Книга: Лестница жизни: десять величайших изобретений эволюции
Назад: Глава 7. Зрение
Дальше: Глава 9. Сознание
Глава 8. Теплокровность
Время мчится, когда поезд я веду”, — так поется в детской песенке. Но кто не испытывал в детстве и обратного ощущения, умирая от скуки на заднем сиденье и поминутно спрашивая: “Пап, мы уже приехали?” Думаю, большинство читателей вспомнят, как тяжело смотреть на дедушек и бабушек или родителей, постепенно переходящих на черепаший шаг и все чаще сидящих неподвижно, так что наши часы пролетают для них как минуты. Обе крайности очень далеки от нормального темпа нашей жизни, этого анданте здорового взрослого человека.
Не надо быть Эйнштейном, чтобы понять: время относительно. Но строгим физическим законам пространства и времени, которые установил Эйнштейн, в биологии, как всегда, соответствует поистине импрессионистская картина. Как сказал знаменитый острослов Клемент Фрейд, “если бросить курить, пить и любить, на самом деле дольше не проживешь — жизнь просто покажется дольше”1. Но в некотором смысле время действительно летит в детстве и замедляется в старости. Это связано с нашими внутренними настройками, с интенсивностью обмена веществ, с частотой сердцебиения и скоростью кислородного “горения” пищи в клетках. И даже среди здоровых взрослых людей встречаются как гиперактивные, так и медлительные. Большинству из нас свойственно постепенно превращаться из первых во вторых. Скорость, с которой мы замедляемся, а иногда и набираем вес, бывает разной и в значительной степени зависит от интенсивности нашего метаболизма, во многом определяемой врожденными особенностями. Два человека, употребляющие одинаковое количество одной и той же пищи и получающие одинаковую физическую нагрузку, вполне могут с разной скоростью сжигать калории в состоянии покоя.
Но интенсивность обмена веществ нигде не бывает важнее, чем в определении разницы между теплокровными и холоднокровными существами. Хотя биологов эти термины раздражают, они наглядны и понятны почти каждому и несут столько же смысла, сколько соответствующие им заумные научные термины “гомойотермные” и “пойкилотермные”. Странное дело: я замечал, что некоторые биологические вопросы вызывают у нас, теплокровных, на удивление живой отклик, отдающий шовинизмом. Сколько, например, сломано копий в журналах и в интернете по поводу того, были ли динозавры теплокровными или холоднокровными. Этому сложно найти рациональное объяснение. Вероятно, здесь включаются некие чувства, возможно, связанные с самомнением, которое тешит мысль о том, что наших предков поедали не медлительные гигантские ящеры, а хитрые, коварные, ловкие чудовища, для выживания в борьбе с которыми им пришлось проявить незаурядные умственные способности. Кажется, что мы, млекопитающие, никак не можем простить динозавров за то, что когда-то нашим маленьким мохнатым предкам приходилось прятаться в тесноте подземных нор от царивших тогда хищников. Но ведь это было около ста двадцати миллионов лет назад, то есть по любым меркам очень давно.
Теплокровность — это, прежде всего, интенсивность обмена веществ, которая определяет темп жизни. Но теплая кровь выгодна и сама по себе, потому что при повышении температуры ускоряются все химические реакции, в том числе биохимические, лежащие в основе жизни. В диапазон температур тела большинства животных (примерно от о до 40° С) укладывается огромная разница в интенсивности биологических процессов. Например, потребление кислорода здесь увеличивается вдвое при повышении температуры на каждые 10° С, сопровождаясь резким увеличением выносливости и силы. Так что животное, температура тела которого 37° С, будет вдвое сильнее животного с температурой тела 27° С и вчетверо сильнее животного с температурой тела 17° С.
Но во многом дело все-таки не в температуре. Кровь теплокровных животных не обязательно теплее крови холоднокровных. Например, большинство рептилий прекрасно умеют нагревать свое тело на солнце, поднимая его температуру настолько, что она оказывается сравнимой с температурой тела млекопитающих и птиц. Разумеется, после захода солнца они уже не могут поддерживать такую высокую температуру тела, но ведь и млекопитающие с птицами ночью часто неактивны. Они могли бы тоже экономить энергию, понижая на это время температуру тела, но почему-то редко это делают — по крайней мере, редко понижают ее существенно (хотя колибри и могут ради экономии энергии впадать в кому). Сейчас, когда так много говорят о необходимости бережно расходовать энергию, защитники природы должны с ужасом смотреть на млекопитающих. Наш термостат заело на отметке 37° С, и он работает в этом режиме семь дней в неделю и двадцать четыре часа в сутки независимо от того, есть ли в этом потребность. Кроме того, здесь нет никаких альтернативных источников энергии: мы, в отличие от ящериц, не отапливаем свой организм за счет солнечных батарей, а в избытке вырабатываем тепло, сжигая горючие материалы во внутриклеточных теплоцентралях, попутно выделяя в больших объемах парниковый углекислый газ. Млекопитающие стали вести себя безответственно задолго до появления человека.
Можно подумать, что млекопитающие, не гася печей ночью, получают то преимущество, что они не нуждаются в разогреве по утрам, но у ящериц на доведение температуры тела до рабочего уровня уходит не так уж много времени. Например, у глухих игуан на макушке имеется полость, наполняемая кровью, которая позволяет им быстро нагреть все тело. По утрам они высовывают из нор головы, внимательно глядя по сторонам, чтобы в случае приближения хищника нырнуть обратно, и уже через полчаса согреваются достаточно, чтобы отправляться по делам. Приятное начало дня, не правда ли? При этом естественный отбор, как это часто бывает, не удовлетворился единственной функцией этого полезного приспособления. Некоторые из игуан, столкнувшись с хищником вдали от родной норы, могут брызгать из глаз кровью (полость у них на макушке связана с веками). Собак, например, это отпугивает.
Крупные размеры тоже позволяют животному поддерживать высокую температуру тела. Не нужно быть великим охотником, чтобы вообразить лежащие на полу шкуры двух животных. Представьте себе, что одна из этих шкур вдвое больше другой как в длину, так и в ширину. Это значит, что площадь шкуры большого животного в четыре раза больше, чем площадь шкуры маленького (2 x 2 = 4). Но при этом большое животное весило в восемь раз больше, чем маленькое, потому что было вдвое больше не только в длину и в ширину, но и в глубину (2 X 2 X 2 = 8). А значит, при каждом увеличении размеров вдвое отношение площади поверхности к массе или объему будет уменьшаться вдвое (4/8 = 1/2). Если предположить, что на каждый килограмм тела вырабатывается одно и то же количество тепла, то большое животное сможет вырабатывать намного больше тепла, чем маленькое. При этом большое животное будет и терять тепло намного медленнее, потому что относительная площадь поверхности его тела (по отношению к количеству вырабатываемого тепла) сравнительно мала. Итак, чем крупнее животное, тем легче ему согреваться. Если холоднокровные животные достигают достаточно больших размеров, они становятся теплокровными. Например, огромные аллигаторы считаются холоднокровными, но могут сохранять тепло так долго, что близки к теплокровности. Даже за ночь внутренняя температура их тела падает лишь на несколько градусов, хотя тепла они при этом вырабатывают мало.
Ясно, что многие динозавры успешно преодолели этот размерный порог. Фактически они были теплокровными, особенно если учесть, какой теплый и мягкий климат был на значительной части суши в благословенные времена их процветания. На полюсах в то время не было ледяных шапок, а уровень углекислого газа в атмосфере был в десять раз больше, чем сегодня. Иными словами, из некоторых простых физических принципов следует, что многие динозавры в любом случае были теплокровными, какой бы ни была интенсивность обмена веществ в их организме. Вполне вероятно, что гигантским растительноядным динозаврам было гораздо важнее охлаждаться, чем согреваться, и некоторые анатомические диковинки, такие как защитные спинные пластины стегозавра, вполне могли дополнительно использоваться для охлаждения, подобно слоновьим ушам.
Но если все было бы так просто, никто бы не спорил, были динозавры теплокровными или нет. В этом вольном понимании термина они точно были теплокровными — по крайней мере, многие из них. Те, кто любит наукообразные слова, могут называть эту разновидность теплокровности инерциальной зндотермией. Гигантские динозавры не только поддерживали высокую внутреннюю температуру тела, но и вырабатывали внутри тепло, причем точно так же, как это делают современные млекопитающие — сжигая пищу. Так в каком же строгом смысле слова “теплокровный” динозавры не были теплокровными? Некоторые из них, предположим, были теплокровными, в чем нам предстоит убедиться, но чтобы разобраться, чем по-настоящему удивительна теплокровность млекопитающих и птиц, нам придется отвлечься от гигантов и посмотреть, что происходит с животными меньшего размера, не достигающими “порога теплокровности”.
