увидим, что у 95 % видов животных имеются глаза: те немногие типы, которые все-таки приобрели глаза, с большим отрывом доминируют в современном мире.
Разумеется, их преобладание могло бы быть простой случайностью. Можно предположить, что планы строения этих видов обладают и другими, не столь явными преимуществами, неучтенными нами и не имеющими никакого отношения к глазам. Но это представляется маловероятным. Судя по всему, появление и развитие настоящих глаз, не только способных отличать свет от тьмы, но и давших их обладателям пространственное зрение, действительно преобразило всю эволюцию жизни на Земле. Древнейшие настоящие глаза появляются в палеонтологической летописи довольно резко, около 540 миллионов лет назад, вскоре после начала “кембрийского взрыва”, этого Большого взрыва эволюции, когда численность и разнообразие ископаемых поразительно резко увеличиваются. В то время в породах, где прежде сотни миллионов лет не сохранялось почти никаких следов животных, чуть ли не в одночасье появляются представители почти всех современных типов.
Изобретение глаз почти наверняка не случайно совпало с взрывом разнообразия животных в палеонтологической летописи, потому что пространственное зрение должно было поставить отношения между хищниками и жертвами на совершенно другую основу. Оно одно вполне могло стать (и, возможно, стало) причиной пристрастия кембрийских животных к тяжелым “доспехам”, сильно увеличившим вероятность успешного захоронения. Биолог Эндрю Паркер из лондонского Музея естественной истории в своей занимательной, хотя местами досадно пристрастной книге правдоподобно обосновал версию, согласно которой именно появление настоящих глаз и вызвало “кембрийский взрыв”. Действительно ли глаза могли возникнуть в ходе эволюции так резко (или же здесь палеонтологическая летопись вводит нас в заблуждение), мы обсудим позже. А сейчас давайте просто отметим, что зрение дает намного больше информации об окружающем мире, чем в принципе могут дать обоняние, слух или осязание, потому что Земля залита светом и остаться незамеченным почти невозможно. Многие из самых замечательных адаптаций в живой природе представляют собой реакцию на возможность быть увиденным. Одни из таких адаптаций способствуют половому размножению, как перья павлина или лепестки цветка, другие выставляют напоказ внушительные латы, как выросты на спине стегозавра, третьи помогают прятаться, как форма тела и окраска палочников. Наше общество придает зрению настолько большое значение, что едва ли есть смысл доказывать здесь его важность.
Проблема эволюции зрения не только важна в связи с его практической пользой, но и имеет культовый статус, связанный с предполагаемым совершенством глаз. Уже во времена Дарвина глаза называли одной из вершин творения, ставящих под сомнение саму концепцию естественного отбора. Могли такой сложный, такой совершенный орган действительно развиться в ходе эволюции без чьей-либо помощи? Критики спрашивали, какая может быть польза от половины глаза. Естественный отбор предполагает, что развитие любой структуры включает тысячи и тысячи постепенных переходов, каждый из которых должен делать организм хоть немного лучше. В противном случае недостроенная структура будет безжалостно уничтожена. Но глаз, по словам таких критиков, совершенен как часы и точно так же неупрощаем. Стоит вынуть несколько деталей, и весь механизм перестанет работать. Часы без стрелок бесполезны, как бесполезен и глаз без хрусталика или сетчатки, — такое утверждение нам часто приходится слышать. А если половина глаза бесполезна, значит, глаз не мог развиться путем естественного отбора или каким-либо другим известным современной биологии способом и должен быть признан доказательством высшего замысла.
Те, кто приводит подобные аргументы, обычно защищают с их помощью давно укрепленные позиции. Сторонники Дарвина возражают на это, что глаз на самом деле далеко не совершенен, и это прекрасно известно всем, кто носит очки или контактные линзы либо вообще теряет зрение. Это действительно так, но это наблюдение уязвимо как теоретический аргумент, потому что здесь не учитываются многие тонкости. Возьмем, например, человеческий глаз. Распространенный аргумент гласит, что в его строении есть принципиальные недостатки, которые на самом деле нужно признать как раз доказательством того, что он кустарно сработан эволюцией из неподходящих, непродуманных структур и испорчен отсутствием у нее дара предвидения. Сторонники этого взгляда утверждают, что профессиональный инженер спроектировал бы глаз гораздо лучше и, более того, что глаз осьминога спроектирован гораздо лучше глаза человека. Но эти поверхностные рассуждения не учитывают одного неудобного правила, которое называют вторым законом Лесли Оргела: эволюция умнее нас.
Давайте коротко обсудим этот случай. Глаз осьминога очень похож на наш и тоже устроен по принципу фотоаппарата: одна линза объектива (хрусталик) впереди и одна светочувствительная поверхность (сетчатка), соответствующая фотопленке, сзади. Поскольку наш последний общий предок с осьминогами был, по-видимому, той или иной формой червя и не имел настоящих глаз, глаза осьминогов и наши собственные глаза должны были развиться независимо, и их сходство обусловлено тем, что эволюция нашла с их помощью похожие решения одной и той же задачи. Этот вывод подтверждают и результаты детального сравнения двух обсуждаемых типов глаз. Глаза человека и осьминога развиваются у зародышей из разных тканей и отличаются по микроскопическому строению. На первый взгляд глаз осьминога устроен гораздо разумнее. Светочувствительные клетки сетчатки смотрят в сторону света, а отходящие от них нервные провода ведут прямо в мозг. Наша же сетчатка, как нередко говорят, включена в сеть не с той стороны, и ее устройство кажется совершенно бредовым. Светочувствительные клетки в ней смотрят как раз в противоположную сторону и покрыты нервными проводами, вначале почему-то идущими в сторону света и только после, кружным путем, приводящими в мозг. Чтобы достичь светочувствительных клеток, свет должен вначале пробираться через лес этих проводов. Более того, все эти провода собраны в пучок, пронзающий сетчатку и образующий зрительный нерв, в результате чего на сетчатке имеется слепое пятно1.
Но не стоит спешить признавать превосходство глаз осьминогов над нашими. Как часто и бывает в биологии, на самом деле ситуация гораздо сложнее. Провода у нас в глазах бесцветны, поэтому не особенно препятствуют прохождению света, да и даже препятствуя ему, могут служить “волноводами”, направляя свет вертикально на светочувствительные клетки и тем самым оптимизируя использование попадающих в глаза фотонов. Кроме того (что, может, еще важнее), у нас есть то преимущество, что наши светочувствительные клетки непосредственно окружены “клетками поддержки” (пигментным эпителием сетчатки), к которым напрямую подходят кровеносные сосуды, обеспечивающие обильное кровоснабжение. Такое устройство позволяет поддерживать постоянный оборот светочувствительных пигментов. Человеческая сетчатка на единицу массы потребляет даже больше кислорода, чем головной мозг. Это самая энергоемкая структура во всем организме, так что подобная организация ее кровоснабжения необычайно ценна. Вероятнее всего, глаз осьминога не смог бы поддерживать столь же интенсивный обмен веществ в сетчатке. Но ему, наверное, это и не нужно. Живя под водой, куда попадает не так уж много света, осьминоги вполне обходятся без такого быстрого, как у нас, оборота светочувствительных пигментов.
Я хочу сказать, что любое биологическое устройство имеет свои достоинства и недостатки и возникает под действием уравновешивающих друг друга сил отбора, которые не всегда просто установить. В том-то и заключается проблема “очевидных” эволюционных объяснений: нам почти неизменно видна лишь половина картины. Аргументы, слишком концептуальные по своей природе, всегда уязвимы для контраргументов. Как и всякий ученый, я предпочитаю следовать фактам. Следует сказать, что бурное развитие молекулярной генетики за последние десятилетия привело к тому, что в нашем распоряжении оказалась масса фактов, дающих вполне конкретные ответы на конкретные вопросы. Из них складывается весьма убедительная картина эволюции глаза и его происхождения, как ни странно, от структур нашего очень далекого зеленого предка. В настоящей главе мы рассмотрим эту картину и увидим, чем полезна половина глаза, как возникли хрусталики и откуда взялись светочувствительные клетки сетчатки. А разобравшись во всем этом, мы убедимся, что изобретение глаза действительно изменило ход и скорость эволюции жизни.
К вопросу, какая может быть польза от половины глаза, легко отнестись насмешливо, уточнив, от какой половины: левой или правой. Я могу понять язвительный ответ Ричарда Докинза: от половины глаза может быть на 1% больше пользы, чем от 49 % глаза. Но если мы хотим ясно представить себе, что такое половина глаза, 49 % глаза нас только запутают. И все же половина глаза, понятая в буквальном смысле, дает нам очень хороший подход к интересующей нас проблеме. Глаз действительно отчетливо делится на две половины: переднюю и заднюю. Любому, кому доводилось бывать на конференциях офтальмологов, известно, что есть два больших клана: работающие с передними структурами глаза (делающие операции на хрусталике и роговице и лечащие бельмо и катаракту) и занимающиеся задними структурами глаза (сетчаткой), лечащие такие часто приводящие к слепоте расстройства, как макулодистрофия. Представители этих кланов взаимодействуют друг с другом неохотно и порой говорят чуть ли не на разных языках. Но это разделение вполне оправданно. Если убрать из глаза все оптические аксессуары, от него останется только голая сетчатка — ничем не покрытый светочувствительный слой. И именно эта голая сетчатка и послужила фундаментом для эволюции глаз.
Идея голой сетчатки может показаться странной, но она прекрасно вписывается в столь же странную среду — мир кислых гидротермальных источников — глубоководных “черных курильщиков”, с которыми мы познакомились в главе 1. Поля таких источников населены поразительными существами: все они так или иначе зависят от бактерий, живущих непосредственно за счет сероводорода, выделяемого источниками, Быть может, самые необычные из них (и определенно самые знаменитые) — гигантские черви рифтии, достигающие почти двух с половиной метров в длину. Рифтии — дальние родственники дождевых червей — особенно удивительны тем, что у них нет ни рта, ни кишечника, а питаются они за счет серных бактерий, которых выращивают внутри собственных тканей. Встречаются на подводных полях и другие гиганты, такие как двустворчатые моллюски.
