Книга: Лестница жизни: десять величайших изобретений эволюции
Назад: Глава 5. Секс
Дальше: Глава 7. Зрение
Глава 6. Движение
Природы зуб и коготь ал” — это, должно быть, одна из самых цитируемых, к месту и не к месту, аллюзий на Дарвина в англоязычной литературе. Но как бы не к месту ее иногда ни цитировали, эта фраза верно передает если не истинную суть естественного отбора, то, по крайней мере, расхожее его понимание. Это строка из глубокомысленной поэмы Теннисона In memoriam, завершенной в 1850 году, за девять лет до публикации “Происхождения видов”. Теннисон посвятил поэму памяти умершего друга — поэта Артура Халлама. Во фрагменте, из которого взята эта строка, описывается чудовищное противоречие между божественной любовью и безжалостной природой. Природа у Теннисона готова губить не только отдельные существа, но и целые группы: “Пропало видов тьмы и тьмы — Пусть гибнут все до одного!” Но если и правда погибнут все до одного, значит, не останется и всего, что нам дорого: целеустремленности, любви, правды, справедливости, Бога. Хотя Теннисон никогда не терял веру окончательно, временами его, похоже, терзали сомнения.
Этот суровый взгляд на природу, впоследствии отнесенный к неумолимым жерновам естественного отбора, подвергался критике со многих позиций. Если понимать слова Теннисона буквально, здесь налицо пренебрежение, по крайней мере, травоядными, растениями, водорослями, грибами, бактериями и так далее, и жизнь сводится лишь к живописной борьбе между хищниками и их жертвами. А если понимать цитату метафорически — как всеобщую борьбу за существование, о которой так много писал Дарвин, — то, рассуждая об этой борьбе, мы часто не отдаем должное сотрудничеству между организмами или видами и даже между генами одного организма — симбиотическим отношениям в природе. Я не хочу подробно останавливаться здесь на сотрудничестве и предлагаю понимать процитированную строчку буквально и обсудить значение хищничества, особенно вопроса о том, как способность живых существ самостоятельно двигаться преобразила мир.
Фраза “зуб и коготь ал” уже намекает на движение. Жертву для начала нужно поймать, а это сложно сделать, не двигаясь с места. Но и чтобы просто сомкнуть челюсти, нужно открыть и закрыть рот, приложив определенную силу, а для этого необходимы мышцы. Когти тоже сложно вонзить в тело жертвы, не орудуя ими со свирепостью, питаемой силой мышц. Если мы попытаемся представить себе неподвижного хищника, то мы, наверное, придем к чему-то вроде хищного гриба. Но и ему придется шевелиться, хотя бы для того, чтобы задушить свою жертву гифами и высосать из нее соки. Главное здесь, что жизнь хищника трудно вообразить без движения. Так что подвижность — это более глубокое, фундаментальное изобретение, чем хищничество. Чтобы ловить и поедать жертвы, хищнику (будь то крошечная амеба, передвигающаяся ползком и глотающая добычу целиком, или стремительный изящный гепард) нужно для начала научиться двигаться.
Движение преобразило жизнь на Земле, но с первого взгляда неочевидно, как именно это повлияло на отдельные ее аспекты, от сложности экосистем до скорости и направлений эволюции растений. Об истории этих перемен нам рассказывает, пусть очень кратко, палеонтологическая летопись. Интересно, что палеонтологическая летопись указывает на довольно резкое изменение степеней сложности живой природы, последовавшее за крупнейшим массовым вымиранием в истории планеты, случившимся в конце пермского периода (примерно двести пятьдесят миллионов лет назад), когда вымерло, по некоторым оценкам, около 95 % видов. После того, как это великое вымирание позволило природе начать свою работу сначала, мир изменился до неузнаваемости.
Разумеется, жизнь на Земле была довольно сложна и до конца пермского периода. Сушу населяли гигантские деревья, папоротники, скорпионы, стрекозы, амфибии и рептилии. Моря занимали трилобиты, рыбы, акулы, аммониты, плеченогие, морские лилии (прикрепленные иглокожие, которых после пермского вымирания осталось совсем немного) и кораллы. На первый взгляд может показаться, что одни виды сменили другие, а экосистемы в целом изменились не так уж заметно. Но детальный анализ говорит об обратном.
Сложность экосистемы можно оценить по сравнительной численности видов: если среди них доминируют очень немногие, а остальные немногочисленны, такую экосистему можно считать простой. Но если множество видов успешно сосуществует и сопоставимо друг с другом по численности, то такую экосистему следует признать гораздо более сложной, и взаимодействия между ее многочисленными компонентами будут куда многообразнее. Подсчитывая суммарное число сосуществовавших в то или иное время видов, сохранившихся в палеонтологической летописи, можно сравнивать получаемые “коэффициенты сложности”, и результаты такого сравнения окажутся неожиданными. Судя по всему, сложность в экосистемах накапливалась не постепенно, а резко проявилась после великого пермского вымирания. До того времени (около трехсот миллионов лет назад) морские экосистемы примерно поровну делились на простые и сложные. После сложных систем стало втрое больше, и за прошедшие с тех пор двести пятьдесят миллионов лет это соотношение осталось неизменным. Так что вместо постепенных изменений мы наблюдаем резкий перепад. Почему?
По мнению палеонтолога Питера Вагнера из Музея естественной истории им. Филда в Чикаго, причина кроется в распространении подвижных организмов. Эта перемена превратила океаны из мира преимущественно прикрепленных организмов (как плеченогие или морские лилии, отфильтровывающие свою скудную пищу из воды) в новый, более подвижный мир, где стали преобладать организмы, активно перемещающиеся, пусть медленно (как брюхоногие моллюски, морские ежи или крабы). Разумеется, многие животные были очень подвижны и до вымирания, однако они стали доминировать в морских экосистемах только после него. Почему резкий рывок произошел после пермского массового вымирания, неизвестно, но, возможно, он был связан с большей “амортизацией” против ударов окружающей среды, с которой сопряжен подвижный образ жизни. Любое активно передвигающееся существо нередко сталкивается с резкими изменениями среды, а значит, должно обладать большей физической стойкостью. Возможно, это давало подвижным животным больше шансов пережить радикальные изменения среды, сопровождавшие пермский апокалипсис (подробнее мы обсудим это в главе 8). Фильтраторам нечем было защитить себя от таких ударов, что и обрекло многих из них на вымирание.
Но каковы бы ни были причины этого события, расцвет подвижных существ, последовавший за пермским вымиранием, преобразил жизнь на планете. Подвижность означала, что животные стали гораздо чаще сталкиваться друг с другом, как в прямом, так и в переносном смысле, что, в свою очередь, сделало возможные взаимодействия между ними намного многообразнее. В мире стало не только больше хищничества. Животные стаи чаще питаться разлагающимися остатками, а также рыться в различных субстратах. У животных всегда было достаточно причин для движения, но многообразие стилей жизни, которое принесла с собой подвижность, сделало задачу быть в нужное время в нужном месте особенно актуальной, как, впрочем, и задачу избегать в ненужное время ненужных мео. Иными словами, это придало им целеустремленности.
Но выгоды подвижности не ограничиваются образом жизни: она диктует и скорость эволюции, то есть скорость, с которой гены, равно как и виды, меняются с течением времени. Хотя быстрее всех эволюционируют паразиты и болезнетворные бактерии, которым приходится иметь дело с бесконечно изобретательной и жестокой травлей со стороны иммунной системы, многие свободноживущие животные не слишком отстают от них. Закрепленные на одном месте фильтраторы и неподвижные растения в целом эволюционируют гораздо медленнее. Идея Черной Королевы, которая должна бежать, чтобы оставаться на одном месте, по крайней мере относительно соперников, едва ли не чужда миру этих организмов, которые остаются почти неизменными на протяжении многих геологических эпох, пока какое-нибудь внезапное бедствие не приводит к их вымиранию. Но из этого общего правила есть важное исключение, которое только подчеркивает значение подвижности: цветковые растения.