Представьте себе ящерицу. Она по определению холоднокровна — не может поддерживать высокую температуру тела в течение ночи. Самые крупные крокодилы недалеки от этого, но чем меньше животное, тем ему сложнее. Теплоизоляция, например шерсть или перья, помогает лишь до определенной степени, а к тому же может и мешать поглощать тепло из окружающей среды. Оденьте ящерицу в меховую шубку (стоит ли говорить, что некоторые добросовестные исследователи именнотан и делали), и она будет постепенно остывать, не имея возможности вырабатывать внутри тела достаточно тепла, чтобы компенсировать недостаток его притока извне. Но с млекопитающими и птицами ничего подобного не происходит, что и приводит нас к строгому определению теплокровности.
Млекопитающие и птицы вырабатывают в 10-15 Раз больше внутреннего тепла, чем ящерицы тех же размеров, причем вне зависимости от обстоятельств. Поместите ящерицу и млекопитающее в очень жаркую среду, и млекопитающее будет по-прежнему вырабатывать в десять раз больше внутреннего тепла, хотя это и не пойдет ему на пользу. Ему понадобится из кожи вон лезть, чтобы охладиться: пить воду, купаться, пыхтеть, прятаться в тень, обмахиваться веером, делать коктейли со льдом или включать кондиционеры. А ящерица будет только радоваться. Не удивительно, что ящерицы и вообще рептилии в целом гораздо лучше чувствуют себя в пустыне, чем млекопитающие.
А теперь попробуйте поместить ящерицу и млекопитающее в холодную среду, например такую, где температура будет около нуля. Ящерица зароется в листья, свернется клубком и погрузится в сон. Надо признаться, что многие мелкие млекопитающие поступили бы точно так же, но по умолчанию у нас установлены совсем другие настройки. Попадая в холод, мы просто сжигаем больше пищи. Млекопитающему, чтобы выбить в холодной среде, приходится сжигать в сто раз больше пищи, чем сжигает ящерица. Даже при температуре, скажем, около 20° С (во многих европейских странах такая темпера- •ура бывает теплым весенним днем) разница остается огромной, примерно тридцатикратной. Для поддержания столь непомерной интенсивности обмена веществ млекопитающему придется сжигать в тридцать раз больше пищи, чем рептилии, каждый день ему потребуется есть столько, сколько ящерица съедает за месяц. Учитывая, что даром еда не дается, это очень высокая цена.
Итак, цена, которую приходится платить за право быть млекопитающим или птицей, начинается примерно с десятикратной цены за то, чтобы быть ящерицей, а часто оказывается и гораздо выше. Что мы получаем взамен столь расточительного образа жизни? Очевидный ответ — возможность расширения экологических ниш. Хотя теплокровность может и не окупаться в пустыне, она позволяет добывать еду по ночам и сохранять активность зимой в умеренном климате, в то время как ящерицам недоступно ни то, ни другое. Еще одна выгода — умственные способности, хотя на первый взгляд не совсем ясно, почему они должны быть связаны с теплокровностью. У млекопитающих головной мозг и вправду крупнее (относительно размеров тела), нему рептилий. Крупный мозг не гарантирует ум и даже сообразительность, но, судя по всему, более интенсивный обмен веществ действительно сопряжен с поддержанием работы более крупного мозга. Ресурсов специально на него обычно не выделяется, но если и ящерица, и млекопитающее направляют, предположим, 3 % своих ресурсов на работу мозга, то у млекопитающего этот показатель окажется раз в десять выше, а, значит, оно сможет позволить себе в десять раз больше мозгов (обычно именно так и бывает). Однако у приматов, и особенно у людей, на работу мозга выделяется намного больше ресурсов. Например, у человека этот показатель составляет около 20 %, хотя наш мозг и составляет лишь несколько процентов от массы тела. Подозреваю, что умственные способности — это во многом лишь бонус теплокровности. Существуют и гораздо более дешевые способы обзавестись более крупным мозгом.
И все же складывается впечатление, что возможности расширения экологических ниш, ночной активности и дополнительных умственных способностей недостаточно для того, чтобы оправдать ту непомерно высокую цену, которую приходится платить за теплокровность. Кажется, чего-то здесь не хватает. Записанная в дебет цена питания, питания и еще раз питания отнюдь не ограничивается болями в животе. Добывание пищи требует серьезных затрат времени и усилий на охоту или собирательство и делает животное уязвимее для хищников и конкурентов. Пища порой заканчивается. И чем быстрее ешь, тем быстрее она заканчивается. В итоге популяции сокращаются. И действительно, интенсивность обмена веществ, как правило, определяет численность популяций, и у рептилий они часто оказываются в десять раз больше, чем у млекопитающих. По тем же причинам млекопитающие обычно оставляют меньшее потомство (хотя на свое немногочисленное потомство они и могут тратить больше ресурсов). Да и продолжительность жизни во многом определяется интенсивностью обмена веществ. Даже если Клемент Фрейд был прав, говоря о людях, его слова нельзя отнести к рептилиям. Возможно, они и вправду живут медленно и скучно, зато они действительно живут дольше. Гигантские черепахи могут жить сотни лет.
Итак, теплокровность собирает с нас обильную дань. Она означает короткую жизнь, полную опасностей, проведенную за едой. Она снижает размеры популяций и численность потомства, - а естественный отбор должен безжалостно карать и то, и другое. Что же мы получаем в порядке компенсации? Возможность не спать по ночам и гулять на морозе? Кажется, оно того не стоит, особенно если нам все равно требуется сон. И все же мы склонны отдавать самые почетные места в пантеоне жизни именно млекопитающим и птицам. Что же у нас с ними есть такое, чего нет у рептилий?
Единственный убедительный ответ — выносливость. Ящерицы вполне могут померяться с млекопитающими скоростью или силой мышц, а на коротких дистанциях даже оказаться быстрее и сильнее, однако они быстро выдыхаются. Попытайтесь схватить ящерицу: она скроется с глаз в одно мгновение. Но после этого ей придется отдыхать, иногда несколько часов, восстанавливая силы. Дело в том, что рептилии сложены не так, как те, чей удел — беготня. Как и бегуны-спринтеры, они полагаются на анаэробное дыхание, а не на атмосферный кислород, которого все равно бы не хватило. Правда, этот способ не позволяет бежать долго. Рептилии вырабатывают энергию (в форме АТФ) необычайно быстро, но с помощью процесса, вызывающего стремительное накопление молочной кислоты, от которой начинает сводить мышцы.
Эта разница и в структуре мышц. Как мы убедились в главе 6, существуют разные типы мышц. Они отличаются в том числе соотношениями трех ключевых составляющих: мышечных волокон, капилляров и митохондрий. Волокна сокращаются, создавая мышечную силу, капилляры поставляют кислород и удаляют отходы, а митохондрии сжигают пищу, окисляя ее кислородом и вырабатывая энергию, требуемую для сокращения мышц. Беда в том, что все они занимают драгоценное пространство, и чем больше в мышцах волокон, тем меньше остается места капиллярам и митохондриям. Мышцы, до отказа наполненные волокнами, могут обладать огромной силой, но будут быстро растрачивать энергию, необходимую для сокращения. Выбор типа мышц имеет самые далеко идущие последствия: большая сила и низкая выносливость или маленькая сила и высокая выносливость. Сравните грузного спринтера с тощим бегуном на длинные дистанции, и вы оцените разницу.
У нас имеются мышцы сразу нескольких типов, и соотношение их меняется в зависимости от обстоятельств, например от того, живем ли мы на уровне моря или высоко в горах. Значительную роль здесь может играть и образ жизни. Если вы будете тренироваться бегать спринт, то у вас разовьются объемные, быстро сокращающиеся мышцы, отличающиеся большой силой, но низкой выносливостью. Если же тренироваться бегать на длинные дистанции, то развиваются мышцы, работающие медленно, зато долго. Различия по этим показателям зависят также от врожденных особенностей отдельных людей или человеческих популяций, поэтому в течение жизни многих поколений на них может влиять отбор, если тому способствуют обстоятельства. Именно поэтому у непальцев, коренного населения Восточной Африки и андских индейцев есть немало общих черт, помогающих им жить высоко в горах. Народы, живущие на равнинах, обычно отличаются более грузным телосложением.
В классической работе 1979 года, которую опубликовали Альберт Беннет и Джон Рубен из Калифорнийского университета в Ирвайне, утверждается, что именно такие отличия лежат в основе теплокровности. Эти исследователи доказывали, что разница между теплокровностью и холоднокровностью вовсе не в температуре, а в выносливости. Их концепция известна как гипотеза аэробной мощности, и даже если она не во всем верна, она радикально изменила представления исследователей, работающих в этой области.
Гипотеза аэробной мощности сводится к двум тезисам. Во-первых, отбор работает не с температурой, а с повышенной активностью, которая во многих обстоятельствах приносит непосредственную пользу. Вот как сформулировали это сами Беннет и Рубен:
Селективные преимущества повышенной активности отнюдь нетрудноуловимы и имеют принципиальное значение для выживания и размножения. Более выносливое животное получает перед менее выносливыми преимущество, которое легко представить в терминах, связанных с отбором. Такое животное может дольше поддерживать высокую скорость при погоне или бегстве, добывая себе пищу или стремясь не стать пищей хищнику. Оно будет превосходить других по способности защищать свою территорию или вторгаться на чужую. Оно будет успешнее осуществлять ухаживание и спаривание.