Все эти гиганты обитают только в Тихом океане. Однако у “черных курильщиков” Атлантики есть свои чудеса, в частности креветки Rimicaris exoculata. Их научное название означает “безглазая рифтовая креветка”, о чем описавшие этот вид исследователи, должно быть, не раз пожалели. Разумеется, как и можно ожидать от существ с таким названием, живущих в беспросветных глубинах океана, эти креветки не имеют глаз в общепринятом смысле слова. У них нет глазных стебельков, как у их родичей, живущих ближе к поверхности, зато на спине есть два больших светлых участка. И хотя на первый взгляд в этих двух полосках нет ничего особенного, в темноте, освещаемые прожекторами подводных аппаратов, они светятся как кошачьи глаза.
Первой эти складки заметила Синди Ван Довер, и ее открытие положило начало одной из самых замечательных научных карьер нашего времени. Ван Довер относится к той породе исследователей, о представителях которой писал в свое время Жюль Верн и которая теперь стала не менее редким видом, чем любые из тех, что она изучает. В настоящее время Синди Ван Довер возглавляет Лабораторию моря в Университете Дьюка. Она посетила почти все известные гидротермальные источники, в том числе немало совершенно неисследованных, и была первой женщиной, управлявшей военно-морским глубоководным пилотируемым аппаратом “Алвин”. Впоследствии именно она открыла, что подобные гигантские моллюски и черви населяют и холодные участки морского дна, где из недр Земли сочится метан. Из этого следует, что движущей силой, обеспечивающей богатство придонной жизни в глубинах океана, служат химические условия, а не температура. Однако в конце 8о-х годов все это было еще впереди, и она, должно быть, чувствовала себя довольно неловко, когда посылала образцы спинных полосок слепой креветки специалисту по глазам беспозвоночных, сопроводив их глуповатым вопросом, не может ли это быть глазами. Но вскоре она получила лаконичный ответ: если изуродовать сетчатку, она может выглядеть примерно так. Хотя у слепых креветок нет обычных атрибутов глаз: роговицы, хрусталиков и так далее, у них обнаружилось что-то вроде голой сетчатки, образующей две полоски на спине — несмотря на то, что живут эти существа в беспросветных океанских глубинах.

Безглазая креветка Rimicaris exoculata. На ее спине видны две светлых полоски голой сетчатки.
Результаты дальнейших исследований оказались еще любопытнее, чем надеялась Синди Ван Довер: в голой сетчатке нашелся пигмент, очень похожий на тот, который улавливает свет в человеческой сетчатке — родопсин. Более того, этот пигмент был упакован в светочувствительные клетки, очень похожие на те, что имеются в глазах обычных креветок, хотя в целом сетчатка выглядела совсем по-другому. Это заставляло предположить, что якобы слепые креветки видят свет. Исследовательница задалась вопросом, не сами ли гидротермальные источники испускают его. В конце концов, нагретые металлические нити в электрических лампочках светятся, а гидротермальные источники не только нагреты, но и полны растворенных металлов.
Прежде на “Алвине” никогда не выключали прожекторов. Никому не приходило в голову это делать в непроглядной тьме океанских глубин, ведь без света аппарат вполне мог налететь на гидротермальный источник и погибнуть вместе с командой или, по крайней мере, повредить оборудование. Синди Ван Довер тогда еще не спускалась сама к “черным курильщикам”, но она смогла убедить геолога Джона Делейни, который собирался их исследовать, выключить свет и сфотографировать гидротермальный источник цифровой камерой. Хотя невооруженному глазу казалось, что за бортом беспросветная тьма, камера запечатлела окружающее источник отчетливое сияние, “висевшее в темноте, словно улыбка Чеширского Кота”. Тем не менее, эти первые фотографии ничего не позволяли сказать о том, какой именно свет испускали источники: какого цвета, какой яркости. Могли ли креветки “видеть” свечение источников, если даже люди совсем ничего не видели?
Предполагалось, что “черные курильщики”, как и нагретый металл, должны были испускать прежде всего красный свет, приближающийся к тепловой (инфракрасной) части спектра. Согласно теоретическим выкладкам, они совсем не должны были испускать лучи с меньшей длиной волны (желтой, зеленой и голубой частей спектра). Первые измерения, казалось бы, подтверждали этот вывод. Правда, они были сделаны грубым методом, с помощью цветных фильтров, надеваемых на объектив. Из этого следовало, что если безглазые креветки могут видеть свечение “черных курильщиков”, то их светочувствительные клетки должны быть “настроены” на красные или на близкие к инфракрасным лучи. Однако результаты первых исследований сетчатки креветок указывали на то, что это не так. Содержащийся в ней пигмент родопсин сильнее всего реагировал на зеленый свет с длиной волны около 500 нанометров. Можно было бы заподозрить, что полученные данные почему-либо искажены, но результаты весьма непростых опытов с регистрацией электрических сигналов, возникающих в сетчатке креветок, также указывали на то, что те могли видеть только зеленый свет. Это было странно. Если гидротермальные источники испускают красный свет, а креветки могут видеть только зеленый, значит, они все равно ничего не видят. Может быть, эта странная голая сетчатка представляет собой нефункциональные, вырожденные органы, вроде незрячих глаз пещерных рыб? Тот факт, что они находятся на спине, а не на голове креветок, заставлял предположить, что они не должны быть вырожденными глазами, но это было всего лишь предположение, а не доказательство.
Доказательство было получено, когда открыли личинок этих креветок. Мир гидротермальных источников не так долговечен, как кажется: отдельные поля таких источников часто перестают работать, забиваясь собственными выбросами, и срок их работы обычно не превышает продолжительности человеческой жизни. Тем временем на других участках дна океана начинают бить новые источники, часто за много миль от старых. Видам, связанным с подобными местообитаниями, жизненно необходимо уметь преодолевать разделяющие такие участки расстояния. Подвижности взрослых особей большинства из этих видов мешает их тесная связь со средой обитания (вспомним хотя бы гигантских рифтий, не имеющих ни рта, ни кишечника), но их личинки могут рассеиваться по океанам. Находят ли они новые поля источников случайно (расселяясь за счет океанических течений) или в результате активных поисков (например, отслеживая химические градиенты) — спорный вопрос. Но, так или иначе, личиночные формы подобных организмов совершенно не приспособлены к среде обитания взрослых форм. В основном они встречаются тоже на большой глубине, но гораздо ближе к поверхности, куда все же могут проникать тускнеющие лучи солнечного света. Иными словами, эти личинки живут в мире, где от глаз может быть польза.
Одним из первых видов, чьи личинки были открыты, стал краб Bythograea thermydron. Примечательно, что у взрослых особей этих крабов, как и у взрослых безглазых креветок, нет настоящих глаз, но имеется пара участков голой сетчатки. При этом у крабов, в отличие от креветок, такие участки расположены на голове — именно там, где обычно у животных находятся глаза. Однако самым поразительным открытием было то, что у личинок этого краба все-таки есть глаза, причем совершенно нормальные, по крайней мере для краба. Таким образом, там, где от глаз могла быть польза, у этих крабов имелись настоящие глаза.
Вскоре был описан целый ряд личинок других видов, связанных с гидротермальными источниками. Рядом с Rimicaris exoculata живет несколько других видов креветок, но их гораздо сложнее обнаружить, потому что они ведут одиночный образ жизни и не образуют таких же больших скоплений, как Rimicaris exoculata. У них тоже есть участки голой сетчатки, но не на спине, а на голове, а у их личинок, как и у личинок краба Bythograea thermydron, имеются совершенно нормальные глаза. Как ни странно, личинки Rimicaris exoculata были обнаружены в последнюю очередь — отчасти потому, что их очень легко спутать с личинками других креветок, а отчасти потому, что у них тоже есть нормальные глаза, расположенные на голове.
Открытие нормальных глаз у личинок креветок, лишенных таких глаз, говорило о многом. Это означало, что голая сетчатка представляет собой не продукт эволюционного упрощения глаз — результат постепенной их утраты, который еще может выполнять какие-то функции, подходящие для жизни в почти непроглядной темноте. У личинок имелись вполне нормальные глаза, исчезновение которых в ходе взросления явно не имело никакого отношения к необратимому постепенному вырождению из поколения в поколение, а было вызвано направленными эволюционными изменениями, каковы бы ни были плюсы и минусы этих изменений. Кроме того, это означало, что голая сетчатка не развилась в царстве вечной ночи из ничего, достигнув минимального качества работы, совершенно не сравнимого с качеством работы настоящего глаза. Напротив, по мере взросления личинок и их спуска на глубину, к гидротермальным источникам, их глаза упрощались и почти исчезали, а все их сложные оптические приспособления постепенно разрушались, пока не оставалась только голая сетчатка. В случае R. exoculata глаза при этом исчезали совсем, а голая сетчатка, судя по всему, развивалась на пустом месте — на спине. В итоге получалось, что для целого ряда животных, связанных с гидротермальными источниками, от голой сетчатки больше пользы, чем от полноценных глаз: это был не единичный случай, не какая-то случайность. Но почему?