До пермского вымирания цветковые растения отсутствовали. Растительный мир был сплошь зеленым, как современный хвойный лес. Яркие краски, которыми заиграли цветы и плоды, стали лишь реакцией растений на изменения в животном мире. Цветы, как известно, привлекают опылителей — подвижных животных, переносящих пыльцу с цветка на цветок и тем самым дающих прикрепленным растениям возможность сполна наслаждаться выгодами секса. Яркие и вкусные плоды тоже взывают к животным, разносящим семена. В результате животные и цветковые растения стали эволюционировать совместно. Те и другие сделались незаменимы друг для друга: растения удовлетворяли важнейшие потребности опылителей и плодоядных, а животные невольно осуществляли тайные замыслы растений — по крайней мере, до тех пор, пока мы, люди, не вывели растения с бессемянными плодами. Это переплетение судеб ускорило эволюцию цветковых, сделав ее темпы сравнимыми с темпами эволюции животных, от которых они зависят.
Итак, подвижность связана с необходимостью иметь дело с быстро меняющейся средой, она подразумевает более тесное взаимодействие растений и животных, новые образы жизни, такие как хищничество, и более сложные экосистемы. Все эти факторы способствовали развитию органов чувств (позволяющих успешнее “прощупывать” окружающий мир) и ускорению эволюции (просто чтобы не отставать от других) не только животных, но и многих растений. В основе всех подобных новшеств лежит лишь одно изобретение: мышцы. На первый взгляд, мышцы не кажутся такими же совершенными органами, как глаза, но если рассмотреть их в микроскоп, нам откроется поразительная совокупность слаженно работающих волокон, рождающая силу и производящая впечатление глубокого замысла. Эти миниатюрные устройства превращают химическую энергию в механическую и достойны самых удивительных изобретений Леонардо да Винчи. Как возникли эти замысловатые устройства? В этой главе мы рассмотрим происхождение и эволюцию молекулярной аппаратуры, обеспечивающей мышечные сокращения, которая позволила животным вызвать перечисленные выше огромные изменения в жизни нашей планеты.
Мало какие свойства производят на нас такое же неизгладимое впечатление, как сильные мышцы. Мускулистые мужчины всегда, со времен Ахиллеса до известного “губернатора-терминатора”, вызывали вожделение и зависть. Но внешность — это еще не все. Параллельно с историей культа мускулатуры продолжались и попытки великих мыслителей и экспериментаторов разобраться в механизме ее работы. Со времен Аристотеля и до Декарта считалось, что мышцы не столько сокращаются, сколько раздуваются, подобно самомнению мускулистых людей. Предполагалось, что от желудочков мозга к мышечным волокнам поступает по полым нервам разряд невидимой и невесомой жизненной силы, отчего мышцы и раздуваются, укорачиваясь. Декарт, придерживавшийся механистических представлений о физиологии, предполагал существование в мышцах крошечных клапанов, не позволяющих жизненной силе утекать, подобно тому, как клапаны в кровеносных сосудах якобы не дают крови течь в обратном направлении.
Однако вскоре после смерти Декарта, в 60-х годах XVII века, было сделано открытие, которое разом перевернуло давно сложившиеся представления. Голландский экспериментатор Ян Сваммердам продемонстрировал, что объем мышц при сокращении не увеличивается, а, напротив, даже немного уменьшается. А раз так, мышцы едва ли могли сокращаться оттого, что раздувались от жизненной силы, как мочевой пузырь от мочи. В 70-х годах XVII века другой голландец — один из первых исследователей микромира Антони ван Левенгук - воспользовался увеличительными стеклами (он сам их великолепно делал), чтобы рассмотреть микроскопическую структуру мышечной ткани. Левенгук описал тонкие волокна, состоящие из “очень маленьких соединенных шариков”, образующих натянутые цепочки. Из тысяч цепочек таких шариков оказались сложены мышцы. Англичанин Уильям Крун предположил, что эти шарики могут быть микроскопическими пузырьками, меняющими форму мышцы, не увеличивая ее общего объема1. Как именно это могло происходить, было сложно проверить экспериментально, но вполне доступно воображению. Несколько ведущих ученых предположили, что при наполнении этих пузырьков происходят в буквальном смысле взрывы. Например, Джон Мейоу предположил, что жизненная сила, поступающая в мышцы из нервов, представляет собой “азотно-воздушные частицы”. Эти частицы, смешиваясь с серными частицами крови, вызывают взрывы, аналогичные пороховым.
Но долго эти версии не продержались. Спустя восемь лет после своего первого исследования мышц Левенгук вновь рассмотрел открытые им “шарики” с помощью нового, более качественного увеличительного стекла. Мышечные волокна оказались вовсе не длинными цепочками крошечных пузырьков: это были именно волокна, разделенные на сегменты упорядоченными “кольцами и морщинами”, и именно эта поперечная полосатость создавала впечатление “шариков”. Более того, раздавив мышечные волокна и исследовав их содержимое под увеличительным стеклом, Левенгук понял, что они, в свою очередь, наполнены нитями еще меньшего размера, которых в каждом волокне около сотни. Сейчас описанные Левенгуком сегменты называют саркомерами, а заключенные в них нити — миофибриллами. Стало ясно, что сокращение мышц не имеет отношения к надувающимся пузырькам и связано исключительно с волокнами.
И все же, хотя ученые и высказали предположение, что мышечные волокна могут каким-то образом “скользить” друг по другу, они по-прежнему совершенно не представляли себе, какая сила заставляет эти волокна двигаться. Прошло почти сто лет, пока не была найдена сила, которая могла претендовать на эту роль, — электричество.
В 1780 году Луиджи Гальвани, профессор анатомии из Болонского университета, был поражен, увидев, как мышца в ноге мертвой лягушки резко сократилась от прикосновения скальпеля, когда в электрической машине на другом конце комнаты пробежала искра. Такой же реакции ему удалось добиться, потерев в ходе препаровки скальпель о медный крючок, а также в ряде других случаев, в том числе во время грозы. Эффект оживления электричеством, вскоре названный гальванизмом, произвел большое впечатление на Мэри Шелли, которая читала Гальвани незадолго до того, как в 1823 году написала готический роман “Франкенштейн”. Более того, одним из прототипов доктора Франкенштейна послужил племянник самого Гальвани — Джованни Альдини. В начале XIX века он ездил по Европе, демонстрируя “гальваническое оживление мертвых”. В ходе одной из самых известных демонстраций, проведенной в Английском королевском колледже хирургов, Альдини на глазах публики, среди которой были не только хирурги, но и герцоги и даже принц Уэльский, пропускал электрический ток через отрубленную голову преступника. По воспоминаниям самого Альдини, когда он приложил электроды ко рту и уху, “челюсть затряслась, прилежащие мышцы жутко скривились, а левый глаз даже открылся”.
Физик Алессандро Вольта из Университета Павии тоже находился под впечатлением от открытия Гальвани, но не соглашался с последним относительно причин этого явления. Вольта настаивал, что в самом организме нет никакого электричества и гальванизм представляет собой лишь реакцию на внешнее раздражение электрическими зарядами, вырабатываемыми металлами. Он утверждал, что хотя нога лягушки и может проводить электричество, как проводит его соляной раствор, это лишь пассивное качество. Гальвани и Вольта вступили в спор, который длился десять лет и в ходе которого их сторонники в лучших итальянских традициях разделились на два лагеря: приверженцев анимализма и металлизма, физиологии и физики, Болоньи и Павии.
Гальвани был убежден, что “животное электричество” действительно вырабатывается в живых организмах, но ему трудно было это доказать — по крайней мере, так, чтобы убедить в этом Вольту. Их спор может служить прекрасной иллюстрацией того, как сила скептицизма гальванизирует научное мышление. Придумывая всевозможные эксперименты, Гальвани установил, что мышцам от природы свойственна раздражимость (он сам это так назвал) — способность совершенно непропорционально реагировать на стимулы. Он даже предположил, что электричество может вырабатываться в самих мышцах за счет накопления положительных и отрицательных зарядов на внутренних поверхностях волокон. По его словам, электрический ток протекает при этом через поры, открывающиеся между двумя поверхностями.
Это была гениальная догадка, но, к сожалению, случай Гальвани может служить иллюстрацией еще и того, что история пишется победителями — даже в науке2. Когда Гальвани отказался присягать Наполеону, войска которого в то время оккупировали Италию, он был изгнан из Болонского университета и в следующем же году умер в бедности. Его идеи были позабыты на десятки лет, а его самого долгое время помнили в основном как адепта оккультных идей “оживления электричеством” и оппонента Вольты. Самому же Вольте Наполеон в 1810 году пожаловал титул ломбардского графа, а впоследствии в его честь была названа единица измерения электрического напряжения — вольт. И все же, хотя Вольта по праву вошел в историю науки как изобретатель первой настоящей электрической батареи (“вольтова столба”), его представления о животном электричестве были глубоко ошибочны.