С этим трудно спорить. Польский зоолог Павел Котея предложил интересное уточнение этого тезиса. Он подчеркнул значение активной заботы о потомстве, связанной с кормлением детенышей в течение нескольких месяцев или даже лет, характерной для млекопитающих и птиц, но не для холоднокровных животных. Такие “вложения” в потомство требуют ощутимой выносливости и могут существенно повысить шансы животного на выживание, когда оно наиболее уязвимо. Но как бы там ни было, именно второй тезис гипотезы аэробной мощности оказывается и проблематичнее, и интереснее. Он касается взаимосвязанности выносливости и отдыха. Беннет и Рубен утверждают, что неизбежно существует связь между максимальной интенсивностью обмена веществ и уровнем его интенсивности в состоянии покоя. Сейчас объясню.
Максимальная интенсивность обмена веществ определяется как количество кислорода, потребляемое во время максимального напряжения, когда силы организма на пределе. Этот показатель зависит в том числе от физической формы и, разумеется, от генов, но в итоге определяется скоростью потребления кислорода конечными пользователями — митохондриями мышечных клеток. Чем быстрее они потребляют кислород, тем больше максимальная интенсивность обмена. Нетрудно догадаться, что скорость потребления кислорода митохондриями должна быть связана с целым рядом взаимосвязанных факторов. Она определяется числом митохондрий, числом поставляющих им кислород капилляров, давлением крови, размерами и строением сердца, числом красных кровяных телец, тонкой молекулярной структурой переносящего кислород пигмента (гемоглобина), размерами и строением легких, диаметром трахеи, силой диафрагмы и так далее. Если хоть один из этих факторов не работает вровень с другими, максимальная интенсивность обмена веществ не будет достигнута.
Так что отбор на выносливость — это то же, что отбор на высокую максимальную интенсивность обмена веществ, который, в свою очередь, сводится к отбору на целый ряд признаков, связанных с дыханием. Беннет и Рубен утверждают, что высокая максимальная интенсивность обмена так или иначе повышает и интенсивность обмена в состоянии покоя. Иными словами, “атлетичное” млекопитающее с высокой выносливостью должно обладать и высокой интенсивностью обмена в состоянии покоя: такое животное должно потреблять довольно много кислорода даже тогда, когда оно просто лежит и ничего не делает. Беннет и Рубен доказывали это эмпирическим путем. Они отмечали, что, каковы бы ни были причины этого соотношения, максимальная интенсивность обмена всех животных, будь то млекопитающие, птицы или рептилии, обычно примерно раз в десять выше, чем интенсивность обмена в состоянии покоя.
А значит, отбор на высокую максимальную интенсивность обмена должен поднимать и интенсивность обмена в покое. Если максимальная интенсивность возрастает в десять раз, что соответствует разнице между млекопитающими и ящерицами, то интенсивность в покое тоже должна увеличивается в десять раз. А когда до этого доходит, животное начинает вырабатывать такое количество тепла, что становится теплокровным в том смысле, что температура его тела постоянно остается высокой.
Эта гипотеза красива и выглядит вполне разумной, но при ближайшем рассмотрении оказывается трудно представить, почему максимальная интенсивность обмена непременно должна быть связана с интенсивностью обмена в покое. Максимальная интенсивность определяется количеством кислорода, доставляемого к мышцам, а в покое вклад мышц в расход кислорода невелик. Главную же роль в его потреблении играют мозг и другие внутренние органы: печень, поджелудочная железа, почки, кишечник, и так далее. Непонятно, почему печень тоже должна потреблять много кислорода, если много кислорода потребляют мышцы. По крайней мере, вполне можно представить себе животное, обладающее очень высокой аэробной мощностью и очень низкой интенсивностью обмена веществ в покое, — своего рода форсированную ящерицу, объединившую в себе лучшие качества теплокровных и холоднокровных животных. Вполне возможно, что динозавры были именно такими форсированными ящерицами. Откровенно говоря, немного неловко, что мы по-прежнему не знаем, почему у современных млекопитающих, рептилий и птиц максимальная интенсивность обмена должна быть связана с интенсивностью обмена в покое и может ли эта связь у каких-то животных разрываться. Известно, что наиболее атлетичные млекопитающие, такие как североамериканское парнокопытное вилорог, обладают очень высокой аэробной мощностью: максимальная интенсивность обмена у них примерно в шестьдесят пять раз выше, чем интенсивность обмена в покое, а значит, связь между этими параметрами все-таки может нарушаться. То же самое относится и к некоторым рептилиям. Например, максимальная интенсивность обмена у миссисипского аллигатора по меньшей мере в сорок раз выше, чем интенсивность обмена в состоянии покоя.
Как бы там ни было, у нас есть основания считать, что Беннет и Рубен правы. Особенно отчетливо на это указывает источник тепла большинства теплокровных животных. Существует много способов непосредственно вырабатывать тепло, но большинство теплокровных животных к ним не прибегают и тепло у них вырабатывается как побочный продукт обмена веществ. Только у мелких млекопитающих, быстро теряющих тепло, таких как крысы, оно вырабатывается напрямую. Крысы (а также детеныши многих других млекопитающих) пользуются для этого специальной тканью, так называемым бурым жиром, со множеством работающих митохондрий. Эти митохондрии и производят тепло, применяя несложный трюк. В норме в митохондриях возникает электрический ток, который представляет собой поток протонов, проходящих через мембрану. Этот ток используется для синтеза АТФ — энергетической валюты клетки (см. главу 1). Для работы данного механизма требуется сплошная мембрана, играющая роль изоляции. Любая брешь в мембране вызывает короткое замыкание, рассеивающее энергию протонов в виде тепла. Именно это и происходит в буром жире: в мембраны его митохондрий специально вставлены белковые поры, создающие в ней течь. В итоге вместо АТФ эти митохондрии вырабатывают тепло.
Итак, если главная задача — выработка тепла, то ее решение — протекающие митохондрии. Если сделать все митохондрии протекающими, как в буром жире, то вся энергия, получаемая из пищи, будет преобразовываться непосредственно в тепловую. Это простой и быстрый процесс, к тому же не требующий много места, потому что даже небольшое количество ткани позволяет вырабатывать довольно много тепла. Но обычно этот механизм не используется. Степень протекания митохондрий у ящериц, млекопитающих и птиц вполне сравнима. Разница между теплокровными и холоднокровными существами касается, прежде всего, размеров органов и числа митохондрий. Например, печень крысы гораздо крупнее печени любой ящерицы такого же размера и содержит намного больше митохондрий. Иными словами, внутренние органы теплокровных существ, по сути, снабжены турбонагревателями. Эти животные потребляют кислород в огромных количествах, но не для того, чтобы напрямую вырабатывать тепло, а для того, чтобы ускорить обмен веществ. Тепло у них первоначально вырабатывалось как побочный продукт и лишь впоследствии, с развитием наружной термоизоляции, такой как шерсть и перья, тоже было поставлено на службу.
Переход к теплокровности в ходе индивидуального развития современных животных свидетельствует в пользу того, что теплокровность связана скорее с работой внутренних органов в качестве турбонагревателей, чем с выработкой собственно тепла. Фрэнк Зебахер, специалист по эволюционной физиологии из Сиднейского университета, недавно занялся поисками генов, осуществляющих переход к теплокровности у зародышей птиц, и обнаружил, что интенсивная работа внутренних органов запускается всего одним “геном-господином” (он кодирует белок РбС-га), заставляющим делиться митохондрии в клетках этих органов. Управлять размерами органов тоже нетрудно, регулируя с помощью других “генов-господ” настройки, определяющие соотношение деления и гибели клеток. Короче говоря, сделать из органов турбонагреватели с помощью генов не так уж сложно (этим процессом может управлять лишь горстка генов), но в энергетическом плане это очень дорогостоящая операция, поэтому отбор может поддерживать ее лишь в том случае, если она будет окупаться с лихвой.
Итак, сценарий, описываемый гипотезой аэробной мощности, в целом убедителен. Нет никаких сомнений в том, что выносливость у теплокровных животных гораздо выше, чем у холоднокровных: обычно раз в десять, если оценивать ее по аэробной мощности. И у млекопитающих, и у птиц эта зашкаливающая аэробная мощность сочетается с повышенным обменом веществ в состоянии покоя, который обеспечивают крупные внутренние органы, богатые митохондриями и работающие как турбонагреватели, так что специально на выработку тепла не тратится особых усилий. Подозреваю, что высокая аэробная мощность действительно сопряжена с усиленной работой системы обеспечения. Эта идея вполне проверяема. Можно проводить искусственный отбор на повышение аэробной мощности и отслеживать, будет ли повышаться интенсивность обмена в покое. Эти два параметра должны по меньшей мере коррелировать, даже если причинно-следственную связь между ними сложно доказать.