Ценность голой сетчатки связана с соотношением разрешения и чувствительности. Разрешением называется способность видеть (различать) детали изображения. Увеличивать разрешение позволяют такие структуры, как хрусталик и роговица, помогающие фокусировать свет на сетчатке, проецируя на него изображение. Чувствительность же — это находящаяся в обратной зависимости от разрешения способность регистрировать фотоны. Когда в глаза попадает мало света, их чувствительность низкая. Человеческие глаза позволяют повышать нашу чувствительность к свету за счет расширения диафрагмы (зрачка) и перехода от использования менее светочувствительных клеток (колбочек) к использованию более светочувствительных (палочек). Но возможности подобных мер ограничены, и имеющиеся в наших глазах приспособления, необходимые для получения изображений хоть с каким-то разрешением, в конечном счете накладывают ограничения на нашу чувствительность к свету. Единственный возможный способ преодолеть эти ограничения предполагает отказ от хрусталика и предельное расширение диафрагмы, что существенно увеличит угол, под которым в глаз может попадать свет. Предельное расширение диафрагмы означает, что она должна совсем исчезнуть, оставив голую сетчатку. Принимая все это во внимание, можно провести расчеты, которые показывают, что голая сетчатка взрослых креветок, живущих у “черных курильщиков”, должна быть по меньшей мере в семь миллионов раз чувствительнее, чем полностью сформированные глаза их собственных личинок.
Итак, жертвуя разрешением, эти креветки приобретают способность регистрировать крайне слабый свет, а также, в какой-то степени, и отслеживать, откуда он поступает — по крайней мере, из какой половины поля зрения: сверху или снизу, спереди или сзади. Способность хоть как-то регистрировать свет может оказаться жизненно важной для обитания между зоной слишком горячей, где креветки могут за секунды свариться, и слишком холодной и удаленной от гидротермального источника, чтобы они могли там выжить. Креветка, потерявшая свой источник, представляется мне похожей на астронавта в открытом космосе, оторвавшегося от своего корабля. Возможно, поэтому у представителей вида R. exoculata, живущих в больших скоплениях на уступах прямо под гидротермальными источниками, “глаза” на спине. Несомненно, что так креветкам удобнее улавливать свет, учитывая, что они часто прячут голову в толпу сородичей. Их родственники, ведущие одиночный образ жизни, судя по всему, решили эту проблему немного иначе, и им ничто не мешает сохранять участки голой сетчатки на обычном месте глаз.
Вопрос, почему креветки видят зеленый свет в красном мире, мы обсудим позже. Сейчас же для нас главное, что половина глаза (голая сетчатка) может оказаться полезнее целого глаза — по крайней мере, в некоторых условиях. А чем половина глаза может быть лучше, чем полное его отсутствие, едва ли есть смысл обсуждать.
С голой сетчатки — большого светочувствительного пятна - обычно начинается обсуждение эволюции глаза. Еще Дарвин так представлял себе начало этого процесса. К прискорбию, его рассуждения по данному вопросу часто вырывают из контекста, причем не только те, кто отказывается признавать реальность естественного отбора, но порой и ученые, пытающиеся решить проблему, с которой великий Дарвин якобы не справился. При этом обычно приводят следующую цитату (вполне точную):
В высшей степени абсурдным, откровенно говоря, может показаться предположение, что путем естественного отбора мог образоваться глаз со всеми его неподражаемыми изобретениями для регуляции фокусного расстояния, для регулирования количества проникающего света, для поправки на сферическую и хроматическую аберрацию.
Что при этом слишком часто опускают — это следующее же предложение, из которого ясно, что на самом деле Дарвин вовсе не считал проблему эволюции глаза затруднением для своей теории:
Но разум мне говорит: если можно показать существование многочисленных градаций от простого и несовершенного глаза к глазу сложному и совершенному, причем каждая ступень полезна для ее обладателя; если, далее, глаз хоть немного варьирует и его вариации наследуются, а это также несомненно; если, наконец, подобные вариации могли оказаться полезными животному при переменах в условиях его жизни, в таком случае затруднение, возникающее при мысли об образовании сложного и совершенного глаза путем естественного отбора, хотя и непреодолимое для нашего воображения, едва ли может быть признано реальным.
Проще говоря, если одни глаза сложнее других, если разное зрение может наследоваться и если плохое зрение может быть помехой, тогда, по словам Дарвина, глаза могут эволюционировать. Все эти условия выполняются в массе случаев. В мире полно простых и несовершенных глаз, начиная от глазных пятен и ямок, не имеющих хрусталика, и заканчивая гораздо более развитыми глазами, которые могут похвастаться многими или всеми “неподражаемыми изобретениями”, о которых писал Дарвин. Зрение, несомненно, бывает разным, что особенно остро ощущает любой, кому приходится носить очки или мучиться, теряя зрение. Очевидно, что у тех, кто плохо видит, больше шансов погибнуть в когтях тигра или под колесами автобуса. При этом “совершенство” глаз, разумеется, относительно. Разрешение, которое обеспечивают глаза орла, в четыре раза превышает разрешающую способность глаз человека и позволяет птице различать детали на расстоянии мили. А наше зрение дает картину в восемьдесят раз более подробную, чем зрение многих насекомых, пикселизированное настолько, что его можно счесть особым жанром искусства.
Полагаю, большинство из нас без колебаний согласится, что оговоренные Дарвином условия выполняются, но все же не так-то просто представить себе все промежуточные стадии, вообразив непрерывный ряд изменений, ведущих к высокоразвитым глазам. Перефразируя Вудхауза, можно было бы сказать, что хотя это затруднение едва ли непреодолимо, одолимым его тоже назвать нельзя2. Как мы убедились, возникновение в ходе эволюции высокоразвитого глаза было бы невозможно, если бы каждая ступень этой эволюции не была полезна сама по себе. Однако на деле это затруднение оказывается вполне преодолимым. Шведские ученые Дан-Эрик Нильссон и Сусанна Пельгер представили искомый ряд в виде последовательности простых изменений (см. рис. на с. 283). Каждый этап этого ряда, начиная голой сетчаткой и заканчивая глазом, похожим на рыбий и не таким уж далеким от нашего, имеет преимущества перед предыдущим. Разумеется, этот ряд мог бы продолжаться (и действительно продолжался). Здесь можно добавить еще радужную оболочку, позволяющую расширять и сжимать зрачок, регулируя количество света, попадающего в глаз, в зависимости оттого, полдень на дворе или сумерки. К хрусталику можно прикрепить мышцы, позволяющие менять его форму, натягивая его и тем самым сплющивая, что даст глазу способность к фокусировке как на близких, так и на далеких предметах (аккомодации). Но эти тонкие усовершенствования имеются далеко не во всех высокоразвитых глазах и могли добавляться в ходе эволюции только к уже выстроенному глазу. Поэтому в данной главе мы ограничимся примерно таким рядом, как показан на рисунке, и обсудим происхождение функционального глаза — грубоватого, но уже достаточного для получения изображений3.
Для этого эволюционного ряда принципиально, что самый что ни на есть примитивный хрусталик лучше, чем отсутствие хрусталика (разумеется, где угодно, только не у “черных курильщиков”), потому что размытое изображение все же лучше, чем никакого. Вспомним также об обратной зависимости между разрешением и чувствительностью. Превосходное изображение можно получить даже с помощью камеры-обскуры, не используя никаких линз. Более того, у некоторых видов действительно имеются глаза, устроенные по типу камеры-обскуры, в частности у наутилусов — живых родичей ископаемых аммонитов4. Проблема, с которой сталкиваются наутилусы, связана с чувствительностью: для получения резкого изображения им нужна очень маленькая диафрагма, а значит, в глаз может попадать очень мало света. При плохом освещении изображение получается очень тусклое, почти неразличимое, и именно это происходит с наутилусами, живущими довольно глубоко под водой, куда проникает мало света. Майкл Лэнд из Университета Сассекса, один из ведущих специалистов по глазам животных, подсчитал, что если добавить хрусталик, не меняя размеров глаза, это позволяет увеличить светочувствительность в четыреста раз, а разрешение — в сто раз. Итак, любое изменение, приводящее к появлению хоть какого-нибудь хрусталика, может сразу же давать огромную выгоду организму, существенно повышая его шансы на выживание.

Последовательность этапов, требуемых для возникновения сложного глаза (по Дану-Эрику Нильссону и Сусанне Пельгер). Для каждого предполагаемого изменения приведена оценка числа поколений, смена которых может его обеспечить. Если предположить, что поколения сменяются раз в год, весь ряд займет меньше полумиллиона лет.
Первый настоящий хрусталик, позволяющий проецировать на сетчатку изображение, появился, по-видимому, у трилобитов — членистоногих, одетых в “латы” и напоминавших средневековых рыцарей. Многочисленные представители этой группы господствовали в морях триста миллионов лет. Древнейшие известные глаза трилобита видны на ископаемых остатках древнейшего известного представителя, жившего около 540 миллионов лет назад, то есть как раз примерно тогда, когда начинался “кембрийский взрыв”. Хотя эти глаза и выглядят довольно скромно по сравнению с шедеврами оптики, развившимися за следующие тридцать миллионов лет, внезапное появление глаз у трилобитов, отраженное в палеонтологической летописи, ставит вопрос: возможно ли, что высокоразвитые глаза действительно возникли так быстро? Если да, то “кембрийский взрыв” и в самом деле мог быть вызван именно эволюцией зрения, как утверждает Эндрю Паркер. Если нет, то высокоразвитые глаза, должно быть, существовали и ранее, но по какой-то причине в ископаемом виде не сохранились, и тогда, конечно, возникновение таких глаз едва ли могло быть причиной биологического Большого взрыва.
Большинство имеющихся данных указывают на то, что "кембрийский взрыв” случился в известное нам время потому, что именно тогда изменения среды позволили животным вырваться из “смирительной рубашки”, ограничивавшей их размеры. Предки кембрийских животных почти наверняка были крошечными и не имели твердых частей тела, чем и объясняется малочисленность их ископаемых остатков. Эти ограничения должны были мешать и развитию глаз. Для пространственного зрения необходим большой хрусталик, обширная сетчатка и мозг, способный интерпретировать информацию, поступающую от глаз, значит, такое зрение могло возникнуть лишь у животных достаточно крупных. Условия для его возникновения (в том числе голая сетчатка и зачаточная нервная система), вероятно, имелись и у мелких животных, существовавших задолго до кембрийского периода, но дальнейшее его развитие почти наверняка делали невозможным их малые размеры. Непосредственным импульсом к возникновению более крупных животных, скорее всего, послужило повышение уровня кислорода в атмосфере и в воде. Крупные размеры и хищничество возможны только при высоких концентрациях кислорода (иначе негде брать энергию: см. главу з), а уровень кислорода стал сопоставим с современным лишь незадолго до кембрия, вскоре после ряда периодов глобального оледенения, описываемых гипотезой “Земли-снежка”. В этой новой среде, насыщенной кислородом, впервые в истории нашей планеты могли появиться крупные животные, живущие за счет хищничества.