Идеи Гальвани снова оказались востребованы лишь в середине XIX века, особенно среди представителей немецкой школы биофизиков, самым известным из которых был великий Герман фон Гельмгольц. Ученые этой школы не только доказали, что мышцы и нервы действительно работают за счет “животного” электричества: Гельмгольц даже рассчитал скорость, с которой электрический импульс передается по нерву. Ученый воспользовался для этого методом, разработанным военными для определения скорости пушечного ядра. Оказалось, что нервные импульсы передаются на удивление медленно, со скоростью всего несколько десятков метров, а не сотни километров в секунду, как обычный электрический ток. Это открытие заставило предположить, что животное электричество какое-то особенное. Как вскоре удалось выяснить, особенность его состояла в том, что оно передается не неуловимыми электронами, которые носятся по проводам, а неповоротливыми заряженными атомами (ионами) калия, натрия и кальция, движущимися в поперечном направлении. Прохождение ионов сквозь мембрану внутрь клетки вызывает волну деполяризации, то есть временного сдвига в сторону большего положительного заряда внутри клетки. Деполяризация действует на соседние участки мембраны, которые тоже начинают пропускать ионы внутрь клетки, и в итоге по нерву или мышце прокатывается так называемый потенциал действия.
Как потенциал действия вызывает сокращение мышц? Чтобы ответить на этот вопрос, для начала нужно было ответить на другой, более общий: каков физический механизм сокращения мышц? Здесь снова помогли достижения микроскопии, позволившие обнаружить в мышечных волокнах упорядоченные полоски, которые, по-видимому, соответствовали материалам разной плотности. С конца 30-х годов XIX века английский хирург и анатом Уильям Боумен тщательно исследовал микроскопическое строение скелетных мышц сорока с лишним видов животных, в том числе человека и ряда других млекопитающих, а также птиц, рептилий, амфибий, рыб, ракообразных и насекомых. И у всех у них мышцы были расчерчены на сегменты — те самые саркомеры, которые еще 160 лет назад описал Левенгук. Но Боумен заметил, что внутри каждого саркомера тоже расположены чередующиеся темные и светлые полоски. При сокращении мышцы саркомеры укорачиваются, и светлые полоски исчезают, вызывая по словам Боумена, “темную волну сокращения”. Он сделал из этого вывод (совершенно правильный), что “сократимость присуща отдельным сегментам”.
Однако впоследствии Боумен отступился от собственного открытия. Он видел, что нервы, заходящие в мышцы, напрямую вовсе не взаимодействуют с саркомерами, поэтому электрический запуск работы последних должен был осуществляться по меньшей мере опосредованно. Особенно его беспокоили гладкие мышцы в сфинктерах и стенках артерий. Они не разделены на полоски, как скелетные мышцы, и все же прекрасно сокращаются. Боумен решил, что полоски не имеют непосредственного отношения к сокращению мышц и что тайна сократимости кроется в невидимой структуре молекул, которая навсегда останется “за пределами доступного пониманию”. Он оказался прав относительно значения молекулярного строения мышечных волокон, однако ошибся относительно полосок, как и пределов понимания. Но в то время с оговорками Боумена по вопросу о строении мышц соглашались почти все исследователи.

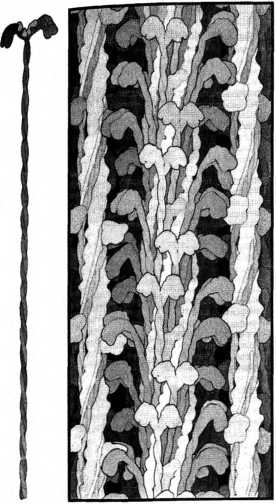
Строение скелетной мышцы, расчерченной на характерные сегменты (саркомеры) и поперечные полоски. Один саркомер занимает участок от одной узкой темной полоски (7-диска) до другой. Самые темные участии саркомера (Л-полосы) содержат миозин, связанный с актином, светлые участки (/-полосы) — актин, а участки промежуточного серого цвета — нити миозина, закрепленные на /И-линии. Когда мышцы сокращаются, миозиновые мостики подтягивают актин из /-полос к /И-линии, укорачивая саркомер и создавая “темную волну” сокращения (где /-полосы поглощаются Л-полосами).
В некотором роде ученые викторианской эпохи уже знали все, но еще не знали ничего. Они знали, например, что мышцы состоят из тысяч волокон, каждое из которых разделено на сегменты (саркомеры), и что эти сегменты представляют собой элементарные единицы сокращения. Они знали, что саркомеры расчерчены на поперечные полосы, соответствующие материалам разной плотности. Некоторые ученые по крайней мере подозревали, что эти полосы состоят из нитей, скользящих друг по другу. Они также знали, что сокращение мышц имеет электрическую природу, что электричество при этом вырабатывается как разность потенциалов между двумя поверхностями, и даже правильно предположили, что ключевую роль здесь может играть кальций. Они выделили главный мышечный белок и назвали его миозин (от греческого слова, означающего “мышцы”). Но глубокие молекулярные тайны, которые Боумен объявил недоступными пониманию, и вправду были недоступны ученым того времени. Они кое-что знали о составляющих механизма, но ничего не знали о том, как эти детали соединены, а также как работает этот механизм. Разобраться в этом удалось только в XX веке, когда виртуозное применение редукционизма существенно расширило горизонты науки. Чтобы оценить подлинное великолепие мускулов и эволюции их компонентов, нам придется оставить викторианских ученых и обратиться к тем, кто занимался уже собственно молекулами.
Кембридж, 1950 год. Недавно открытое в Кавендишской лаборатории отделение структурной биологии. Поворотный момент в истории науки. Два физика и два химика пытаются усовершенствовать метод, которому суждено преобразить биологию второй половины XX века, — рентгеноструктурный анализ. Даже применительно к однородным, геометрически правильным кристаллам этот метод довольно сложен. Применительно же к свернутым сложным образом большим биологическим молекулам он и сегодня требует математических расчетов, напоминающих о черной магии.
Возглавлял отделение Макс Перуц. Он и его заместитель Джон Кендрю первыми определили структуру больших белков, таких как гемоглобин и миоглобин, причем исключительно по узорам, образуемым рентгеновскими лучами, которые при прохождении сквозь кристалл белка рассеиваются, попадая на атомы, соединенные в сложные колеблющиеся цепочки3. Фрэнсис Крик, к которому вскоре присоединится молодой американец Джеймс Уотсон, применял тот же метод, пытаясь разобраться в структуре ДНК. Но в 1950 году четвертым сотрудником отделения был не Уотсон, а человек сравнительно малоизвестный, по крайней мере для неспециалистов, и единственный из этой команды, кто не получил впоследствии Нобелевской премии. Но Хью Хаксли, несомненно, был ее достоин, потому что он сделал больше, чем кто-либо, для того, чтобы разобраться, как работают мышцы на уровне молекулярных рычагов, и на протяжении полувека совершал открытия в этой области. Хорошо, что хотя бы Лондонское королевское общество в 1997 году удостоило его самой высокой своей награды — медали Копли. Сейчас он заслуженный профессор в Университете Брандейса в штате Массачусетс и по-прежнему публикуется, хотя ему уже восемьдесят три года.
Должно быть, одна из причин, почему Хью Хаксли не так известен, связана с тем, что его путают со знаменитым однофамильцем — нобелевским лауреатом Эндрю Хаксли, внуком “бульдога Дарвина” — неистового и красноречивого Томаса Генри Хаксли (Гекели). Эндрю Хаксли прославился в послевоенные годы своими исследованиями передачи нервных импульсов, после чего, в начале 50-х годов, обратился к изучению мышц и в последующие годы по праву считался одним из ведущих специалистов в этой области. Оба Хаксли, работая независимо, пришли к одному и том же выводу и в 1954 году договорились о том, чтобы их результаты были опубликованы в одном и том же номере журнала “Нейчур”. Идея, которую они оба высказали, стала известна как теория скользящих нитей. Хью Хаксли (которому в ту пору было всего двадцать) сыграл особенно важную роль в развитии этой теории, используя замечательные возможности таких методов, как рентгеноструктурный анализ и электронная микроскопия. Эта комбинация оказалась необычайно удачной и позволила исследователям в следующие десятилетия узнавать о работе мышц все больше и больше.