С тех пор, как эта гипотеза была выдвинута, прошло почти тридцать лет. Было предпринято немало попыток проверить ее экспериментально, но ни одна не увенчалась полным успехом. Между максимальной интенсивностью обмена и интенсивностью обмена в покое в целом действительно наблюдается корреляция — но не более того, причем из данного правила есть немало исключений. Возможно, на каком-то этапе эволюции эти два параметра и вправду были связаны, даже если физиологические причины и не требуют строгой связи между ними. Не зная подробностей эволюционной истории теплокровности, трудно сказать точно. Но на сей раз палеонтологическая летопись, оказывается, все-таки может дать нам ключ к ответу. Вполне вероятно, что недостающее звено кроется не в физиологии, а в превратностях судьбы.
Теплокровность — это, по сути, усиленный режим работы внутренних органов, например печени. Но время беспощадно к мягким тканям, и даже шерсть в ископаемом состоянии нечасто сохраняется. Поэтому долгое время по палеонтологической летописи сложно было разобраться в происхождении теплокровности, и даже сегодня об этой проблеме не так уж редко идут ожесточенные споры. Но пересмотр палеонтологической летописи в свете гипотезы аэробной мощности — задача вполне выполнимая, поскольку и строение скелета может о многом нам рассказать.
Предполагаемые предки млекопитающих и птиц известны еще из триасового периода, начавшегося около двухсот пятидесяти миллионов лет назад. Этот период последовал непосредственно за крупнейшим массовым вымиранием за всю историю нашей планеты — пермским вымиранием, которое, как полагают, привело к исчезновению примерно 95 % существовавших тогда видов. Среди тех немногих, кто пережил тот апокалипсис, были две группы рептилий: терапсиды (“звероподобные рептилии”), предки современных млекопитающих, и архозавры (от греческого “правящие ящеры”), предки птиц и крокодилов, а также динозавров и птерозавров.
Учитывая последующее господство динозавров, может показаться странным, что в начале триаса именно терапсиды были наиболее успешной из этих двух групп. Их потомки, млекопитающие, уменьшились в размерах и стали жить в норах еще до того, как начался расцвет динозавров. Но в раннем триасе господствующими рептилиями были представители рода листрозавров (греч. Lystrosaurus — “ящер-лопата”) — растительноядные животные размером со свинью, с двумя короткими и толстыми бивнями, широкой мордой и грудью колесом. Какой образ жизни вели листрозавры, трудно сказать. Многие годы их считали полуводными животными, некрупными рептильными бегемотами, но теперь полагают, что они обитали в довольно сухом климате и предположительно рыли норы, как и многие другие терапсиды. Мы вернемся к значению этого позже, но так или иначе, начало триаса было периодом доминирования листрозавров, ничего подобного которому никогда впоследствии не наблюдалось. Утверждают, что на некотором отрезке времени листрозавры составляли 95 % всех наземных позвоночных. Вот что писал об этом американский поэт и натуралист Кристофер Кокинос: “Представьте, что вы проснетесь завтра и отправитесь в путешествие по континентам, и везде вам будут встречаться, скажем, одни белки”.
Листрозавры были растительноядными (может быть, единственными растительноядными рептилиями того времени) и во времена своего господства не боялись никаких хищников. В более поздние эпохи триасового периода листрозавров начала вытеснять другая, родственная им группа терапсид — так называемые цинодонты (“собачьи зубы”), и в конце триаса, около двухсот миллионов лет назад, листрозавры вымерли. Среди цинодонтов были как растительноядные, так и хищные. Именно от цинодонтов и произошли млекопитающие, возникшие ближе к концу триаса. Цинодонты уже обладали многими признаками высокой аэробной мощности, в том числе твердым нёбом (отделяющим дыхательные каналы от ротовой полости и позволяющим одновременно дышать и жевать), широкой грудью с видоизмененной грудной клеткой и, по-видимому, мускулистой диафрагмой. Более того, их носовые ходы были увеличены и содержали тонкие решетчатые кости — так называемые носовые раковины. Вполне возможно, цинодонты были даже покрыты шерстью, хотя они откладывали яйца, как большинство рептилий.
Итак, вполне вероятно, что цинодонты уже обладали повышенной аэробной мощностью, которая должна была обеспечивать им высокую выносливость. Но какова была интенсивность их обмена веществ в состоянии покоя? Были ли они уже теплокровными? Джон Рубен утверждает, что одним из немногих надежных признаков повышенного уровня обмена в состоянии покоя могут служить носовые раковины. Эти структуры уменьшают потерю воды при дыхании, что может быть принципиально, если высокая интенсивность дыхания поддерживается постоянно, а не сопровождает непродолжительные периоды активности. Поскольку у современных рептилий интенсивность обмена в состоянии покоя невелика, в этом состоянии они дышат очень сдержанно и не нуждаются в приспособлениях, снижающих потерю воды. В итоге ни у одной современной рептилии носовых раковин нет, в то время как у всех настоящих теплокровных существ такие раковины имеются, хотя из этого правила есть исключения: приматы и некоторые птицы. Ясно, что носовые раковины при теплокровности полезны, хотя в них и нет абсолютной необходимости, и их наличие у ископаемых может служить важным ключом к разгадке происхождения теплокровности. Учитывая также предполагаемое наличие шерсти (имеются лишь косвенные указания, потому что в ископаемом виде она не сохранилась), судя по всему, цинодонты на определенном этапе эволюционной линии, ведущей к млекопитающим, действительно стали теплокровными.
Но, несмотря на все это, цинодонты вскоре отошли на второй план, и в результате позднетриасового “переворота”, осуществленного воинственными архозаврами, они были вынуждены перейти к скрытному, ночному образу жизни. Если у цинодонтов к тому времени уже выработалась теплокровность, то как обстояли дела у победителей — группы, из которой вскоре возникли динозавры? Последние дожившие до наших дней потомки архозавров — крокодилы и птицы — соответственно холоднокровны и теплокровны. Где-то на пути к первым птицам у архозавров должна была выработаться теплокровность. Но у каких и почему? И были ли среди них динозавры?
Здесь ситуация осложняется, и некоторые вопросы вызывают среди ученых жесточайшие споры. Птицы, как и динозавры, привлекают к себе столько внимания, что иногда об их происхождении высказываются версии, которые лишь с натяжкой можно назвать научными. Уже давно считается, что птицы так или иначе родственны динозаврам, особенно одной их группе — так называемым тероподам, к которым относится, в частности, знаменитый тираннозавр (Tyrannosaurus rex). С 80-х годов XX века был проведен целый ряд сравнительно-анатомических систематических исследований (в рамках подхода, именуемого кладизмом), результаты которых довольно убедительно показывают, что эволюционная линия птиц восходит непосредственно к одной из ветвей теропод. Это означает, что птицы не просто родственны динозаврам: они сами представляют собой динозавров — птичью линию теропод. Хотя большинство специалистов согласны с этими выводами, существует и меньшинство, представители которого доказывают их ошибочность. Их лидером считается выдающийся палеоорнитолог Алан Федуччиа из Университета Северной Каролины. Он утверждает, что птицы произошли от более ранней (какой точно, неизвестно) группы архозавров, отделившейся от их ствола еще до возникновения теропод. Согласно этим представлениям, птицы не динозавры, а особая, отдельная группа архозавров.
Последние данные, полученные по этой проблеме, вызвали особенно громкий резонанс. Они касаются не морфологических признаков, а белков. Удивительное открытие, которое сделали в 2007 году Джон Асара и его коллеги из Гарвардской медицинской школы, состояло в том, что одна необычайно хорошо сохранившаяся кость тираннозавра возрастом около шестидесяти восьми миллионов лет по-прежнему содержит фрагменты коллагена — главного органического компонента костей. Исследователям удалось прочитать последовательность аминокислот нескольких фрагментов, а затем соединить их и получить небольшой участок последовательности аминокислот коллагена тираннозавра. В 2008 году ученые сравнили этот участок с соответствующими участками последовательностей аминокислот в коллагене млекопитающих, птиц и аллигаторов. Этот участок невелик, так что восстановленная на его основе картина может и не соответствовать действительности, но на первый взгляд ближайшим современным родственником тираннозавра из всех исследованных видов оказалась курица, а следующим, тоже довольно близким — африканский страус. Неудивительно, что публикация этих данных была встречена бурей одобрения в газетах, читателям которых давно хотелось знать, каково было мясо тираннозавра на вкус. Но гораздо существеннее было то, что исследование коллагена в целом подтвердило выводы кладистов о том, что птицы представляют собой одну из ветвей динозавров-теропод.