Но если до кембрийского периода настоящих глаз ни у кого не было, вновь встает вопрос, могли ли они развиться путем естественного отбора так быстро, и вопрос этот оказывается еще актуальнее. Могли ли высокоразвитые глаза уже возникнуть 540 миллионов лет назад, если 544 миллиона лет назад их еще не было? На первый взгляд может показаться, что палеонтологическая летопись противоречит дарвиновскому требованию миллиона тонких градаций, каждая ступень которых должна быть полезна для ее обладателя. Но на самом деле это кажущееся противоречие во многом объясняется несоответствием временных масштабов: с одной стороны, привычных нам продолжительности жизни и периодов смены поколений, а с другой — невообразимого масштаба геологических эпох. По меркам неспешно текущих сотен миллионов лет любые изменения, занимающие всего миллион лет, кажутся до неприличия поспешными, но для жизни отдельных организмов миллион лет — непомерно большой срок. Например, все современные породы собак развились из волка (надо признать, не без помощи человека) всего за сотую долю этого срока.
В геологическом масштабе времени “кембрийский взрыв" был мгновенным: он длился всего несколько миллионов лет. Однако в эволюционном масштабе это очень долго: даже полумиллиона лет могло оказаться более чем достаточно для возникновения высокоразвитых глаз. Предлагая свой вариант последовательности этапов их эволюции (рис. на с. 283), Нильссон и Пельгер также оценили вероятную продолжительность каждого этапа. Ученые исходили из осторожного предположения, что каждый этап предполагает изменение соответствующей структуры не более чем на 1 % (слегка более глубокое глазное яблоко, немного более плотный хрусталик, и так далее). Сложив длительность всех этапов, они с удивлением обнаружили, что путь от голой сетчатки до полностью сформированного глаза должен был потребовать всего около четырехсот тысяч элементарных изменений (гораздо меньше произвольного миллиона, о котором я упоминал). Далее они предположили, что на каждое поколение приходилось одно такое изменение (хотя их вполне могло быть и несколько, так что данное предположение тоже следует признать очень осторожным). И наконец, они предположили, что “средние” морские животные, организм которых эволюционировал, размножались один раз в год. Исследователи рассчитали, что развитие настоящего глаза должно было потребовать менее полумиллиона лет5.
Если эти расчеты верны, то возникновение высокоразвитых глаз действительно могло стать искрой, которая вызвала “кембрийский взрыв”. А если это так, то их изобретение определенно должно считаться одним из самых важных событий за всю историю жизни на Земле.
Схема Нильссона и Пельгер содержит лишь один сомнительный этап — первую стадию развития хрусталика. Нетрудно догадаться, как естественный отбор мог видоизменять и совершенствовать хрусталик, когда он уже возник, но как он вообще сформировался? Если части, требуемые для построения хрусталика, сами по себе ни для чего не нужны, не должен ли был естественный отбор просто-напросто отбросить их еще до того, как строительство могло начаться? Не может ли именно этой трудностью объясняться отсутствие хрусталика у наутилусов, хотя им и было бы выгодно этой структурой обзавестись?
На самом деле никакой трудности здесь нет, и глазам наутилусов, по крайней мере пока, придется оставаться необъясненной диковинкой, поскольку большинство глазастых животных (в том числе и ближайшие родственники наутилусов — осьминоги и кальмары) хрусталик приобрели, причем некоторые проявили при этом поразительную изобретательность. Хотя хрусталик явно представляет собой специализированную ткань, он неоднократно возникал в ходе эволюции из чего придется, конструировался из “строительных блоков”, прихваченных из любого источника, оказывавшегося под рукой — от минералов и кристаллов до ферментов и даже отдельных фрагментов клетки6.
Трилобиты дают нам отличный пример такого приспособленчества. У этих уникальных окаменелостей и при жизни были каменные глаза, потому что их хрусталики были из кристаллов кальцита (карбоната кальция). Известняк представляет собой кальците большим количеством примесей, мел — гораздо более чистую разновидность того же минерала. Белые скалы Дувра сложены из почти чистого кальцита, образованного беспорядочно расположенными крошечными кристаллами, отражающими во всех направлениях свет. Именно этим и обусловлен белый цвет мела. Однако если такие кристаллы растут медленно (это нередко бывает в минеральных жилах), кальцит может образовывать тонкие полупрозрачные структуры, немного отклоняющиеся от кубической формы, — ромбоэдры. У ромбоэдров есть одно любопытное оптическое свойство, естественным образом вытекающее из геометрических особенностей расположения атомов: они преломляют свет, падающий под любыми углами, кроме одного, соответствующего так называемой с-оси, проходящей через их середину. Если свет попадает в ромбоэдр параллельно этой оси, он проходит кристалл насквозь, не меняя направления, как почетный гость по ковровой дорожке. Трилобиты использовали это любопытное свойство себе на пользу. Каждая из множества фасеток в их глазах содержит собственный хрусталик из кальцита, сориентированный так, что его привилегированная с-ось направлена вглубь фасетки. Только свет, падающий на каждый хрусталик параллельно его с-оси, может проходить его насквозь, достигая лежащей под ним сетчатки.
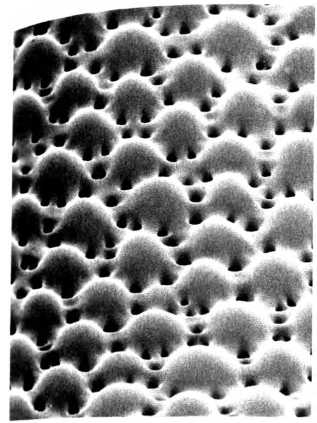
Как именно трилобиты выращивали кристаллы для своих хрусталиков, располагая их все под строго определенными углами, неизвестно — и, скорее всего, известно не будет, потому что последние трилобиты исчезли во времена пермского вымирания (около 250 миллионов лет назад). Но гибель трилобитов, последовавшая после столь продолжительного периода процветания, не означает, что у нас нет возможности узнать, как вообще могла возникнуть подобная система. Один ключ к разгадке этой тайны был получен в 2001 году из довольно неожиданного источника. Судя по всему, хрусталики трилобитов не уникальны, как считалось: одному современному морскому животному из класса офиур, как оказалось, тоже помогают видеть хрусталики из кальцита.

Внутренние поверхности кристаллических хрусталиков из глаз трилобита Dalmanitina socialis из ордовикских отложений Богемии (Чехия). Диаметр одного хрусталика около полмиллиметра.
Существует около двух тысяч видов офиур. У каждой офиуры имеется пять рук, как и у большинства морских звезд — их родственников, но, в отличие от морских звезд, руки у офиур очень длинные и тонкие, похожие на змеиные хвосты (отсюда и их название, означающее по-гречески “змеехвостки”). Скелет офиур сложен из сцепленных друг с другом кальцитовых пластин, которые, кроме того, образуют у них на руках шипы, помогающие этим хищникам хватать своих жертв. Большинство офиур нечувствительны к свету, но один вид, Ophiocoma wendtii, ставил исследователей в тупик своей способностью удирать при приближении опасности в какую-нибудь темную щель, находясь даже на расстоянии метра от нее. Загадка была в том, что у этих офиур нет глаз — по крайней мере, так все считали, пока исследователи из “Белл лабораториз” не заметили на руках этого животного группы кальцитовых бугорков, напоминающих хрусталики трилобитов. Ученые показали, что эти бугорки действительно играют роль хрусталиков, фокусируя свет на расположенных под ними светочувствительных клетках. Оказалось, что хотя у этих офиур нет настоящего мозга, у них все-таки имеются функциональные глаза. В журнале “Нэшнл джиографик” об этом открытии написали так: “По прихоти природы в морях у звезд есть глаза”.
Как образуются хрусталики у офиур? Хотя многие детали еще предстоит выяснить, ясно, что образуются они в целом так же, как и другие минерализованные структуры живых организмов, например иглы морских ежей (тоже состоящие из кальцита). Процесс начинается внутри клеток, где ионы кальция, накопленные в большой концентрации, взаимодействуют с белками, закрепляющими их в определенном положении, создавая “закваску” (точно так же в Советском Союзе один-единственный оптимист, вставший у входа в пустой продуктовый магазин, порой играл роль “отца-основателя” длинной очереди). Достаточно одному человеку или атому встать на определенном месте, как к нему начинают пристраиваться другие.
К вящей славе редукционизма, если очистить белки, отвечающие за создание “закваски” кристаллов кальцита, и нанести их на лист бумаги, а затем поместить в насыщенный раствор карбоната кальция, то прямо на этом листке вырастут безупречные кристаллы в форме ромбоэдров с оптической с-осью, направленной перпендикулярно бумаге, совсем такие же, как хрусталики трилобитов. Косвенные данные даже указывают, как изначально могли возникнуть такие хрусталики. Дело в том, что выбор конкретного белка не имеет особого значения: достаточно, чтобы из его молекулы торчали кислые боковые группы. Еще в 1992 году, за десять лет до того, как были открыты хрусталики офиур, биоминералоги Лия Аддади и Стивен Вайнер сумели вырастить на листе бумаги очаровательные кальцитовые ромбоэдры, используя для этого кислые белки, выделенные из раковин моллюсков, у которых определенно нет никаких глаз. Иными словами, каким бы удивительным ни был этот процесс, он может от начала до конца идти самопроизвольно, если смешать обычные белки с обычными неорганическими веществами. Это и в самом деле удивительно, но не требует никаких чудес, кроме тех, благодаря которым образовались фантастические друзы кристаллов, встречающиеся во многих подземных полостях (вспомним, например, Пещеру Мечей в Мексике).