Во время войны Хью Хаксли занимался конструированием радаров. Демобилизовавшись, он вернулся на учебу в Кембридж. Как и многие физики того поколения, Хаксли, узнав об ужасных возможностях атомного оружия, решил оставить физику и заняться чем-то не столь пугающим и не столь спорным в моральном плане. Это стало потерей для физики, но большим приобретением для биологии. В 1948 году Хаксли вошел в состав маленькой исследовательской группы Перуца. С удивлением узнав, как мало известно биологам о строении и работе мышц, он посвятил всю свою оставшуюся жизнь заполнению этого пробела. Поначалу он, как некогда Гальвани, работал с мышцами ног лягушек, но первые полученные им результаты принесли разочарование. Рентгенограммы мышечных волокон, добытых из содержавшихся в лаборатории лягушек, оказались слишком нечеткими. Однако вскоре Хаксли обнаружил, что с дикими лягушками результат получается гораздо лучше. В итоге ему не раз пришлось по утрам, еще до завтрака, ездить на велосипеде по холодку на болото за лягушками. Дикие лягушки позволили ему получить гораздо более подробные рентгенограммы, которые, тем не менее, можно было интерпретировать по-разному. По иронии судьбы в 1952 году, сдавая экзамен на получение докторской степени, Хаксли познакомился с Дороти Ходжкин — выдающимся кристаллографом и одним из первых специалистов по рентгеноструктурному анализу. Прочитав диссертацию Хаксли, она сразу решила, что полученные результаты могут указывать на скользящие нити, и стала увлеченно обсуждать эту идею с Фрэнсисом Криком, с которым столкнулась на лестнице. Но Хаксли со свойственной юности воинственностью принялся доказывать, что она недостаточно внимательно прочитала раздел его диссертации, посвященный методам, и что полученные им данные не подтверждают ее предположений. Однако два года спустя, с помощью электронно-микроскопических изображений, ему удалось самому прийти к похожим выводам, на этот раз действительно получившим убедительное экспериментальное подтверждение.
Но хотя Хаксли и отказался делать преждевременные выводы, отодвинув на два года свое открытие скользящих нитей, он довольно рано уверился в том, что разобраться в молекулярных подробностях механизма сокращения мышц ему позволит рентгеноструктурный анализ в сочетании с электронной микроскопией. Оба метода были несовершенны. Вот что писал об этом сам Хаксли: “Электронная микроскопия давала изображения вполне отчетливые, но содержавшие всевозможные артефакты, в то время как дифракция рентгеновских лучей давала настоящие данные, но в загадочной форме”. Замечательным было то, что, как понял Хаксли, первый метод позволял преодолеть недостатки второго, а второй — недостатки первого, Отчасти успех Хаксли объяснялся удачей, потому что в то время никто еще не мог предвидеть замечательных достижений второй половины столетия — особенно в области рентгеноструктурного анализа. Главную трудность здесь составляла сила луча. Чтобы получить информативную рентгенограмму той или иной структуры, отражающую рассеивание (дифракцию) проходящих сквозь нее рентгеновских лучей, таких лучей должно быть очень много. А на это требуется время (в 50-х годах, когда Хаксли и другим исследователям приходилось работать по ночам, охлаждая слабые рентгеновские аппараты, на это уходил не один час, а иногда не один день) либо исключительно сильный источник, позволяющий сразу получать мощный поток рентгеновских лучей. Работа биологов, как это часто бывало, зависела от достижений физики, особенно от разработок синхротронов — огромных кольцевых ускорителей субатомных частиц, в которых синхронизированные магнитное и электрическое поля используются для ускорения до астрономических скоростей и столкновения друг с другом протонов или электронов. Для биологов синхротрон ценен именно тем эффектом, который у физиков вызывает досаду. Когда субатомные частицы носятся в синхротроне по кругу, они испускают электромагнитное излучение (“синхротронный свет”), значительная доля которого приходится на диапазон рентгеновских лучей. Эти потрясающе мощные лучи позволяют за ничтожные доли секунды получать дифракционные картины, на получение которых в 50-х годах обычно уходило несколько часов или дней. А это имело принципиальное значение, потому что молекулярные механизмы, обеспечивающие сокращение мышц, срабатывают за сотые доли секунды. Поэтому исследовать изменения, происходящие в молекулярной структуре мышечных волокон во время их сокращения, имело смысл только с помощью синхротронного света.
Когда Хаксли впервые сформулировал свою концепцию скользящих нитей, она была еще только гипотезой, основанной на довольно невнятных данных. Однако с тех пор, благодаря усовершенствованию тех же методов, которые теперь позволяют за доли секунды получать картины с атомарным разрешением, Хаксли и другим исследователям удалось доказать справедливость многих его предположений о точном молекулярном механизме мышечных сокращений. Там, где ученые викторианской эпохи могли наблюдать лишь довольно грубые микроскопические структуры, Хаксли удалось разглядеть тонкие молекулярные подробности и постулировать вероятный механизм. Теперь, хотя кое-какие неясности еще остаются, мы знаем, как именно сокращаются мышцы, с точностью почти до отдельных атомов.
Сокращение мышц обусловлено свойствами двух веществ: актина и миозина. Оба они представлены многочисленными одинаковыми белковыми субъединицами, образующими длинные нити (полимеры). Толстые нити состоят из миозина, названного так еще в викторианскую эпоху, тонкие — из актина. Нити этих двух типов, толстые и тонкие, идут параллельно друг другу в пучках, где они соединены перпендикулярными мостиками (которые в 50-х годах Хаксли первым увидел в электронный микроскоп). Эти мостики не остаются неподвижными, а совершают взмахи — и с каждым взмахом немного сдвигают актиновые нити относительно миозиновых, как викинги, гребущие на своей ладье, подгоняя ее бег по волнам. Причем сходство с ладьей этим не ограничивается: взмахи весел здесь тоже не упорядочены и не подчиняются единому ритму. В электронный микроскоп можно увидеть, что из многих тысяч мостиков меньше половины движутся в унисон, а большинство “гребут” вразнобой. Однако расчеты показывают, что крошечные взмахи этих мостиков, даже машущих неслаженно, вместе дают достаточную силу, чтобы их работой полностью объяснялось сокращение мышц.
Все эти машущие мостики торчат из толстых нитей и входят в состав миозиновых субъединиц. По молекулярным меркам молекулы миозина огромны — они в восемь раз больше, чем белки средних размеров, например гемоглобин. Каждая молекула миозина похожа по форме на сперматозоид — точнее, на два сперматозоида, головки которых торчат рядом, а хвосты переплетены так, что не развести. Каждая толстая нить состоит из ступенчато расположенных молекул, переплетенных в ней, как волокна в канате. Из этого каната во все стороны равномерно торчат головки. Они-то и образуют машущие мостики, взаимодействующие с актиновыми нитями.
Миозиновые мостики работают так. Мостик связывается с актиновой нитью, а прикрепившись к ней, связывается с молекулой АТФ, которая обеспечивает весь процесс энергией. Сразу после этого мостик отделяется от актиновой нити. Теперь, освободившись, он совершает взмах (вокруг гибкого участка - “шейки”), описывая угол около 70°, а затем вновь связывается с актиновой нитью. Когда это происходит, остатки использованной молекулы АТФ высвобождаются, и мостик упруго возвращается в исходное положение, при этом работая как рычаг и протягивая тонкую нить вдоль толстой. Весь цикл (отделение, взмах, связывание, протягивание) действительно аналогичен работе весла, и каждый его повтор продвигает тонкую нить вдоль толстой на несколько миллионных долей миллиметра. Молекула АТФ играет здесь ключевую роль. Без нее головка не может отделиться от актина и совершить взмах, и мышцы коченеют, как это и происходит, например, при трупном окоченении, возникающем из-за отсутствия притока АТФ. (Где-то через сутки после смерти трупное окоченение начинает ослабевать из-за разложения мышечной ткани.)