Другой камень преткновения представляет оперение. Алан Федуччиа и ряд других ученых долгое время утверждали, что перья развились у птиц как приспособление для полета, которым они овладели настолько, что оно кажется граничащим с чудом. Но если перья развились как приспособление для полета, их никак не должно было быть у нелетающих теропод, таких как тираннозавр. Федуччиа именно это и доказывал, но за последнее десятилетие в Китае удалось раскопать целый ряд оперенных динозавров. Хотя некоторые из их остатков и вызывают сомнения в правильности интерпретации, в данном случае большинство специалистов также убеждено, что нелетающие тероподы, в том числе вероятный маленький предок тираннозавров, действительно имели оперение.
Альтернативная точка зрения, согласно которой “перья” нелетающих теропод были не тем, чем кажутся, и на самом деле представляют собой раздавленные коллагеновые волокна, отдает подгонкой под ответ. Если это действительно были просто коллагеновые волокна, сложно объяснить, почему они встречаются преимущественно у представителей одной группы теропод — так называемых рапторов, к числу которых относятся и велоцирапторы, прославленные авторами “Парка юрского периода”, а также почему они выглядят точно так же, как перья самых настоящих птиц, ископаемые остатки которых сохранились в тех же отложениях. Мало того, что эти перья похожи на перья: некоторые из рапторов, особенно микрорапторы, могли, судя по всему, планировать с дерева на дерева с помощью перьев, обильно покрывавших у них все четыре конечности (или, если можно так выразиться, все четыре крыла). Мне сложно поверить, что эти прекрасно сохранившиеся перья - на самом деле не перья, и даже Федуччиа уже начал в этом сомневаться. Однако вопрос, имеет ли планирующий полет микрорапторов с дерева на дерево отношение к возникновению полета у настоящих птиц или у их ближайших родственников - археоптериксов, — остается спорным.
Вывод о том, что перья возникли у динозавров-теропод еще до возникновения полета, подтверждают результаты исследований зародышевого развития птичьих перьев, особенно в сравнении с развитием кожи у зародышей крокодилов. Напомним, что крокодилы тоже представляют собой сохранившуюся ветвь архозавров, появившихся в триасе. Расхождение ветвей крокодилов и динозавров (в том числе птиц) началось в середине триаса, около 230 миллионов лет назад. Однако как бы давно предки крокодилов ни разошлись с предками птиц, в их организме уже были “семена” перьев. Даже у современных крокодилов в коже зародышей сохранилисьте же слои, из которых у птиц развиваются перья, а также белки (так называемые кератины пера), которые, естественно, отличаются легкостью, гибкостью и прочностью.
Кератины пера присутствуют в основном в тех слоях кожи зародышей крокодилов, которые сбрасываются после выхода из яйца, обнажая развивающееся под ними покрытие из чешуи (при этом остатки кератинов пера присутствуют и в чешуе взрослых крокодилов). У птиц подобная чешуя имеется на ногах, и слои кожи, покрывающие ее у зародыша, тоже сбрасываются после вылупления. Лоренцо Алибарди, специалист по эволюционному развитию перьев из Болонского университета, показал, что перья развиваются из тех самых слоев кожи зародыша, которые сбрасываются при образовании чешуи. Зародышевая чешуя при этом удлиняется, образуя трубчатые нити — бородки. Эти полые структуры напоминают волосы, однако имеют стенки из живых клеток, происходящих из определенных слоев зародышевой кожи, так что в любой части бородок могут отрастать ветви7. Самые простые перья (пуховые) представляют собой пучки бородок, растущих из одной точки, в то время как маховые перья оснащены множеством ветвистых бородок, плотными рядами отходящих от центрального стержня. Живые клетки стенок бородок откладывают кератин, а затем дегенерируют, обнажая построенную из кератина ветвящуюся структуру. Так развивается перо. У крокодилов имеются не только слои кожи и белки, задействованные в росте перьев, но даже гены, необходимые для управления этим процессом, что заставляет предположить наличие этих генов и у тех архозавров, которые были общими предками крокодилов и птиц. Изменились только программы развития. Близкое эмбриологическое родство между птичьими перьями и чешуей выдает также одна в высшей степени странная мутация, приводящая к тому, что у птиц на ногах на месте чешуй вырастают перья. Однако оперенных крокодилов еще никто пока не встречал.
В итоге складывается впечатление, что примитивные перья имелись чуть ли не у древнейших архозавров, и нет ничего удивительного в том, что у теропод начали отрастать “придатки эпидермиса”, по-видимому, весьма разнообразные, от щетинок (как у птерозавров) до простых ветвящихся структур вроде пуховых перьев. Но для чего они, если не для полета? На этот вопрос есть целый ряд правдоподобных ответов, отнюдь не взаимоисключающих: привлечение полового партнера, участие в работе органов чувств, защитные функции (придатки эпидермиса могли увеличивать видимые размеры тела или образовывать иглы, как у дикобраза), а также, разумеется, теплоизоляция. Перья, в изобилии обнаруженные у теропод, несомненно, повышают вероятность того, что эти животные были теплокровными, как и их современные родичи — птицы.
Имеются и другие данные, указывающие на то, что тероподы были довольно активными динозаврами — по крайней мере, они отличались выносливостью. В частности, это сведения, касающиеся строения сердца. У крокодилов и птиц, в отличие от ящериц и большинства других рептилий, мощное четырехкамерное сердце. Это заставляет предположить, что сердце такого типа унаследовано птицами и крокодилами от общих предков и что им обладали и другие архозавры, в том числе динозавры. Четырехкамерное сердце важно тем, что оно разделяет кровообращение на два круга, по одному из которых кровь поступает в легкие, а по второму — ко всем остальным органам. Эта схема имеет два важных преимущества. Во-первых, она позволяет под большим давлением закачивать кровь в мышцы, мозг и другие органы, не вызывая повреждений в тонких тканях легких (из-за которых может развиваться отек, нередко приводящий к смертельному исходу). Ясно, что более высокое давление крови позволяет поддерживать более высокий уровень активности, а также достигать намного более крупных размеров. Гигантские динозавры с длинными шеями не смогли бы снабжать свой мозг кровью без четырехкамерного сердца. Во-вторых, полное разделение системы кровообращения на два круга означает, что насыщенная кислородом кровь больше не смешивается с ненасыщенной, когда возвращается в сердце из легких, откуда она сразу же поступает в другие органы, с предельной эффективностью доставляя кислород туда, где он нужен. Хотя четырехкамерное сердце не обязательно означает теплокровность (в конце концов, крокодилы холоднокровны), без него добиться высокой аэробной мощности почти невозможно.
Дыхательная система теропод, судя по всему, была похожа на птичью и позволяла поддерживать высокий уровень активности. Механизм работы легких у птиц отличается от нашего. Он гораздо эффективнее даже на небольших высотах. На больших же высотах разница оказывается колоссальной. Птицы могут извлекать из разреженного воздуха втрое больше кислорода, чем млекопитающие. Именно поэтому гуси способны совершать перелеты, поднимаясь в небо на тысячи футов выше вершины Эвереста, в то время как млекопитающие начинают задыхаться и на куда меньших высотах.
Наши легкие устроены как дерево с глубоким разветвленным дуплом, в которое воздух попадает через отверстие у основания ствола (соответствующее трахее) и заходит в две ветви (соответствующих главным бронхам), где “распускается”. Но “веточки” наших легких заканчиваются не острыми концами, а маленькими надувными шариками — альвеолами, стенки которых пронизаны капиллярами. В этих шариках происходит газообмен: здесь гемоглобин, содержащийся в красных кровяных тельцах, отдает углекислый газ и связывает кислород, после чего насыщенная кислородом кровь возвращается в сердце. Вся система шариков при вдохе и выдохе надувается и сдувается, как кузнечные мехи, за счет работы диафрагмы и мышц грудной клетки. Неизбежный недостаток этой системы состоит в том, что разветвленное дупло заканчивается слепыми отростками, где воздух едва перемешивается, хотя именно туда и требуется обеспечить прежде всего приток воздуха. И даже тот воздух, который все-таки достигает альвеол, попутно смешивается с выдыхаемым воздухом и оказывается не таким уж и свежим.
Птичьи же легкие представляют собой превосходную модификацию легких рептилий. Легкие у рептилий обычно простые: это всего лишь большие надувные мешки, центральная полость которых разделена перегородками из легочной ткани — так называемыми спетами. Легкие рептилий, как и млекопитающих, работают подобно кузнечным мехам — либо за счет расширения грудной клетки, либо (у крокодилов) за счет действующей по принципу поршня диафрагмы, соединенной с печенью и двигаемой мышцами, прикрепленными к лобковой кости. Это делает легкие крокодила похожими на шприц, который тоже наполняется за счет того, что его поршень отводится назад. Хотя этот способ дыхания вполне эффективен, птицы зашли намного дальше, превратив половину своего тела в сложную однонаправленную систему взаимосвязанных воздушных мешков. Воздух входит в их легкие не напрямую, а вначале поступает в воздушные мешки, после чего, уже на выходе, проходит сквозь легкие. В результате воздух в птичьих легких не застаивается, как в наших, а постоянно замещается. Свежий воздух в птичьих легких проходит мимо септ (тоже усовершенствованных) как на вдохе, так и на выдохе. Это происходит за счет движений нижней части грудной клетки и системы воздушных мешков задней части тела. Диафрагмы у птиц нет, что тоже принципиально. Более того, воздух в птичьих легких движется в одну сторону, а кровь — в другую, противоположную, что, согласно принципу противотока, позволяет осуществлять газообмен с максимальной эффективностью.