Кристаллические хрусталики офиуры Ophiocoma wendtii, расположенные на скелетных пластинках на верхней стороне каждой руки, защищающих ее сочленения.

Ромбоэдрические кристаллы кальцита, выросшие на помещенном в насыщенный раствор карбоната кальция листке бумаги с нанесенными на нее кислыми белками из раковины моллюска. Оптическая с-ось, соответствующая единственному направлению, в котором свет проходит сквозь кристалл, не рассеиваясь, направлена перпендикулярно бумаге.
Хотя глаза с кристаллическими хрусталиками обеспечивали трилобитам довольно острое зрение, это был тупиковый путь.
Они важны прежде всего в историческом плане, как первые настоящие глаза, а не как один из долговечных шедевров эволюции. Другие животные научились использовать с той же целью другие кристаллы, в частности гуанин (один из “строительных блоков” ДНК), который может образовывать плоские кристаллы, позволяющие фокусировать свет. Кристаллы гуанина придают рыбьей чешуе ее серебристый блеск, и именно поэтому гуанин добавляют во многие косметические средства. Его можно найти и в гуано (от которого гуанин и получил свое название)- засохших экскрементах птиц или летучих мышей. Похожие органические кристаллы работают в живой природе как зеркала: именно они, например, придают кошачьим глазам зеркальный блеск. Такие кристаллы улучшают ночное зрение, отражая свет обратно на сетчатку и тем самым давая рецепторам еще один шанс уловить немногие попадающие в глаза фотоны. Другие зеркала отражают свет на сетчатку, создавая на ней изображение. Именно так устроены многочисленные прекрасные глаза морских гребешков, выглядывающие из-под щупалец у краев створок их раковины. Для фокусировки света в них используется вогнутое зеркальце, расположенное под сетчаткой. В фасеточных глазах многих ракообразных, в том числе раков и креветок, свет также фокусируется за счет зеркал, тоже состоящих из органических кристаллов, аналогичных гуаниновым.
Однако в целом эволюция больше полагалась на хрусталики, сделанные из особых белков, и именно они принесли ей больше всего славы. Были ли они тоже изготовлены кустарным способом из попавшихся под руку материалов, которые уже выполняли в организме какие-то другие функции? Хотя про эволюционную биологию иногда говорят, что, будучи исторической наукой, она не знает сослагательного наклонения, на самом деле она позволяет формулировать вполне конкретные и проверяемые предположения. В данном случае теория предполагает, что белки хрусталика должны были первоначально использоваться в организме для чего-то другого: специализированные белки хрусталика никак не могли возникнуть прежде, чем возник сам хрусталик.
Хрусталик человеческого глаза явно образован исключительно специализированной тканью: он прозрачен, в нем нет кровеносных сосудов, а его клетки утратили почти все свои нормальные функции. Они занимаются лишь тем, что концентрируют белки в жидкокристаллическую смесь, позволяющую хрусталику преломлять свет и проецировать на сетчатку отчетливое изображение. Хрусталик, кроме того, способен менять форму, меняя глубину резкости. Более того, степень преломления света оказывается разной в разных частях хрусталика. Это позволяет компенсировать такие нежелательные эффекты, как сферическая аберрация (она приводит к тому, что свет, проходящий через центр и через края линзы, фокусируется в разных точках). Учитывая все это, мы могли бы предположить, что белки, требуемые для получения такой замечательной смеси, будут единственными в своем роде и их оптические свойства не будут встречаться ни у каких из обычных белков. И оказались бы совершенно не правы.
Белки хрусталика человеческого глаза назвали кристаллинами, ожидая, что они окажутся единственными в своем роде. Кристаллины составляют около 90 % всех белков в хрусталике. Поскольку хрусталики разных видов позвоночных похожи и внешне, и функционально, казалось разумным предположить, что все они сделаны из похожих белков. Однако когда стали широко доступны технологии сравнения последовательностей “строительных блоков” в молекулах белков (с 8о-х годов XX века), ученые с удивлением обнаружили, что кристаллины вовсе не относятся к структурным белкам, причем большинство из них встречаются не только в хрусталике и выполняют в организме и другую работу. Еще большей неожиданностью стало то, что многие из кристаллинов оказались ферментами (биологическими катализаторами), выполняющими “хозяйственные" функции в других частях организма. Например, больше всего в человеческом хрусталике так называемого альфа-кристаллина: он близок к белку теплового шока, впервые найденному у плодовых мушек дрозофил и, как теперь известно, широко распространенному у животных. У людей он играет роль шаперона, то есть оберегает другие белки от повреждений. Его можно найти не только в глазах, но и в мозге, печени, легких, селезенке, коже и тонкой кишке.
К настоящему времени описаны одиннадцать разновидностей кристаллинов. Только три имеются в глазах у всех позвоночных, остальные же есть лишь у некоторых групп, а значит, по-видимому, были “призваны на службу” в хрусталик, совершенно независимо, что, опять же, и следует из предположения, что компоненты хрусталика были подобраны эволюцией оппортунистически. Мы не станем останавливаться на названиях и функциях этих белков. Отметим лишь, что все они, как ни странно, выполняют в клетках метаболические функции, причем разные. “Ополчение” кристаллинов как будто набрано исключительно из ремесленников, к тому же из разных гильдий. Но каковы бы ни были причины этого странного отбора рекрутов, он отнюдь не указывает на то, что эволюции было сколько-нибудь сложно набирать белки для хрусталиков.
В целом в белках хрусталика нет ничего особенного: они позаимствованы из каких-то других частей организма и отправлены служить в глазах. Почти все белки прозрачны, поэтому сих цветом проблем не было (лишь некоторые белки, соединенные с пигментами, такие как гемоглобин, отличаются собственной яркой окраской). Изменение оптических свойств, например степени преломления света (рефракции), различающейся в разных частях хрусталика, достигается просто за счет варьирования концентраций разных белков. Это определенно требует тонкой работы, но едва ли сопряжено с принципиальными трудностями. Почему среди белков хрусталика так много ферментов, неизвестно, но с чем бы это ни было связано, ясно, что белки хрусталика не могли появиться сразу в готовом виде, как Афина вышла из головы Зевса.
На возникновение всего этого проливает свет одно из морских беспозвоночных, называемых оболочниками, а именно Ciona intestinalis (в буквальном переводе “кишечный столб” — Карл Линней неласково обошелся с этим существом). По взрослым особям этого вида трудно судить об их происхождении: они представляют собой прикрепленные к подводным камням полупрозрачные бочонки с двумя нежесткими желтоватыми сифонами, через которые входит и выходит вода. В прибрежных водах Великобритании их так много, что они считаются вредителями. Но их личинки выдают глубокую тайну, показывающую, что эти животные заслуживают большего, чем звание обыкновенных вредителей. Личинки оболочников немного похожи на головастиков и могут активно плавать, пользуясь при этом слаборазвитой нервной системой и парой примитивных глаз, не имеющих хрусталика. Когда маленький оболочник находит подходящее место, он надежно прикрепляется к субстрату, после чего, больше не нуждаясь в собственных мозгах, частично переваривает их (Стив Джонс шутит, что этот трюк вызывает восхищение у многих университетских профессоров).
Хотя во взрослых оболочниках трудно узнать наших родственников, их личинки, похожие на головастиков, выдают тайное родство с нами: эти существа относятся к примитивным хордовым и действительно обладают хордой, на основе которой у позвоночных животных формируется позвоночник. На генеалогическом древе хордовых, в основном представленном позвоночными, оболочники составляют одну из нижних ветвей. Основание этой ветви отделилось от ствола хордовых еще до того, как они обзавелись хрусталиками. А это значит, что оболочники с их примитивными глазами вполне могут пролить свет на происхождение хрусталика позвоночных.
И действительно проливают: в 2005 году Себастиан Шимелд и его коллеги из Оксфорда выяснили, что хотя у C. intestinalis нет настоящего хрусталика, у него есть один полноценный белок из числа кристаллинов (только находится он не в глазах, а спрятан в нервной системе). Сложно сказать, что он там делает, но это и не имеет отношения к интересующему нас вопросу. А имеет к нему отношение тот факт, что те же самые гены, которые направляют развитие хрусталика у позвоночных, также управляют активностью этого белка, и у оболочника они работают как в нервной системе, так и в глазах. Так что весь аппарат, необходимый для построения хрусталика, уже имелся у общего предка позвоночных и оболочников. Произошедшее у позвоночных небольшое изменение в регуляции работы генов привело к тому, что этот белок стал синтезироваться в глазах. Подобные браконьерские набеги на другие органы позволяли набирать в армию кристаллинов и другие белки. Некоторые из них были набраны еще у общих предков всех позвоночных, некоторые — позже, у предков отдельных групп. Почему зволюционная линия оболочников так и не смогла осуществить простое перераспределение ресурсов, требуемое для построения хрусталика, остается загадкой: найти подходящий для прикрепления подводный камень нетрудно и без хрусталика. В любом случае оболочники остались в меньшинстве. Большинство хордовых успешно проделало подобный трюк, причем происходило это не менее одиннадцати раз. Так что в последовательности изменений, требуемых для возникновения высокоразвитого глаза, нет ни одного особенно сложного этапа.