Акварельные рисунки Дэвида Гудселла, изображающие миозин. Слева: отдельная молекула миозина с двумя головками вверху, от которых отходят два сплетенных друг с другом хвоста. Справа: плотная миозиновая нить, образованная переплетенными, как волокна в канате, хвостами, из которой во все стороны торчат головки, взаимодействующие с актином.
Существует множество разновидностей миозиновых мостиков. Они в целом похожи друг на друга, но различаются скоростью работы. Вместе они составляют большое “надсемейство”, включающее тысячи членов. Лишь у людей имеется около сорока разновидностей таких мостиков. Скорость и сила мышечных сокращений зависят от скорости работы миозина: быстрые молекулы миозина быстро расщепляют молекулы АТФ и осуществляют цикл сокращения, медленные молекулы — медленно. В каждом организме есть мышцы нескольких типов, и каждому из них свойственны своя разновидность миозина и своя скорость сокращения4. Подобные различия отмечаются и между видами. Самые быстрые миозины — в летательных мышцах насекомых, таких как плодовая мушка дрозофила. Циклы их работы совершаются с частотой несколько сотен раз в секунду — почти на порядок выше, чем у большинства млекопитающих. У маленьких животных, как правило, более быстрые миозины, так что мышиные мышцы сокращаются примерно втрое быстрее, чем человеческие, а крысиные — вдвое быстрее. Самые медленные миозины работают в исключительно медлительных мышцах ленивцев и сухопутных черепах. Их миозины расщепляют АТФ со скоростью раз в двадцать медленнее, чем человеческие.
Хотя скорость сокращения наших мышц диктуется скоростью расщепления миозином АТФ, они устают вовсе не оттого, что в них заканчивается АТФ. Если бы это было так, после занятий в спортзале у нас всегда наступало бы окоченение, похожее на трупное, и нас приходилось бы разносить по домам на носилках. Утомление мышц наступает задолго до того, как весь АТФ в них может израсходоваться. Считается, что этот механизм выработался как адаптация, оберегающая нас от окоченения. Начало и конец мышечных сокращений определяются уровнем кальция в клетках. Именно это и связывает сокращение мышц с “животным электричеством” Гальвани. Когда к мышце приходит возбуждающий импульс, он быстро распространяется по сети внутриклеточных трубочек, из которых в клетку поступают ионы кальция. Запуская каскад реакций, которые мы не станем здесь разбирать, кальций вызывает обнажение тех участков актиновых нитей, с которыми связываются миозиновые мостики, и это позволяет мышце сокращаться. Но как только мышечная клетка наполняется кальцием, каналы, по которым он поступает, закрываются и запускаются насосы, закачивающие кальций обратно во внутриклеточные трубочки, где он будет ждать нового сигнала, чтобы запустить следующий цикл сокращения. Когда уровень кальция падает, связывающиеся с миозином участки актиновых нитей снова оказываются закрытыми, мостики больше не могут к ним присоединяться, и сокращения не происходит. После этого естественная упругость саркомеров обеспечивает их скорое возвращение в исходное расслабленное состояние.
Разумеется, это очень упрощенное описание. Загляните в любой учебник, и вы найдете не одну страницу с изложением этих подробностей, где будут описаны белок за белком, каждый из которых играет свою собственную тонкую регуляторную или структурную роль. Биохимия мышц ужасно сложна, но сквозь нее просвечивает простота лежащего в ее основе механизма. Эта простота не только помогает понять суть процесса, но и имеет ключевое значение для эволюции сложных организмов. В различных тканях и у различных видов работает множество разных способов управления связыванием миозина с актином. Все эти биохимические детали сродни пышным украшениям барочной церкви, благодаря которым любая такая церковь может быть архитектурным шедевром, в то же время оставаясь одной из многих барочных церквей. И точно так же, несмотря на все пышные украшения, которыми отличаются разные варианты механизма работы мышц, миозин всегда связывается с актином, причем всегда в одном и том же месте, а АТФ всегда обеспечивает энергией скольжение одних нитей относительно других.
Возьмем, например, гладкие мышцы, способность которых сужать сфинктеры и артерии ставила в тупик Уильяма Боумена и других ученых викторианской эпохи. Гладким мышцам совершенно не свойственна та поперечная полосатость, которой отличаются скелетные мышцы. И все же они тоже сокращаются за счет актина и миозина, только актиновые и миозиновые нити расположены в них довольно неупорядоченно и не создают ни малейшей видимости микроскопического порядка. Взаимодействия актина и миозина здесь тоже упрощены. Приток кальция активирует миозиновые головки напрямую, а не кружным путем, как в скелетных мышцах. В остальном же сокращение гладких мышц похоже на сокращение скелетных. В обоих случаях оно осуществляется за счет миозина, который связывается с актином, проворачивая все тот же цикл, осуществляемый за счет энергии все того же АТФ.
Эта сравнительная простота, казалось бы, говорит о том, что гладкие мышцы представляют собой один из этапов эволюции скелетных мышц. Гладкая мускулатура тоже способна «сокращению и, несмотря на отсутствие сложной микроскопической структуры, работает совсем неплохо. Однако исследования мышечных белков различных видов животных показывают, что эволюция мускулатуры была вовсе не так проста. Одно тщательное исследование, которое провели генетики Оота Сатоси и Сайтоу Наруя из японского Национального института генетики в Мисиме, показало, что набор белков, действующих в скелетных мышцах млекопитающих, так похож на набор белков в поперечнополосатых летательных мышцах насекомых, что они явно унаследованы от общего предка позвоночных и беспозвоночных, жившего около шестисот миллионов лет назад. У этого существа уже были поперечнополосатые мышцы, хотя и не было скелета. То же самое относится и к белкам гладких мышц, эволюция которых восходит к другому давнему общему предку. Гладкая мускулатура не была промежуточным этапом на пути к более сложной поперечнополосатой мускулатуре — это просто другая эволюционная ветвь.
Это весьма примечательный факт. Миозин наших собственных скелетных мышц состоит в более близком родстве с миозином, за счет которого летает назойливая комнатная муха, которая вьется у вас над головой, чем с миозином из ваших собственных сфинктеров, сжимающихся от отвращения. Как ни удивительно, они разделились даже задолго до нашего общего предка и, судя по всему, еще до возникновения билатеральной (двусторонней) симметрии, свойственной как насекомым, так и позвоночным. Похоже, что даже у медуз есть поперечнополосатые мышцы, во многих деталях похожие на наши. Итак, и гладкие, и поперечнополосатые мышцы сокращаются с помощью похожей системы из актина и миозина, ноте и другие, судя по всему, развились независимо и унаследованы нами от общего предка, у которого были оба типа мышечных клеток. Причем этот общий предок был одним из древнейших животных, существовавших в те времена, когда венцом творения были какие-нибудь медузы.
И все же, несмотря на неожиданно продолжительное эволюционное разделение поперечнополосатой и гладкой мускулатуры, ясно, что все бесчисленные формы миозина восходят к одной предковой форме. Все они обладают одной и той же базовой структурой, все они связываются с актином и АТФ в одних и тех же местах, и все они проворачивают один и тот же двигательный цикл. Если миозины поперечнополосатой и гладкой мускулатуры происходят от общего предка, значит, этот предок был еще примитивнее медуз, и у него, по-видимому, не было ни поперечнополосатых, ни гладких мышц. И все же он находил какое-то применение и актину, и миозину. Какое? Ответ на этот вопрос был получен еще в 60-х годах XX века — благодаря неожиданному открытию. Но хотя это открытие было сделано уже довольно давно, его по праву можно считать одним из самых поразительных в истории биологии. Оно удивляет не только своей наглядностью, но и тем, сколько света оно пролило на эволюцию мышц. Совершил это открытие Хью Хаксли. Он обнаружил, что актин можно “украсить” миозиновыми головками и рассматривать в электронный микроскоп. Сейчас я все объясню.