Вопрос, разделивший исследователей на два враждовавшие не одно десятилетие лагеря, касается того, какие легкие были у древних теропод: поршневые, как у крокодилов, или вентилируемые сквозным путем, как у птиц. Птичья система воздушных мешков заходит не только в мягкие ткани внутри брюшной полости и грудной клетки, но даже в кости, в том числе в ребра и в позвоночник. Давно было известно, что у ископаемых теропод в костях имелись полости, причем в тех же местах, где и у птиц. Одним из главных разжигателей вражды между специалистами был американский палеонтолог Роберт Бэккер, в 70-х годах на основании этих и других сведений реконструировавший динозавров как активных теплокровных животных. Именно те революционные представления и вдохновили Майкла Крайтона на написание книги “Парк юрского периода”, по которой впоследствии был снят одноименный фильм. Однако Джон Рубен и его коллеги реконструировали легкие ископаемых теропод иначе. Они пришли к выводу, что легкие этих динозавров были гораздо ближе к крокодильим, чем к птичьим, и работали по принципу шприца. На одном или двух ископаемых образцах исследователи даже нашли предполагаемые следы диафрагмы. Рубен не отрицал существования воздушных полостей в костях ископаемых теропод, но иначе интерпретировал их назначение. Он утверждал, что полости служили не для того, чтобы помогать вентиляции легких, а, возможно, для уменьшения веса костей — или чтобы двуногим животным было легче удерживать равновесие. Споры не утихали, и без новых данных не было возможности их разрешить. И вот в 2005 году Патрик О’Коннор и Леон Классенс, работавшие тогда в Университете Огайо и в Гарварде соответственно, опубликовали в журнале “Нейчур” статью, ставшую новой вехой в исследовании этой проблемы.
О’Коннор и Классенс начали с того, что подробно исследовали системы воздушных мешков нескольких сотен видов современных птиц (на материале, как они писали, “предназначенных на выброс экземпляров” из центров лечения диких животных и из музеев). Они наполняли воздушные мешки мертвых птиц латексом, что помогало лучше разобраться в анатомии дыхательной системы. Первое, в чем убедились исследователи, — это что система воздушных мешков проникает гораздо глубже, чем они предполагали, и занимает не только часть шеи и груди, но и немалую долю брюшной полости, откуда воздушные мешки проникают в нижние отделы позвоночника. Последняя деталь имела принципиальное значение для интерпретации строения скелетов ископаемых теропод. Работу всей дыхательной системы птиц определяет задний (хвостовой) воздушный мешок. В процессе дыхания он сжимается, проталкивая воздух вперед, в легкие, а затем расширяется, засасывая воздух из связанных с ним воздушных мешков в груди и шее. (Можно привести заумный термин — аспирационный насос). Работа такой системы отчасти похожа на игру на волынке, в которую регулярно подкачивают воздух, чтобы он мог непрерывно выходить из трубок.
О’Коннор и Классенс использовали результаты своего исследования для анализа строения костей ископаемых теропод, в том числе прекрасно сохранившегося скелета динозавра Majungatholus atopus — представителя группы теропод, состоящей в довольно далеком родстве с птицами. Прежде ученые, изучавшие возможность существования у динозавров воздушных мешков, уделяли особое внимание их шейным позвонкам и ребрам, а О’Коннор и Классенс занялись поиском полостей в задних отделах позвоночника. Наличие таких полостей должно было свидетельствовать о том, что у ископаемых теропод имелся задний воздушный мешок. И действительно, исследователям удалось обнаружить именно такие полости, расположенные в точности там же, где и у птиц. Более того, анатомическое строение позвоночника, грудной клетки и грудины в целом указывало на работу аспирационного насоса: повышенная подвижность задних ребер и грудины позволяла сжимать хвостовой воздушный мешок, проталкивавший воздух вперед, и вентилировать легкие. Эти результаты оставляли мало места для сомнений в том, что у ископаемых теропод и впрямь действовал аспирационный насос, подобный птичьему — самая эффективная система дыхания из всех, которыми пользуются позвоночные.
Итак, у ископаемых теропод были перья, четырехкамерное сердце и воздушные мешки, обеспечивавшие сквозную вентиляцию легких, и все это в целом указывает на их активный образ жизни, требующий высокой выносливости. Но действительно ли тероподы могли достичь такой выносливости исключительно за счет настоящей теплокровности, как предполагает гипотеза аэробной мощности, или же они представляли собой “компромиссный” вариант, промежуточный между современными крокодилами и птицами? Хотя наличие перьев и может указывать на теплоизоляцию, а, значит, и теплокровность, перья древних теропод вполне могли выполнять какие-то иные функции. При этом поиски других свидетельств теплокровности, в том числе носовых раковин, дают результаты, однозначно интерпретировать которые еще сложнее.
У большинства птиц, как и у большинства млекопитающих, имеются носовые раковины, однако они состоят не из костей, как у человека, а из хрящей, которые в ископаемом виде плохо сохраняются. До сих пор у теропод не обнаружено никаких признаков носовых раковин, хотя образцов достаточно хорошей сохранности, чтобы судить об этом, известно немало. Джон Рубен отмечает, что у птиц, имеющих носовые раковины, всегда увеличены носовые ходы. Причина здесь предположительно в том, что тонкие завитки носовых раковин в некоторой степени затрудняют прохождение воздуха, а увеличение носовых ходов позволяет компенсировать это неудобство. Впрочем, носовые ходы ископаемых теропод не особенно велики, и мы не находим у них носовых раковин не оттого, что те не сохранились, а оттого, что их не было. Могли ли тероподы быть теплокровными, не имея носовых раковин? Ну, учитывая, что мы, сами будучи теплокровными, не имеем таких раковин, следует ответить на этот вопрос утвердительно, но это допущение ставит перед нами еще несколько вопросов.
Сам Рубен считает, что ископаемые тероподы все-таки обладали высокой аэробной мощностью, но не были теплокровными, хотя его собственная гипотеза аэробной мощности предполагает, что одно должно быть связано с другим. Мы знаем еще слишком мало, чтобы ответить на этот вопрос уверенно, но сложившееся к настоящему времени общее мнение (если его можно считать общим), таково: уровень обмена веществ в состоянии покоя у ископаемых теропод был, по-видимому, повышен, но настоящей теплокровности у них все же не было. По крайней мере, именно на это указывают ископаемые остатки. Однако горные породы содержат не только ископаемых, но и многое другое, в том числе свидетельства о климате и составе атмосферы прошлого. И эти свидетельства говорят нам об атмосфере триасового периода нечто такое, в свете чего данные палеонтологической летописи принимают совсем другой оборот. Эти сведения помогают не только объяснить высокую аэробную мощность цинодонтов и теропод, но и ответить на вопрос, почему господствующее положение в итоге заняли динозавры.
Проблемы физиологии обычно обсуждают так, будто участники обсуждения пребывают в историческом вакууме: исходя из того, что в прошлом все было так же, как теперь, и что силы отбора неизменны, как сила тяготения. Но это не так, доказательством чему служат великие вымирания. Самое грандиозное произошло в конце пермского периода, около 250 миллионов лет назад, и послужило прелюдией к начавшемуся сразу после него неудержимому взлету “правящих ящеров” и эре динозавров.
Пермское вымирание часто называют одной из великих загадок жизни (наряду с целым рядом других явлений, на изучение которых охотнее выделяют гранты), но условия среды, тот фон, на котором оно происходило, пока описаны лишь в общих чертах. На самом деле это было даже не одно вымирание, а два, разделенные промежутком почти в десять миллионов лет — временем отчаянного упадка. Оба вымирания соответствуют продолжительным периодам повышенной вулканической активности — самым масштабным излияниям лавы в истории Земли, похоронившим обширные территории (чуть ли не целые континенты) под толстым слоем базальтов. Эрозия застывших потоков этой лавы привела к появлению участков со ступенчатым рельефом — так называемых траппов. Первый всплеск вулканической активности, случившийся около 260 миллионов лет назад, привел к образованию Эмэйшаньских траппов в Китае. Восемь миллионов лет спустя последовал второй всплеск, еще более масштабный, в результате которого возникли Сибирские траппы. Принципиально здесь то, что и Эмэйшаньские, и сибирские вулканические потоки извергались, пробиваясь сквозь слои, содержавшие карбонатные породы и уголь. Это важно потому, что при реакции раскаленной лавы с такими породами в атмосферу в огромных количествах выделялись углекислый газ и метан, и это происходило при каждом извержении на протяжении не одной тысячи лет. А в результате менялся климат.