Белки сетчатки сильно отличаются от разношерстных команд из набранных отовсюду белков и кристаллов, составляющих хрусталики разных видов животных. Значение одного из таких белков особенно велико. Это родопсин — вещество, ответственное за восприятие света. Вспомним живущих у “черных курильщиков” креветок Rimicaris exoculata с их голой сетчаткой. Несмотря на необычность мира “черных курильщиков”, на странность ношения на спине двух участков голой сетчатки, на способность этих креветок улавливать невидимое нашему глазу слабое свечение, на их питание за счет серных бактерий, голубую кровь и отсутствие позвоночника, а также на то, что наш последний общий с ними предок жил около шестисот миллионов лет назад, задолго до “кембрийского взрыва”, видеть этим креветкам позволяет тот же белок, что и нам. Чем обусловлена эта глубокая связь, проходящая сквозь время и пространство: поразительной случайностью или чем-то более важным?
Родопсин креветок немного отличается от нашего, но тот и другой так похожи, что если бы вам пришлось доказывать в суде, что ваша версия этого белка — оригинальное произведение, а не плохо прикрытый плагиат, у вас было бы мало шансов на победу. Более того, вас бы подняли на смех, потому что родопсин свойствен отнюдь не только глубоководным креветкам и людям: в животном мире он присутствует почти повсеместно. Например, хотя мы мало знаем о тонком механизме глаз трилобитов (он, в отличие от их хрусталиков, не сохранился), мы знаем о родственниках этих животных достаточно, чтобы довольно уверенно сказать, что и у них в глазах содержался родопсин. Всем животным, за редкими исключениями, позволяет видеть один и тот же белок. Пытаться убедить суд, что ваш родопсин — не плагиат, было бы все равно что доказывать, что ваш телевизор принципиально отличается от все* остальных только потому, что он больше, или потому, что у него плоский экран.
Это поразительное единообразие теоретически могло возникнуть несколькими путями. Оно может означать, что мы все унаследовали от общего предка один и тот же белок. За последние шестьсот миллионов лет наши родопсины, конечно, претерпели немало изменений, но, по сути, они остались тем же белком. Кроме того, это единообразие может означать, что существуют объективные ограничения, требующие, чтобы молекулы, позволяющие воспринимать свет, были принципиально устроены именно так и никак иначе, и в итоге нам всем пришлось прийти именно к такому строению светочувствительного белка. Это было бы похоже на просмотр телепрограмм на экране компьютера — случай, когда разные технологии дают похожие решения. Наконец, единообразие родопсинов может означать, что ген этого белка свободно передавался от одного вида другому и достался многим не по наследству, а в результате бесстыдного воровства.
Третью возможность легко отвергнуть. Виды действительно иногда воруют гены друг у друга (например, гены могут передаваться от одного вида другому с вирусами), но эта практика широко распространена только у бактерий, и когда такое воровство происходит, его очень сложно скрыть. Чтобы его выявить, достаточно сопоставить наборы тонких различий между белками разных видов с их известными генеалогическими связями. Например, если бы родопсин креветок был украден у людей, он выделялся бы на фоне других их белков, как нелегальный иммигрант, всем видом выдавая свое происхождение от людей, а не от креветок. Если же родопсин креветок достался им от общего предка с их близкими родственниками, раками, то он должен был накопить гораздо меньше отличий от их родопсинов, чем можно найти между ним и родопсинами более далеких родственников креветок, таких как мы. Это именно так.
Если родопсин не был никем украден, то был ли он изобретен неоднократно, в силу чисто технической необходимости? На этот вопрос ответить сложнее, потому что после сравнения разных родопсинов действительно может сложиться впечатление, что они были изобретены независимо, по крайней мере дважды. Родопсин креветок едва ли не так далек от нашего, как только возможно для двух очень похожих друг на друга молекул. Существует целый спектр родопсинов, занимающих промежуточное положение, однако этот ряд не непрерывен. Все родопсины можно разделить на две группы, более или менее соответствующие с одной стороны позвоночным, с другой — и беспозвоночным (в том числе креветкам). Разница между этими группами усугубляется целым рядом различий между контекстами, в которые вписан родопсин. Светочувствительные клетки в обоих случаях представляют собой модифицированные нервные клетки, но этим их сходство и ограничивается. У креветок и других беспозвоночных родопсин вставлен в мембраны выростов (микроворсинок), торчащих на вершине светочувствительной клетки как колючки, в то время как у позвоночных на вершине клетки имеется единственный вырост (ресничка), высящийся там как радиомачта. На этой мачте есть ряд глубоких горизонтальных складок, делающих ее похожей на стопку дисков, лежащую на клетке.
Внутри светочувствительных клеток эти различия дополняются другими, биохимическими. У позвоночных при поглощении родопсином света возникает каскад сигналов, увеличивающих электрическое напряжение на мембране клетки. У беспозвоночных же происходит ровно обратное: при поглощении родопсином света запускается совершенно другой каскад, приводящий к тому, что электрическое напряжение на мембране совсем пропадает, и именно это и вызывает передачу по зрительному нерву в мозг сигнала “свет”. Так что довольно похожие друг на друга родопсины позвоночных и беспозвоночных работают в клетках совершенно разного типа. Означает ли все это, что светочувствительные клетки возникли в ходе эволюции дважды: один раз у беспозвоночных и один раз у позвоночных?
Это кажется вполне правдоподобным, и такое мнение разделяло большинство работавших в данной области ученых до середины 90-х годов XX века, когда все неожиданно изменилось. Все, что было известно тогда, соответствовало действительности, однако оказалось, что это далеко не вся история. Теперь складывается впечатление, что все животные используют родопсин именно потому, что унаследовали его от общего предка. Похоже, что первоначальный прототип глаза возник входе эволюции лишь однажды.
Пересмотром господствующих представлений мы во многом обязаны революционным достижениям швейцарского специалиста по биологии развития Вальтера Геринга из Базельского университета. Геринг был в числе первооткрывателей так называемых гомеозисных генов (ответственных за реализацию планов строения), а в 1995 году сделал еще одно эпохальное открытие, поставив один из самых поразительных экспериментов в истории биологии. Исследователи из команды Геринга взяли один из генов мыши и встроили его в геном плодовой мушки дрозофилы. Это был необычный ген: он обладал комплексным действием, и под его вредным влиянием на ногах, на крыльях и даже на усиках мушек начали расти глаза. Эти неуместные крошечные глаза были устроены не по принципу фотоаппарата, как знакомые нам глаза мышей и людей, а примерно так же, как нормальные фасеточные глаза, состоящие из множества фасеток, характерные для насекомых и ракообразных. Пугающие результаты эксперимента красноречиво свидетельствовали о том, что у мышей и мух для формирования глаз требуются одни и те же гены, сохранившиеся за шестьсот миллионов лет эволюции — со времен последнего общего предка позвоночных и беспозвоночных, - и столь мало изменившиеся за это время, что они по-прежнему в состоянии подменять друг друга. Стоило поместить мышиный ген в геном дрозофилы, и он подчинял себе ее механизмы развития, и в местах, где он работал, гены дрозофилы по его команде тут же начинали формировать глаза.

Голова плодовой мушки дрозофилы под сканирующим электронным микроскопом. На конце усика виден крошечный дополнительный глаз, сформировавшийся под влиянием внедренного в геном дрозофилы гена Pax6. Тот же самый ген управляет развитием глаз и у позвоночных, и у беспозвоночных и, должно быть, выполнял эту же функцию у их общего предка, жившего около шестисот миллионов лет назад.
В Базеле когда-то преподавал Ницше. Геринг — быть может, в память о его “морали господ” — обозначил этот мышиный ген термином “ген-господин” (master gene). По-моему, здесь подошел бы другой термин — “ген-маэстро” (maestro gene). По крайней мере, он звучит менее напыщенно и, быть может, оказался бы уместнее и в ряде других случаев. Подобно дирижеру, по мановению палочки которого возникает прекраснейшая музыка, хотя сам он не издает ни звука, этот ген вызывает формирование структур глаза, управляя игрой других генов, каждый из которых отвечает за собственную партию. Различные варианты этого гена были уже известны по мутациям в нем, происходящим у дрозофил, мышей и людей. У мышей и дрозофил его называли соответственно “маленький глаз” (Small eye) и “безглазый” (Eyeless), указывая (генетики питают ужасную склонность к выворачиванию слов наизнанку) на различные результаты прекращения его нормальной работы. У нас мутации в этом гене вызывают болезнь аниридию, при которой у человека не развивается радужная оболочка. Хотя симптомы этой болезни малоприятны и она нередко приводит к слепоте, это на удивление умеренные последствия для выключения “гена-господина”, который, как предполагается, руководит развитием всего глаза. Но так бывает только в случаях, когда повреждена лишь одна из двух его копий. Если повреждены или утрачены обе, у зародыша вообще не формируется голова.
С тех пор, как Геринг провел свой революционный эксперимент, нам открылась более сложная картина, чем казалось поначалу. Его “ген-господин” теперь известен под названием Pax6, и выяснилось, что он, с одной стороны, еще влиятельнее, чем представлялось, а с другой стороны, не так одинок в своем величии. Pax6 удалось обнаружить почти у всех позвоночных и беспозвоночных, в том числе у креветок, а близкородственный ему ген имеется даже у медуз. Причем оказалось, что Pax6 стоит за формированием не только глаз, но и значительной части мозга, поэтому если у зародышей не работают обе копии этого гена, у них не развивается голова. В то же время Pax6 — не единственный подобный ген. Есть и другие гены, под действием которых у дрозофил могут формироваться целые глаза — судя по всему, это на удивление простой трюк. Все эти гены явно родственны друг другу и существуют очень давно. По большей части они имеются и у беспозвоночных, и у позвоночных, хотя и играют немного разные роли и вписаны в разные контексты. Как это ни грустно, прекрасной музыкой жизни управляет не дирижер, а небольшой комитет.
Итак, формированием глаз и у позвоночных, и у беспозвоночных управляет один и тот же “комитет” генов. В отличие от родопсина, этот комитет не обязан быть именно таким по “техническим” причинам. Все входящие в него гены — безликие бюрократы, место которых с тем же успехом могла бы занимать кучка других безликих бюрократов. Тот факт, что формированием глаз (в отличие, например, от синтеза белков хрусталика) всегда руководит одна и та же кучка генов, свидетельствует о том, что история действовала по прихоти случая, а не в силу необходимости. И история эта заставляет нас предположить, что светочувствительные клетки животных возникли входе эволюции лишь однажды, у общего предка позвоночных и беспозвоночных, под управлением одного и того же небольшого “комитета” генов.