Актиновые и миозиновые нити можно извлечь из мышечных волокон и разделить на составляющие. Например, головки миозина можно отделить от их длинных хвостов и соединить в пробирке с актином. Актин может сам быстро собираться в длинные нити. Способность к полимеризации при соответствующих условиях — одно из его неотъемлемых качеств. К таким нитям миозиновые головки тоже прикрепляются сами, как и в работающих мышцах, выстраиваясь вдоль актиновых нитей как стрелки, которые рисуют на схемах. Все эти стрелки указывают одно направление, отражающее полярность актиновых нитей, всегда собирающихся в одной и той же конфигурации. Миозин тоже всегда связывается с ними в одном и том же направлении, что позволяет создавать мышечную силу. (В саркомерах актиновые волокна, расходящиеся в стороны от середины, имеют противоположную полярность, так что при сокращении оба края саркомера приближаются к середине и весь саркомер сжимается. Такое сокращение множества расположенных в ряд саркомеров приводит к укорачиванию всей мышцы.)
Миозиновые головки связываются только с актином и ни с чем иным, поэтому, добавляя их к содержимому любых клеток, можно проверять, есть ли в них актиновые нити. До 60-х годов считалось, что актин — это специализированный мышечный белок, неизменно присутствующий в мышцах разных видов животных, но отсутствующий в их клетках других типов. Эти общепринятые представления лишь недавно были поставлены под сомнение биохимическими данными, указывавшими на то, что у одного из многих организмов, не имеющих мышц, — у пивных дрожжей, — актин, возможно, все-таки есть, но когда стало известно, что актин можно украшать миозиновыми головками, открылся настоящий ящик Пандоры. Первым его открыл Хаксли. Он добавил кроличий миозин к актиновым нитям, выделенным из слизевика — представителя очень примитивной группы организмов, — и обнаружил, что эти белки прекрасно подходят друг к другу.
Актин есть везде. У всех сложных клеток имеется внутренний скелет (цитоскелет) из актиновых (и других) нитей. Во всех клетках нашего организма, как и во всех клетках любых других животных, а также любых растений, грибов, водорослей и простейших, есть актиновый цитоскелет. И то, что кроличий миозин связывается с актином слизевика, заставляет предположить, что актиновые нити из принципиально разных типов клеток очень похожи по своей тонкой структуре. Это предположение, как ни удивительно, совершенно справедливо: например, теперь нам известно, что генетические последовательности, кодирующие актин у человека и у дрожжей, совпадают на 95 %5. В свете этого эволюция мышц выглядит совсем по-другому. Те же самые нити, которые работают в наших мышцах, работают и во всех сложных клетках. Они по-настоящему отличаются только тем, что по-разному организованы.
Среди всех музыкальных форм мне почему-то особенно дороги вариации. Утверждают, что когда Бетховен в юности демонстрировал свою игру Моцарту, она не произвела на Моцарта особого впечатления, если не считать мастерства импровизации — умения извлекать бесконечные ритмические и мелодические вариации из одной простой темы. В зрелые годы это мастерство достигло у Бетховена апогея в его великих Вариациях на тему Диабелли. Как и предшествовавшие им замечательные баховские Вариации Гольдберга, бетховенские вариации отличаются строгостью формы. Положенная в их основу гармоническая схема сохраняется на протяжении всего произведения, сообщая ему безошибочно ощущаемое единство. В последующие времена композиторы часто отказывались от подобной строгости, что позволяло им задерживаться на различных настроениях и впечатлениях, но лишало их произведения ореола математического совершенства. Такие вариации не создают ощущения, что все скрытые нюансы мелодии в них извлечены на свет, все тайные измерения обрели реальность, все возможности были использованы.
Актиновые нити, добытые из слизевика Physarum polycephalym, украшенные актиновыми “стрелками" из мышц кролика.

Актиновый цитоскелет в клетке хрящевой ткани коровы, помеченный флуоресцентным красителем фаллоидином-ФИТЦ.
Эта способность взять определенную тему и проиграть ее во всевозможных вариациях, неизменно сохраняя верность основным элементам ее структуры, характерна и для живой природы. Например, на такую заданную тему, как двигательные взаимодействия миозина и актина, естественный отбор с бесконечной изобретательностью писал вариации, породив захватывающий дух набор форм и функций. Внутренний мир любой сложной клетки наглядно подтверждает, с какой удивительной легкостью естественному отбору даются строгие вариации.
Взаимодействие двигательных белков с нитями цитоскелета порождает целый мир движений сложных клеток, как внутренних, так и внешних. Многие клетки способны без видимых усилий скользить по твердым поверхностям, не прибегая ни к помощи конечностей, ни к изменениям формы тела. Другие образуют выросты (так называемые ложноножки), которые вытягиваются и подтягивают за собой клетку или помогают ей заглатывать жертв, обволакивая их протоплазмой. У третьих есть реснички или жгутики, которые, ритмично извиваясь, помогают клетке двигаться. Внутри клеток бурлит цитоплазма, поддерживая непрерывную циркуляцию их содержимого. В этом микромире беспрестанно суетятся крупные тельца, такие как митохондрии, и танцуют, прежде чем разойтись к разным полюсам, свой изысканный гавот хромосомы. А вскоре после этого клетка делится надвое, перешнуровываясь посередине, как будто безжалостно перетягивая свою талию. Все эти движения обеспечиваются молекулярным инструментарием, характерные элементы которого составляют актин и миозин. И все эти процессы представляют собой строгие вариации на одну и ту же тему.
Представьте, что вы уменьшились до размеров, скажем, молекулы АТФ и что клетка стала для вас огромным футуристическим городом. Куда ни кинешь взор, везде протянуты замысловатые системы проводов, поддерживаемых другими проводами. Одни кажутся слабыми и тонкими, другие отличаются немалым диаметром. Земное тяготение не играет в этом городе будущего никакой роли: повсюду вязкая среда и беспорядочно трясущиеся атомы. Попытайтесь сдвинуться с места, и вы почувствуете, что застряли, как муха в варенье, в то время как со всех сторон на вас сыплются удары и толчки. Вы вдруг замечаете, как через этот головокружительный город с поразительной скоростью движется необычайная машина, перебирающая механическими руками, которыми она держится за один из множества натянутых проводов. К этой машине с помощью громоздкого соединительного блока прикреплен поистине огромный предмет, который она быстро тянет за собой. Если вы окажетесь у него на пути, вы почувствуете такой удар, будто с вами столкнулась летающая электростанция. На самом деле это и есть электростанция — то есть митохондрия, транспортируемая в другой конец клеточного города, чтобы обеспечить энергией затеваемую там стройку. Теперь вы замечаете, что и другие разнообразные объекты движутся в том же направлении: одни быстрее, другие медленнее, но все их тянут, держась за натянутые в небе провода, похожие машины. Когда митохондрия со свистом проносится мимо, вас подхватывает создаваемый ею вихрь, и вы тоже начинаете кружиться вместе со всеми. Вы сами становитесь частью перемешивающего содержимое любой сложной клетки непрерывного круговорота — цитоплазматического потока.
Все это обеспечивается нанотехнологиями столь сложными, что мы еще только начали в них разбираться, и все же, несмотря на всю странность этого города будущего, его инфраструктура на удивление похожа на инфраструктуру бесчисленного множества других подобных городов. Клетка, внутренности которой я описал, могла оказаться одной из клеток вашего собственного организма, но с тем же успехом могла быть и клеткой растения, или гриба, или одноклеточного простейшего, плавающего в пруду возле дома. Миру клеток свойственно изумительное единообразие, создающее глубокое ощущение взаимосвязанности и братства живой природы. С точки зрения клетки вы представляете собой лишь очередную вариацию на тему плана строения, еще один способ построить нечто замечательное из похожих кирпичиков. Но каких кирпичиков! У всех эукариот (организмов, состоящих из сложных клеток, наделенных ядром; см. главу 4) каждый из таких кирпичиков выглядит как процветающий мегаполис, в отличие от гораздо проще устроенных клеток бактерий. И в немалой степени эта разница определяется богатствами цитоскелета и постоянно осуществляемой им транспортировки, благодаря которой содержимое клетки непрерывно развозится по ее внутренностям. Без этого неиссякаемого транспортного потока клеточный город не смог бы существовать, как не смогли бы существовать и наши прекрасные города без их оживленных магистралей.