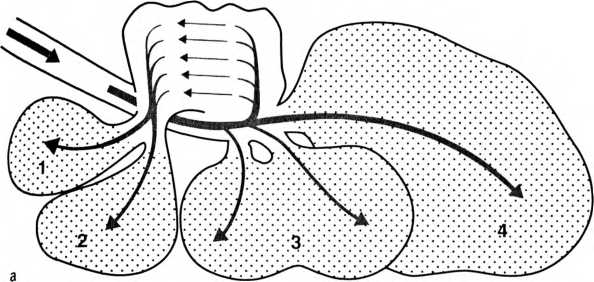
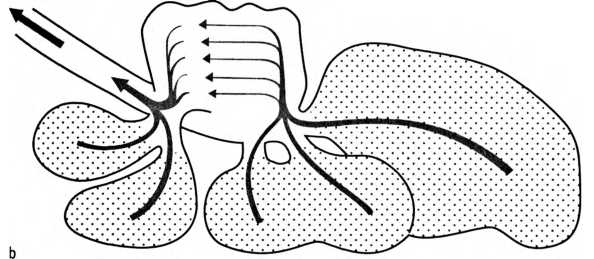
Поток воздуха, проходящий через птичьи легкие на вдохе (о) и на выдохе (Ь). Воздушные мешки: 1 — ключичный, 2 — черепной, 3 — хвостовой-грудной, 4 — брюшной. Воздух непрерывно проходит сквозь легкие в одном и том же направлении, в то время как кровь течет в противоположную сторону, чем по принципу противотока обеспечивается высокоэффективный газообмен.
Было предпринято немало попыток вычислить “убийцу”, стоявшего за пермскими вымираниями. Приводились убедительные аргументы в пользу того, что им было глобальное потепление, истощение озонового слоя, выделение метана и углекислого газа, кислородное голодание, отравление сероводородом, и так далее. Единственная возможная причина, которая более или менее исключена — это столкновение с метеоритом: нет почти никаких данных, указывающих на то, что в конце пермского периода происходили какие-либо столкновения, похожие на последовавшее почти двести миллионов лет спустя и ознаменовавшее собой финал продолжительного господства динозавров. Однако все остальные из перечисленных причин вполне правдоподобны, и большим достижением последних лет стало открытие, что все они неразрывно связаны друг с другом. Любой всплеск вулканической активности, сравнимый по масштабам с тем, что привел к образованию Эмэйшаньских траппов, неизбежно должен был запустить целый каскад неотвратимых взаимозависимых изменений — леденящую сердце последовательность. Сходные каскады угрожают нам и сегодня, хотя (пока) они несопоставимы по масштабам с пермскими.
Извержения вулканов вызывали выброс в атмосферу метана, углекислого газа и других вредных газов, разрушавших озоновый слой и в итоге приведших к установлению на планете гораздо более теплого и сухого климата. Через весь суперконтинент Пангею протянулись обширные сухие равнины. Великие угленосные болота предшествующих периодов, каменноугольного и пермского, высохли, и отложившийся в них уголь полетел по ветру, реагируя с кислородом и делая сам воздух малопригодным для жизни. За десять миллионов лет уровень кислорода медленно, но неуклонно падал — с 30 % до минимума, составившего менее 15 %. Сочетание потепления воды (снижавшего растворимость кислорода), падения уровня атмосферного кислорода и повышения уровня углекислого газа привело к гибели большинства морских организмов. Процветали лишь ядовитые бактерии, преобладавшие на нашей планете задолго до эпохи животных и растений и теперь вновь г огромных количествах выделявшие токсичный сероводород. Газы, испускаемые мертвеющими океанами, отравляли атмосферу еще сильнее, вызывая гибель населения суши. И тогда судьба нанесла еще ряд сокрушительных ударов: раз за разом на протяжении пяти миллионов лет несли смерть извержения — те самые, что вызвали образование Сибирских траппов. В морях и на суше оставалось мало живого, и надежда забрезжила лишь потом.
Кто же выжил? Любопытно, что и в морях, и на суше это удалось тем, кто обладал одним и тем же свойством — лучше других умел дышать, перенося низкий уровень кислорода, высокий уровень углекислого газа и присутствие жуткой смеси из вредоносных газов. Это были те, кто мог задерживать дыхание, но сохранять активность: те, кто обитал в пещерах и норах, в грязи, в болотах, в иле, кто питался чем придется и жил в местах, где никому другому жить не хотелось. “Слизких тварей миллион” выжил, и с ними выжили наши предки. Именно поэтому знаменательно, что первыми наземными животными, восстановившимися после того великого вымирания, были листрозавры — норные рептилии с грудью колесом, с мускулистой диафрагмой, с твердым нёбом, с расширенными дыхательными каналами и с носовыми раковинами. Они вылезли, пыхтя, из своих душных нор и разбрелись по опустевшей суше как белки по лесу.
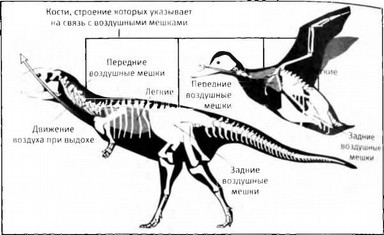
Реконструкция системы воздушных мешков динозавра Majungatholus atopus в сравнении с системой воздушных мешков современных птиц. В обоих случаях легкие обслуживаются передними и задними воздушными мешками, следы которых, сохранившиеся в костях динозавров, полностью соответствуют расположению этих мешков у птиц. Воздушные мешки работают как мехи, прокачивая воздух сквозь неподвижные легкие.
Эта удивительная история, записанная в горных породах на языке химии, длилась миллионы лет и определила ход триасового периода. Токсичные газы исчезли из атмосферы, но уровень углекислого газа по-прежнему зашкаливал — он был в десять раз выше, чем в наши дни. Концентрация кислорода в атмосфере упорно держалась ниже отметки 15 %, и климат оставался засушливым. Даже тем, кто жил на уровне моря, приходилось довольствоваться воздухом, в одном глотке которого было столько же кислорода, сколько сегодня бывает лишь высоко в горах. Именно в таком мире жили первые динозавры, вставшие на задние ноги и освободившие свои легкие от тех ограничений, с которыми сталкиваются ползающие по земле ящерицы, неспособные одновременно бегать и дышать. Прибавьте к этому воздушные мешки и аспирационный насос, и расцвет динозавров покажется неизбежным. Обо всем этом подробно и убедительно рассказывает палеонтолог Питер Уорд из Вашингтонского университета в Сиэтле в своей замечательной книге “Порождения скудного воздуха”. Уорд утверждает (и я ему верю), что секрет успеха архозавров, в итоге вытеснивших цинодонтов, состоял в том, что их легкие были разделены на септы: это давало архозаврам возможность преобразовать их в удивительную систему со сквозным потоком воздуха, которой сегодня пользуются птицы. Тероподы стали единственными животными на планете, не страдавшими от недостатка кислорода. Поэтому и носовые раковины были им не особенно нужны.
Итак, повышенная выносливость была не дополнительным бонусом, а жизненно важным свойством, счастливым лотерейным билетом, позволившим тем, кому он достался, пережить тяжелейшие времена. Но здесь я вынужден поспорить с Уордом. Я согласен, что высокая аэробная мощность должна была оказаться ключом к выживанию, но действительно ли она должна быть сопряжена с высокой интенсивностью обмена веществ в состоянии покоя? Уорд, ссылаясь на гипотезу аэробной мощности, считает именно так, но почему-то ничего подобного не происходит в наши дни с животными, обитающими высоко в горах. Напротив, их мышечная масса обычно уменьшается, и в выигрыше оказываются самые жилистые из этих существ. Хотя их аэробная мощность и бывает повышена, интенсивность обмена в покое вовсе не повышается вместе с ней - напротив, она может даже снижаться. В тяжелые времена физиология должна быть экономнее.
Действительно ли в триасе, когда главной задачей было выжить, животные без нужды повышали интенсивность своего обмена в покое? В этом трудно не усомниться. Судя по всему, тероподы повысили свою аэробную мощность, не сделавшись при этом по-настоящему теплокровными, по крайней мере сначала. При этом побежденные цинодонты, похоже, все-таки стали теплокровными. Было ли это одной из последних отчаянных попыток преуспеть в борьбе с грозными архозаврами? Или же это помогло им сохранять активность, уменьшившись в размерах и перейдя к ночному образу жизни? Оба варианта выглядят вполне правдоподобными, но мне еще больше нравится третий, который, возможно, помогает ответить также на вопрос, почему динозавры поступили прямо обратным образом: выросли в гигантов, подобных которым свет не видывал ни до, ни после.
В вегетарианцах меня часто раздражает их чувство морального превосходства. Хотя, быть может, во мне говорит чувство вины, связанное с собственной плотоядностью. Но если верить одной не самой известной, но весьма важной статье, опубликованной в 2008 году в не самом известном журнале “Эколоджи леттерс”, у вегетарианцев могут иметься и более весомые основания для гордости, чем общеизвестные. Если бы не вегетарианцы (не нынешние, а их предшественники — древние растительноядные животные), наши предки, может, никогда бы не пришли к теплокровности и связанному с ней быстрому темпу жизни. Марсель Классен и Барт Нолет из Нидерландского института экологии — авторы упомянутой статьи — весьма успешно применили счетоводческий (по-научному — стехиометрический) подход к выяснению различий между животной и растительной пищей.