Для предположения, что светочувствительные клетки животных возникли лишь однажды, есть и еще одно основание — непосредственное свидетельство живого ископаемого. Это крошечный многощетинковый червь Platynereis из семейства нереид. Черви этого рода, достигающие в длину несколько сантиметров, живут в заиленных лагунах, и рыболовы охотно используют их как наживку. Интересно, многие ли из этих рыболовов знают, что форма тела и морфология этих червей почти не изменилась с кембрийского периода? Похожий на них червь, живший еще раньше, был общим предком и позвоночных, и беспозвоночных. Как и все позвоночные и многие беспозвоночные, Platynereis обладает билатеральной симметрией, то есть его правая сторона выглядит как зеркальное отражение левой. Некоторым беспозвоночным, например морским звездам, свойствен другой тип симметрии. Билатерально симметричных животных, от насекомых до нас с вами, объединяют в группу билатерий. Принципиально, что эволюционная линия нашего червя возникла до того, как этот тип строения начал реализовывать свой огромный потенциал, породив все свои удивительные воплощения, которые мы наблюдаем повсюду. Так что Platynereis может считаться живым ископаемым, представляющим первоначальных билатерий, протобилатерий, и именно поэтому Детлев Арендт и его коллеги из Европейской молекулярно-биологической лаборатории в Гейдельберге заинтересовались его светочувствительными клетками.
Они знали, что глаза этого червя похожи по устройству на глаза других беспозвоночных, а не на глаза позвоночных (в частности, используют соответствующую разновидность родопсина). Но в 2004 году исследователи из Гейдельберга обнаружили у этого червя еще одну группу светочувствительных клеток, спрятанных в глубине нервной системы. Эти клетки вообще не идут на нужды зрения, а задействованы в работе внутренних часов, определяющих суточный ритм, который управляет сном и бодрствованием и помогает отличать день от ночи - даже у бактерий. Оказалось, что клетки внутренних часов червя Platynereis не только используют родопсин, но и представляют собой вполне узнаваемые (по крайней мере, для таких специалистов, как Арендт) светочувствительные клетки позвоночных. Этот вывод впоследствии подтвердился в ходе более подробных биохимических и генетических исследований. Арендт заключил, что у протобилатерий имелись оба типа светочувствительных клеток, а значит, клетки каждого типа не возникли независимо друг от друга в двух совершенно разных эволюционных линиях, а были когда-то “сестринскими” клетками, эволюционировавшими вместе у одних и тех же организмов — предков протобилатерий.
Разумеется, если у этого общего предка позвоночных и беспозвоночных были оба типа светочувствительных клеток, то мы тоже вполне могли унаследовать клетки обоих типов, у нас тоже можно найти и клетки другого типа (конечно, если знать, где искать). Похоже, что так и есть. В том же году, когда живые ископаемые открыли ученым свою тайну, Сатчин Панда и его коллеги из Института Солка в Сан-Диего попытались проверить свои подозрения относительно некоторых клеток человеческого глаза - ганглионарных клеток сетчатки, влияющих на наш суточный ритм. Хотя эти клетки и не специализируются на регистрации света, в них тоже содержится родопсин. Он представлен необычной формой — так называемым меланопсином, которая, как выяснилось, характерна для светочувствительных клеток беспозвоночных. Весьма примечательно, что этот связанный с суточным ритмом родопсин наших глаз по структуре ближе к родопсину участков голой сетчатки креветок, живущих у “черных курильщиков”, чем к другому типу родопсина, работающему вместе с ним в человеческой сетчатке.
Это означает, что светочувствительные клетки позвоночных и беспозвоночных появились из одного и того же источника. Это не два разных изобретения, а сестринские клетки, у которых была общая праматерь. И эта праматерь, первичная светочувствительная клетка, прародительница глаз всех животных, возникла в ходе эволюции лишь однажды.
Складывается следующая картина. У общих предков позвоночных и беспозвоночных развились светочувствительные клетки одного типа, содержащие зрительный пигмент родопсин и формирующиеся под управлением небольшого комитета из генов. Впоследствии эти светочувствительные клетки разделились на два типа, которые стали специализироваться либо на зрении, либо на суточном ритме. По неизвестным причинам (может быть, случайно) клетки этих двух типов у позвоночных и у беспозвоночных выбрали для специализации противоположные функции, так что глаза у тех и у других развивались на основе разных тканей. Это обусловило принципиальные различия в эмбриональном развитии глаз между такими представителями позвоночных и беспозвоночных, как человек и осьминог. Первой остановкой на пути к высокоразвитому глазу стала голая сетчатка — слой светочувствительных клеток одного или другого типа, в зависимости от эволюционной линии. У некоторых организмов сохранились простые, плоские участки голой сетчатки, в то время как у других эти участки прогнулись, и сетчатка погрузилась в ямки, так что на нее теперь могла падать тень, позволяя определять, откуда идет свет. Когда эти ямки стали достаточно глубокими, на их работе начала сказываться обратная зависимость между чувствительностью и разрешением, означавшая, что любой хрусталик лучше, чем никакого, и для выполнения функций хрусталика были собраны подвернувшиеся материалы, от минералов до ферментов. Сходные процессы имели место в различных эволюционных линиях, породив неразбериху. Но оптические законы позволяют построить лишь несколько принципиальных разновидностей функциональных глаз, что накладывало ограничения на пути их развития. Поэтому все сложившееся на молекулярном уровне разнообразие на макроскопическом уровне свелось к немногим принципиальным структурам: от наших собственных глаз, устроенных по типу фотоаппарата, до фасеточных глаз насекомых.
Разумеется, многие подробности здесь опущены, но в общих чертах эволюция глаза именно такова. Неудивительно, что у нас и у креветок, живущих на “черных курильщиках”, один итотже родопсин: мы все унаследовали его от одного и того же древнего предка. Но у нас по-прежнему остается еще один вопрос: кем был этот предок? Ответ нам дают тоже гены.
Когда Синди Ван Довер спускалась к “черным курильщикам”, ее неотступно преследовала мысль о свете. Креветки, которых она изучала, судя по всему, отлично улавливали зеленый свет с помощью родопсина, похожего на человеческий. При этом имевшиеся тогда измерения показывали, что “черные курильщики” не светятся зеленым. В чем же дело?
Один выдающийся исследователь в выступлении, посвященном своему уходу на пенсию, дал молодым ученым шуточный совет: ни в коем случае не пытаться повторить успешный эксперимент, потому что результаты таких попыток неизменно приносят горькое разочарование8. Не так уж очевидно, что может быть верно и обратное (всегда без колебаний повторять неудавшиеся эксперименты), но у Синди Ван Довер были все основания попробовать. Родопсины, как и покойники, не лгут. Синди Ван Довер рассуждала так: если родопсин креветок поглощает зеленый свет, значит, там должен быть зеленый свет. Оставалось предположить, что примитивное оборудование, использованное для первых измерений, было не так чувствительно, как голая сетчатка креветок.
Был заказан новый, намного более совершенный фотометр, спроектированный учеными из НАСА, знавшими все о регистрации излучений в черноте космического пространства. Этот прибор назывался ALIS5 (Ambient Light Imaging and Spectral System — “система визуализации и спектрометрии окружающего света”), и он действительно позволил зарегистрировать свет с другими длинами волн. С помощью ALISS в стране чудес “черных курильщиков” удалось выявить небольшой пик в зеленой части спектра, где интенсивность света была на несколько порядков выше, чем предсказывала теория. Результаты новых измерений вскоре подтвердились и на других полях “черных курильщиков”. Хотя источник этого мистического света по-прежнему остается загадкой, нет недостатка в остроумных гипотезах. Одна из них предполагает, что видимый свет могут испускать выходящие из гидротермальных источников пузырьки газа, сжимаемые высоким давлением воды, подобно тому, как это может происходить при образовании и разрушении кристаллов под высоким давлением при высокой температуре.
Синди Ван Довер не зря верила в родопсин: она знала, что что-то тут не так. У родопсинов есть удивительная способность соответствовать условиям окружающей среды. Море не зря называют синим: этот свет проходит сквозь воду дальше, чем свет с другими длинами волн. Море быстро поглощает красный свет, и он не может проникать глубоко под воду. Желтый заходит немного глубже, оранжевый — еще глубже. Но начиная с глубины около двадцати метров под водой остается в основном зеленый, голубой и синий свет, и чем глубже, тем больше доля синего. Этот свет рассеивается в воде, на больших глубинах окрашивая все в разные опенки синего. Родопсины рыб прекрасно отслеживают этот сдвиг в синюю часть спектра, пользуясь так называемой спектральной настройкой. В итоге на глубине восьмидесяти метров мы находим рыб, родопсины которых лучше всего поглощают зеленый цвет (с длиной волны около 520 нанометров), но на глубине двухсот метров, куда проникают в основном лишь тусклые лучи синего цвета, у рыб имеются родопсины, поглощающие синий цвет (с длиной волны около 450 нанометров). Интересно, что уже знакомый нам краб В. thermydron, живущий у “черных курильщиков”, спускаясь на дно, совершает обратный переход. Личинки этого краба живут на синих глубинах и пользуются родопсином, лучше всего поглощающим синий цвет (длина волны около 450 нанометров). А голая сетчатка взрослых крабов содержит родопсин, поглощающий свет с длиной волны 490 нанометров: он ближе к зеленой части спектра. Учитывая, что родопсин креветок, живущих у “черных курильщиков”, тоже поглощает зеленый свет, длина волны которого около 500 нанометров, Синди Ван Довер имела все основания подозревать, что это неслучайно.