Весь внутриклеточный транспорт осуществляется с помощью белковых моторов. Первый из них — миозин, скользящий вдоль актиновых нитей точно так же, как он делает это в мышцах. А дальше начинаются вариации. В мышцах миозиновые головки почти постоянно отделены от актиновых нитей. Если бы они не были отделены, а оставались связанными, это физически мешало бы другим головкам совершать взмахи. Такая система была бы похожа на лодку, гребцы которой отказываются поднимать весла из воды. В мышцах механизм с отделенными большую часть времени миозиновыми головками прекрасно работает, потому что сплетенные друг с другом длинные хвосты, на которых закреплены эти головки, удерживают их рядом с актиновыми нитями. Но с актиновыми проводами, пересекающими клетку вдоль и поперек, осуществлять работу подобного механизма было бы гораздо сложнее. Как только двигательные головки отделялись бы от актиновой нити, их начинало бы дергать из стороны в сторону, и им трудно было бы вновь ухватиться за провод (хотя в ряде случаев миозиновые головки все же удерживаются возле актиновых проводов за счет электрических взаимодействий).
Лучшим решением этой проблемы будет “процессивный" двигатель, остающийся прикрепленным к актину и в то же время каким-то образом марширующий вдоль актиновой нити, как уличная процессия. Именно это мы и наблюдаем. Несколько небольших изменений в структуре миозина делают из него как раз такой процессивный двигатель, способный перемещаться вдоль актиновой нити, все время за нее держась. Какие это изменения? Одно из них — удлинение шейки. Вспомните, что в мышцах две миозиновые головки торчат рядом, прочно соединенные друг с другом хвостами и шейками, но в остальном, судя по всему, не особенно координируют свою работу. Стоит немного удлинить шейки, и взаимозависимость головок еще ослабнет. Это позволит одной головке оставаться прикрепленной, пока вторая совершает взмах, в результате чего миозиновый двигатель сможет перемещаться вдоль провода, перебирая “руками”6. Другие варианты предполагают соединение трех или даже четырех головок. Также, разумеется, нужно избавиться от хвостов, чтобы миозиновые головки не торчали из толстой нити, а могли свободно “расхаживать” по клетке. И, наконец, к двигательным головкам нужно прикреплять другие предметы. Это происходит за счет “соединительных” белков, каждый из которых соответствует какой-то одной разновидности груза. В итоге мы получаем целое племя процессивных двигателей, способных по актиновым путям развозить грузы в клетке во всех направлениях.
Как возник этот великий парад двигательных белков? В мире бактерий и близко нет ничего подобного. При этом актин и миозин — не единственный двигательный “дуэт” в эукариотических клетках. У двигательных белков другого семейства, так называемых кинезинов, принцип работы примерно такой же, как у миозинов: они тоже перемещаются по проводам цитоскелета, перебирая “руками”. Но кинезины пользуются при этом не актиновыми нитями, а проводами большего калибра — так называемыми микротрубочками, собираемыми из субъединиц еще одного белка — тубулина. Одна из многих функций кинезинов состоит в том, что во время деления клеток они обеспечивают расхождение хромосом к полюсам по веретену из микротрубочек. Существуют и другие разновидности двигательных белков, но мы не будем их разбирать, чтобы не завязнуть в избыточных подробностях.
Для всех этих двигательных белков, как и для их путеводных проводов, известны бактериальные прототипы, хотя родственные связи между теми и другими далеко не всегда очевидны, а бактериальные белки обычно выполняют совсем другую работу7. Здесь тоже помогли методы рентгеноструктурного анализа, позволившие установить родство, которое, может, вообще не удалось бы выявить, пользуясь только генетическими последовательностями.
На уровне подробных генетических последовательностей между двигательными белками двух основных разновидностей, миозинами и кинезинами, нет почти ничего общего. Здесь и там попадаются похожие участки, но долгое время считалось, что такие участки возникли случайно или в результате конвергентной эволюции. Кинезины и миозины и вправду казались классическим примером конвергенции: двумя неродственными группами белков, которые специализировались на выполнении сходных функций и у которых поэтому выработались черты структурного сходства (точно так же, как между крыльями летучих мышей и птиц, возникших независимо и похожих друг на друга оттого, что в них были сходным образом решены одни и те же задачи, связанные с полетом).
Но затем с помощью рентгеноструктурного анализа удалось с атомарным уровнем разрешения установить их трехмерную структуру. Генетическая последовательность дает лишь двумерную последовательность букв — либретто без музыки, - в то время как рентгеноструктурный анализ кристаллов позволяет определить трехмерную форму белка — оперу целиком во всем ее великолепии. Вагнер однажды заметил, что оперная музыка должна вырастать из слов, что слова первичны. Но Вагнера помнят не за одни словесные выражения тевтонских страстей — именно музыка вдохнула жизнь в его произведения, которыми наслаждается уже не одно поколение ценителей. Точно так же и последовательность генов представляет собой Слово природы, но настоящая музыка белков заключена в их форме, и именно она позволяет белкам выживать под давлением естественного отбора. Отбору нет дела до последовательностей генов: его заботят только функции. Хотя гены и определяют функции кодируемых ими белков, они часто делают это именно за счет того, что диктуют форму, в которую белки сворачиваются по правилам, по-прежнему плохо нам понятным. В результате последовательности генов, происходящих от общего предка, могут постепенно расходиться так далеко, что между ними не останется никакого сходства, как и получилось с генами миозинов и кинезинов. Но вырастающая из них музыка белковых шариков никуда не делась, и ее по-прежнему можно выявить с помощью рентгеноструктурного анализа.
Итак, исходя из данных рентгеноструктурного анализа, мы знаем, что миозины и кинезины действительно происходят от одного и того же белка, несмотря на очень слабое сходство кодирующих эти белки генов. Их трехмерное строение обнаруживает множество соответствующих друг другу складок и других структур, вплоть до ключевых аминокислот, сохранившихся в одном и том же положении на одних и тех же местах. Это поразительное достижение отбора: он сохранил строение белковых структур и промежутков между ними неизменными в течение миллиардов лет, хотя сам материал этих структур и даже определяющие его последовательности со временем исказились до неузнаваемости. И все эти структуры показывают, что и миозины, и кинезины родственны более многочисленному семейству белков, которые явно происходят от бактериальных предшественников8. Эти бактериальные белки выполняли (и по-прежнему выполняют) работу, включающую те или иные движения, требующие приложения физической силы, например переходы из одной конформации в другую, но ни один из них не обеспечивал настоящей подвижности. Таким образом, рентгеноструктурный анализ показывает нам строение “костей” белка, как рентгеновский снимок птичьего крыла демонстрирует строение его скелета. И так же, как строение костей и суставов выдает происхождение крыльев, развившихся из конечностей бескрылых рептилий, строение двигательных белков свидетельствует о том, что они явно произошли от бактериальных белков, способных менять конформацию, но не обладавших настоящей подвижностью.
Рентгеноструктурный анализ позволил сделать удивительные открытия и об эволюции цитоскелета — тех самых протянутых по всей клетке актиновых и тубулиновых проводов. Можно задаться вопросом, зачем клетке понадобилось вырабатывать целую сеть таких проводов — скоростных магистралей для двигательных белков, когда в ней еще не было самих этих двигательных белков. Не означало ли это, что эволюция поставила телегу впереди лошади? Нет, если цитоскелет был ценен сам по себе. Ценность цитоскелета определяется его структурными свойствами. Форма всех эукариотических клеток, от длинных и тонких нервных клеток до плоских клеток эндотелия, поддерживается именно нитями цитоскелета, и оказывается, что примерно то же самое относится и к бактериям. Многие поколения биологов приписывали многочисленные формы бактериальных клеток (палочковидную, спиралевидную, серповидную и так далее) окружающей эти клетки жесткой клеточной стенке. Поэтому, когда в середине 90-х годов XX века выяснилось, что у бактерий тоже есть цитоскелет, это стало большой неожиданностью. Бактериальный цитоскелет состоит из тонких нитей, очень похожих на актиновые и тубулиновые. Как мы теперь знаем, эти нити отвечают за поддержание сложной формы бактериальных клеток. (Мутации в генах цитоскелета приводят к тому, что обладающие сложной формой бактериальные клетки превращаются в простые шарики.)