Услышав словосочетание “белковая пища”, многие наверняка представят себе сочный бифштекс. Действительно, белки и мясо в нашем сознании крепко связаны друг с другом (благодаря, надо полагать, бесконечным кулинарным телепередачам и руководствам по похудению). Чтобы снабжать организм белками, нужно есть мясо, а если вы вегетарианец — то орехи и семена, особенно семена бобовых. В целом для вегетарианцев характерно большее внимание к составу своего рациона, чем для мясоедов. Белковую пищу нужно есть, чтобы получать достаточно азота, а это, в свою очередь, необходимо для синтеза наших собственных белков, а также ДНК, которая тоже содержит много азота. На самом деле поддерживать сбалансированный рацион не так уж сложно даже вегетарианцам, но ведь мы — животные теплокровные, а это означает, что мы в любом случае едим много. Классен и Нолет отмечают, что к холоднокровным животным это не относится. Их холоднокровность означает, что они едят не так уж много, и из-за этого у них возникает одна интересная проблема.
Современных растительноядных ящериц очень мало, а среди 3,4 тысячи видов змей нет и вовсе ни одного растительноядного. Некоторые ящерицы все же специализируются на растительной пище, но для них обычно характерны или довольно крупные размеры (зеленые игуаны), или более высокая активность, а также более высокая температура тела по сравнению с плотоядными ящерицами. В отличие от плотоядных ящериц, в случае надобности быстро снижающих температуру тела и погружающихся в оцепенение, растительноядные ящерицы не обладают подобной гибкостью и не позволяют себе расслабляться. Традиционно это связывали с трудностями переваривания растительной пищи, которое осуществляется с помощью кишечных микробов, способных расщеплять ее особо стойкие компоненты (этот процесс гораздо лучше идет при высоких температурах). Но Классен и Нолет утверждают, что здесь может быть и другая причина, связанная с низким содержанием азота в обычной растительной пище. Исследователи рассчитали количество азота, содержащегося в различных продуктах питания ящериц, и убедились, что у растительноядных ящериц с этим действительно связана серьезная проблема.
Представьте, что вы питаетесь только растительной пищей, содержащей мало азота. Как обеспечить себя азотом? Можно, конечно, попытаться разнообразить рацион, искать падаль, грызть семечки и так далее, но и этого может оказаться недостаточно. Но можно просто есть больше. Если, съедая корзину листьев, вы удовлетворяете, например, пятую часть суточной потребности организма в азоте, значит, нужно съедать ежедневно пять корзин листьев. Но тогда вы будете получать избыток углеводов, которыми богата растительная пища, и от них нужно будет как-то избавляться. Как? Сжигать, — предлагают Классен и Нолет. Теплокровные животные легко могут себе позволить чисто растительный рацион, потому что так или иначе все время сжигают углеводы корзинами, но для холоднокровных это всегда проблематично. Теперь вернемся к листрозаврам, которые были растительноядными, и цинодонтам, среди которых были и растительноядные, и плотоядные. Не могла ли теплокровность развиться у цинодонтов оттого, что они уже обладали высокой аэробной мощностью (которая в те трудные времена была им жизненно необходима) в сочетании с рационом, богатым растительной пищей? Когда же у этих древних растительноядных выработалась теплокровность, они вполне могли воспользоваться преимуществами связанного с ней дополнительного притока энергии, чтобы быстро нарастить свою численность, преодолевая многие мили по засушливым триасовым равнинам в поисках пищи или спасаясь от хищников. У хищников, вероятно, не было столь насущной потребности в теплокровности, связанной с питанием, но им приходилось держаться наравне с оснащенными турбонагревателем травоядными. Возможно, что теплокровность понадобилась им просто потому, что иначе им было не угнаться за Черной Королевой, обернувшейся теплокровной вегетарианкой.
А как обстояли дела у гигантских динозавров — самых известных из древних растительноядных? Можно предположить, что они решили ту же проблему иначе. Если съедать пять корзин листьев в день, но не сжигать их постоянно, можно просто накапливать где-то потребляемую пищу, то есть увеличивать размер тела! Гиганты не только обладают большей “вместимостью”, но и всегда отличаются более низкой интенсивностью обмена веществ, а значит и более медленным оборотом белков и ДНК и, следовательно, меньшей потребностью в азоте. Итак, есть два способа успешно придерживаться рациона, богатого растительной пищей: крупные размеры в сочетании с замедленным обменом веществ или маленькие размеры в сочетании с ускоренным обменом веществ. О многом говорит факт, что и современные растительноядные ящерицы всегда выбирают одну из этих двух стратегий, хотя низкая аэробная мощность не дает им стать по-настоящему теплокровными. (Как предки этих ящериц пережили пермское вымирание — вопрос, который мы здесь обсуждать не будем.)
Но почему только динозавры стали такими огромными? На этот вопрос никто пока не дал достаточно убедительного ответа, хотя многие пытались. Джаред Даймонд и его коллеги в статье, опубликованной в 2001 году и посвященной другим вопросам, попутно высказали идею, что ответ вполне может крыться в повышенном уровне углекислого газа в атмосфере тех времен, что, по-видимому, увеличивало первичную продукцию, то есть скорость роста растений. Однако Даймонд не рассматривал сторону вопроса, связанную с азотом, которую осветили Классен и Нолет. Высокий уровень углекислого газа действительно повышает первичную продукцию, но одновременно и понижает содержание азота в тканях растений. Этим вопросом занимается недавно возникшая отрасль исследований, связанных с возможным влиянием нынешнего повышения уровня углекислого газа на режим питания нашей планеты. Итак, проблема, стоявшая перед цинодонтами и динозаврами, была еще острее, чем та, что стоит сегодня перед ящерицами: в те времена растительноядным животным для удовлетворения потребности в азоте нужно было есть еще больше. Строгим же вегетарианцам, должно быть, и вовсе приходилось поглощать пищу в огромных количествах.
Может быть, именно поэтому тероподам не была нужна теплокровность. Они были плотоядными, а значит, проблема баланса азота перед ними не стояла. Но, в отличие от пыхтящих цинодонтов, которым приходилось на равных соревноваться с растительноядными, оснащенными турбонагревателем, тероподы были выше этого. У них имелись необычайно эффективные легкие с аспирационным насосом, позволявшие им ловить все, что движется.
Лишь позже, уже в меловом периоде, некоторые из рапто- ров сделались вегетарианцами. Одним из первых, судя по всему, был манираптор Falcarius utahensis, описанный в 2005 году в журнале “Нейчур” группой исследователей из Юты. Линдси Занно, которая была в числе авторов, формально описавших этот вид, неформально охарактеризовала его как “предел несуразности: помесь страуса, гориллы и Эдварда Руки-ножницы”. Так или иначе, это было настоящее недостающее звено, наполовину раптор, наполовину травоядное, жившее примерно в то самое время, когда на Земле распространились цветковые растения, сделавшие переход к вегетарианству заманчивым как никогда. Но для предмета нашего обсуждения самый важный факт, касающийся этого вида, состоит, пожалуй, в том, что он относился к группе манирапторов, от которой предположительно произошли птицы. Не могло ли возникновение теплокровности у птиц тоже быть связано с переходом к вегетарианскому рациону, потребовавшему сжигать гораздо больше пищи для удовлетворения потребности в азоте?
Мы завершим эту главу умозрительными рассуждениями. Но от умозрительных рассуждений легко перейти к гипотезе — тому самому мысленному прыжку в неизвестность, о котором писал Питер Медавар, а именно это и есть основа настоящей науки. Здесь остается еще многое исследовать и проверять. Но если мы хотим разобраться в причинах ускоренного темпа нашей жизни, нам может понадобиться учитывать не только физиологические принципы, но и саму историю жизни на планете, причем тот ее период, когда особенно большую роль играли исключительные обстоятельства. Возможно, эта проблема скорее историческая, чем естественнонаучная, — в том смысле, что события могли пойти иначе, но случилось то, что случилось. Могла ли высокая аэробная мощность никогда не стать вопросом жизни и смерти, если бы не было пермского вымирания и последовавшего за ним периода пониженного уровня кислорода в атмосфере? Могла ли эволюция ничего не сделать с примитивными легкими рептилий? И могла ли теплокровность никогда не возникнуть, если бы несколько животных, обладавших высокой аэробной мощностью, не стали вегетарианцами? Даже если это исторические, а не естественнонаучные вопросы, чтение летописей этой истории все-таки составляет предмет естественных наук, и результаты, которые приносят исследования в этой области, существенно обогащают наши представления о природе жизни.
Назад: Глава 7. Зрение
Дальше: Глава 9. Сознание