Наше цветовое зрение тоже зависит от способности родопсинов поглощать свет с разными длинами волн. В нашей сетчатке имеются светочувствительные клетки двух типов: палочки и колбочки. Строго говоря, только палочки содержат родопсин, а колбочки содержат один из трех собственных, колбочковых опсинов. Но эта классификация ничего нам не дает, поэтому что все зтизрительные пигменты обладают примерно одинаковой принципиальной структурой: все они состоят из того или иного белка (опсина), встроенного в клеточную мембрану, где он образует семикратный зигзаг, и связанного с ним производного витамина А — так называемого ретиналя. Ретиналь — пигмент, отвечающий за поглощение света. Поглотив фотон, молекула ретиналя меняет свою форму с изогнутой на прямую, и этого оказывается достаточно, чтобы запустить работу всего биохимического каскада, в итоге посылающего в мозг сигнал “свет”.
Хотя свет поглощает именно ретиналь, важнейшим фактором спектральной настройки оказывается как раз структура белка опсина. Небольшие изменения в ней могут менять цвет поглощаемого ретиналем света с ультрафиолетового (около 350 нанометров), как бывает у насекомых и птиц, до красного около 625 нанометров), как у хамелеонов. В итоге, используя несколько разных опсинов, поглощающих свет с разными длинами волн, можно получить цветовое зрение. Наши собственные колбочковые опсины поглощают преимущественно свет синей (433 нанометра), зеленой (535 нанометров) и красной (564 нанометра) частей спектра, вместе обеспечивая привычный диапазон видимого света.
Хотя общая структура опсинов похожа, по различиям между ними можно восстановить интереснейшую историю эволюции их обладателей. Все они возникли за счет удвоения и последующего эволюционного расхождения генов, и у истоков их всех стоял один и тот же ген опсина. Ясно, что одни удвоения происходили раньше, другие — позже. Например, наши “красный” и “зеленый” опсины состоят в довольно близком родстве: породивший их ген удвоился лишь у общего предка приматов. Это удвоение дало приматам три типа колбочковых опсинов (по крайней мере, впоследствии, когда полученные два гена немного разошлись) вместо двух, благодаря чему и большинство людей обладают трехцветным (трихроматическим) зрением. У тех немногих, которые, к несчастью, родились дальтониками, не способными различать красный и зеленый цвета, один из этих генов вновь утрачен, что делает их зрение дихроматическим, как почти у всех остальных млекопитающих. Слабость цветового зрения большинства млекопитающих, возможно, связана с ночным образом жизни их сравнительно недавних предков, который помогал им прятаться от господствовавших в те времена динозавров. Почему приматы вернули себе трехцветное зрение — вопрос спорный. Самая популярная теория предполагает, что оно помогало им замечать красные плоды на фоне зеленой листвы. Согласно другой теории, придающей большее значение социальным факторам, трехцветное зрение возникло у приматов потому, что помогало им различать эмоции, признаки угрозы и сигналы потенциальных половых партнеров — от краски стыда до неприкрытого вранья (интересно, что у всех приматов, обладающих трихроматическим зрением, лица не прикрыты шерстью).
Я сказал, что приматы “вернули себе” трехцветное зрение, но на самом деле в том, что касается зрения, мы по-прежнему остаемся бедными родственниками многих других позвоночных. У рептилий, птиц, амфибий и акул зрение четырехцветное, и вполне вероятно, что общий предок всех позвоночных тоже обладал тетрахроматическим зрением и способностью видеть ультрафиолетовый свет10. Эту возможность подтвердили результаты одного очаровательного эксперимента: сравнив ДНК-последовательности генов современных позвоночных, Ши Юншэн и Сёдзо Ёкояма из Сиракузского университета в штате Нью-Йорк реконструировали последовательность одного из генов, которым обладал общий предок всех позвоночных. По одной последовательности мы пока не можем судить, какой была длина волны света, поглощаемого соответствующим опсином. Но это не остановило Ши и Ёкояму: они воспользовались методами генной инженерии, чтобы синтезировать кодируемый данным геном белок, а затем непосредственно измерили спектр поглощаемого света. Как и ожидалось, пик поглощения пришелся на ультрафиолетовую часть спектра (360 нанометров).
Ствол генеалогического древа опсинов, как мы убедились, восходит к общим предкам позвоночных и беспозвоночных. Но даже у такого живого ископаемого, как близкий к протобилатериям многощетинковый червь Platynereis, имеются уже два разных опсина, соответствующих опсинам позвоночных и беспозвоночных. На что же был похож самый древний предок опсинов всех животных, и откуда он взялся? На этот счет существует несколько гипотез. Но нашими главными проводниками в прошлое до сих пор были сами гены, и с их помощью мы смогли преодолеть промежуток в 600 миллионов лет. Не укажут ли они нам дорогу в еще более далекое прошлое? Петер Хегеманн и его коллеги из Регенсбургского университета в Германии утверждают, что гены действительно позволяют ответить на интересующий нас вопрос, и ответ этот оказывается совершенно неожиданным. Если верить выводам исследователей, самый первый прототип глаза возник даже не у животных, а у водорослей!
Водоросли, как и растения, — мастера фотосинтеза, осуществлять который им помогают самые разные светочувствительные пигменты. У многих водорослей есть пигменты, содержащиеся в простом глазном пятне и используемые, чтобы отслеживать интенсивность света и при необходимости как-то на нее реагировать. Например, удивительно красивая микроскопическая водоросль вольвокс образует состоящие из сотен клеток полые сферы до миллиметра в диаметре. У каждой клетки имеются два жгутика, торчащие наружу, как весла. Они работают в темноте, но останавливаются на свету, благодаря чему вся сфера перемещается в сторону Солнца и может находить наилучшие условия для фотосинтеза. Команда остановить работу жгутиков поступает от глазных пятен. Самое же удивительное то, что работающий в глазных пятнах вольвокса пигмент оказался родопсином.
Еще большей неожиданностью оказалось то, что родопсин вольвокса производит впечатление предкового по отношению к опсинам всех животных. Место молекулы этого родопсина, где ретиналь связывается с белком, содержит участки, точно совпадающие с соответствующими участками опсинов позвоночных и беспозвоночных, и производит впечатление промежуточного между теми и другими. Общая структура гена этого родопсина, представляющего собой пеструю смесь кодирующих и некодирующих последовательностей (известных в науке как интроны и экзоны), также выдает его древнюю связь с опсинами как позвоночных, так и беспозвоночных. Это еще ничего не доказывает, но именно такими особенностями должен был обладать белок, от которого произошли оба семейства опсинов. А значит, вполне вероятно, что прародительницей глаз всех животных была, как ни странно, фотосинтезирующая водоросль.
Но здесь, разумеется, возникает вопрос, как родопсин водорослей мог достаться животным? Вольвокс никак не может быть предком животных. Но если взглянуть на устройство его глазного пятна, можно сразу увидеть возможный ключ к разгадке: родопсин у вольвокса встроен в мембрану хлоропласта - одной из крошечных структур, отвечающих в клетках водорослей и растений за фотосинтез. Предки хлоропластов, жившие миллиард лет назад, были свободноживущими фотосинтетическими бактериями — цианобактериями, которых впоследствии поглотила более крупная сложная клетка (см. главу 3). А это значит, что глазное пятно не должно быть уникальной особенностью клеток вольвокса и вполне может быть свойственно и другим хлоропластам, а может быть, и их предкам — цианобактериям11. А хлоропласты можно найти и во многих других клетках, в том числе у некоторых простейших, к которым действительно принадлежали предки животных.
Простейшие — это группа одноклеточных организмов, среди которых особенно известны амебы. Голландец Антони ван Левенгук, один из первых исследователей микромира, первым увидел простейших (и свои собственные сперматозоиды) и назвал их ярким термином “анималькули” (“крошечные животные"), противопоставив их микроскопическим водорослям, которые он относил к растениям, зеленым и неподвижным. Но это радикальное подразделение на животных и растений глубоко несовершенно: если мы увеличим некоторых из “крошечных животных” до наших собственных размеров, то с ужасом убедимся, что на самом деле эти монстры представляют полуживотных-полурастения, напоминающие фантастических существ Арчимбольдо. Строго говоря, некоторые подвижные простейшие, активно плавающие в погоне за жертвами, обладают хлоропластами, что сближает их с водорослями. И действительно, они обзавелись хлоропластами, как водоросли — поглотив другие клетки. Иногда проглоченные хлоропласты остаются в рабочем состоянии, помогая своим хозяевам удовлетворять их потребность в пище. В других случаях хлоропласты дегенерируют, и тогда об их славном прошлом напоминают только характерные мембраны и гены, или же, как это бывает с разнообразным хламом в руках кустаря, могут лечь в основу нового изобретения, например глаза. Одна из таких микроскопических химер, а не собственно вольвокс, по мнению некоторых ученых (в том числе такого авторитетного, как Вальтер Геринг), могла быть прародительницей всех животных, от которой они унаследовали свои глаза.
Какой была эта крошечная химера? Это неизвестно, однако у нас, возможно, есть ключи к разгадке тайны, и еще многое предстоит выяснить. У некоторых простейших (динофлагеллят) имеются поразительно сложные миниглаза, имеющие аналоги сетчатки, хрусталика и роговицы — все это у одной-единственной клетки. Судя по всему, эти глаза развились из дегенерировавших хлоропластов, и в них тоже используется родопсин. Вопрос, развились ли глаза животных в этом кишащем жизнью и недостаточно изученном микромире напрямую из таких структур или опосредованно (через симбиоз), остается открытым. Было ли это эволюционное событие закономерным, или же оно произошло по прихоти судьбы, трудно сказать. Но такого рода вопросы, одновременно общие и конкретные, и составляют существо науки. Я надеюсь, что поисками ответов на них займутся новые поколения ученых, вдохновляемые мерцающими глазами звезд.
Назад: Глава 6. Движение
Дальше: Глава 8. Теплокровность