Как и в случае с двигательными белками, генетическое сходство между белками бактериального и эукариотического цитоскелетов невелико. Но сходство их трехмерного строения, установленное лет десять назад с помощью рентгеноструктурного анализа, оказалось еще сильнее, чем у двигательных белков. По сути, бактериальные и эукариотические цитоскелетные белки почти точно накладываются друг на друга, так что одни и те же структуры и промежутки, а также несколько одинаковых ключевых аминокислот полностью совпадают. Ясно, что цитоскелет эукариотических клеток развился из бактериального. При этом эукариотические белки сохранили не только форму, но и функции далеких предшественников. И те, и другие играют общую структурную роль, но в обоих случаях цитоскелет способен на нечто большее, чем обеспечение неподвижной опоры. В отличие от нашего жесткого костного скелета клеточный скелет всегда динамичен, непрерывно меняется и перестраивается, непостоянен и всеобъемлющ, как грозовые облака. Он позволяет прикладывать силу, передвигая хромосомы, разделяя клетки пополам в ходе их удвоения, а также (по крайней мере, у эукариот) формировать наружные выросты и без помощи двигательных белков. Короче говоря, цитоскелет уже сам по себе обладает подвижностью. Как такое могло получиться?
Как актиновые, так и тубулиновые нити состоят из белковых субъединиц, собирающихся в длинные цепочки — полимеры. Эта способность к полимеризации не так уж удивительна: в конце концов, пластмассы тоже представляют собой полимеры, состоящие из субъединиц, образующих длинные молекулярные цепочки. Необычно в цитоскелете то, что его структура находится в состоянии динамического равновесия — переменчивого баланса присоединяющихся и отпадающих субъединиц, полимеризации и деполимеризации. В результате цитоскелет вечно перестраивается, надстраиваясь и снова разбираясь. Однако “строительные блоки” цитоскелета могут присоединяться к другим только с одного конца цепочки (как детали конструктора “Лего”, или — это, может быть, точнее, — как воланы, вложенные один в другой), а отделяться — только с другого. Это и дает цитоскелету возможность создавать механическую силу. И вот почему.
Если скорость добавления субъединиц на одном конце цепочки равна скорости их отделения на другом конце, то полимерная цепочка в целом сохраняет постоянную длину. В этом случае кажется, что она движется в направлении того конца, к которому добавляются субъединицы. Если на пути такой цепочки оказывается тот или иной предмет, она может физически двигать его вперед. В действительности его при этом двигает не сама цепочка. Этот предмет толкают беспорядочные молекулярные силы, но каждый раз, когда между ним и растущим концом цепочки образуется небольшой промежуток, туда может протиснуться и пристроиться к цепочке еще одна субъединица. Тем самым рост цепочки не дает предмету двигаться назад, а беспорядочные толчки двигают его вперед.
Самый наглядный пример работы этого механизма, вероятно, наблюдается при некоторых бактериальных инфекциях, возбудители которых нарушают сборку цитоскелета. Например, листерии, которые могут вызывать менингит у новорожденных, выделяют два или три белка, вместе подчиняющие себе работу цитоскелета зараженной клетки. Это позволяет бактериям активно двигаться внутри клетки за счет актиновых “комет”, собирающихся и разбирающихся за ними. Полагают, что сходный процесс происходит и в клетках самих бактерий, помогая им отделять друг от друга хромосомы и плазмиды (маленькие кольцевые молекулы ДНК) в ходе клеточного деления. Причем нечто похожее происходит и у амеб (а также в некоторых клетках нашей собственной иммунной системы, таких как макрофаги). Клеточные выросты (ложноножки) образуются за счет динамичной сборки и разборки актиновых нитей. Ни в каких сложных двигательных белках просто нет нужды.
Работа динамичного цитоскелета может показаться очень сложной, но биохимик Тим Митчисон из Гарварда утверждает, что это отнюдь не так. В основе этого явления лежат самопроизвольные физические процессы, которые происходят и без особой эволюции. Белки, не играющие вообще никакой структурной роли, могут сами по себе внезапно полимеризоваться, образовывая скелетные структуры, способные к приложению силы, после чего так же быстро разбираться, возвращаясь в исходное состояние. Такое поведение может показаться пугающим. Оно и правда обычно не приводит ни к чему хорошему. Например, при серповидноклеточной анемии, если уровень кислорода падает, то особая форма гемоглобина внезапно поляризуется, образуя своего рода каркас. Этот каркас и придает красным кровяным тельцам серповидную форму, давшую болезни ее название, то есть прикладывает силу и совершает движения. Когда уровень кислорода вновь повышается, этот аномальный цитоскелет разбирается (также самопроизвольно) и красные кровяные тельца опять обретают нормальную дискоидальную форму. Хотя возникающий при этом цитоскелет и не делает ничего хорошего, это настоящий динамичный цитоскелет, способный к приложению физической силы9.
Что-то подобное, должно быть, произошло в очень давние времена и с нормальным цитоскелетом. Субъединицы актина и тубулина происходят от обычных белков, выполняющих в клетке другие функции. Несколько несложных изменений в их структуре, подобных тем, что приводят к образованию аномальной формы гемоглобина, позволили им спонтанно собираться в нити. Однако, в отличие от изменений в гемоглобине при серповидноклеточной анемии, эти изменения должны были сразу оказаться полезными для клетки, поскольку их поддержал естественный отбор. Их польза могла быть и непрямой и даже не связанной с движением. В конце концов, ведь и мутацию, вызывающую серповидноклеточную анемию, естественный отбор поддерживает в тех регионах, где распространена малярия, поскольку обладание единственной копией мутантного гена защищает от этого недуга. Несмотря на то, что аномальный гемоглобин вызывает длительные болезненные приступы (серповидные клетки негибки и закупоривают капилляры), естественный отбор сохранил самопроизвольную сборку вредного цитоскелета, потому что у нее есть ценный, хотя и непрямой побочный эффект — устойчивость к малярии.
Итак, величием подвижности, от его простейших начал до прославленной силы скелетных мышц, мы обязаны горстке белков и бесконечным вариациям на тему механизма их работы. Задача, которую нам еще предстоит решить, состоит в том, чтобы разобраться во всех этих изумительных вариациях и найти исходную тему, тот простой хорал, с которого все началось. Поиски этого “хорала” составляют одно из самых увлекательных и дискуссионных направлений современных исследований, ведь его напевала еще праматерь всех эукариотических клеток, по-видимому, около двух миллиардов лет назад, и отголоски столь давних аккордов теперь не так-то просто расслышать. Как именно древнейшая эукариотическая клетка обрела подвижность, точно неизвестно. Мы не знаем, сыграло ли здесь ключевую роль сотрудничество (симбиоз) между клетками, что долгое время доказывала Линн Маргулис, или же клеточный скелет развился из генов, уже имевшихся у клетки-хозяина. Вероятно, мы сможем пролить больше света на эту проблему, когда отгадаем несколько интересных загадок. Например, у бактерий хромосомы расходятся при делении клетки с помощью актиновых нитей, в то время как перешнуровка, разделяющая материнскую клетку на две дочерние, осуществляется с помощью тубулиновых микротрубочек. При делении эукариотических клеток все происходит наоборот. Здесь “строительные леса” веретена деления, по которым хромосомы расходятся в разные стороны, построены из микротрубочек, а шнуровка, разделяющая клетку, состоит из актина. Когда мы узнаем, как и почему эти белки поменялись ролями, мы получим лучшее представление об истории жизни на Земле.
Но все эти серьезные проблемы, стоящие перед исследователями, на самом деле относятся лишь к деталям общей схемы, которая в целом теперь ясна. Мы знаем, из каких белков возникли белки цитоскелета и двигательные белки, и для обобщенной схемы не так уж важно, достались они эукариотам от симбиотической бактерии или от клетки-хозяина. Обе возможности вполне правдоподобны, и когда мы узнаем ответ на этот вопрос, он в любом случае не пошатнет оснований современной биологии. Один факт не вызывает сомнений. Если когда-то и существовали эукариоты, не способные передвигаться, не имевшие динамичного цитоскелета и двигательных белков, этих источников механической силы, теперь их уже не найти: все они давным-давно вымерли. Предок всех современных эукариот обладал подвижностью, которая предположительно давала ему большие преимущества. Поэтому вполне возможно, что расцвет подвижных организмов не только навсегда изменил степень сложности экосистем, но и способствовал в свое время изменению облика нашей планеты и превращению ее из простого мира, где правили бактерии, в тот богатый и изумительный мир, который мы знаем сейчас.
Назад: Глава 5. Секс
Дальше: Глава 7. Зрение

